
|
|
|
|
 Far Far |
| WinNavigator |
| Frigate |
| Norton
Commander |
| WinNC |
| Dos
Navigator |
| Servant
Salamander |
| Turbo
Browser |
|
|
| Winamp,
Skins, Plugins |
| Необходимые
Утилиты |
| Текстовые
редакторы |
| Юмор |
|
|

|
File managers and best utilites |
Реферат: Мейоз, его цитогенетические механизмы и биологическое значение. Реферат мейоз
Реферат - Мейоз
МЕЙОЗ
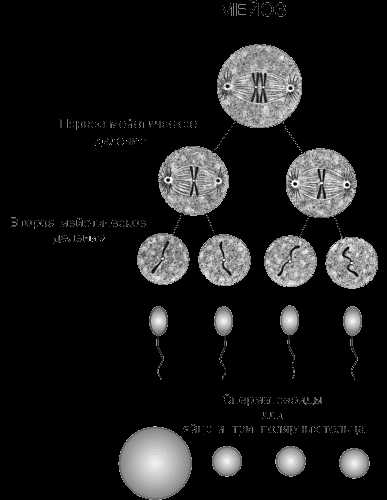
МЕЙОЗ – это деление клетки, при котором происходит редукция числа хромосом и их перекомбинация у дочерних клеток по сравнению с материнской. Мейоз – основа полового размножения, при котором потомство не идентично родителям. Важнейшая его эволюционная роль – барьер на пути нежизнеспособных комбинаций хромосом и генов. Мейоз протекает в два деления, первое из которых называется редукционным (в процессе именно этого деления количество хромосом у дочерних клеток уменьшается в два раза), а второе – эквационным (в результате него происходит равномерное распределение хромосом по дочерним клеткам, оно аналогично митозу).
МЕЙОЗ
Половое размножение организмов осуществляется с помощью специализированных клеток, т.н. гамет, – яйцеклетки (яйца) и спермия (сперматозоида). Гаметы, сливаясь, образуют одну клетку – зиготу. Каждая гамета гаплоидна, т.е. имеет по одному набору хромосом. Внутри набора все хромосомы разные, однако каждой хромосоме яйцеклетки соответствует одна из хромосом спермия. Зигота, таким образом, содержит уже пару таких соответствующих друг другу хромосом, которые называют гомологичными. Гомологичные хромосомы сходны, поскольку имеют одни и те же гены или их варианты (аллели), определяющие специфические признаки. Например, одна из парных хромосом может иметь ген, кодирующий группу крови А, а другая – его вариант, кодирующий группу крови В. Хромосомы зиготы, происходящие из яйцеклетки, являются материнскими, а происходящие из спермия – отцовскими.
В результате многократных митотических делений из образовавшейся зиготы возникает либо многоклеточный организм, либо многочисленные свободноживущие клетки, как это происходит у обладающих половым размножением простейших и у одноклеточных водорослей.
При образовании гамет диплоидный набор хромосом, имевшийся у зиготы, должен наполовину уменьшиться (редуцироваться). Если бы этого не происходило, то в каждом поколении слияние гамет приводило бы к удвоению набора хромосом. Редукция до гаплоидного числа хромосом происходит в результате редукционного деления – т.н. мейоза, который представляет собой вариант митоза.
(23.48 Кб)
Расщепление и рекомбинация. Особенность мейоза состоит в том, что при клеточном делении экваториальную пластинку образуют пары гомологичных хромосом, а не удвоенные индивидуальные хромосомы, как при митозе. Парные хромосомы, каждая из которых осталась одинарной, расходятся к противоположным полюсам клетки, клетка делится, и в результате дочерние клетки получают половинный, по сравнению с зиготой, набор хромосом.
Для примера предположим, что гаплоидный набор состоит из двух хромосом. В зиготе (и соответственно во всех клетках организма, продуцирующего гаметы) присутствуют материнские хромосомы А и В и отцовские А' и В'. Во время мейоза они могут разделиться следующим образом:
Наиболее важен в этом примере тот факт, что при расхождении хромосом вовсе не обязательно образуется исходный материнский и отцовский набор, а возможна рекомбинация генов, как в гаметах АВ' и А'В в приведенной схеме.
Теперь предположим, что пара хромосом АА' содержит два аллеля – a и b – гена, определяющего группы крови А и В. Сходным образом пара хромосом ВВ' содержит аллели m и n другого гена, определяющего группы крови M и N. Разделение этих аллелей может идти следующим образом:
Очевидно, что получившиеся гаметы могут содержать любую из следующих комбинаций аллелей двух генов: am, bn, bm или an.
Если имеется большее число хромосом, то пары аллелей будут расщепляться независимо по тому же принципу. Это означает, что одни и те же зиготы могут продуцировать гаметы с различными комбинациями аллелей генов и давать начало разным генотипам в потомстве.
Мейотическое деление. Оба приведенных примера иллюстрируют принцип мейоза. На самом деле мейоз – значительно более сложный процесс, так как включает два последовательных деления. Главное в мейозе то, что хромосомы удваиваются только один раз, тогда как клетка делится дважды, в результате чего происходит редукция числа хромосом и диплоидный набор превращается в гаплоидный.
Во время профазы первого деления гомологичные хромосомы конъюгируют, т. е. сближаются попарно. В результате этого очень точного процесса каждый ген оказывается напротив своего гомолога на другой хромосоме. Обе хромосомы затем удваиваются, но хроматиды остаются связанными одна с другой общей центромерой.
В метафазе четыре соединенные хроматиды выстраиваются, образуя экваториальную пластинку, как если бы они были одной удвоенной хромосомой. В противоположность тому, что происходит при митозе, центромеры не делятся. В результате каждая дочерняя клетка получает пару хроматид, все еще связанных цетромерой. Во время второго деления хромосомы, уже индивидуальные, опять выстраиваются, образуя, как и в митозе, экваториальную пластинку, но их удвоения при этом делении не происходит. Затем центромеры делятся, и каждая дочерняя клетка получает одну хроматиду.
Деление цитоплазмы. В результате двух мейотических делений диплоидной клетки образуются четыре клетки. При образовании мужских половых клеток получается четыре спермия примерно одинаковых размеров. При образовании же яйцеклеток деление цитоплазмы происходит очень неравномерно: одна клетка остается крупной, тогда как остальные три настолько малы, что их почти целиком занимает ядро. Эти мелкие клетки, т.н. полярные тельца, служат лишь для размещения избытка хромосом, образовавшихся в результате мейоза. Основная часть цитоплазмы, необходимой для зиготы, остается в одной клетке – яйцеклетке.
Конъюгация и кроссинговер. Во время конъюгации хроматиды гомологичных хромосом могут разрываться и затем соединяться в новом порядке, обмениваясь участками следующим образом:
Этот обмен участками гомологичных хромосом называется кроссинговером (перекрестом). Как показано выше, кроссинговер ведет к возникновению новых комбинаций аллелей сцепленных генов. Так, если исходные хромосомы имели комбинации АВ и ab, то после кроссинговера они будут содержать Ab и aB. Этот механизм появления новых генных комбинаций дополняет эффект независимой сортировки хромосом, происходящей в ходе мейоза. Различие состоит в том, что кроссинговер разделяет гены одной и той же хромосомы, тогда как независимая сортировка разделяет только гены разных хромосом.
studfiles.net
Доклад - Мейоз. Рекомбинация генетического материала
АО «МЕДИЦИНСКИЙ УНИВЕРСИТЕТ АСТАНА»
Кафедра __________________________________________________
СРС
Тема: Мейоз. Рекомбинация генетического материала.
Выполнил: _______________
Проверила: _______________
Астана 2010
План:
1.Понятие.
2.Фазы мейоза.
3.Рекомбинация генетического материала.
4.Биологическое значение рекомбинации.
Мейоз.
Мейоз (от греч. meiosis — уменьшение) — особый способ деления клеток, деление созревания, в результате которого происходит редукция (уменьшение) числа хромосом и переход клеток их диплоидного состояния в гаплоидное. Мейоз — это особый тип дифференцировки, специализации клеток, который приводит к образованию половых клеток. Этот процесс занимает два клеточных цикла при отсутствии синтеза ДНК во втором мейотическом делении. Необходимо отметить, что мейоз представляет собой универсальное явление, характерное для всех эукариотических организмов. При мейозе происходит не только редукция числа хромосом до гаплоидного их числа, но происходит чрезвычайно важный генетический процесс — обмен участками между гомологичными хромосомами, процесс, получивший название кроссинговера.
Существует несколько разновидностей мейоза. При зиготном (характерном для аскомицетов, базимицетов, некоторых водорослей, споровиков и др.), для которых в жизненном цикле преобладает гаплоидная фаза, две клетки — гаметы сливаются, образуя зиготу с двойным (диплоидным) набором хромосом. В таком виде диплоидная зигота (покоящаяся спора) приступает к мейозу, дважды делиться, и образуется четыре гаплоидные клетки, которые продолжают размножаться.
Споровый тип мейоза встречается у высших растений, клетки которых имеют диплоидный набор хромосом. В данном случае в органах размножения растений, образовавшиеся после мейоза гаплоидные клетки еще несколько раз делятся. Другой тип мейоза, гаметный, происходит во время созревания гамет — предшественников зрелых половых клеток. Он встречается у многоклеточных животных, среди некоторых низших растений.
В случае гаметного мейоза характерно при развитии организма выделение клонов герминативных клеток, которые впоследствии будут дифференцироваться в половые клетки. И только клетки этих клонов будут при созревании подвергаться мейозу и превращаться в половые клетки. Следовательно, все клетки развивающихся многоклеточных животных организмов можно разделить на две группы: соматические — из которых будут образовываться клетки всех тканей и органов, и герминативные, которые дадут начало половым клеткам.
Такое выделение герминативных клеток (гоноцитов) обычно происходит на ранних стадиях эмбрионального развития. Так, детерминация гоноцитов у рачка циклопа происходит уже на первом делении зиготы: одна из двух клеток дает начало герминальным клеткам. У аскариды герминативные клетки или клетки «зародышевого пути» (А.Вейсман) выделяются на стадии 16 бластомеров, у дрозофилы — на стадии бластоцисты, у человека — первичные половые клетки (гонобласты) появляются на 3-ей неделе эмбрионального развития в стенке желточного мешка в каудальном отделе эмбриона.
Как и все клетки развивающегося организма, клетки зародышевого пути диплоидны. Они могут увеличиваться в числе путем обычного митоза, повторяя все стадии обычного клеточного цикла, где происходит чередование уровней количества ДНК и хромосом на клетку:
2n (2c) ® S-период® 4n (4c) ® 2 клетки 2n (2c) и т.д.
Однако на определенных стадиях развития при половом созревании особей этот обычный ход смены событий меняется. Герминативные клетки превращаются в гониальные (оогонии — женские и сперматогонии — мужские клетки — предшественники), и они вступают в процесс мейоза. При этом как для женских, так и для мужских клеток наступает первый цикл мейоза. На этой и следующей стадии половые клетки получили название сперматоцитов и ооцитов (I и II порядка).
В первом клеточном цикле мейоза происходит целый ряд событий, который его значительно отличает от обычного клеточного цикла. После вступления в I цикл созревания и сперматоциты I и ооциты I порядков синтезируют ДНК, её количеств удваивается, так же как удваивается за счет репликации количество хромосом. Следовательно, после S-периода эти клетки нужно считать (также как и соматические клетки после синтеза ДНК) тетраплоидными. После короткого G2-периода наступает профаза I мейотического деления, которая резко отличается от обычной мейотический профазы.
Особенности профазы I мейотического деления
Во-первых, эта стадия занимает большой отрезок времени (от суток до годов !). Во-вторых, она состоит из нескольких структурно-функциональных фаз (лептотена, зиготена, пахитена, диплотена, диакинез). Далее, в этот период происходит объединение, конъюгация, гомологичных (родительских) удвоенных в результате репликации хроматид, при этом образуются т.н. тетрады, т.е. хромосомные комплексы, состоящие из четырех хроматид (удвоенные материнские и удвоенные отцовские), которые соединены вместе с помощью специальной структуры — синаптинемного комплекса. В это же время происходит обмен участками между хроматидами гомологичных хромосом (но не между сестринскими хроматидами одного гомолога) — кроссинговер. Кроме того, в процессе конъюгации и обмена происходит синтез примерно 1,5% хромосомной ДНК.
В профазе I мейотического деления наблюдается рост объема ооцитов, в которых накапливаются запасные вещества, обеспечивающие ранние стадии развития будущего зародыша.
Эта профаза отличается также длительностью во времени, необходимого для прохождения перечисленных выше событий. Обычная соматическая профаза длится 0,5-1,5 часа. Мейотическая профаза сперматоцита I порядка у самцов мыши длится 12 суток, у человека — 24 дня (плюс еще около двух месяцев до полного созревания сперматозоида). Среди женских половых клеток профаза I порядка тритона обыкновенного длится около 1 года, у мыши от 4 месяцев до 3 лет, у человека профаза I ооцитов начинается на 3-ем месяце внутриутробного развития и может продолжаться до 50-летнего возраста женщины. При этом у человека происходит постепенная гибель заложенных ооцитов: у 3-х месячного эмбриона их около7х106 клеток, к рождению ребенка их остается около 2х106, к половому созреванию — 3х105, всего же созревает (овулируют) примерно 5х102 ооцитов.
Другой особенностью профазы I меойза является то, что в отличие от обычного митоза, хромосомы сохраняют ряд функциональных нагрузок, а именно: они способны к синтезу РНК, частичному синтезу ДНК, претерпевают ряд структурных перестроек. Другими словами, профазные хромосомы I мейотического деления не находятся в состоянии функционального покоя, а участвуют в целом ряде событий.
Стадии профазы I мейотического деления
Вся профаза I мейотического деления состоит из нескольких стадий: лептотена — стадия тонких нитей (хромосом), зиготена — стадия сливающихся (объединяющихся, конъюгирующих) нитей, пахитена — стадия толстых нитей, диплотена — стадия двойных нитей, диакинез — стадия расходящихся нитей.
Из всех стадий профазы I самой длительной является стадия пахитены, в ряде случаев она занимает до 50% времени.
Так, у человека при спермиогенезе стадии лептотены с зиготеной занимают 6,5 сут, пахатина- 15, диплотена и диакинез — 0,8; у мыши лептотена с зиготеной длятся около 3 суток, пахитена — 7 суток, диплотена с диакинезом — около 2 суток; у тритона лептотена занимает 5 сут, зиготена — 8, пахитена — 4-5, диплотена — 2 сут; у домашнего сверчка лептотена и зиготена занимают 2-3 сут, пахитена — 6-9, диплотена — 2. По сравнению с обычным митозом продолжительность деления клеток в процессе мейоза несравнимо длительнее. Это особенно наглядно видно при созревании женских половых клеток у животных, у которых яйцеклетки могут останавливаться в развитии на несколько месяцев и даже лет в стадии диплотены профазы I-го мейотического деления.
У растений мейоз также намного длиннее митоза по времени. Так, у традесканции весь мейоз занимает около 5 сут, из которых на профазу I-го деления приходится 4 сут, но встречаются виды, у которых мейоз идет со скоростью, соизмеримой с митозом.
Лептотена, или стадия тонких нитей, морфологически напоминает раннюю профазу митоза, но отличается тем, что при мейозе ядра обычно крупнее и хромосомы очень тонкие, так что проследить их по всей длине очень трудно.
Длина каждой мейотической хромосомы на ранних стадиях мейоза может быть в 10-100 раз больше длины соответствующих митотических хромосом. Следовательно, мейотические хромосомы имеют меньшую степень компактизации, они примерно в 30 раз менее компактны, чем хромосомы в метафазе мейоза. В лептотене хромосомы удвоены, но сестринские хроматиды в них далеко не всегда удается различить (так же как в хромосомах в ранней профазе митоза). Таким образом, в лептотене содержится диплоидное количество (2n) сдвоенных сестринских хроматид, общее количество последних, как и при митозе, равно 4n вследствие редупликации в S-периоде.
Расположение хромосом в лептотене часто повторяет телофазную поляризацию ядра. При этом у некоторых животных хромосомы образуют так называемую фигуру «букета» — дугообразно изогнутые сближенные хромосомы, связанные своими теломерами с ядерной оболочкой. У некоторых растений в конце лептотены хромосомы собираются в клубок (синезис).
Характерным для лептотены является появление на тонких хромосомах сгустков хроматина — хромомеров, которые как бы нанизаны в виде бусинок и располагаются по всей длине хромосомы. Число, размер и расположение таких хромомерных участков характерны для каждой хромосомы. Это позволяет составлять морфологические карты хромосом и использовать их для цитологического анализа. Число хромомеров различно у разных объектов: всего у тритона на 12 хромосомах их 2,5 тыс., у сверчка — около 200, у риса на 24 хромосомы — 645.
В лептотене начинает выявляться следующий, чрезвычайно важный и характерный для мейоза процесс конъюгации гомологичных хромосом, их сближение, которое начинается в теломерных участках, связанных с ядерной оболочкой. В этих местах образуется сложная специальная структура — тяж белковой природы, синаптонемный комплекс, который позже, в зиготене свяжет гомологичные удвоенные хроматиды по всей их длине.
Рекомбинация генетического материала.
Генетическая рекомбинация — это перераспределение генетического материала (ДНК), приводящее к возникновению новых комбинаций генов. Рекомбинация может происходить путем обмена клеточными ядрами, целыми молекулами ДНК или частями молекул. В то время как процессы репликации и репарации ДНК обеспечивают воспроизведение и сохранение генетического материала, рекомбинация приводит к генетической изменчивости. Биологическое значение рекомбинации столь велико, что она получила развитие у всех живых организмов. Она может происходить у эукариот (как при образовании половых клеток — гамет, так и в соматических клетках), у бактерий и даже при размножении вирусов, в том числе таких, генетический материал которых состоит из РНК. Перетасовка хромосом в мейозе, приводящая к огромному разнообразию гамет, случайность слияния гамет при оплодотворении, обмен частями между гомологичными хромосомами — все это (и далеко не только это) относится к рекомбинации.
Понятно, что из широкого круга рекомбинационных явлений интерес молекулярных биологов в первую очередь вызывает рекомбинация, заключающаяся в обменах частями между молекулами ДНК, ведь здесь можно применять весь арсенал методов генетики и молекулярной биологии, и эти исследования перекрываются с изучением других важных генетических процессов, прежде всего репликации и репарации ДНК. Но даже в таком виде, суженном до обменов частями молекул ДНК, понятие «рекомбинация» включает большой набор разных по своей природе явлений. При этом для всех рекомбинационных процессов общим является этап, на котором молекулы ДНК вступают в контакт в участке, где произойдет обмен полинуклеотидными цепями. Этот этап получил название «синапсис». Однако механизм синапсиса при разных типах рекомбинации принципиально различен. Более того, он является одним из критериев при классификации рекомбинационных явлений.
Прежде чем перейти к их рассмотрению, напомним некоторые термины и понятия, которыми мы будем пользоваться. Молекула ДНК представляет собой дуплекс — структуру из двух закрученных в спираль полинуклеотидных цепей. Последовательность нуклеотидов в цепях определяет специфичность ДНК и несет генетическую информацию. Молекулы, имеющие общее происхождение и состоящие из одинаковых нуклеотидных последовательностей, называют гомологичными. Однако их идентичность нарушается из-за мутаций, накапливающихся в течение поколений. По большей части мутации приводят к заменам единичных нуклеотидов, реже к выпадениям и вставкам отдельных нуклеотидов. Поэтому нарушения гомологии в результате мутаций не очень существенны по сравнению с основной массой идентичных нуклеотидов, и в таких случаях можно говорить об общей гомологии молекул. Каждая новая мутация приводит к образованию нового аллеля в том гене, где она возникла. Следовательно, новые аллели обычно отличаются от исходной формы одним нуклеотидом. Если мутация приводит к изменению фенотипа у исходной формы, то к ней можно применять также термин «генетический маркер».
Две цепи, составляющие дуплекс ДНК, антипараллельны, то есть имеют разную полярность: одна цепь имеет направление 5'-3', другая — 3'-5'. Цепи удерживаются вместе водородными связями между парами комплементарных оснований А-Т и G-C. Поэтому обе цепи в дуплексе являются также комплементарными. Процесс расхождения цепей в результате разрыва водородных связей есть денатурация, обратная реакция — ренатурация. Все это сказано для того, чтобы подвести читателя к отправной идее статьи: поскольку отдельные цепи ДНК, полученные от разных родителей, гомологичны и, следовательно, комплементарны, они могут ренатурировать, формируя новый дуплекс. Иными словами, гомологичные ДНК могут узнавать друг друга по комплементарности их нуклеотидной последовательности. Новый дуплекс, состоящий из цепей от разных молекул, называется гетеродуплексом.
А теперь можно дать классификацию основных типов рекомбинации. Все, что говорилось о гомологии ДНК и комплементарности полинуклеотидных цепей, относится к гомологичной, или общей, рекомбинации (она же кроссинговер), основанной на спаривании комплементарных цепей ДНК. От других типов рекомбинационных процессов ее отличают необходимость в общей (по всей длине молекул) гомологии между рекомбинирующими ДНК и участие большого набора специальных белков. Гомологичная рекомбинация начинается с возникновения в одном или обоих дуплексах участков из одиночных цепей ДНК, которые затем с помощью специальных белков находят комплементарные последовательности в гомологичном дуплексе и образуют с ними гетеродуплекс — ключевой промежуточный продукт (интермедиат) рекомбинации. Конечным результатом рекомбинации будет обмен равными частями гомологичных молекул (рис. 1).
Из общей рекомбинации можно выделить как частный случай так называемую эктопическую рекомбинацию. Она заключается в обменах (кроссинговерах) между отдельными участками гомологичной ДНК, разбросанными по геному. К ним относятся разнообразные подвижные элементы, названные так за способность перемещаться по геному, гены транспортных и рибосомных РНК, гистонов и многие другие повторяющиеся последовательности (повторы) ДНК. Такая локальная гомологичная рекомбинация интересна прежде всего тем, что она может приводить к хромосомным перестройкам, хотя ее биологическая роль этим не исчерпывается. На рис. 2 в качестве примера приведены схемы возникновения инверсий (поворотов внутренних участков хромосом на 180?), утрат (делеций) и удвоений (дупликаций) частей хромосом в результате эктопической рекомбинации. Это только часть возможных перестроек хромосом. Другие их типы могут возникать в зависимости от того, какова ориентация повторов ДНК по отношению друг к другу (прямая или обратная), и от того, где они расположены: внутри одной хромосомы, в сестринских хроматидах или разных хромосомах. Несмотря на то, что обмены происходят между локальными участками гомологии, эктопическая рекомбинация осуществляется в основном теми же белками, что и гомологичная. Принципиально иными являются три других типа рекомбинации, которые основаны не на взаимодействии комплементарных цепей ДНК, а на совершенно иных механизмах и участии иных белков.
Биологическое значение гомологичной рекомбинации огромно. Прежде всего она вносит большой вклад в лежащую в основе эволюции генетическую изменчивость, позволяющую организмам постоянно приспосабливаться к среде обитания. Преимущества перекомбинаций генов настолько велики, что рекомбинационные системы появились у вирусов и бактерий, которые размножаются вегетативно. У эукариот они достигли большего разнообразия и сложности, особенно в соматических клетках. Эктопическая рекомбинация приводит к перестройкам хромосом, с которыми (прежде всего с дупликациями) связывают эволюцию генетического аппарата. Считается, что дупликации участков хромосом обеспечили материал для дивергенции нуклеотидных последовательностей, приводящей к возникновению новых генов.
Однако биологическое значение гомологичной, и в том числе эктопической, рекомбинации нельзя свести к их роли в эволюции. Большую роль они играют и в разнообразных онтогенетических перестройках генетического материала, участвующих в регуляции работы генов. Например, конверсия гена (коррекция гетеродуплекса), которая в мейотических клетках является одним из этапов общего процесса кроссинговера, в соматических клетках эукариот и клетках бактерий может не сопровождаться кроссинговером по внешним генам и выступать как самостоятельное явление. Такая конверсия выполняет важные функции в онтогенезе бактерий, дрожжей, животных. Известно много примеров, когда определенный ген расположен в локусе, где он имеет собственный промотор и может функционировать, в то время как в других локусах находятся последовательности, в основном гомологичные этому гену, но заметно отличающиеся по нуклеотидному составу из-за накопившихся в них мутаций. Они лишены промотора и не могут выполнять функции генов. Эти «молчащие» последовательности могут вступать в синапсис с работающим геном и служить матрицей для его конверсии. Таким образом, работающий ген может менять свою нуклеотидную последовательность. Подобным способом клетки гомоталличных штаммов дрожжей меняют свой половой тип.
У некоторых патогенных микроорганизмов этот же механизм, позволяющий их клеткам менять свои поверхностные антигены, участвует в процессах, описанных ниже. Так, многие патогенные бактерии (спирохета Borrelia bormsei, гонококки и др.) и простейшие (африканские трипаносомы), с одной стороны, и животные, в которых они паразитируют, — с другой, используют в борьбе друг против друга в сущности сходные приемы. Животные продуцируют в огромном ассортименте антитела, обеспечивающие им иммунитет, а патогенные микроорганизмы в ответ на это образуют на своей поверхности все новые и новые антигены, позволяющие им уходить от иммунного ответа хозяйского организма. В основе данных процессов лежат рекомбинационные перестройки в локусах, кодирующих антигены (или антитела). Рекомбинационные перестройки включают одни и выключают другие гены либо создают новые гены. В этих сложных процессах участвуют разные типы рекомбинации, но гомологичная и эктопическая рекомбинации (и в том числе конверсия гена) играют здесь не последнюю роль. Помимо описанных процессов у бактерий и низших эукариот известны и другие рекомбинационные системы, участвующие в регуляции работы генов. Но это тема следующей статьи.
Список литературы:
Слюсарев А.А., Жукова С.В. Биология – Киев. Вища школа. 1987
Лобашов М.Е. Генетика – Л. Изд. Ленинградского унив., 1967
Биология.: Учебник для мед. спец. ВУЗ-ов. Под ред. В.Н. Ярыгина М., Высшая Школа, 1997
Албертс Б., Брей Д., Льюис Дж. и др. Молекулярная биология клетки:
Пер. с англ. М.: Мир, 1994. Т. 1, ч. 2. С. 301-310.
Инге-Вечтомов С.Г. Введение в молекулярную генетику. М.: Высш. шк., 1983. С. 120-136.
Льюин Б. Гены. Пер. с англ. М.: Мир, 1987. С. 443-453.
www.ronl.ru
Реферат - Мейоз. Овогенез. Сперматогенез
Содержание
Введение. 2
1. Половое размножение многоклеточных. 3
2. Гаметогенез. 4
2.1 Сперматогенез. 4
2.2 Овогенез. 5
2.3 Мейоз. 6
Вывод. 11
Список использованной литературы… 12
Огромное влияние на жизнь человечества в наши дни оказывают генетика и связанные с ней исследования физиологических особенностей размножения человека.
Современные люди часто не помнят или забывают о прошлом и считают, что интерес к генетике и связанным с ней вопросам возник недавно. Но можно утверждать, что интерес к вопросам размножения столь же древен, как и наш собственный биологический вид.
Размножение — основное проявление жизнедеятельности для любого вида, даже если его представители этого и не осознают.
Мендель открыл основные законы наследственности приблизительно в то же время, когда другие биологи принялись изучать строение клетки. Поэтому когда законы генетики открыли заново в 1900-х годах, ученые уже смогли понять, что так называемые факторы должны соответствовать наблюдаемым в реальности структурам — хромосомам. Отождествление абстрактных понятий и реальных структур — крупнейшее достижение генетики на ранней стадии ее развития, и оно было сделано в основном благодаря исследованию природы пола. [1]
Развитие гамет у многоклеточных животных происходит в половых железах — гонадах (греч. gone — семя). Различают два типа половых клеток: мужские (сперматозоиды) и женские (яйцеклетки). Сперматозоиды развиваются в семенниках, яйцеклетки в яичниках.
Процесс формирования половых клеток (гамет) известен под общим названием гаметогенеза. Он характеризуется рядом весьма важных биологических процессов и протекает несколько по-разному при созревании сперматозоидов (сперматогенез) и яйцеклеток (овогенез). [2]
Семенник состоит из многочисленных канальцев. На поперечном разрезе через каналец видно, что его стенка имеет несколько слоев клеток. Они представляют собой последовательные стадии развития сперматозоидов.
Наружный слой составляют сперматогонии — клетки округлой формы; у них относительно большое ядро и значительное количество цитоплазмы. В период эмбрионального развития и после рождения до полового созревания сперматогонии делятся путем митоза, благодаря чему увеличивается число этих клеток и семенник. Период интенсивного деления сперматогоний называют периодом размножения. После наступления половой зрелости часть сперматогониев также продолжает делиться митотически и образовывать такие же клетки, но некоторые из сперматогониев перемещаются в следующую зону роста, расположенную ближе к просвету канальца. Здесь происходит значительное возрастание размеров клеток за счет увеличения количества цитоплазмы. В этой стадии они называются сперматоцидами первого порядка.
Третий период развития мужских гамет называется периодом созревания. В этот период происходят два быстро наступающих одно вслед за другим деления. Из каждого сперматоцида первого порядка сначала образуются два сперматоцида второго порядка, а затем четыре сперматиды, имеющие овальную форму и значительно меньшие размеры. Деление клеток во время периода созревания сопровождается перестройкой хромосомного аппарата (происходит мейоз). Сперматиды перемещаются в зону, ближайшую к просвету канальцев, где из них формируются сперматозоиды.
У большинства диких животных сперматогенез происходит лишь в определенные периоды года. В промежутках между ними в канальцах семенников содержатся лишь сперматогонии. Но у человека и большинства домашних животных сперматогенез происходит в течение всего года. [3]
Фазы овогенеза сопоставимы с таковыми при сперматогенезе. В этом процессе также имеется период размножения, когда интенсивно делятся овогонии — мелкие клетки с относительно крупным ядром и небольшим количеством цитоплазмы. У млекопитающих и человека этот период заканчивается еще до рождения. Сформировавшиеся к этому времени овоциты первого порядка сохраняются далее без изменений многие годы. С наступлением половой зрелости периодически отдельные овоциты вступают в период роста. Овоциты увеличиваются, в них накапливаются желток, жир, пигменты. В цитоплазме клетки в ее органоидах и мембранах происходят сложные морфологические биохимические преобразования. Каждый овоцит окружается мелкими фолликулярными клетками, обеспечивающими его питание.
Далее наступает период созревания, в процессе которого происходят два последовательных деления, связанных с преобразованием хромосомного аппарата (мейоз). Кроме того, эти деления сопровождаются неравномерным разделением цитоплазмы между дочерними клетками. При делении овоцита первого порядка образуется одна крупная клетка — овоцит второго порядка, содержащая почти всю цитоплазму, и маленькая клетка, получившая название полярного, или редукционного тельца.
При втором делении созревания цитоплазма снова распределяется неравномерно. Образуется одна крупная овотида и второе редукционное тельце. В это время первое редукционное тельце также может разделиться на две клетки. Таким образом, из одного овоцита первого порядка образуются одна овотида и три редукционных тельца.
Далее из овотиды формируется яйцо, а редукционные тельца рассасываются или сохраняются на поверхности яйца, но не принимают участия в дальнейшем развитии. Неравномерное распределение цитоплазмы обеспечивает яйцу получение значительного количества цитоплазмы и питательных веществ, которые потребуются в будущем для развития зародыша.
У млекопитающих и человека периоды размножения и роста яйцевых клеток проходят в фолликулах. Фолликул заполнен жидкостью, внутри него находится яйцеклетка. Во время овуляции стенка фолликула лопается, яйцеклетка попадает в брюшную полость, а затем, как правило, в яйцеводы (маточные трубы). Период созревания яйцевых клеток протекает в трубах, здесь же происходит оплодотворение.
У многих животных овогенез и созревание яиц совершаются лишь в определенные сезоны года. У женщин обычно ежемесячно созревает одно яйцо, а за весь период половой зрелости около 400 яиц.
В ядрах незрелых половых клеток также, как и ядрах соматических клеток, все хромосомы парные, набор хромосом двойной (2 n), диплоидный. В процессе созревания половых клеток происходит редукционное деление (мейоз), при котором число хромосом уменьшается, становится одинарным (n), гаплоидным. Мейоз (от греч. meiosis — уменьшение) происходит во время гаметогенеза. Этот процесс совершается во время двух следующих одно за другим делений периода созревания, называемых соответственно первым и вторым мейотическим делением. Каждое из этих делений имеет фазы, аналогичные митозу.
Схематично эти фазы можно изобразить так:
Интерфаза I
Профаза I
Мейоз Деление первое Прометофаза I
Метафаза I
Анафаза I
Телофаза I
Интерфаза II — ин — Профаза II
терокинез Метафаза II
Деление второе Анафаза II
Телофаза II
В интерфазе I (по-видимому, еще в период роста) происходит удвоение количества хромосомного материала путем редупликации молекул ДНК.
Из всех фаз наиболее продолжительна и сложна по протекающим в ней процессам профаза I. В ней различают 5 последовательных стадий. Лептонема — стадия длинных, тонких, слабо спирализованных хромосом, на которых видны утолщения — хромомеры. Зигонема — стадия попарного соединения гомологичных хромосом, при котором хромомеры одной гомологичной хромосомы точно прикладываются к соответствующим хромомерам другой (это явление называется конъюгацией, или синапсисом). Пахинема — стадия толстых нитей. Гомологичные хромосомы соединены в пары — биваленты. Число бивалентов соответствует гаплоидному набору хромосом. На этой стадии каждая из хромосом, входящих в бивалент, состоит уже из двух хроматид, поэтому каждый бивалент включает в себя четыре хроматиды. В это время конъюгирующие хромосомы переплетаются, что приводит к обмену участками хромосом (происходит так называемый перекрест, или кроссинговер). Диплонема — стадия, когда гомологичные хромосомы начинают отталкиваться друг от друга, но в ряде участков, где происходит кроссинговер, они продолжают быть еще связанными. Диакинез — стадия, на которой отталкивание гомологичных хромосом продолжается, но они еще остаются соединенными в биваленты своими концами, образуя характерные фигуры — кольца и кресты. На этой стадии хромосомы максимально спирализованы, укорочены и утолщены. Непосредственно после диакинеза ядерная оболочка растворяется.
В прометафазе I спирализация хромосом достигает наибольшей степени. Они перемещаются в области экватора.
В метафазе I биваленты располагаются по экватору, так что центромеры гомологичных хромосом обращены к противоположным полюсам и отталкиваются друг от друга.
В анафазе Iначинают расходиться к полюсам не хроматиды, а целые гомологичные хромосомы каждой пары, так как в отличие от митоза центромера не делится и хроматиды не разъединяются. Этим первое мейотическое деление принципиально отличается от митоза. Деление заканчивается телофазой I.
Таким образом, во время первого мейотического деления происходит расхождение гомологичных хромосом. В каждой дочерней клетке уже содержится гаплоидное число хромосом, но содержание ДНК еще равно диплоидному их набору. Вслед за короткой интерфазой, во время которой синтеза ДНК не происходит, клетки вступают во второе мейотическое деление.
Профаза II продолжается недолго. Во время метафазы II хромосомы выстраиваются по экватору, центромеры делятся. В анафазе II сестринские хроматиды направляются к противоположным полюсам. Деление заканчивается телофазой II. После этого деления хроматиды, попавшие в ядра дочерних клеток, называются хромосомами.
Итак, при мейозе гомологичные хромосомы соединяются в пары, затем в конце первого мейотического деления расходятся по одной в дочерние клетки. Во время вторго мейотического деления гомологичные хромосомы расщепляются и расходятся в новые дочерние клетки. Следовательно, в результате двух последовательных мейотических делений из одной клетки с диплоидным набором хромосом образуются четыре клетки с гаплоидным набором хромосом. В зрелых гаметах количество ДНК вдвое меньше, чем соматических клетках.
При образовании как мужских, так и женских половых клеток, происходят принципиально одни и те же процессы, хотя в деталях они несколько различаются.
Значение мейотического деления в следующем:
Это тот механизм, которым обеспечивается поддержание постоянства числа хромосом. Если бы не происходило редукции числа хромосом при гаметогенезе, то из поколения в поколение возрастало бы их число и был бы утрачен один из существенных признаков каждого вида — постоянство числа хромосом.
При мейозе образуется большое число различных новых комбинаций негомологичных хромосом. Ведь в диплоидном наборе они двойного происхождения: в каждой гомологичной паре одна из хромосом от отца, другая — от матери. Что же происходит при мейозе? Это можно проследить на рис.2. Изображенные в ядрах сперматогонии и овогонии хромосомы отцовского происхождения обозначены черным цветом, материнские белым. Как видно на рисунке, в сперматозоидах и яйцеклетках они образуют новые сочетания, причем таких сочетаний даже при том же числе хромосом (три пары) получится больше чем изображено. Следовательно, благодаря такому механизму достигается большое число новых сочетаний наследственной информации, а именно 2, где n — число пар хромосом. Следовательно, у организма, имеющего три пары хромосом, этих сочетаний окажется 2, т.е.8; у дрозофилы, имеющей 4 пары хромосом, их будет 2, т.е.16, а у человека — 2, что составляет 8388608.
В процессе кроссинговера также происходит рекомбинация генетического материала. Практически все хромосомы, попадающие в гаметы, имеют участки, происходящие как от первоначально отцовских, так и от первоначально материнских хромосом. Этим достигается еще большая степень перекомбинации наследственного материала. В этом одна из причин изменчивости организма, дающий материал для отбора. [4]
Таким образом, принципиальное отличие полового размножения от бесполого заключается в том, что оно приводит к огромной изменчивости, образованию форм с новыми наследственными свойствами в результате перекомбинации различных свойств обоих родителей. В то время как при бесполом размножении дочерние организмы повторяют наследственную информацию единственной родительской особи. [5]
1. Гуттман Б., Гриффитс Э., Сузуки Д., Кулис Т. Генетика. М.: ФАИР — ПРЕСС, 2004., 448 с.
2. Слюсарев А.А. Биология с общей генетикой. М.: Медицина, 1978., 472с.
[1] Гуттман Б., Гриффитс Э., Сузуки Д., Кулис Т. Генетика. М.: ФАИР – ПРЕСС, 2004., 5-6.с., 115с.
[2] Слюсарев А.А. Биология с общей генетикой. М.: Медицина, 1978., — 98с.
[3] Слюсарев А.А. Биология с общей генетикой. М.: Медицина, 1978., — 100-101c.
[4] Слюсарев А.А. Биология с общей генетикой. М.: Медицина, 1978., — 102-105 с
[5] Слюсарев А.А. Биология с общей генетикой. М.: Медицина, 1978., — 105 с
www.ronl.ru
Реферат - Мейоз. Рекомбинация генетического материала
АО «МЕДИЦИНСКИЙ УНИВЕРСИТЕТ АСТАНА»
Кафедра __________________________________________________
СРС
Тема: Мейоз. Рекомбинация генетического материала.
Выполнил: _______________
Проверила: _______________
Астана 2010
План:
1.Понятие.
2.Фазы мейоза.
3.Рекомбинация генетического материала.
4.Биологическое значение рекомбинации.
Мейоз.
Мейоз (от греч. meiosis — уменьшение) — особый способ деления клеток, деление созревания, в результате которого происходит редукция (уменьшение) числа хромосом и переход клеток их диплоидного состояния в гаплоидное. Мейоз — это особый тип дифференцировки, специализации клеток, который приводит к образованию половых клеток. Этот процесс занимает два клеточных цикла при отсутствии синтеза ДНК во втором мейотическом делении. Необходимо отметить, что мейоз представляет собой универсальное явление, характерное для всех эукариотических организмов. При мейозе происходит не только редукция числа хромосом до гаплоидного их числа, но происходит чрезвычайно важный генетический процесс — обмен участками между гомологичными хромосомами, процесс, получивший название кроссинговера.
Существует несколько разновидностей мейоза. При зиготном (характерном для аскомицетов, базимицетов, некоторых водорослей, споровиков и др.), для которых в жизненном цикле преобладает гаплоидная фаза, две клетки — гаметы сливаются, образуя зиготу с двойным (диплоидным) набором хромосом. В таком виде диплоидная зигота (покоящаяся спора) приступает к мейозу, дважды делиться, и образуется четыре гаплоидные клетки, которые продолжают размножаться.
Споровый тип мейоза встречается у высших растений, клетки которых имеют диплоидный набор хромосом. В данном случае в органах размножения растений, образовавшиеся после мейоза гаплоидные клетки еще несколько раз делятся. Другой тип мейоза, гаметный, происходит во время созревания гамет — предшественников зрелых половых клеток. Он встречается у многоклеточных животных, среди некоторых низших растений.
В случае гаметного мейоза характерно при развитии организма выделение клонов герминативных клеток, которые впоследствии будут дифференцироваться в половые клетки. И только клетки этих клонов будут при созревании подвергаться мейозу и превращаться в половые клетки. Следовательно, все клетки развивающихся многоклеточных животных организмов можно разделить на две группы: соматические — из которых будут образовываться клетки всех тканей и органов, и герминативные, которые дадут начало половым клеткам.
Такое выделение герминативных клеток (гоноцитов) обычно происходит на ранних стадиях эмбрионального развития. Так, детерминация гоноцитов у рачка циклопа происходит уже на первом делении зиготы: одна из двух клеток дает начало герминальным клеткам. У аскариды герминативные клетки или клетки «зародышевого пути» (А.Вейсман) выделяются на стадии 16 бластомеров, у дрозофилы — на стадии бластоцисты, у человека — первичные половые клетки (гонобласты) появляются на 3-ей неделе эмбрионального развития в стенке желточного мешка в каудальном отделе эмбриона.
Как и все клетки развивающегося организма, клетки зародышевого пути диплоидны. Они могут увеличиваться в числе путем обычного митоза, повторяя все стадии обычного клеточного цикла, где происходит чередование уровней количества ДНК и хромосом на клетку:
2n (2c) ® S-период® 4n (4c) ® 2 клетки 2n (2c) и т.д.
Однако на определенных стадиях развития при половом созревании особей этот обычный ход смены событий меняется. Герминативные клетки превращаются в гониальные (оогонии — женские и сперматогонии — мужские клетки — предшественники), и они вступают в процесс мейоза. При этом как для женских, так и для мужских клеток наступает первый цикл мейоза. На этой и следующей стадии половые клетки получили название сперматоцитов и ооцитов (I и II порядка).
В первом клеточном цикле мейоза происходит целый ряд событий, который его значительно отличает от обычного клеточного цикла. После вступления в I цикл созревания и сперматоциты I и ооциты I порядков синтезируют ДНК, её количеств удваивается, так же как удваивается за счет репликации количество хромосом. Следовательно, после S-периода эти клетки нужно считать (также как и соматические клетки после синтеза ДНК) тетраплоидными. После короткого G2-периода наступает профаза I мейотического деления, которая резко отличается от обычной мейотический профазы.
Особенности профазы I мейотического деления
Во-первых, эта стадия занимает большой отрезок времени (от суток до годов !). Во-вторых, она состоит из нескольких структурно-функциональных фаз (лептотена, зиготена, пахитена, диплотена, диакинез). Далее, в этот период происходит объединение, конъюгация, гомологичных (родительских) удвоенных в результате репликации хроматид, при этом образуются т.н. тетрады, т.е. хромосомные комплексы, состоящие из четырех хроматид (удвоенные материнские и удвоенные отцовские), которые соединены вместе с помощью специальной структуры — синаптинемного комплекса. В это же время происходит обмен участками между хроматидами гомологичных хромосом (но не между сестринскими хроматидами одного гомолога) — кроссинговер. Кроме того, в процессе конъюгации и обмена происходит синтез примерно 1,5% хромосомной ДНК.
В профазе I мейотического деления наблюдается рост объема ооцитов, в которых накапливаются запасные вещества, обеспечивающие ранние стадии развития будущего зародыша.
Эта профаза отличается также длительностью во времени, необходимого для прохождения перечисленных выше событий. Обычная соматическая профаза длится 0,5-1,5 часа. Мейотическая профаза сперматоцита I порядка у самцов мыши длится 12 суток, у человека — 24 дня (плюс еще около двух месяцев до полного созревания сперматозоида). Среди женских половых клеток профаза I порядка тритона обыкновенного длится около 1 года, у мыши от 4 месяцев до 3 лет, у человека профаза I ооцитов начинается на 3-ем месяце внутриутробного развития и может продолжаться до 50-летнего возраста женщины. При этом у человека происходит постепенная гибель заложенных ооцитов: у 3-х месячного эмбриона их около7х106 клеток, к рождению ребенка их остается около 2х106, к половому созреванию — 3х105, всего же созревает (овулируют) примерно 5х102 ооцитов.
Другой особенностью профазы I меойза является то, что в отличие от обычного митоза, хромосомы сохраняют ряд функциональных нагрузок, а именно: они способны к синтезу РНК, частичному синтезу ДНК, претерпевают ряд структурных перестроек. Другими словами, профазные хромосомы I мейотического деления не находятся в состоянии функционального покоя, а участвуют в целом ряде событий.
Стадии профазы I мейотического деления
Вся профаза I мейотического деления состоит из нескольких стадий: лептотена — стадия тонких нитей (хромосом), зиготена — стадия сливающихся (объединяющихся, конъюгирующих) нитей, пахитена — стадия толстых нитей, диплотена — стадия двойных нитей, диакинез — стадия расходящихся нитей.
Из всех стадий профазы I самой длительной является стадия пахитены, в ряде случаев она занимает до 50% времени.
Так, у человека при спермиогенезе стадии лептотены с зиготеной занимают 6,5 сут, пахатина- 15, диплотена и диакинез — 0,8; у мыши лептотена с зиготеной длятся около 3 суток, пахитена — 7 суток, диплотена с диакинезом — около 2 суток; у тритона лептотена занимает 5 сут, зиготена — 8, пахитена — 4-5, диплотена — 2 сут; у домашнего сверчка лептотена и зиготена занимают 2-3 сут, пахитена — 6-9, диплотена — 2. По сравнению с обычным митозом продолжительность деления клеток в процессе мейоза несравнимо длительнее. Это особенно наглядно видно при созревании женских половых клеток у животных, у которых яйцеклетки могут останавливаться в развитии на несколько месяцев и даже лет в стадии диплотены профазы I-го мейотического деления.
У растений мейоз также намного длиннее митоза по времени. Так, у традесканции весь мейоз занимает около 5 сут, из которых на профазу I-го деления приходится 4 сут, но встречаются виды, у которых мейоз идет со скоростью, соизмеримой с митозом.
Лептотена, или стадия тонких нитей, морфологически напоминает раннюю профазу митоза, но отличается тем, что при мейозе ядра обычно крупнее и хромосомы очень тонкие, так что проследить их по всей длине очень трудно.
Длина каждой мейотической хромосомы на ранних стадиях мейоза может быть в 10-100 раз больше длины соответствующих митотических хромосом. Следовательно, мейотические хромосомы имеют меньшую степень компактизации, они примерно в 30 раз менее компактны, чем хромосомы в метафазе мейоза. В лептотене хромосомы удвоены, но сестринские хроматиды в них далеко не всегда удается различить (так же как в хромосомах в ранней профазе митоза). Таким образом, в лептотене содержится диплоидное количество (2n) сдвоенных сестринских хроматид, общее количество последних, как и при митозе, равно 4n вследствие редупликации в S-периоде.
Расположение хромосом в лептотене часто повторяет телофазную поляризацию ядра. При этом у некоторых животных хромосомы образуют так называемую фигуру «букета» — дугообразно изогнутые сближенные хромосомы, связанные своими теломерами с ядерной оболочкой. У некоторых растений в конце лептотены хромосомы собираются в клубок (синезис).
Характерным для лептотены является появление на тонких хромосомах сгустков хроматина — хромомеров, которые как бы нанизаны в виде бусинок и располагаются по всей длине хромосомы. Число, размер и расположение таких хромомерных участков характерны для каждой хромосомы. Это позволяет составлять морфологические карты хромосом и использовать их для цитологического анализа. Число хромомеров различно у разных объектов: всего у тритона на 12 хромосомах их 2,5 тыс., у сверчка — около 200, у риса на 24 хромосомы — 645.
В лептотене начинает выявляться следующий, чрезвычайно важный и характерный для мейоза процесс конъюгации гомологичных хромосом, их сближение, которое начинается в теломерных участках, связанных с ядерной оболочкой. В этих местах образуется сложная специальная структура — тяж белковой природы, синаптонемный комплекс, который позже, в зиготене свяжет гомологичные удвоенные хроматиды по всей их длине.
Рекомбинация генетического материала.
Генетическая рекомбинация — это перераспределение генетического материала (ДНК), приводящее к возникновению новых комбинаций генов. Рекомбинация может происходить путем обмена клеточными ядрами, целыми молекулами ДНК или частями молекул. В то время как процессы репликации и репарации ДНК обеспечивают воспроизведение и сохранение генетического материала, рекомбинация приводит к генетической изменчивости. Биологическое значение рекомбинации столь велико, что она получила развитие у всех живых организмов. Она может происходить у эукариот (как при образовании половых клеток — гамет, так и в соматических клетках), у бактерий и даже при размножении вирусов, в том числе таких, генетический материал которых состоит из РНК. Перетасовка хромосом в мейозе, приводящая к огромному разнообразию гамет, случайность слияния гамет при оплодотворении, обмен частями между гомологичными хромосомами — все это (и далеко не только это) относится к рекомбинации.
Понятно, что из широкого круга рекомбинационных явлений интерес молекулярных биологов в первую очередь вызывает рекомбинация, заключающаяся в обменах частями между молекулами ДНК, ведь здесь можно применять весь арсенал методов генетики и молекулярной биологии, и эти исследования перекрываются с изучением других важных генетических процессов, прежде всего репликации и репарации ДНК. Но даже в таком виде, суженном до обменов частями молекул ДНК, понятие «рекомбинация» включает большой набор разных по своей природе явлений. При этом для всех рекомбинационных процессов общим является этап, на котором молекулы ДНК вступают в контакт в участке, где произойдет обмен полинуклеотидными цепями. Этот этап получил название «синапсис». Однако механизм синапсиса при разных типах рекомбинации принципиально различен. Более того, он является одним из критериев при классификации рекомбинационных явлений.
Прежде чем перейти к их рассмотрению, напомним некоторые термины и понятия, которыми мы будем пользоваться. Молекула ДНК представляет собой дуплекс — структуру из двух закрученных в спираль полинуклеотидных цепей. Последовательность нуклеотидов в цепях определяет специфичность ДНК и несет генетическую информацию. Молекулы, имеющие общее происхождение и состоящие из одинаковых нуклеотидных последовательностей, называют гомологичными. Однако их идентичность нарушается из-за мутаций, накапливающихся в течение поколений. По большей части мутации приводят к заменам единичных нуклеотидов, реже к выпадениям и вставкам отдельных нуклеотидов. Поэтому нарушения гомологии в результате мутаций не очень существенны по сравнению с основной массой идентичных нуклеотидов, и в таких случаях можно говорить об общей гомологии молекул. Каждая новая мутация приводит к образованию нового аллеля в том гене, где она возникла. Следовательно, новые аллели обычно отличаются от исходной формы одним нуклеотидом. Если мутация приводит к изменению фенотипа у исходной формы, то к ней можно применять также термин «генетический маркер».
Две цепи, составляющие дуплекс ДНК, антипараллельны, то есть имеют разную полярность: одна цепь имеет направление 5'-3', другая — 3'-5'. Цепи удерживаются вместе водородными связями между парами комплементарных оснований А-Т и G-C. Поэтому обе цепи в дуплексе являются также комплементарными. Процесс расхождения цепей в результате разрыва водородных связей есть денатурация, обратная реакция — ренатурация. Все это сказано для того, чтобы подвести читателя к отправной идее статьи: поскольку отдельные цепи ДНК, полученные от разных родителей, гомологичны и, следовательно, комплементарны, они могут ренатурировать, формируя новый дуплекс. Иными словами, гомологичные ДНК могут узнавать друг друга по комплементарности их нуклеотидной последовательности. Новый дуплекс, состоящий из цепей от разных молекул, называется гетеродуплексом.
А теперь можно дать классификацию основных типов рекомбинации. Все, что говорилось о гомологии ДНК и комплементарности полинуклеотидных цепей, относится к гомологичной, или общей, рекомбинации (она же кроссинговер), основанной на спаривании комплементарных цепей ДНК. От других типов рекомбинационных процессов ее отличают необходимость в общей (по всей длине молекул) гомологии между рекомбинирующими ДНК и участие большого набора специальных белков. Гомологичная рекомбинация начинается с возникновения в одном или обоих дуплексах участков из одиночных цепей ДНК, которые затем с помощью специальных белков находят комплементарные последовательности в гомологичном дуплексе и образуют с ними гетеродуплекс — ключевой промежуточный продукт (интермедиат) рекомбинации. Конечным результатом рекомбинации будет обмен равными частями гомологичных молекул (рис. 1).
Из общей рекомбинации можно выделить как частный случай так называемую эктопическую рекомбинацию. Она заключается в обменах (кроссинговерах) между отдельными участками гомологичной ДНК, разбросанными по геному. К ним относятся разнообразные подвижные элементы, названные так за способность перемещаться по геному, гены транспортных и рибосомных РНК, гистонов и многие другие повторяющиеся последовательности (повторы) ДНК. Такая локальная гомологичная рекомбинация интересна прежде всего тем, что она может приводить к хромосомным перестройкам, хотя ее биологическая роль этим не исчерпывается. На рис. 2 в качестве примера приведены схемы возникновения инверсий (поворотов внутренних участков хромосом на 180?), утрат (делеций) и удвоений (дупликаций) частей хромосом в результате эктопической рекомбинации. Это только часть возможных перестроек хромосом. Другие их типы могут возникать в зависимости от того, какова ориентация повторов ДНК по отношению друг к другу (прямая или обратная), и от того, где они расположены: внутри одной хромосомы, в сестринских хроматидах или разных хромосомах. Несмотря на то, что обмены происходят между локальными участками гомологии, эктопическая рекомбинация осуществляется в основном теми же белками, что и гомологичная. Принципиально иными являются три других типа рекомбинации, которые основаны не на взаимодействии комплементарных цепей ДНК, а на совершенно иных механизмах и участии иных белков.
Биологическое значение гомологичной рекомбинации огромно. Прежде всего она вносит большой вклад в лежащую в основе эволюции генетическую изменчивость, позволяющую организмам постоянно приспосабливаться к среде обитания. Преимущества перекомбинаций генов настолько велики, что рекомбинационные системы появились у вирусов и бактерий, которые размножаются вегетативно. У эукариот они достигли большего разнообразия и сложности, особенно в соматических клетках. Эктопическая рекомбинация приводит к перестройкам хромосом, с которыми (прежде всего с дупликациями) связывают эволюцию генетического аппарата. Считается, что дупликации участков хромосом обеспечили материал для дивергенции нуклеотидных последовательностей, приводящей к возникновению новых генов.
Однако биологическое значение гомологичной, и в том числе эктопической, рекомбинации нельзя свести к их роли в эволюции. Большую роль они играют и в разнообразных онтогенетических перестройках генетического материала, участвующих в регуляции работы генов. Например, конверсия гена (коррекция гетеродуплекса), которая в мейотических клетках является одним из этапов общего процесса кроссинговера, в соматических клетках эукариот и клетках бактерий может не сопровождаться кроссинговером по внешним генам и выступать как самостоятельное явление. Такая конверсия выполняет важные функции в онтогенезе бактерий, дрожжей, животных. Известно много примеров, когда определенный ген расположен в локусе, где он имеет собственный промотор и может функционировать, в то время как в других локусах находятся последовательности, в основном гомологичные этому гену, но заметно отличающиеся по нуклеотидному составу из-за накопившихся в них мутаций. Они лишены промотора и не могут выполнять функции генов. Эти «молчащие» последовательности могут вступать в синапсис с работающим геном и служить матрицей для его конверсии. Таким образом, работающий ген может менять свою нуклеотидную последовательность. Подобным способом клетки гомоталличных штаммов дрожжей меняют свой половой тип.
У некоторых патогенных микроорганизмов этот же механизм, позволяющий их клеткам менять свои поверхностные антигены, участвует в процессах, описанных ниже. Так, многие патогенные бактерии (спирохета Borrelia bormsei, гонококки и др.) и простейшие (африканские трипаносомы), с одной стороны, и животные, в которых они паразитируют, — с другой, используют в борьбе друг против друга в сущности сходные приемы. Животные продуцируют в огромном ассортименте антитела, обеспечивающие им иммунитет, а патогенные микроорганизмы в ответ на это образуют на своей поверхности все новые и новые антигены, позволяющие им уходить от иммунного ответа хозяйского организма. В основе данных процессов лежат рекомбинационные перестройки в локусах, кодирующих антигены (или антитела). Рекомбинационные перестройки включают одни и выключают другие гены либо создают новые гены. В этих сложных процессах участвуют разные типы рекомбинации, но гомологичная и эктопическая рекомбинации (и в том числе конверсия гена) играют здесь не последнюю роль. Помимо описанных процессов у бактерий и низших эукариот известны и другие рекомбинационные системы, участвующие в регуляции работы генов. Но это тема следующей статьи.
Список литературы:
Слюсарев А.А., Жукова С.В. Биология – Киев. Вища школа. 1987
Лобашов М.Е. Генетика – Л. Изд. Ленинградского унив., 1967
Биология.: Учебник для мед. спец. ВУЗ-ов. Под ред. В.Н. Ярыгина М., Высшая Школа, 1997
Албертс Б., Брей Д., Льюис Дж. и др. Молекулярная биология клетки:
Пер. с англ. М.: Мир, 1994. Т. 1, ч. 2. С. 301-310.
Инге-Вечтомов С.Г. Введение в молекулярную генетику. М.: Высш. шк., 1983. С. 120-136.
Льюин Б. Гены. Пер. с англ. М.: Мир, 1987. С. 443-453.
www.ronl.ru
Реферат - Мейоз, его цитогенетические механизмы и биологическое значение.
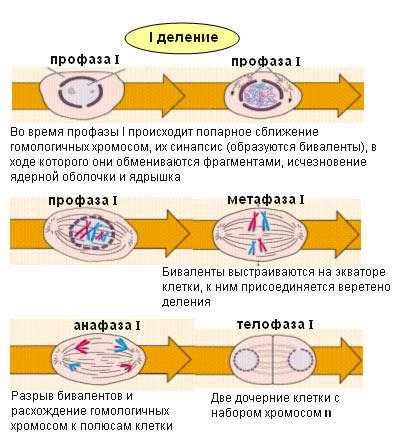
Мейоз — процесс деления клеточного ядра с образованием четырех дочерних ядер, каждое из которых содержит вдвое меньше хромосом, чем исходное ядро. Мейоз — редукционное деление: происходит уменьшение числа хромосом в клетке с диплоидного (2n) до гаплоидного (n). Мейоз сопровождает образование гамет у животных и образование спор у растений. В результате мейоза получаются гаплоидные ядра, при слиянии которых во время оплодотворения восстанавливается диплоидный набор хромосом Мейоз включает два последовательных деления. В каждом мейотическом делении выделяют четыре стадии: профазу, метафазу, анафазу и телофазу.Первое мейотическое деление называют редукционным. В результате из одной клетки с диплоидным набором хромосом образуются две с гаплоидным набором.
Профаза I — профаза первого мейотического деления – самая продолжительная. Ее условно делят на пять стадий: лептотену, зиготену, пахитену, диплотену и диакинез.
Первая стадия — лептотена — характеризуется увеличением ядра. В ядре виден диплоидный набор хромосом. Хромосомы представляют собой длинные, тонкие
нити. Каждая хромосома состоит из двух хроматид. Хроматиды имеют хромомерное строение. Начинается спирализация хромосом.
Во время второй стадии профазы 1 — го мейотического деления — зиготене — происходит конъюгация гомологичных хромосом. Гомологичными называют хромосомы, имеющие одинаковую форму и размер: одна из них получена от матери, другая от отца. Гомологичные хромосомы притягиваются и прикладываются друг к другу по всей длине. Центромера одной из парных хромосом точно прилегает к центромере другой, и каждая хроматида прилегает к гомологичной хроматиде другой.
Третья стадия — пахитена — стадия толстых нитей. Конъюгирующие хромосомы тесно прилегают друг к другу. Такие сдвоенные хромосомы называют бивалентами. Каждый бивалент состоит из четверки (тетрады) хроматид. Число бивалентов равно гаплоидному набору хромосом. Происходит дальнейшая спирализация. Тесный контакт между хроматидами дает возможность обмениваться идентичными участками в гомологичных хромосомах. Это явление называется кроссинговер
Четвертая стадия — диплотена — характеризуется возникновением сил отталкивания. Хромосомы, составляющие биваленты, начинают отходить друг от друга. Расхождение начинается в области центромер. Хромосомы соединены между собой в нескольких точках. Эти точки называют хиазмами т. е. местами, где произойдет кроссинговер. В каждой хиазме
осуществляется обмен участками хроматид. Хромосомы спирализуются и укорачиваются.
Пятая стадия — диакинез — характеризуется максимальной спирализацией, укорочением и утолщением хромосом. Отталкивание хромосом продолжается, но они остаются соединенными в биваленты своими концами. Ядрышко и ядерная оболочка растворяются. Центриоли расходятся к полюсам. Таким образом, в профазе 1 — го мейотического деления происходят три основных процесса:
1) конъюгация гомологичных хромосом;
2) образование бивалентов хромосом или тетрад хроматид;
3) кроссинговер.
Метафаза I. В метафазе первого мейотического деления биваленты хромосом
располагаются по экватору клетки, образуя метафазную пластинку. К ним
прикрепляются нити веретена деления. Анафаза I. В анафазе первого мейотического деления к полюсам клетки расходятся хромосомы, а не хроматиды. В дочерние клетки попадают только по одной из пары гомологичных хромосом.
Телофаза I. В телофазе первого мейотического деления число хромосом в каждой клетке становится гаплоидным. Хромосомы состоят из двух хроматид. Вследствие кроссинговера при образовании хиазм, хроматиды генетически не однородны. На короткое время образуется ядерная оболочка, хромосомы деспирализуются, ядро становится интерфазным. Затем у животной клетки начинается деление цитоплазмы, а у растительной клетки формирование клеточной стенки. У многих растений нет телофазы I, клеточная стенка не образуется, нет интерфазы II, клетки сразу переходят из анафазы I в профазу II.
Интерфаза II. Эта стадия есть только у животных клеток. Во время интерфазы
между первым и вторым делением в S период не происходит редупликация молекул
ДНК.
Второе мейотическое деление называют эквационным. Оно похоже на митоз.
Из хромосом, имеющих две хроматиды, образуются хромосомы, состоящие из одной
хроматиды.
Профаза II. В профазе второго мейотического деления хромосомы утолщаются и укорачиваются. Ядрышко и ядерная оболочка разрушаются. Образуется веретено деления.
Метафаза II. В метафазе второго мейотического деления хромосомы выстраиваются вдоль экватора. Нити ахроматинового веретена отходят к полюсам. Образуется метафазная пластинка.
Анафаза II. В анафазе второго мейотического деления центромеры делятся и тянут за собой к противоположным полюсам отделившиеся друг от друга хроматиды, называемые хромосомами.
Телофаза II, В телофазе второго мейотического деления хромосомы деспирализуются, становятся невидимыми. Нити веретена исчезают. Вокруг ядер формируется ядерная оболочка. Ядра содержат гаплоидный набор хромосом. Происходит деление цитоплазмы и образование клеточной стенки у растений. Из
одной исходной клетки образуются четыре гаплоидных клетки.
ЗНАЧЕНИЕ МЕЙОЗА
1. Поддержание постоянства числа хромосом. Если бы не возникало редукции числа хромосом при гаметогенезе, и половые клетки имели гаплоидный набор хромосом, то из поколения в поколение возрастало бы их число.
2. При мейозе образуется большое число новых комбинаций негомологичных
хромосом.
3. В процессе кроссинговера имеют место рекомбинации генетического
материала. Практически все хромосомы, попадающие в гаметы, содержат участки,
происходящие как первоначально от отцовской, так и от материнской хромосомы.
Этим достигается большая степень перекомбинации наследственного материала. В
этом одна из причин изменчивости организмов, дающая материал для отбора.
ОТЛИЧИЯ МИТОЗА ОТ МЕЙОЗА
При митозе в профазе нет конъюгации гомологичных хромосом и
кроссинговера
Удвоение хромосом соответствует каждому делению клетки. В метафазе при митозе на экваторе выстраиваются хромосомы, состоящие из двух хроматид. В анафазе при митозе к полюсам расходятся хроматиды. В телофазе дочерние клетки содержат то же число хромосом, что и материнские. При мейозе в профазе I происходит конъюгация гомологичных хромосом, имеет место кроссинговер. Образуются биваленты хромосом.
В метафазе I при мейозе на экваторе располагаются биваленты хромосом. При мейозе в анафазе I к полюсам расходятся хромосомы, состоящие из двух
хроматид. В телофазе I мейоза число хромосом в дочерних клетках вдвое меньше, чем в
материнских. Между I и II делениями мейоза в интерфазе нет синтеза ДНК. Мейоз
осуществляется в диплоидных и полиплоидных клетках. В результате мейоза из одной клетки образуются четыре гаплоидных. Мейоз у человека имеет место во время овогенеза и сперматогенеза.
www.ronl.ru
МЕЙОЗ МЕЙОЗ - Реферат - Мейоз
Реферат - Мейоз (95.1 kb.)Доступные файлы (1):содержаниеМЕЙОЗ.doc
- МЕЙОЗ


МЕЙОЗ
Половое размножение организмов осуществляется с помощью специализированных клеток, т.н. гамет, – яйцеклетки (яйца) и спермия (сперматозоида). Гаметы, сливаясь, образуют одну клетку – зиготу. Каждая гамета гаплоидна, т.е. имеет по одному набору хромосом. Внутри набора все хромосомы разные, однако каждой хромосоме яйцеклетки соответствует одна из хромосом спермия. Зигота, таким образом, содержит уже пару таких соответствующих друг другу хромосом, которые называют гомологичными. Гомологичные хромосомы сходны, поскольку имеют одни и те же гены или их варианты (аллели), определяющие специфические признаки. Например, одна из парных хромосом может иметь ген, кодирующий группу крови А, а другая – его вариант, кодирующий группу крови В. Хромосомы зиготы, происходящие из яйцеклетки, являются материнскими, а происходящие из спермия – отцовскими.
В результате многократных митотических делений из образовавшейся зиготы возникает либо многоклеточный организм, либо многочисленные свободноживущие клетки, как это происходит у обладающих половым размножением простейших и у одноклеточных водорослей.
При образовании гамет диплоидный набор хромосом, имевшийся у зиготы, должен наполовину уменьшиться (редуцироваться). Если бы этого не происходило, то в каждом поколении слияние гамет приводило бы к удвоению набора хромосом. Редукция до гаплоидного числа хромосом происходит в результате редукционного деления – т.н. мейоза, который представляет собой вариант митоза.
(23.48 Кб)
^ Особенность мейоза состоит в том, что при клеточном делении экваториальную пластинку образуют пары гомологичных хромосом, а не удвоенные индивидуальные хромосомы, как при митозе. Парные хромосомы, каждая из которых осталась одинарной, расходятся к противоположным полюсам клетки, клетка делится, и в результате дочерние клетки получают половинный, по сравнению с зиготой, набор хромосом.
Для примера предположим, что гаплоидный набор состоит из двух хромосом. В зиготе (и соответственно во всех клетках организма, продуцирующего гаметы) присутствуют материнские хромосомы А и В и отцовские А' и В'. Во время мейоза они могут разделиться следующим образом:
Наиболее важен в этом примере тот факт, что при расхождении хромосом вовсе не обязательно образуется исходный материнский и отцовский набор, а возможна рекомбинация генов, как в гаметах АВ' и А'В в приведенной схеме.
Теперь предположим, что пара хромосом АА' содержит два аллеля – a и b – гена, определяющего группы крови А и В. Сходным образом пара хромосом ВВ' содержит аллели m и n другого гена, определяющего группы крови M и N. Разделение этих аллелей может идти следующим образом:
Очевидно, что получившиеся гаметы могут содержать любую из следующих комбинаций аллелей двух генов: am, bn, bm или an.
Если имеется большее число хромосом, то пары аллелей будут расщепляться независимо по тому же принципу. Это означает, что одни и те же зиготы могут продуцировать гаметы с различными комбинациями аллелей генов и давать начало разным генотипам в потомстве.
^ Оба приведенных примера иллюстрируют принцип мейоза. На самом деле мейоз – значительно более сложный процесс, так как включает два последовательных деления. Главное в мейозе то, что хромосомы удваиваются только один раз, тогда как клетка делится дважды, в результате чего происходит редукция числа хромосом и диплоидный набор превращается в гаплоидный.
Во время профазы первого деления гомологичные хромосомы конъюгируют, т. е. сближаются попарно. В результате этого очень точного процесса каждый ген оказывается напротив своего гомолога на другой хромосоме. Обе хромосомы затем удваиваются, но хроматиды остаются связанными одна с другой общей центромерой.
В метафазе четыре соединенные хроматиды выстраиваются, образуя экваториальную пластинку, как если бы они были одной удвоенной хромосомой. В противоположность тому, что происходит при митозе, центромеры не делятся. В результате каждая дочерняя клетка получает пару хроматид, все еще связанных цетромерой. Во время второго деления хромосомы, уже индивидуальные, опять выстраиваются, образуя, как и в митозе, экваториальную пластинку, но их удвоения при этом делении не происходит. Затем центромеры делятся, и каждая дочерняя клетка получает одну хроматиду.
^ В результате двух мейотических делений диплоидной клетки образуются четыре клетки. При образовании мужских половых клеток получается четыре спермия примерно одинаковых размеров. При образовании же яйцеклеток деление цитоплазмы происходит очень неравномерно: одна клетка остается крупной, тогда как остальные три настолько малы, что их почти целиком занимает ядро. Эти мелкие клетки, т.н. полярные тельца, служат лишь для размещения избытка хромосом, образовавшихся в результате мейоза. Основная часть цитоплазмы, необходимой для зиготы, остается в одной клетке – яйцеклетке.
^ Во время конъюгации хроматиды гомологичных хромосом могут разрываться и затем соединяться в новом порядке, обмениваясь участками следующим образом:
Этот обмен участками гомологичных хромосом называется кроссинговером (перекрестом). Как показано выше, кроссинговер ведет к возникновению новых комбинаций аллелей сцепленных генов. Так, если исходные хромосомы имели комбинации АВ и ab, то после кроссинговера они будут содержать Ab и aB. Этот механизм появления новых генных комбинаций дополняет эффект независимой сортировки хромосом, происходящей в ходе мейоза. Различие состоит в том, что кроссинговер разделяет гены одной и той же хромосомы, тогда как независимая сортировка разделяет только гены разных хромосом.
www.studmed.ru
Реферат - Мейоз как центральный механизм гаметогенеза, цитологическая и цитогенетическая характеристики. Биологическое значение мейоза.
Мейоз – это процесс деления специализированных клеток приводящей к редукции числа хромосом вдвое. Происходит в процессе спорогенеза у растений и гаметогенеза у животных, у растений был открыт в 1888 г. Стразбургером, а у животных в 1882 г. Флемингом.
Мейоз состоит из двух быстро следующих одно за другим делений, происходящих в периоде созревания. Удвоение ДНК для этих делений осуществляется однократно в периоде роста. Второе деление мейоза следует за первым практически сразу так, что наследственный материал не синтезируется в промежутке между ними.
Первое мейотическое деление называют редукционным, так как оно приводит к образованию из диплоидных клеток (2n2с) гаплоидных клеток п2с. Такой результат обеспечивается благодаря особенностям профазы первого деления мейоза. В профазе I мейоза, так же как в обычном митозе, наблюдается компактная упаковка генетического материала (спирализация хромосом). Одновременно происходит событие, отсутствующее в митозе: гомологичные хромосомы конъюгируют друг с другом, т.е. тесно сближаются соответствующими участками.
В результате конъюгации образуются хромосомные пары, или биваленты, числом п. Так как каждая хромосома, вступающая в мейоз, состоит из двух хроматид, то бивалент содержит четыре хроматиды. Формула генетического материала в профазе I остается 2n4c. К концу профазы хромосомы в бивалентах, сильно спирализуясь, укорачиваются. Так же как в митозе, в профазе I мейоза начинается формирование веретена деления, с помощью которого хромосомный материал будет распределяться между дочерними клетками. Отцовские хромосомы обозначены черным цветом, материнские —неокрашснные. На рисунке не изображена метафаза I, в которой биваленты располагаются в плоскости экватора веретена деления, и телофаза I, быстро переходящая в профазу II.
Профаза I самая продолжительная и подразделяется на 5 стадий:
Ø Лептотена – конденсация, т.е. набухание хромосомы
Ø Зиготена – конъюгация
Ø Пахитена – кроссинговер
Ø Диплотена – образуются хиазмы – это мостики между гомологичными хромосомами
Ø Диакинез – соединение хроматид
Таким образом, несмотря на возникающие между гомологичными хромосомами силы отталкивания, в профазе I не происходит окончательного разрушения бивалентов. Особенностью мейоза в овогенезе является наличие специальной стадии — диктиотены, отсутствующей в сперматогенезе. На этой стадии, достигаемой у человека еще в эмбриогенезе, хромосомы, приняв особую морфологическую форму «ламповых щеток», прекращают какие-либо дальнейшие структурные изменения на многие годы. По достижении женским организмом репродуктивного возраста под влиянием лютеинизирующего гормона гипофиза, как правило, один овоцит ежемесячно возобновляет мейоз.
В метафазе I мейоза завершается формирование веретена деления. Его нити прикрепляются к центромерам хромосом, объединенных в биваленты, таким образом, что от каждой центромерыидет лишь одна нить к одному из полюсов веретена. В результате нити, связанные с центромерами гомологичных хромосом, направляясь к разным полюсам, устанавливают бивалентны в плоскости экватора веретена деления.
В телофазе I мейоза у полюсов веретена собирается одинарный, гаплоидный набор хромосом, каждая из них содержит удвоенное количество ДНК.
Формула генетического материала образующихся дочерних клеток соответствует п2с.
Второе мейотическое (эквационное) деление приводит к образованию клеток, в которых содержание генетического материала в хромосомах будет соответствовать их однонитчатой структуре nс. Это деление протекает, как митоз, только клетки, вступающие в него, несут гаплоидный набор хромосом. В процессе такого деления материнские двунитчатые хромосомы, расщепляясь, образуют дочерние однонитчатые.
Одна из главных задач мейоза — создание клеток с гаплоидным набором однонитчатых хромосом —достигается благодаря однократной редупликации ДНК для двух последовательных делений мейоза, а также благодаря образованию в начале первого мейотического деления пар гомологичных хромосом и дальнейшего их расхождения в дочерние клетки.
Процессы, протекающие в редукционном делении, обеспечивают также не менее важное следствие — генетическое разнообразие гамет, образуемых организмом. К таким процессам относят кроссинговер, расхождение гомологичных хромосом в разные гаметы и независимое поведение бивалентов в первом мейотическом делении.
Кроссинговер обеспечивает перекомбинацию отцовских и материнских аллелей в группах сцепления. Ввиду того что перекрест хромосом может происходить в разных участках, кроссинговер в каждом отдельном случае приводит к обмену разным по количеству генетическим материалом. Необходимо отметить также возможность возникновения нескольких перекрестов между двумя хроматидами и участия в обмене более чем двух хроматид бивалента. Отмеченные особенности кроссинговера делают этот процесс эффективным механизмом перекомбинации аллелей.
Расхождение гомологичных хромосом в разные гаметы в случае гетерозиготности приводит к образованию гамет, различающихся по аллелям отдельных генов.
Случайное расположение бивалентов в плоскости экватора веретена деления и последующее их расхождение в анафазе I мейоза обеспечивают перекомбинацию родительских групп сцепления в гаплоидном наборе гамет.
Биологическое значение мейоза:
Ø Обеспечивает генетическую, комбинативную изменчивость.
Ø Способствует формированию гаплоидных клеток (спор, гамет).
www.ronl.ru



|
|
..:::Счетчики:::.. |
|
|
|
|
|
|
|
|