
|
|
|
|
 Far Far |
| WinNavigator |
| Frigate |
| Norton
Commander |
| WinNC |
| Dos
Navigator |
| Servant
Salamander |
| Turbo
Browser |
|
|
| Winamp,
Skins, Plugins |
| Необходимые
Утилиты |
| Текстовые
редакторы |
| Юмор |
|
|

|
File managers and best utilites |
Клетки прокариот и эукариот. Реферат эукариоты
Реферат Эукариоты
скачатьРеферат на тему:
План:
- Введение
- 1 Строение эукариотической клетки
- 2 Деление на царства
- 3 Отличия эукариот от прокариот
- 4 Источники, ссылки
Введение
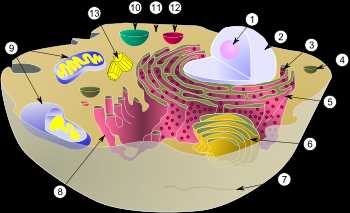
Диаграмма типичной клетки животного. Отмеченные органоиды (органеллы) 1. Ядрышко 2. Ядро 3. Рибосома 4. Везикула 5. Шероховатый (гранулярный) эндоплазматический ретикулум 6. Аппарат Гольджи 7. Клеточная стенка 8. Гладкий (агранулярный) эндоплазматический ретикулум 9. Митохондрия 10. Вакуоль 11. Гиалоплазма 12. Лизосома 13. Центросома (Центриоль)
Эукарио́ты, или Я́дерные (лат. Eukaryota от греч. εύ- — хорошо и κάρυον — ядро) — домен (надцарство) живых организмов, клетки которых содержат ядра. Все организмы, кроме бактерий и архей, являются ядерными (вирусы и вироиды также не являются эукариотами, но не все биологи считают их живыми организмами).
Животные, растения, грибы, а также группы организмов под общим названием протисты — все являются эукариотическими организмами. Они могут быть одноклеточными и многоклеточными, но все имеют общий план строения клеток. Считается, что все эти столь несхожие организмы имеют общее происхождение, поэтому группа ядерных рассматривается как монофилетический таксон наивысшего ранга. Согласно наиболее распространённым гипотезам, эукариоты появились 1,5–2 млрд. лет назад. Важную роль в эволюции эукариот сыграл симбиогенез — симбиоз между эукариотической клеткой, видимо, уже имевшей ядро и способной к фагоцитозу, и проглоченными этой клеткой бактериями — предшественниками митохондрий и пластидов.
1. Строение эукариотической клетки
Эукариотические клетки в среднем намного крупнее прокариотических, разница в объёме достигает тысяч раз. Клетки эукариот включают около десятка видов различных структур, известных как органоиды (или органеллы, что, правда, несколько искажает первоначальное значение этого термина), из которых многие отделены от цитоплазмы одной или несколькими мембранами. В прокариотических клетках всегда присутствуют клеточная мембрана, рибосомы (существенно отличные от эукариотических рибосом) и генетический материал — бактериальная хромосома, или генофор, однако внутренние органоиды, окруженные мембраной, встречаются редко. Ядро — это часть клетки, окружённая у эукариот двойной мембраной (двумя элементарными мембранами) и содержащая генетический материал: молекулы ДНК, «упакованные» в хромосомы. Ядро обычно одно, но бывают и многоядерные клетки.
2. Деление на царства
Существует несколько вариантов деления надцарства эукариот на царства. Первыми были выделены царства растений и животных. Затем было выделено царство грибов, которые из-за биохимических особенностей, по мнению большинства биологов, не могут быть причислены ни к одному из этих царств. Также некоторые авторы выделяют царства простейших, миксомицетов, хромистов. Некоторые системы насчитывают до 20 царств. По системе Томаса Кавалир-Смита все эукариоты подразделяются на два монофилетических таксона — Unikonta и Bikonta.
3. Отличия эукариот от прокариот
Важнейшая, основополагающая особенность эукариотических клеток связана с расположением генетического аппарата в клетке. Генетический аппарат всех эукариот находится в ядре и защищён ядерной оболочкой (по-гречески «эукариот» значит имеющий ядро). ДНК эукариот линейная (у прокариот ДНК кольцевая и находится в особой области клетки — нуклеоиде, который не отделён мембраной от остальной цитоплазмы). Она связана с белками-гистонами и другими белками хромосом, которых нет у бактерий.
В жизненном цикле эукариот обычно присутствуют две ядерные фазы (гаплофаза и диплофаза). Первая фаза характеризуется гаплоидным (одинарным) набором хромосом, далее, сливаясь, две гаплоидные клетки (или два ядра) образуют диплоидную клетку (ядро), содержащую двойной (диплоидный) набор хромосом. Иногда при следующем делении, а чаще спустя несколько делений клетка вновь становится гаплоидной. Такой жизненный цикл и в целом диплоидность для прокариот не характерны.
Третье, пожалуй, самое интересное отличие, — это наличие у эукариотических клеток особых органелл, имеющих свой генетический аппарат, размножающихся делением и окружённых мембраной. Эти органеллы — митохондрии и пластиды. По своему строению и жизнедеятельности они поразительно похожи на бактерий. Это обстоятельство натолкнуло современных учёных на мысль, что подобные организмы являются потомками бактерий, вступившими в симбиотические отношения с эукариотами. Прокариоты характеризуются малым количеством органелл, и ни одна из них не окружена двойной мембраной. В клетках прокариот нет эндоплазматического ретикулума, аппарата Гольджи, лизосом.
Ещё одно важное различие между прокариотами и эукариотами — наличие у эукариот эндоцитоза, в том числе у многих групп — фагоцитоза. Фагоцитозом (дословно «поедание клеткой») называют способность эукариотических клеток захватывать, заключая в мембранный пузырёк, и переваривать самые разные твёрдые частицы. Этот процесс обеспечивает в организме важную защитную функцию. Впервые он был открыт И. И. Мечниковым у морских звёзд. Появление фагоцитоза у эукариот скорее всего связано со средними размерами (далее о размерных различиях написано подробнее). Размеры прокариотических клеток несоизмеримо меньше, и поэтому в процессе эволюционного развития эукариот у них возникла проблема снабжения организма большим количеством пищи. Как следствие среди эукариот появляются первые настоящие, подвижные хищники.
Большинство бактерий имеет клеточную стенку, отличную от эукариотической (далеко не все эукариоты имеют её). У прокариот это прочная структура, состоящая главным образом из муреина (у архей из псевдомуреина). Строение муреина таково, что каждая клетка окружена особым сетчатым мешком, являющимся одной огромной молекулой. Среди эукариот клеточную стенку имеют многие протисты, грибы и растения. У грибов она состоит из хитина и глюканов, у низших растений — из целлюлозы и гликопротеинов, диатомовые водоросли синтезируют клеточную стенку из кремниевых кислот, у высших растений она состоит из целлюлозы, гемицеллюлозы и пектина. Видимо, для более крупных эукариотических клеток стало невозможно создавать клеточную стенку из одной молекулы высокую по прочности. Это обстоятельство могло заставить эукариот использовать иной материал для клеточной стенки. Другое объяснение состоит в том, что общий предок эукариот в связи с переходом к хищничеству утратил клеточную стенку, а затем были утрачены и гены, отвечающие за синтез муреина. При возврате части эукариот к осмотрофному питанию клеточная стенка появилась вновь, но уже на другой биохимической основе.
Разнообразен и обмен веществ у бактерий. Вообще всего выделяют четыре типа питания, и среди бактерий встречаются все. Это фотоавтотрофные, фотогетеротрофные, хемоавтотрофные, хемогетеротрофные (фототрофные используют энергию солнечного света, хемотрофные используют химическую энергию). Эукариоты же либо сами синтезируют энергию из солнечного света, либо используют готовую энергию такого происхождения. Это может быть связано с появлением среди эукариотов хищников, необходимость синтезировать энергию для которых отпала.
Ещё одно отличие — строение жгутиков. У бактерий они тонкие — всего 15–20 нм в диаметре. Это полые нити из белка флагеллина. Строение жгутиков эукариот гораздо сложнее. Они представляют собой вырост клетки, окруженный мембраной, и содержат цитоскелет (аксонему) из девяти пар периферических микротрубочек и двух микротрубочек в центре. В отличие от вращающихся прокариотическох жгутиков жгутики эукариот изгибаются или извиваются. Две группы рассматриваемых нами организмов, как уже было сказано, сильно отличаются и по своим средним размерам. Диаметр прокариотической клетки составляет обычно 0,5–10 мкм, когда тот же показатель у эукариот составляет 10–100 мкм. Объём такой клетки в 1000–10000 раз больше, чем прокариотической. У прокариот рибосомы мелкие (70S-типа). У эукариот рибосомы более крупные (80S-типа).
Видимо, различается и время возникновения этих групп. Первые прокариоты возникли в процессе эволюции около 3,5 млрд. лет назад, от них около 1,2 млрд. лет назад произошли эукариотические организмы.
4. Источники, ссылки
- Галицкий В. А. Возникновение эукариотических клеток и происхождение апоптоза // Цитология, 2005, том 47, вып. 2, с. 103—120.
- Биологический энциклопедический словарь / под редакцией М. С. Гилярова. — М., 1989.
- Мирабдуллаев И. М. Проблема происхождения эукариот // Успехи совр. биол. 1989а. Т. 107. С. 341—356.
- Марков А. В. Проблема происхождения эукариот // Палеонтологический журнал 2 (2005): 3—12.
- Подборка статей по проблеме происхождения эукариот
- Вся Биология — Эукариоты
- Б. М. Медников. Биология: формы и уровни жизни. — Просвещение, 1995.
- Д. Тейлор, Н. Грин, У. Стаут. Биология (в трёх томах)
wreferat.baza-referat.ru
Клетки прокариот и эукариот | Цитология. Реферат, доклад, сообщение, кратко, презентация, лекция, шпаргалка, конспект, ГДЗ, тест
Форма и размеры клеток очень разнообразны. Несмотря на их многообразие, встречается лишь два типа структурной организации клеток. К одним относятся клетки бактерий, цианобактерий и архебактерий — это прокариоты (лат. pro — «перед», «раньше»; греч. karion — «ядро»). К другим относятся клетки грибов, растений и животных — это эукариоты (лат. eu — «полностью», «хорошо»; греч. karion — «ядро»). Прокариоты обладают более простым строением, а эукариоты — более сложным и разнообразным.
Прокариотические клетки свойственны древним одноклеточным организмам. Лишь некоторые виды бактерий и цианобактерий образуют небольшие цепочки или комочки из нескольких клеток. Эукариотические клетки свойственны и одноклеточным, и различным многоклеточным организмам. Разность между прокариотами и эукариотами столь велика, что послужила основанием разделить их на два надцарства: Доядерные организмы, или Прокариоты, и Ядерные организмы, или Эукариоты.
Клетки прокариот имеют ядерное вещество и одну хромосому (кольцевую нить ДНК), но в них нет оформленного ядра. ДНК-содержащую зону в клетке прокариот называют нуклеоидом (лат. nucleus — «ядро»; греч. eidos — «вид»), т. е. похожим на ядро. Обычно нуклеоид находится в центре клетки, но он не отграничен мембранами от цитоплазмы.
Клетки прокариот отличаются очень малыми размерами (от 0,5 до 5 мкм) и простейшим строением (рис. 36). Они имеют неподвижную цитоплазму, плазматическую мембрану и клеточную стенку. Цитоплазма содержит немного мелких рибосом и различные включения в виде гранул липидов и других веществ. Генетический материал (ДНК) не отделён мембранами от цитоплазмы, нет и оформленных хромосом, а «хромосомой» условно называют единственную кольцевую молекулу ДНК.
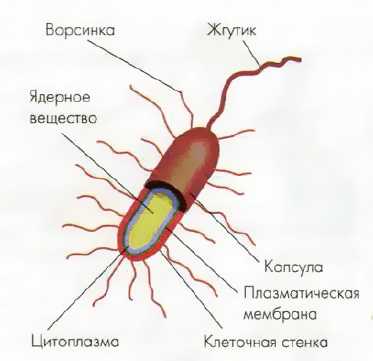 |
| Рис. 36. Схема строения клетки прокариот (бактерии) |
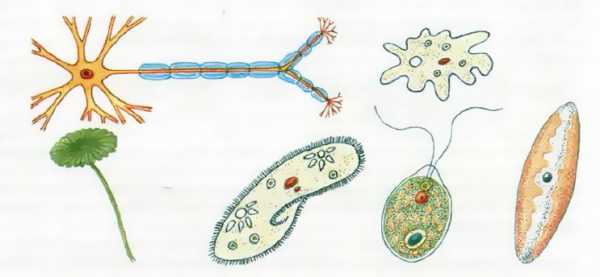 |
| Рис. 37. Разнообразие форм клеток эукариот |
Клетки эукариот являются очень сложными единицами живой природы и характеризуются большим структурно-функциональным разнообразием (рис. 37). При этом форма клеток часто зависит от выполняемых ими функций у многоклеточного организма. Однако общий план строения всех клеток эукариот обладает принципиальным сходством. В клетках эукариот присутствуют хорошо оформленное ядро, отграниченное от цитоплазмы оболочкой из двух мембран; хромосомы из длинных скрученных нитей ДНК; полный набор различных органоидов.
Различие клеток прокариот и эукариот особенно хорошо видно при сопоставлении их основных признаков (таблица).
|
Признаки |
Прокариоты |
Эукариоты |
|
Размер клеток |
От 0,5 до 5 мкм |
10-100 мкм |
|
Дыхание |
Аэробное или анаэробное |
Аэробное |
|
Генетический материал |
Кольцевая ДНК находится в цитоплазме и ничем не защищена |
Линейные молекулы ДНК, связанные с белками и РНК, образуют хромосомы внутри ядра |
|
Синтез РНК и белка |
И то и другое — в цитоплазме |
Синтез РНК в ядре, а белка — в цитоплазме |
|
Органоиды |
Мало |
Много |
|
Мембранные органоиды |
Клеточная (редко) и плазматическая |
Много различных мембранных органоидов |
|
Немембранные органоиды — рибосомы |
Есть в цитоплазме |
Есть в цитоплазме, в митохондриях и хлоропластах Материал с сайта http://doklad-referat.ru |
|
Внутриклеточное переваривание |
Нет |
Есть |
|
Деление клеток |
Прямое деление надвое |
Мейоз и митоз |
|
Хромосомы |
Одна кольцевая, содержит мало белка |
Много линейных, содержат белок |
У клеток эукариот и прокариот сходно функционирует клеточная мембрана. О чём свидетельствует этот факт?
Сравните клетки эукариот и прокариот.
doklad-referat.ru
Реферат - Строение клеток прокариот и эукариот
О.Д. Лопина
По строению клетки живые организмы делят на прокариот и эукариот. Клетки и тех и других окружены плазматической мембраной, снаружи от которой во многих случаях имеется клеточная стенка. Внутри клетки находится полужидкая цитоплазма. Однако клетки прокариот устроены значительно проще, чем клетки эукариот.
Рис. 1. Строение клетки прокариот
Основной генетический материал прокариот (от греч. про – до и карион – ядро) находится в цитоплазме в виде кольцевой молекулы ДНК. Эта молекула (нуклеоид) не окружена ядерной оболочкой, характерной для эукариот, и прикрепляется к плазматической мембране (рис.1). Таким образом, прокариоты не имеют оформленного ядра. Кроме нуклеоида в прокариотической клетке часто встречается небольшая кольцевая молекула ДНК, называемая плазмидой. Плазмиды могут перемещаться из одной клетки в другую и встраиваться в основную молекулу ДНК.
Некоторые прокариоты имеют выросты плазматической мембраны: мезосомы, ламеллярные тилакоиды, хроматофоры. В них сосредоточены ферменты, участвующие в фотосинтезе и в процессах дыхания. Кроме того, мезосомы ассоциированы с синтезом ДНК и секрецией белка.
Клетки прокариот имеют небольшие размеры, их диаметр составляет 0, 3–5 мкм. С наружной стороны плазматической мембраны всех прокариот (за исключением микоплазм) находится клеточная стенка. Она состоит из комплексов белков и олигосахаридов, уложенных слоями, защищает клетку и поддерживает ее форму. От плазматической мембраны она отделена небольшим межмембранным пространством.
В цитоплазме прокариот обнаруживаются только немембранные органоиды рибосомы. По структуре рибосомы прокариот и эукариот сходны, однако рибосомы прокариот имеют меньшие размеры и не прикрепляются к мембране, а располагаются прямо в цитоплазме.
Рис. 2. Строение клеток эукариот
Многие прокариоты подвижны и могут плавать или скользить с помощью жгутиков.
Размножаются прокариоты обычно путем деления надвое (бинарным). Делению предшествует очень короткая стадия удвоения, или репликации, хромосом. Так что прокариоты – гаплоидные организмы.
К прокариотам относятся бактерии и синезеленые водоросли, или цианобактерии. Прокариоты появились на Земле около 3, 5 млрд лет назад и были, вероятно, первой клеточной формой жизни, дав начало современным прокариотам и эукариотам.
Эукариоты (от греч. эу – истинный, карион – ядро) в отличие от прокариот, имеют оформленное ядро, окруженное ядерной оболочкой – двуслойной мембраной. Молекулы ДНК, обнаруживаемые в ядре, незамкнуты (линейные молекулы). Кроме ядра часть генетической информации содержится в ДНК митохондрий и хлоропластов. Эукариоты появились на Земле примерно 1, 5 млрд лет назад.
В отличие от прокариот, представленных одиночными организмами и колониальными формами, эукариоты могут быть одноклеточными (например, амеба), колониальными (вольвокс) и многоклеточными организмами. Их делят на три больших царства: Животные, Растения и Грибы.
Диаметр клеток эукариот составляет 5–80 мкм. Как и прокариотические клетки, клетки эукариот окружены плазматической мембраной, состоящей из белков и липидов. Эта мембрана работает как селективный барьер, проницаемый для одних соединений и непроницаемый для других. Снаружи от плазматической мембраны расположена прочная клеточная стенка, которая у растений состоит главным образом из волокон целлюлозы, а у грибов – из хитина. Основная функция клеточной стенки – обеспечение постоянной формы клеток. Поскольку плазматическая мембрана проницаема для воды, а клетки растений и грибов обычно соприкасаются с растворами меньшей ионной силы, чем ионная сила раствора внутри клетки, вода будет поступать внутрь клеток. За счет этого объем клеток будет увеличиваться, плазматическая мембрана начнет растягиваться и может разорваться. Клеточная стенка препятствует увеличению объема и разрушению клетки.
У животных клеточная стенка отсутствует, но наружный слой плазматической мембраны обогащен углеводными компонентами. Этот наружный слой плазматической мембраны клеток животных называют гликокаликсом. Клетки многоклеточных животных не нуждаются в прочной клеточной стенке, поскольку есть другие механизмы, обеспечивающие регуляцию клеточного объема. Так как клетки многоклеточных животных и одноклеточные организмы, живущие в море, находятся в среде, в которой суммарная концентрация ионов близка к внутриклеточной концентрации ионов, клетки не набухают и не лопаются. Одноклеточные животные, живущие в пресной воде (амеба, инфузория туфелька), имеют сократительные вакуоли, которые постоянно выводят наружу поступающую внутрь клетки воду.
Структурные компоненты эукариотической клетки
Внутри клетки под плазматической мембраной находятся цитоплазма. Основное вещество цитоплазмы (гиалоплазма) представляет собой концентрированный раствор неорганических и органических соединений, главными компонентами которого являются белки. Это коллоидная система, которая может переходить из жидкого в гелеобразное состояние и обратно. Значительная часть белков цитоплазмы является ферментами, осуществляющими различные химические реакции. В гиалоплазме располагаются органоиды, выполняющие в клетке различные функции. Органоиды могут быть мембранными (ядро, аппарат Гольджи, эндоплазматический ретикулум, лизосомы, митохондрии, хлоропласты) и немембранными (клеточный центр, рибосомы, цитоскелет).
Мембранные органоиды
Основным компонентом мембранных органоидов является мембрана. Биологические мембраны построены по общему принципу, но химический состав мембран разных органоидов различен. Все клеточные мембраны – это тонкие пленки (толщиной 7–10 нм), основу которых составляет двойной слой липидов (бислой), расположенных так, что заряженные гидрофильные части молекул соприкасаются со средой, а гидрофобные остатки жирных кислот каждого монослоя направлены внутрь мембраны и соприкасаются друг с другом (рис. 3). В бислой липидов встроены молекулы белков (интегральные белки мембраны) таким образом, что гидрофобные части молекулы белка соприкасаются с жирнокислотными остатками молекул липидов, а гидрофильные части экспонированы в окружающую среду. Кроме этого часть растворимых (немембранных белков) соединяется с мембраной в основном за счет ионных взаимодействий (периферические белки мембраны). Ко многим белкам и липидам в составе мембран присоединены также углеводные фрагменты. Таким образом, биологические мембраны – это липидные пленки, в которые встроены интегральные белки.
Рис. 3. Структура биологических мембран
Одна из основных функций мембран – создание границы между клеткой и окружающей средой и различными отсеками клетки. Липидный бислой проницаем в основном для жирорастворимых соединений и газов, гидрофильные вещества переносятся через мембраны с помощью специальных механизмов: низкомолекулярные – с помощью разнообразных переносчиков (каналов, насосов и др.), а высокомолекулярные – с помощью процессов экзо- и эндоцитоза (рис. 4).
Рис. 4. Схема переноса веществ через мембрану
При эндоцитозе определенные вещества сорбируются на поверхности мембраны (за счет взаимодействия с белками мембраны). В этом месте образуется впячивание мембраны внутрь цитоплазмы. Затем от мембраны отделяется пузырек, внутри которого содержится переносимое соединение. Таким образом, эндоцитоз – это перенос в клетку высокомолекулярных соединений внешней среды, окруженных участком мембраны. Обратный процесс, то есть экзоцитоз – это перенос веществ из клетки наружу. Он происходит путем слияния с плазматической мембраной пузырька, заполненного транспортируемыми высокомолекулярными соединениями. Мембрана пузырька сливается с плазматической мембраной, а его содержимое изливается наружу.
Каналы, насосы и другие переносчики – это молекулы интегральных белков мембраны, обычно образующие в мембране пору.
Кроме функций разделения пространства и обеспечения избирательной проницаемости мембраны способны воспринимать сигналы. Эту функцию осуществляют белки-рецепторы, связывающие сигнальные молекулы. Отдельные белки мембраны являются ферментами, осуществляющими определенные химические реакции.
Ядро – крупный органоид клетки, окруженный ядерной оболочкой и имеющий обычно шаровидную форму. Ядро в клетке одно, и хотя встречаются многоядерные клетки (клетки скелетных мышц, некоторых грибов) или не имеющие ядра (эритроциты и тромбоциты млекопитающих), но эти клетки возникают из одноядерных клеток-предшественников.
Основная функция ядра – хранение, передача и реализация генетической информации. Здесь происходит удвоение молекул ДНК, в результате чего при делении дочерние клетки получают одинаковый генетический материал. В ядре с использованием в качестве матрицы отдельных участков молекул ДНК (генов) происходит синтез молекул РНК: информационных (иРНК), транспортных (тРНК) и рибосомальных (рРНК), необходимых для синтеза белка. В ядре осуществляется сборка субъединиц рибосом из молекул рРНК и белков, которые синтезируются в цитоплазме и переносятся в ядро.
Ядро состоит из ядерной оболочки, хроматина (хромосом), ядрышка и нуклеоплазмы (кариоплазмы).
Рис. 5. Структура хроматина: 1 – нуклеосома, 2 – ДНК
Под микроскопом внутри ядра видны зоны плотного вещества – хроматина. В неделящихся клетках он равномерно заполняет объем ядра или конденсируется в отдельных местах в виде более плотных участков и хорошо окрашивается основными красителями. Хроматин представляет собой комплекс ДНК и белков (рис. 5), большей частью положительно заряженных гистонов.
| Рис. 6. Упаковка молекулы ДНК в хромосоме |
Количество молекул ДНК в ядре равно числу хромосом. Количество и форма хромосом являются уникальной характеристикой вида. В состав каждой из хромосом входит одна молекула ДНК, состоящая из двух связанных между собой нитей и имеющая вид двойной спирали толщиной 2 нм. Длина ее значительно превышает диаметр клетки: она может достигать нескольких сантиметров. Молекула ДНК заряжена отрицательно, поэтому сворачиваться (конденсироваться) она может только после связывания с положительно заряженными белками-гистонами (рис. 6).
Сначала двойная нить ДНК закручивается вокруг отдельных блоков гистонов, в каждый из которых входит 8 молекул белка, образуя структуру в виде «бусин на нитке» толщиной около 10 нм. Бусины называются нуклеосомами. В результате формирования нуклеосом длина молекулы ДНК уменьшается примерно в 7 раз. Далее нить с нуклеосомами сворачивается, формируя структуру в виде каната толщиной около 30 нм. Затем такой канат, изогнутый в виде петель, прикрепляется к белкам, образующим основу хромосомы. В результате образуется структура с толщиной около 300 нм. Дальнейшая конденсация этой структуры приводит к образованию хромосомы.
В период между делениями хромосома частично разворачивается. В результате этого отдельные участки молекулы ДНК, которые должны экспрессироваться в данной клетке, освобождаются от белков и вытягиваются, что делает возможным считывание с них информации путем синтеза молекул РНК.
Ядрышко – это тип матричной ДНК, отвечающей за синтез рРНК и собранной в отдельных участках ядра. Ядрышко – наиболее плотная структура ядра, оно не является отдельным органоидом, а представляет собой один из локусов хромосомы. В нем образуется рРНК, которая затем образует комплекс с белками, формируя субъединицы рибосом, которые уходят в цитоплазму.
Негистоновые белки ядра образуют внутри ядра структурную сеть. Она представлена слоем фибрилл, подстилающим ядерную оболочку. К ней прикрепляется внутриядерная сеть фибрилл, к которой присоединены фибриллы хроматина.
Ядерная оболочка состоит из двух мембран: внешней и внутренней, разделенных межмембранным пространством. Внешняя мембрана соприкасается с цитоплазмой, на ней могут находиться полирибосомы, а сама она может переходить в мембраны эндоплазматического ретикулума. Внутренняя мембрана связана с хроматином. Таким образом, ядерная оболочка обеспечивает фиксацию хромосомного материала в трехмерном пространстве ядра.
Оболочка ядра имеет круглые отверстия – ядерные поры (рис. 7). В области поры внешняя и внутренняя мембраны смыкаются и образуют отверстия, заполненные фибриллами и гранулами. Внутри поры располагается сложная система из белков, обеспечивающих избирательное связывание и перенос макромолекул. Количество ядерных пор зависит от интенсивности метаболизма клетки.
| Рис. 7. Поры в ядерной мембране | Рис. 8. Эндоплазматический ретикулум |
Эндоплазматический ретикулум, или эндоплазматическая сеть (ЭПР), представляет собой причудливую сеть каналов, вакуолей, уплощенных мешков, соединенных между собой и отделенных от гиалоплазмы мембраной (рис. 8).
Различают шероховатый и гладкий ЭПР. На мембранах шероховатого ЭПР находятся рибосомы (рис. 9), которые синтезируют белки, экскретируемые из клетки или встраивающиеся в плазматическую мембрану. Вновь синтезированный белок сходит с рибосомы и проходит через специальный канал внутрь полости эндоплазматического ретикулума, где он подвергается посттрансляционной модификации, например связыванию с углеводами, протеолитическому отщеплению части полипептидной цепи, образованию S–S-связей между остатками цистеина в цепи. Далее эти белки транспортируются в комплекс Гольджи, где входят либо в состав лизосом, либо секреторных гранул. В обоих случаях эти белки оказываются внутри мембранного пузырька (везикулы).
Рис. 9. Схема синтеза белка в шероховатом ЭПР: 1 – малая и
2 – большая субъединицы рибосомы; 3 – молекула рРНК;
4 – шероховатый ЭПР; 5 – вновь синтезируемый белок
Гладкий ЭПР лишен рибосом. Его основная функция – синтез липидов и метаболизм углеводов. Он хорошо развит, например, в клетках коркового вещества надпочечников, где содержатся ферменты, обеспечивающие синтез стероидных гормонов. В гладком ЭПР в клетках печени находятся ферменты, осуществляющие окисление (детоксикацию) чужеродных для организма гидрофобных соединений, например лекарств.
Рис. 10. Аппарат Гольджи: 1 – пузырьки; 2 – цистерны
Комплекс Гольджи (рис. 10) состоит из 5–10 плоских ограниченных мембраной полостей, расположенных параллельно. Концевые части этих дискообразных структур имеют расширения. Таких образований в клетке может быть несколько. В зоне комплекса Гольджи находится большое количество мембранных пузырьков. Часть из них отшнуровывается от концевых частей основной структуры в виде секреторных гранул и лизосом. Часть мелких пузырьков (везикул), переносящих синтезированные в шероховатом ЭПР белки, перемещается к комплексу Гольджи и сливается с ним. Таким образом комплекс Гольджи участвует в накоплении и дальнейшей модификации продуктов, синтезированных в шероховатом ЭПР, и их сортировке.
Рис. 11. Образование и функции лизосом: 1 – фагосома; 2 – пиноцитозный пузырек; 3 – первичная лизосома; 4 – аппарат Гольджи; 5 – вторичная лизосома
Лизосомы – это вакуоли (рис. 11), ограниченные одной мембраной, которые отпочковываются от комплекса Гольджи. Внутри лизосом достаточно кислая среда (рН 4, 9–5, 2). Там располагаются гидролитические ферменты, расщепляющие различные полимеры при кислых рН (протеазы, нуклеазы, глюкозидазы, фосфатазы, липазы). Эти первичные лизосомы сливаются с эндоцитозными вакуолями, содержащими компоненты, которые должны расщепляться. Вещества, попавшие во вторичную лизосому, расщепляются до мономеров и переносятся через мембрану лизосомы в гиалоплазму. Таким образом, лизосомы участвуют в процессах внутриклеточного переваривания.
Митохондрии окружены двумя мембранами: наружной, отделяющей митохондрию от гиалоплазмы, и внутренней, отграничивающей ее внутреннее содержимое. Между ними располагается межмембранное пространство шириной 10–20 нм. Внутренняя мембрана образует многочисленные выросты (кристы). В этой мембране располагаются ферменты, обеспечивающие окисление образовавшихся за пределами митохондрий аминокислот, сахаров, глицерина и жирных кислот (цикл Кребса) и осуществляющие перенос электронов в дыхательной цепи (схема). За счет переноса электронов по дыхательной цепи с высокого на более низкий энергетический уровень часть освобождающейся свободной энергии запасается в виде АТФ – универсальной энергетической валюты клетки. Таким образом, основная функция митохондрий – это окисление различных субстратов и синтез молекул АТФ.
Схема переноса двух электронов по дыхательной цепи
Внутри митохондрии находится кольцевая молекула ДНК, которая кодирует часть белков митохондрии. Во внутреннем пространстве митохондрий (матриксе) находятся рибосомы, похожие на рибосомы прокариот, которые и обеспечивают синтез этих белков.
Тот факт, что митохондрии имеют свою кольцевую ДНК и прокариотические рибосомы, привел к возникновению гипотезы, согласно которой митохондрия является потомком древней прокариотической клетки, когда-то попавшей внутрь эукариотической и в процессе эволюции взявшей на себя отдельные функции.
Рис. 12. Хлоропласты (А) и тилакоидные мембраны (Б)
Пластиды – органоиды растительной клетки, которые содержат пигменты. В хлоропластах содержится хлорофилл и каротиноиды, в хромопластах – каротиноиды, в лейкопластах пигментов нет. Пластиды окружены двойной мембраной. Внутри них располагается система мембран, имеющая форму плоских пузырьков, называемых тилакоидами (рис. 12). Тилакоиды уложены в стопки, напоминающие стопки тарелок. Пигменты встроены в мембраны тилакоидов. Их основная функция – поглощение света, энергия которого с помощью ферментов, встроенных в мембрану тилакоида, преобразуется в градиент ионов Н+ на мембране тилакоида. Как и митохондрии, пластиды имеют собственную кольцевую ДНК и рибосомы прокариотического типа. По-видимому, пластиды также являются прокариотическим организмом, живущим в симбиозе с клетками эукариот.
Рибосомы – это немембранные клеточные органоиды, встречающиеся как в клетках про-, так и эукариот. Рибосомы эукариот больше по размеру, чем прокариотические, их размер составляет 25х20х20 нм. Состоит рибосома из большой и малой субъединиц, прилегающих друг к другу. Между субъединицами в функционирующей рибосоме располагается нить иРНК.
Каждая субъединица рибосомы построена из рРНК, плотно упакованной и связанной с белками. Рибосомы могут располагаться в цитоплазме свободно или быть связанными с мембранами ЭПР. Свободные рибосомы могут быть единичными, но могут образовывать полисомы, когда на одной нити иРНК располагается последовательно несколько рибосом. Основная функция рибосом – синтез белка.
Цитоскелет – это опорно-двигательная система клетки, включающая белковые нитчатые (фибриллярные) образования, являющиеся каркасом клетки и выполняющие двигательную функцию. Структуры цитоскелета динамичны, они возникают и распадаются. Цитоскелет представлен тремя типами образований: промежуточными филаментами (нити диаметром 10 нм), микрофиламенты (нити диаметром 5–7 нм) и микротрубочками. Промежуточные филаменты – неветвящиеся белковые структуры в виде нитей, часто расположенные пучками. Их белковый состав различен в разных тканях: в эпителии они состоят из кератина, в фибробластах – из виментина, в мышечных клетках – из десмина. Промежуточные филаменты выполнят опорно-каркасную функцию.
| Рис. 13. Строение микроворсинки животной клетки (кишечного эпителия) |
Микрофиламенты – это фибриллярные структуры, расположенные непосредственно под плазматической мембраной в виде пучков или слоев. Они хорошо видны в ложноножках амебы, в движущихся отростках фибробластов, в микроворсинках кишечного эпителия (рис. 13). Микрофиламенты построены из сократительных белков актина и миозина и являются внутриклеточным сократительным аппаратом.
Микротрубочки входят в состав как временных, так и постоянных структур клетки. К временным относится веретено деления, элементы цитоскелета клеток между делениями, а к постоянным – реснички, жгутики и центриоли клеточного центра. Микротрубочки – это прямые полые цилиндры с диаметром около 24 нм, их стенки образованы округлыми молекулами белка тубулина. Под электронными микроскопом видно, что сечение микротрубочки образовано 13 субъединицами, соединенными в кольцо. Микротрубочки присутствуют в гиалоплазме всех эукариотических клеток. Одна из функций микротрубочек – создание каркаса внутри клеток. Кроме того, по микротрубочкам, как по рельсам, перемещаются мелкие везикулы.
Клеточный центр состоит из двух центриолей, расположенных под прямым углом друг к другу и связанных с ними микротрубочек. Эти органеллы в делящихся клетках принимают участие в формировании веретена деления. Основой центриоли являются расположенные по окружности 9 триплетов микротрубочек, образующих полый цилиндр, шириной 0, 2 мкм и длиной 0, 3–0, 5 мкм. При подготовке клеток к делению центриоли расходятся и удваиваются. Перед митозом центриоли участвуют в образовании микротрубочек веретена деления. Клетки высших растений не имеют центриолей, но у них есть аналогичный центр организации микротрубочек.
www.ronl.ru
Реферат Эукариота
скачатьРеферат на тему:
План:
- Введение
- 1 Строение эукариотической клетки
- 2 Деление на царства
- 3 Отличия эукариот от прокариот
- 4 Источники, ссылки
Введение
Диаграмма типичной клетки животного. Отмеченные органоиды (органеллы) 1. Ядрышко 2. Ядро 3. Рибосома 4. Везикула 5. Шероховатый (гранулярный) эндоплазматический ретикулум 6. Аппарат Гольджи 7. Клеточная стенка 8. Гладкий (агранулярный) эндоплазматический ретикулум 9. Митохондрия 10. Вакуоль 11. Гиалоплазма 12. Лизосома 13. Центросома (Центриоль)
Эукарио́ты, или Я́дерные (лат. Eukaryota от греч. εύ- — хорошо и κάρυον — ядро) — домен (надцарство) живых организмов, клетки которых содержат ядра. Все организмы, кроме бактерий и архей, являются ядерными (вирусы и вироиды также не являются эукариотами, но не все биологи считают их живыми организмами).
Животные, растения, грибы, а также группы организмов под общим названием протисты — все являются эукариотическими организмами. Они могут быть одноклеточными и многоклеточными, но все имеют общий план строения клеток. Считается, что все эти столь несхожие организмы имеют общее происхождение, поэтому группа ядерных рассматривается как монофилетический таксон наивысшего ранга. Согласно наиболее распространённым гипотезам, эукариоты появились 1,5–2 млрд. лет назад. Важную роль в эволюции эукариот сыграл симбиогенез — симбиоз между эукариотической клеткой, видимо, уже имевшей ядро и способной к фагоцитозу, и проглоченными этой клеткой бактериями — предшественниками митохондрий и пластидов.
1. Строение эукариотической клетки
Эукариотические клетки в среднем намного крупнее прокариотических, разница в объёме достигает тысяч раз. Клетки эукариот включают около десятка видов различных структур, известных как органоиды (или органеллы, что, правда, несколько искажает первоначальное значение этого термина), из которых многие отделены от цитоплазмы одной или несколькими мембранами. В прокариотических клетках всегда присутствуют клеточная мембрана, рибосомы (существенно отличные от эукариотических рибосом) и генетический материал — бактериальная хромосома, или генофор, однако внутренние органоиды, окруженные мембраной, встречаются редко. Ядро — это часть клетки, окружённая у эукариот двойной мембраной (двумя элементарными мембранами) и содержащая генетический материал: молекулы ДНК, «упакованные» в хромосомы. Ядро обычно одно, но бывают и многоядерные клетки.
2. Деление на царства
Существует несколько вариантов деления надцарства эукариот на царства. Первыми были выделены царства растений и животных. Затем было выделено царство грибов, которые из-за биохимических особенностей, по мнению большинства биологов, не могут быть причислены ни к одному из этих царств. Также некоторые авторы выделяют царства простейших, миксомицетов, хромистов. Некоторые системы насчитывают до 20 царств. По системе Томаса Кавалир-Смита все эукариоты подразделяются на два монофилетических таксона — Unikonta и Bikonta.
3. Отличия эукариот от прокариот
Важнейшая, основополагающая особенность эукариотических клеток связана с расположением генетического аппарата в клетке. Генетический аппарат всех эукариот находится в ядре и защищён ядерной оболочкой (по-гречески «эукариот» значит имеющий ядро). ДНК эукариот линейная (у прокариот ДНК кольцевая и находится в особой области клетки — нуклеоиде, который не отделён мембраной от остальной цитоплазмы). Она связана с белками-гистонами и другими белками хромосом, которых нет у бактерий.
В жизненном цикле эукариот обычно присутствуют две ядерные фазы (гаплофаза и диплофаза). Первая фаза характеризуется гаплоидным (одинарным) набором хромосом, далее, сливаясь, две гаплоидные клетки (или два ядра) образуют диплоидную клетку (ядро), содержащую двойной (диплоидный) набор хромосом. Иногда при следующем делении, а чаще спустя несколько делений клетка вновь становится гаплоидной. Такой жизненный цикл и в целом диплоидность для прокариот не характерны.
Третье, пожалуй, самое интересное отличие, — это наличие у эукариотических клеток особых органелл, имеющих свой генетический аппарат, размножающихся делением и окружённых мембраной. Эти органеллы — митохондрии и пластиды. По своему строению и жизнедеятельности они поразительно похожи на бактерий. Это обстоятельство натолкнуло современных учёных на мысль, что подобные организмы являются потомками бактерий, вступившими в симбиотические отношения с эукариотами. Прокариоты характеризуются малым количеством органелл, и ни одна из них не окружена двойной мембраной. В клетках прокариот нет эндоплазматического ретикулума, аппарата Гольджи, лизосом.
Ещё одно важное различие между прокариотами и эукариотами — наличие у эукариот эндоцитоза, в том числе у многих групп — фагоцитоза. Фагоцитозом (дословно «поедание клеткой») называют способность эукариотических клеток захватывать, заключая в мембранный пузырёк, и переваривать самые разные твёрдые частицы. Этот процесс обеспечивает в организме важную защитную функцию. Впервые он был открыт И. И. Мечниковым у морских звёзд. Появление фагоцитоза у эукариот скорее всего связано со средними размерами (далее о размерных различиях написано подробнее). Размеры прокариотических клеток несоизмеримо меньше, и поэтому в процессе эволюционного развития эукариот у них возникла проблема снабжения организма большим количеством пищи. Как следствие среди эукариот появляются первые настоящие, подвижные хищники.
Большинство бактерий имеет клеточную стенку, отличную от эукариотической (далеко не все эукариоты имеют её). У прокариот это прочная структура, состоящая главным образом из муреина (у архей из псевдомуреина). Строение муреина таково, что каждая клетка окружена особым сетчатым мешком, являющимся одной огромной молекулой. Среди эукариот клеточную стенку имеют многие протисты, грибы и растения. У грибов она состоит из хитина и глюканов, у низших растений — из целлюлозы и гликопротеинов, диатомовые водоросли синтезируют клеточную стенку из кремниевых кислот, у высших растений она состоит из целлюлозы, гемицеллюлозы и пектина. Видимо, для более крупных эукариотических клеток стало невозможно создавать клеточную стенку из одной молекулы высокую по прочности. Это обстоятельство могло заставить эукариот использовать иной материал для клеточной стенки. Другое объяснение состоит в том, что общий предок эукариот в связи с переходом к хищничеству утратил клеточную стенку, а затем были утрачены и гены, отвечающие за синтез муреина. При возврате части эукариот к осмотрофному питанию клеточная стенка появилась вновь, но уже на другой биохимической основе.
Разнообразен и обмен веществ у бактерий. Вообще всего выделяют четыре типа питания, и среди бактерий встречаются все. Это фотоавтотрофные, фотогетеротрофные, хемоавтотрофные, хемогетеротрофные (фототрофные используют энергию солнечного света, хемотрофные используют химическую энергию). Эукариоты же либо сами синтезируют энергию из солнечного света, либо используют готовую энергию такого происхождения. Это может быть связано с появлением среди эукариотов хищников, необходимость синтезировать энергию для которых отпала.
Ещё одно отличие — строение жгутиков. У бактерий они тонкие — всего 15–20 нм в диаметре. Это полые нити из белка флагеллина. Строение жгутиков эукариот гораздо сложнее. Они представляют собой вырост клетки, окруженный мембраной, и содержат цитоскелет (аксонему) из девяти пар периферических микротрубочек и двух микротрубочек в центре. В отличие от вращающихся прокариотическох жгутиков жгутики эукариот изгибаются или извиваются. Две группы рассматриваемых нами организмов, как уже было сказано, сильно отличаются и по своим средним размерам. Диаметр прокариотической клетки составляет обычно 0,5–10 мкм, когда тот же показатель у эукариот составляет 10–100 мкм. Объём такой клетки в 1000–10000 раз больше, чем прокариотической. У прокариот рибосомы мелкие (70S-типа). У эукариот рибосомы более крупные (80S-типа).
Видимо, различается и время возникновения этих групп. Первые прокариоты возникли в процессе эволюции около 3,5 млрд. лет назад, от них около 1,2 млрд. лет назад произошли эукариотические организмы.
4. Источники, ссылки
- Галицкий В. А. Возникновение эукариотических клеток и происхождение апоптоза // Цитология, 2005, том 47, вып. 2, с. 103—120.
- Биологический энциклопедический словарь / под редакцией М. С. Гилярова. — М., 1989.
- Мирабдуллаев И. М. Проблема происхождения эукариот // Успехи совр. биол. 1989а. Т. 107. С. 341—356.
- Марков А. В. Проблема происхождения эукариот // Палеонтологический журнал 2 (2005): 3—12.
- Подборка статей по проблеме происхождения эукариот
- Вся Биология — Эукариоты
- Б. М. Медников. Биология: формы и уровни жизни. — Просвещение, 1995.
- Д. Тейлор, Н. Грин, У. Стаут. Биология (в трёх томах)
wreferat.baza-referat.ru
Реферат eukaryota
скачатьРеферат на тему:
План:
- Введение
- 1 Строение эукариотической клетки
- 2 Деление на царства
- 3 Отличия эукариот от прокариот
- 4 Источники, ссылки
Введение
Диаграмма типичной клетки животного. Отмеченные органоиды (органеллы) 1. Ядрышко 2. Ядро 3. Рибосома 4. Везикула 5. Шероховатый (гранулярный) эндоплазматический ретикулум 6. Аппарат Гольджи 7. Клеточная стенка 8. Гладкий (агранулярный) эндоплазматический ретикулум 9. Митохондрия 10. Вакуоль 11. Гиалоплазма 12. Лизосома 13. Центросома (Центриоль)
Эукарио́ты, или Я́дерные (лат. Eukaryota от греч. εύ- — хорошо и κάρυον — ядро) — домен (надцарство) живых организмов, клетки которых содержат ядра. Все организмы, кроме бактерий и архей, являются ядерными (вирусы и вироиды также не являются эукариотами, но не все биологи считают их живыми организмами).
Животные, растения, грибы, а также группы организмов под общим названием протисты — все являются эукариотическими организмами. Они могут быть одноклеточными и многоклеточными, но все имеют общий план строения клеток. Считается, что все эти столь несхожие организмы имеют общее происхождение, поэтому группа ядерных рассматривается как монофилетический таксон наивысшего ранга. Согласно наиболее распространённым гипотезам, эукариоты появились 1,5–2 млрд. лет назад. Важную роль в эволюции эукариот сыграл симбиогенез — симбиоз между эукариотической клеткой, видимо, уже имевшей ядро и способной к фагоцитозу, и проглоченными этой клеткой бактериями — предшественниками митохондрий и пластидов.
1. Строение эукариотической клетки
Эукариотические клетки в среднем намного крупнее прокариотических, разница в объёме достигает тысяч раз. Клетки эукариот включают около десятка видов различных структур, известных как органоиды (или органеллы, что, правда, несколько искажает первоначальное значение этого термина), из которых многие отделены от цитоплазмы одной или несколькими мембранами. В прокариотических клетках всегда присутствуют клеточная мембрана, рибосомы (существенно отличные от эукариотических рибосом) и генетический материал — бактериальная хромосома, или генофор, однако внутренние органоиды, окруженные мембраной, встречаются редко. Ядро — это часть клетки, окружённая у эукариот двойной мембраной (двумя элементарными мембранами) и содержащая генетический материал: молекулы ДНК, «упакованные» в хромосомы. Ядро обычно одно, но бывают и многоядерные клетки.
2. Деление на царства
Существует несколько вариантов деления надцарства эукариот на царства. Первыми были выделены царства растений и животных. Затем было выделено царство грибов, которые из-за биохимических особенностей, по мнению большинства биологов, не могут быть причислены ни к одному из этих царств. Также некоторые авторы выделяют царства простейших, миксомицетов, хромистов. Некоторые системы насчитывают до 20 царств. По системе Томаса Кавалир-Смита все эукариоты подразделяются на два монофилетических таксона — Unikonta и Bikonta.
3. Отличия эукариот от прокариот
Важнейшая, основополагающая особенность эукариотических клеток связана с расположением генетического аппарата в клетке. Генетический аппарат всех эукариот находится в ядре и защищён ядерной оболочкой (по-гречески «эукариот» значит имеющий ядро). ДНК эукариот линейная (у прокариот ДНК кольцевая и находится в особой области клетки — нуклеоиде, который не отделён мембраной от остальной цитоплазмы). Она связана с белками-гистонами и другими белками хромосом, которых нет у бактерий.
В жизненном цикле эукариот обычно присутствуют две ядерные фазы (гаплофаза и диплофаза). Первая фаза характеризуется гаплоидным (одинарным) набором хромосом, далее, сливаясь, две гаплоидные клетки (или два ядра) образуют диплоидную клетку (ядро), содержащую двойной (диплоидный) набор хромосом. Иногда при следующем делении, а чаще спустя несколько делений клетка вновь становится гаплоидной. Такой жизненный цикл и в целом диплоидность для прокариот не характерны.
Третье, пожалуй, самое интересное отличие, — это наличие у эукариотических клеток особых органелл, имеющих свой генетический аппарат, размножающихся делением и окружённых мембраной. Эти органеллы — митохондрии и пластиды. По своему строению и жизнедеятельности они поразительно похожи на бактерий. Это обстоятельство натолкнуло современных учёных на мысль, что подобные организмы являются потомками бактерий, вступившими в симбиотические отношения с эукариотами. Прокариоты характеризуются малым количеством органелл, и ни одна из них не окружена двойной мембраной. В клетках прокариот нет эндоплазматического ретикулума, аппарата Гольджи, лизосом.
Ещё одно важное различие между прокариотами и эукариотами — наличие у эукариот эндоцитоза, в том числе у многих групп — фагоцитоза. Фагоцитозом (дословно «поедание клеткой») называют способность эукариотических клеток захватывать, заключая в мембранный пузырёк, и переваривать самые разные твёрдые частицы. Этот процесс обеспечивает в организме важную защитную функцию. Впервые он был открыт И. И. Мечниковым у морских звёзд. Появление фагоцитоза у эукариот скорее всего связано со средними размерами (далее о размерных различиях написано подробнее). Размеры прокариотических клеток несоизмеримо меньше, и поэтому в процессе эволюционного развития эукариот у них возникла проблема снабжения организма большим количеством пищи. Как следствие среди эукариот появляются первые настоящие, подвижные хищники.
Большинство бактерий имеет клеточную стенку, отличную от эукариотической (далеко не все эукариоты имеют её). У прокариот это прочная структура, состоящая главным образом из муреина (у архей из псевдомуреина). Строение муреина таково, что каждая клетка окружена особым сетчатым мешком, являющимся одной огромной молекулой. Среди эукариот клеточную стенку имеют многие протисты, грибы и растения. У грибов она состоит из хитина и глюканов, у низших растений — из целлюлозы и гликопротеинов, диатомовые водоросли синтезируют клеточную стенку из кремниевых кислот, у высших растений она состоит из целлюлозы, гемицеллюлозы и пектина. Видимо, для более крупных эукариотических клеток стало невозможно создавать клеточную стенку из одной молекулы высокую по прочности. Это обстоятельство могло заставить эукариот использовать иной материал для клеточной стенки. Другое объяснение состоит в том, что общий предок эукариот в связи с переходом к хищничеству утратил клеточную стенку, а затем были утрачены и гены, отвечающие за синтез муреина. При возврате части эукариот к осмотрофному питанию клеточная стенка появилась вновь, но уже на другой биохимической основе.
Разнообразен и обмен веществ у бактерий. Вообще всего выделяют четыре типа питания, и среди бактерий встречаются все. Это фотоавтотрофные, фотогетеротрофные, хемоавтотрофные, хемогетеротрофные (фототрофные используют энергию солнечного света, хемотрофные используют химическую энергию). Эукариоты же либо сами синтезируют энергию из солнечного света, либо используют готовую энергию такого происхождения. Это может быть связано с появлением среди эукариотов хищников, необходимость синтезировать энергию для которых отпала.
Ещё одно отличие — строение жгутиков. У бактерий они тонкие — всего 15–20 нм в диаметре. Это полые нити из белка флагеллина. Строение жгутиков эукариот гораздо сложнее. Они представляют собой вырост клетки, окруженный мембраной, и содержат цитоскелет (аксонему) из девяти пар периферических микротрубочек и двух микротрубочек в центре. В отличие от вращающихся прокариотическох жгутиков жгутики эукариот изгибаются или извиваются. Две группы рассматриваемых нами организмов, как уже было сказано, сильно отличаются и по своим средним размерам. Диаметр прокариотической клетки составляет обычно 0,5–10 мкм, когда тот же показатель у эукариот составляет 10–100 мкм. Объём такой клетки в 1000–10000 раз больше, чем прокариотической. У прокариот рибосомы мелкие (70S-типа). У эукариот рибосомы более крупные (80S-типа).
Видимо, различается и время возникновения этих групп. Первые прокариоты возникли в процессе эволюции около 3,5 млрд. лет назад, от них около 1,2 млрд. лет назад произошли эукариотические организмы.
4. Источники, ссылки
- Галицкий В. А. Возникновение эукариотических клеток и происхождение апоптоза // Цитология, 2005, том 47, вып. 2, с. 103—120.
- Биологический энциклопедический словарь / под редакцией М. С. Гилярова. — М., 1989.
- Мирабдуллаев И. М. Проблема происхождения эукариот // Успехи совр. биол. 1989а. Т. 107. С. 341—356.
- Марков А. В. Проблема происхождения эукариот // Палеонтологический журнал 2 (2005): 3—12.
- Подборка статей по проблеме происхождения эукариот
- Вся Биология — Эукариоты
- Б. М. Медников. Биология: формы и уровни жизни. — Просвещение, 1995.
- Д. Тейлор, Н. Грин, У. Стаут. Биология (в трёх томах)
wreferat.baza-referat.ru
Реферат Эукариот
скачатьРеферат на тему:
План:
- Введение
- 1 Строение эукариотической клетки
- 2 Деление на царства
- 3 Отличия эукариот от прокариот
- 4 Источники, ссылки
Введение
Диаграмма типичной клетки животного. Отмеченные органоиды (органеллы) 1. Ядрышко 2. Ядро 3. Рибосома 4. Везикула 5. Шероховатый (гранулярный) эндоплазматический ретикулум 6. Аппарат Гольджи 7. Клеточная стенка 8. Гладкий (агранулярный) эндоплазматический ретикулум 9. Митохондрия 10. Вакуоль 11. Гиалоплазма 12. Лизосома 13. Центросома (Центриоль)
Эукарио́ты, или Я́дерные (лат. Eukaryota от греч. εύ- — хорошо и κάρυον — ядро) — домен (надцарство) живых организмов, клетки которых содержат ядра. Все организмы, кроме бактерий и архей, являются ядерными (вирусы и вироиды также не являются эукариотами, но не все биологи считают их живыми организмами).
Животные, растения, грибы, а также группы организмов под общим названием протисты — все являются эукариотическими организмами. Они могут быть одноклеточными и многоклеточными, но все имеют общий план строения клеток. Считается, что все эти столь несхожие организмы имеют общее происхождение, поэтому группа ядерных рассматривается как монофилетический таксон наивысшего ранга. Согласно наиболее распространённым гипотезам, эукариоты появились 1,5–2 млрд. лет назад. Важную роль в эволюции эукариот сыграл симбиогенез — симбиоз между эукариотической клеткой, видимо, уже имевшей ядро и способной к фагоцитозу, и проглоченными этой клеткой бактериями — предшественниками митохондрий и пластидов.
1. Строение эукариотической клетки
Эукариотические клетки в среднем намного крупнее прокариотических, разница в объёме достигает тысяч раз. Клетки эукариот включают около десятка видов различных структур, известных как органоиды (или органеллы, что, правда, несколько искажает первоначальное значение этого термина), из которых многие отделены от цитоплазмы одной или несколькими мембранами. В прокариотических клетках всегда присутствуют клеточная мембрана, рибосомы (существенно отличные от эукариотических рибосом) и генетический материал — бактериальная хромосома, или генофор, однако внутренние органоиды, окруженные мембраной, встречаются редко. Ядро — это часть клетки, окружённая у эукариот двойной мембраной (двумя элементарными мембранами) и содержащая генетический материал: молекулы ДНК, «упакованные» в хромосомы. Ядро обычно одно, но бывают и многоядерные клетки.
2. Деление на царства
Существует несколько вариантов деления надцарства эукариот на царства. Первыми были выделены царства растений и животных. Затем было выделено царство грибов, которые из-за биохимических особенностей, по мнению большинства биологов, не могут быть причислены ни к одному из этих царств. Также некоторые авторы выделяют царства простейших, миксомицетов, хромистов. Некоторые системы насчитывают до 20 царств. По системе Томаса Кавалир-Смита все эукариоты подразделяются на два монофилетических таксона — Unikonta и Bikonta.
3. Отличия эукариот от прокариот
Важнейшая, основополагающая особенность эукариотических клеток связана с расположением генетического аппарата в клетке. Генетический аппарат всех эукариот находится в ядре и защищён ядерной оболочкой (по-гречески «эукариот» значит имеющий ядро). ДНК эукариот линейная (у прокариот ДНК кольцевая и находится в особой области клетки — нуклеоиде, который не отделён мембраной от остальной цитоплазмы). Она связана с белками-гистонами и другими белками хромосом, которых нет у бактерий.
В жизненном цикле эукариот обычно присутствуют две ядерные фазы (гаплофаза и диплофаза). Первая фаза характеризуется гаплоидным (одинарным) набором хромосом, далее, сливаясь, две гаплоидные клетки (или два ядра) образуют диплоидную клетку (ядро), содержащую двойной (диплоидный) набор хромосом. Иногда при следующем делении, а чаще спустя несколько делений клетка вновь становится гаплоидной. Такой жизненный цикл и в целом диплоидность для прокариот не характерны.
Третье, пожалуй, самое интересное отличие, — это наличие у эукариотических клеток особых органелл, имеющих свой генетический аппарат, размножающихся делением и окружённых мембраной. Эти органеллы — митохондрии и пластиды. По своему строению и жизнедеятельности они поразительно похожи на бактерий. Это обстоятельство натолкнуло современных учёных на мысль, что подобные организмы являются потомками бактерий, вступившими в симбиотические отношения с эукариотами. Прокариоты характеризуются малым количеством органелл, и ни одна из них не окружена двойной мембраной. В клетках прокариот нет эндоплазматического ретикулума, аппарата Гольджи, лизосом.
Ещё одно важное различие между прокариотами и эукариотами — наличие у эукариот эндоцитоза, в том числе у многих групп — фагоцитоза. Фагоцитозом (дословно «поедание клеткой») называют способность эукариотических клеток захватывать, заключая в мембранный пузырёк, и переваривать самые разные твёрдые частицы. Этот процесс обеспечивает в организме важную защитную функцию. Впервые он был открыт И. И. Мечниковым у морских звёзд. Появление фагоцитоза у эукариот скорее всего связано со средними размерами (далее о размерных различиях написано подробнее). Размеры прокариотических клеток несоизмеримо меньше, и поэтому в процессе эволюционного развития эукариот у них возникла проблема снабжения организма большим количеством пищи. Как следствие среди эукариот появляются первые настоящие, подвижные хищники.
Большинство бактерий имеет клеточную стенку, отличную от эукариотической (далеко не все эукариоты имеют её). У прокариот это прочная структура, состоящая главным образом из муреина (у архей из псевдомуреина). Строение муреина таково, что каждая клетка окружена особым сетчатым мешком, являющимся одной огромной молекулой. Среди эукариот клеточную стенку имеют многие протисты, грибы и растения. У грибов она состоит из хитина и глюканов, у низших растений — из целлюлозы и гликопротеинов, диатомовые водоросли синтезируют клеточную стенку из кремниевых кислот, у высших растений она состоит из целлюлозы, гемицеллюлозы и пектина. Видимо, для более крупных эукариотических клеток стало невозможно создавать клеточную стенку из одной молекулы высокую по прочности. Это обстоятельство могло заставить эукариот использовать иной материал для клеточной стенки. Другое объяснение состоит в том, что общий предок эукариот в связи с переходом к хищничеству утратил клеточную стенку, а затем были утрачены и гены, отвечающие за синтез муреина. При возврате части эукариот к осмотрофному питанию клеточная стенка появилась вновь, но уже на другой биохимической основе.
Разнообразен и обмен веществ у бактерий. Вообще всего выделяют четыре типа питания, и среди бактерий встречаются все. Это фотоавтотрофные, фотогетеротрофные, хемоавтотрофные, хемогетеротрофные (фототрофные используют энергию солнечного света, хемотрофные используют химическую энергию). Эукариоты же либо сами синтезируют энергию из солнечного света, либо используют готовую энергию такого происхождения. Это может быть связано с появлением среди эукариотов хищников, необходимость синтезировать энергию для которых отпала.
Ещё одно отличие — строение жгутиков. У бактерий они тонкие — всего 15–20 нм в диаметре. Это полые нити из белка флагеллина. Строение жгутиков эукариот гораздо сложнее. Они представляют собой вырост клетки, окруженный мембраной, и содержат цитоскелет (аксонему) из девяти пар периферических микротрубочек и двух микротрубочек в центре. В отличие от вращающихся прокариотическох жгутиков жгутики эукариот изгибаются или извиваются. Две группы рассматриваемых нами организмов, как уже было сказано, сильно отличаются и по своим средним размерам. Диаметр прокариотической клетки составляет обычно 0,5–10 мкм, когда тот же показатель у эукариот составляет 10–100 мкм. Объём такой клетки в 1000–10000 раз больше, чем прокариотической. У прокариот рибосомы мелкие (70S-типа). У эукариот рибосомы более крупные (80S-типа).
Видимо, различается и время возникновения этих групп. Первые прокариоты возникли в процессе эволюции около 3,5 млрд. лет назад, от них около 1,2 млрд. лет назад произошли эукариотические организмы.
4. Источники, ссылки
- Галицкий В. А. Возникновение эукариотических клеток и происхождение апоптоза // Цитология, 2005, том 47, вып. 2, с. 103—120.
- Биологический энциклопедический словарь / под редакцией М. С. Гилярова. — М., 1989.
- Мирабдуллаев И. М. Проблема происхождения эукариот // Успехи совр. биол. 1989а. Т. 107. С. 341—356.
- Марков А. В. Проблема происхождения эукариот // Палеонтологический журнал 2 (2005): 3—12.
- Подборка статей по проблеме происхождения эукариот
- Вся Биология — Эукариоты
- Б. М. Медников. Биология: формы и уровни жизни. — Просвещение, 1995.
- Д. Тейлор, Н. Грин, У. Стаут. Биология (в трёх томах)
wreferat.baza-referat.ru
: :
:
“
”
:
10- , N55
: ..
. 1999
1 . :
1.1 ;
;
;
;
;
2. ;
3. :
;
;
;
;
:
;
;
;
:
6.1 ;
: ;
2 :
;
;
.
- . , . - 1 . , , 800 1000 . . , , - . . : , , , , . . .
, 3 : , . - - 4 , 500 . , . . . , - , .
, . , - - ,.. , - , , . , , .. . , - , , - . , , , , - . - : , , , , , . , , - : (Monera, ) (Protoctista, Animalia, Fungi , Plantae). - , , . , , . - - . , Protoctista , - , , , , . - , , , .
- , , . ( ) , . 0,02 , , . , . . , ( ) . ( - , ) , . , , , . - . - - . . - . - . , . , , - . . . , . , , - . . , . , . , , . . , , , .
- , . .
. .. , - , , . . . , , - . , : , - , , .
, . . , , , . - , . : , . - - . - . - - . - , . , - . , , . , , . , . , - . - .
- : 2h3X+CO2=Ch3O+2X+h3O . , 2 , . , . . - . , 2 - . , - , . 2 , . , , , .
2 - . , . : , - , , -- , . - , , , .
.
, . , , - . - , , , . - . - , - : - , , , - - , - , . , - . , , - - --. , - , . , ? , , Paracoccus denitrificans. , - . - , , . - . , , , . . , . , , - : , - . . - , . - . , . - . - , . , , . - - , . - . , . . 400 . , - .
.
.
- , - - .
, , - , - , - - . . . . , - . , . , . , . , , - , . - . , - , , . . , , , , .
, - , . , - , - . , , , - .
, ()
()
:
.
:
( ) ( - )
.
, .
.
, .
.
Š: , ; .
:
: .
( ) ( , , ) ( ).
( - ), - , - - ( ).
.
, . .
.
, , .
: . - , “ “ . - , .
. - . , - . , - , - , , .
, , - ( ancestral phytomonads ) . , Cyanidium Cyanophora, . , . , , , . , , , , , . , , ( , , - ) , , . - , , , , - . , , . , , , - . , , 2 . , , . - 2 , . , . - . , - . , , , . : , - , , . : , , . , - . , , - : 0,024 . , - , , , . , . - , , , - . , , , - . , ,- . , - , , , . . , , , , - , . .
.
- - - - .
. 80- , . , , , Paracoccus .
“ “ .
. . , , , . . , , , . . , , , , , , - , .
. . - , , , . , , , - . - . , , - . , - , .
:
-
1.5 2 , . - . - ( ) .
. , , , , . N 5 1998
. . . ., . 1983
. . “ “ 1992 .
.. .. .; , 1981 , . 165 181 .
. ; . . . 1986 .1
stud-baza.ru



|
|
..:::Счетчики:::.. |
|
|
|
|
|
|
|
|