
Спинной мозг — это самый древний отдел ЦНС. Он расположен в позвоночном канале и имеет сегментарное строение. К моменту рождения из всех отделов ЦНС спинной мозг наиболее развит. К этому времени его масса составляет 3—4 г, к 6 месяцам она удваивается. У человека к 20 годам масса спинного мозга в 8 раз больше, чем у новорожденного, и равна 30 г. На ранних стадиях онтогенеза плода спинной мозг заполняет всю полость позвоночного канала. В дальнейшем позвоночник растет быстрее, чем спинной мозг. Вследствие этого он не заполняет весь канал. К моменту рождения спинной мозг достигает 3-го поясничного позвонка, а у новорожденного находится на уровне 2—3-го поясничных позвонков. К концу первого года жизни его положение такое же, как у взрослых, на уровне 1—2-го поясничных позвонков. В связи с несоответствием размеров спинного мозга и позвоночного столба, корешки, прежде чем выйти из позвоночного канала, проходят вдоль спинного мозга в нисходящем направлении. В самом нижнем отделе они образуют «конский хвост», который состоит из пояснично-крестцовых корешковых волокон и конечной нити спинного мозга.
Клеточные структуры спинного мозга характеризуются тем, что у 5—6 месячного плода имеется большое количество еще неразвитых нервных клеток, которые различны по форме и расположению. К моменту рождения все нервные и глиальные клетки хорошо развиты и не отличаются от клеток детей дошкольного возраста. У старших детей они становятся крупнее.
Рефлекторная функция спинного мозга начинается еще в эмбриональном периоде, что связано с морфологическим развитием нервной системы. Показано, что раньше всех созревают спинномозговые рефлексы: сначала появляется стадия обобщенных (генерализованных) рефлексов, которые затем переходят в специализированные акты. Все стадии рефлекторной деятельности спинного мозга обнаруживаются уже у плода и после рождения. Некоторые рефлексы, такие как хватательный, рефлекс Бабинского (подошвенный) у новорожденных сохраняются до 6-месячного возраста, а затем исчезают. Указанные рефлексы свидетельствуют о готовности ЦНС новорожденного к выполнению рефлекторных двигательных актов (рефлекс шагания, плавания, почесывания и др.). Такой контроль функции нейронного аппарата спинного мозга со стороны вышележащих отделов ЦНС создает условия для координированной и согласованной деятельности всего организма.
Спинной мозг разделяется на шейный, грудной, поясничный и крестцовый отделы, в каждый из которых входит разное количество сегментов. От сегмента отходят две пары корешков: задние и передние (рис. 1).
Задние корешки образованы аксонами первичных афферентных нейронов, тела которых лежат в спинномозговых чувствительных ганглиях; передние корешки состоят из отростков двигательных нейронов, они направляются к соответствующим эффекторам (закон Белла—Мажанди). Каждый корешок представляет собой множество нервных волокон. К примеру, дорсальный корешок кошки включает 12 тыс., а вентральный — 6 тыс. нервных волокон.
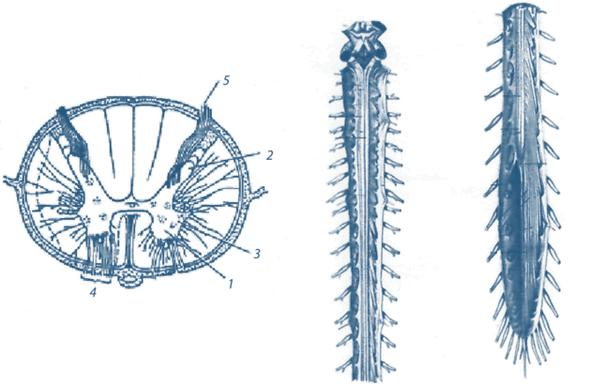
Рис. 1. Спинной мозг (поперечный и продольный срез)
(Р.И. Айзман и соавт., 2010):
1 — передний рог, 2 — задний рог серого вещества, 3 — боковой рог серого вещества,
4 — передний корешок спинного мозга, 5 — задний корешок
На поперечном разрезе спинного мозга видно, что в центре располагается серое вещество, состоящее из тел нейронов и напоминающее форму бабочки, а по периферии лежит белое вещество, представляющее собой систему отростков нейронов: восходящих (нервные волокна направляются к различным отделам головного мозга) и нисходящих (нервные волокна направляются в те или иные отделы спинного мозга). В грудном и поясничном отделах имеются боковые рога спинного мозга. В них локализованы ядра преганглионарных нейронов симпатических нервов. Их аксоны через передние рога выходят на периферию, составляя симпатическую нервную систему.
Появление и усложнение спинного мозга связано с развитием локомоции (передвижения). Локомоция, обеспечивая перемещения человека или животного в окружающей среде, создает возможность их существования.
Спинной мозг является центром многих рефлексов. Их можно разделить на три группы: защитные, вегетативные и тонические.
1. Защитно-болевые рефлексы характеризуются тем, что действие раздражителей, как правило, на кожную поверхность, вызывает защитную реакцию, которая приводит к удалению раздражителя с поверхности тела или удалению тела или его частей от раздражителя. Защитные реакции выражаются в отдергивании конечности или убегании от раздражителя (сгибательные и разгибательные рефлексы). Эти рефлексы осуществляются посегментно, но при более сложных рефлексах, таких как чесание в труднодоступных местах возникают сложные многосегментные рефлексы.
2. Вегетативные рефлексы обеспечиваются нервными клетками, расположенными в боковых рогах спинного мозга, которые являются центрами симпатической нервной системы. Здесь осуществляются сосудодвигательные, мочеиспускательный рефлексы, рефлексы дефекации, потоотделения и др.
3. Очень важное значение имеют тонические рефлексы. Они обеспечивают образование и сохранение тонуса скелетной мускулатуры. Тонус — это постоянное, невидимое сокращение (напряжение) мышц без явления утомления. Тонус обеспечивает позу и положение тела в пространстве. Поза — это фиксированное положение тела (головы и других частей тела) человека или животных в пространстве в условиях земного притяжения. Поза может быть исходным состоянием для движения, выражения эмоций (угроза, радость и т.д.). Тонус создается рефлекторным механизмом, рефлекторная дуга которого начинается с проприорецепторов, расположенных в самих мышцах (мышечные веретена). Адекватным раздражителем для них является растяжение (удлинение) мышцы. При удлинении мышцы импульсы, возникающие в рецепторах, направляются в спинной мозг, а затем, после переработки, по эфферентным нервам к этой же мышце, вызывая сокращение некоторого количества ее волокон. Это и создает тонус мышц.
Кроме того, спинной мозг выполняет проводниковую функцию, которая осуществляется восходящими и нисходящими волокнами белого вещества спинного мозга (табл. 1). В составе проводящих путей проходят как афферентные, так и эфферентные волокна. Поскольку часть этих волокон проводит интероцептивные импульсы от внутренних органов, это позволяет использовать их с целью обезболивания при внутриполостных операциях путем введения анестезирующего вещества в спинномозговой канал (спинномозговая анестезия).
Таблица 1. Проводниковые пути спинного мозга и их физиологическое значение.
| Восходящие (чувствительные) пути | Физиологическое значение |
|
|
|
Спинальный шок — это моментальное снижение рефлекторной деятельности, вызванное перерезкой или повреждением спинного мозга человека или позвоночных животных. Длительность спинального шока зависит от вида животного: у лягушек полное восстановление рефлекторной деятельности наступает через 5—10 минут, у собак и кошек — через несколько часов, а у человека спинальный шок длится недели и даже месяцы. Полученные данные позволяют считать, что продолжительность спинального шока зависит от уровня энцефализации, т.е. чем выше из спинного мозга в вышележащие отделы ЦНС перемешаются регуляторные центры, тем дольше сохраняется спинальный шок.
Имеются два предположения о причине его развития.
1. Шок развивается вследствие выпадения возбуждающего влияния со стороны вышележащих отделов головного мозга.
2. Шок возникает потому, что повреждение спинного мозга устраняет подавляющее влияние высших моторных центров на спинномозговое торможение.
referatwork.ru
Введение
Глава 1. Морфофункциональная организация
Глава 2. Рефлекторная деятельность
Заключение
Список использованной литературы
Введение
Спинной мозг – вытянутый в длину цилиндрической формы орган, заключен в позвоночном канале. Верхняя граница его у человека соответствует перекрестку пирамид, где он сливается с продолговатым мозгом. На уровне двух первых поясничных позвонков спинной мозг заканчивается мозговым конусом, который переходит в длинную терминальную нить. Конец ее фиксируется к надкостнице второго копчикового позвонка,
Спинной мозг обладает метамерией и представляет собою многосегментарное образование с билатеральной симметрией. Две его половины разделены передней срединной щелью и задней срединной бороздой. Условная линия, соединяющая их, проходит через центральный канал. У человека симметричность в строении спинного мозга в значительной степени нарушена из-за сложности меж- и надсегментарных связей.
Под сегментом понимают участок спинного мозга, соответствующий половине выше- и нижележащих межкорешковых промежутков, заключенных между входом задних и выходом передних корешков.
В шейном и пояснично-крестцовом отделах имеются соименные утолщения, которые образовались с появлением конечностей, их оптимальным мышечным вооружением. Масса и длина спинного мозга зависит от возраста и пола.
С функциональной стороны спинной мозг представлен сегментарными (рабочими) центрами, связанными прямо с исполнительными органами. Хотя это положение в связи с открытием интегрирующих механизмов его ретикулярной формации подверглось пересмотру, тем не менее, представление о спинном мозге как органе сегментарных функций остается в силе.
Спинной мозг, как и головной, развивается из нервной трубки, в которой зачатки чувствительных и двигательных зон серого вещества имеют дорсо-вентральную ориентацию и в процессе гистогенеза дают начало нейронам, группирующимся в 10 слоях или пластинах.
Для серого вещества спинного мозга характерны мультиполярные нейроны, среди которых выделяют три основных типа.
Филогенетически более древним типом считается ретикулярный или изодендритический нейрон, с немногочисленными длинными и прямыми слабоветвящимися дендритами. Эти нейроны господствуют в промежуточной зоне, хотя в небольшом количестве имеются в передних и задних рогах.
Специфические – идиодендритические нейроны снабжены большим числом часто ветвящихся дендритов. Они тесно переплетаются и образуют фигуру, имеющую вид куста или клубка. Эти нейроны характерны для передних рогов, особенно двигательных ядер, задних рогов, в частности студневидного вещества и ядра Кларка.
Третий переходный или аллодендритический тип по степени развития дендритической кроны занимает промежуточное положение. Эти нейроны имеются в дорсальной части передних и вентральной части задних рогов; они типичны для собственного ядра заднего рога.
Релейные нейроны, на которые переключается сенсорная информация с волокон задних корешков, находятся в студневидном веществе заднего рога, его собственном ядре, ядре Кларка и ядрах задних канатиков, которые лежат на границе Спинного и продолговатого мозга и рассматриваются как продолжение задних рогов.
Спинной мозг – наиболее древний отдел ЦНС позвоночных, появившийся уже у наиболее примитивных представителей хордовых – ланцетника – еще до сформирования головного мозга. Поэтому он более, чем другие отделы ЦНС, сохранил черты древней организации, выражающейся в метамерном строении.
Спинной мозг является самым каудальным отделом ЦНС. Он помещается в позвоночном канале и имеет неодинаковую длину у разных представителей позвоночных. У большинства рыб, хвостатых амфибий, рептилий и птиц он заполняет весь позвоночный канал, в то время как у бесхвостых амфибий и млекопитающих – только его часть. Поэтому в каудальном отделе позвоночного канала собираются корешки каудальных сегментов спинного мозга, образуя так называемый конский хвост.
Глава 1. Морфофункциональная организация
Спинной мозг характеризуется выраженным сегментарным строением, отражающим сегментарное строение тела позвоночных. От каждого спинномозгового сегмента отходят две пары вентральных и дорсальных корешков. Дорсальные корешки формируют афферентные входы спинного мозга. Они образованы центральными отростками волокон первичных афферентных нейронов, тела которых вынесены на периферию и находятся в спинномозговых ганглиях. Вентральные корешки образуют эфферентные выходы спинного мозга. В них проходят аксоны a и g-мотонейронов, а также преганглионарных нейронов вегетативной нервной системы. Такое распределение афферентных и эфферентных волокон было установлено еще в начале прошлого века и получило название закона Белла – Мажанди. После перерезки передних корешков на одной стороне наблюдается полное выключение двигательных реакций; но чувствительность этой стороны тела сохраняется. Перерезка задних корешков выключает чувствительность, но не приводит к утрате двигательных реакций мускулатуры.
Нейроны спинномозговых ганглиев относятся к простым униполярным, или псевдоуниполярным, нейронам. Название «псевдоуниполярный» объясняется тем, что в эмбриональном периоде первичные афферентные нейроны происходят от биполярных клеток, отростки которых затем сливаются. Нейроны спинномозговых ганглиев можно подразделить на клетки малых и больших размеров. Тело крупных нейронов имеет диаметр порядка 60–120 мкм, в то время как у мелких нейронов он колеблется от 14 до 30 мкм.
Крупные нейроны дают начало толстым миелинизированным волокнам. От мелких начинаются как тонкие миелинизированные, так и немиелинизированные волокна. После бифуркации оба отростка направляются в противоположные направления: центральный входит в дорсальный корешок и в его составе – в спинной мозг, периферический – в различные соматические и висцеральные нервы, подходящие к рецепторным образованиям кожи, мышц и внутренних органов.
Иногда центральные отростки первичных афферентных нейронов заходят в вентральной корешок. Это происходит при трифуркации аксона первичного афферентного нейрона, в результате которой его отростки проецируются в спинной мозг и через дорсальный и вентральный корешки.
Из всей популяции клеток дорсальных ганглиев примерно 60–70% относится к мелким нейронам. Это соответствует тому, что число немиелинизированных волокон в дорсальном корешке превышает число миелинизированных волокон.
Тела нейронов спинномозговых ганглиев не имеют дендритных отростков и не получают синоптических входов. Их возбуждение происходит в результате прихода потенциала действия по периферическому отростку, контактирующему с рецепторами.
Клетки спинномозговых ганглиев содержат высокие концентрации глутаминовой кислоты – одного из предполагаемых медиаторов. Их поверхностная мембрана содержит рецепторы, специфически чувствительные к g-аминомасляной кислоте, что совпадает с высокой чувствительностью к g-аминомасляной кислоте центральных окончаний первичных афферентных волокон. Малые нейроны ганглиев содержат вещество Р или соматостатин. Оба этих полипептида также являются вероятными медиаторами, высвобождаемыми окончаниями первичных афферентных волокон.
Каждая пара корешков соответствует одному из позвонков и покидает позвоночный канал через отверстие между ними. Поэтому сегменты спинного мозга принято обозначать по тому позвонку, возле которого из спинного мозга выходят соответствующие корешки. Спинной мозг принято также разделять на несколько отделов: шейный, грудной, поясничный и крестцовый, каждый из которых содержит по нескольку сегментов. В связи с развитием конечностей нейронный аппарат тех сегментов спинного мозга, которые их иннервируют, получил наибольшее развитие. Это нашло свое отражение в образовании шейного и поясничного утолщений. В области утолщений спинного мозга корешки содержат наибольшее количество волокон и имеют наибольшую толщину.
На поперечном срезе спинного мозга ясно выделяется центрально расположенное серое вещество, образованное скоплением нервных клеток, и окаймляющее его белое вещество, образованное нервными волокнами. В сером веществе различают вентральные и дорсальные рога, между которыми лежит промежуточная зона. Кроме того, в грудных сегментах различают также боковое выпячивание серого вещества – боковые рога.
Все нейронные элементы спинного мозга могут быть подразделены на 4 основные группы: эфферентные нейроны, вставочные нейроны, нейроны восходящих трактов и интраспинальные волокна чувствительных афферентных нейронов. Моторные нейроны сосредоточены в передних рогах, где они образуют специфические ядра, все клетки которых посылают свои аксоны к определенной мышце. Каждое двигательное ядро обычно тянется на несколько сегментов. Поэтому и аксоны мотонейронов, иннервирующих одну и ту же мышцу, покидают спинной мозг в составе нескольких вентральных корешков.
Кроме моторных ядер, расположенных в вентральных рогах, выделяются большие скопления нервных клеток в промежуточной зоне серого вещества. Это основное ядро вставочных нейронов спинного мозга. Аксоны вставочных нейронов распространяются как внутри сегмента, так и в ближайшие соседние сегменты.
Характерное скопление нервных клеток занимает также дорсальную часть дорсального рога. Эти клетки образуют густые переплетения, а указанная зона получила название желатинозной субстанции Роланда.
Наиболее точное и систематизированное представление о топографии нервных клеток серого вещества спинного мозга дает разделение его на последовательные слои, или пластины, в каждой из которых группируются главным образом однотипные нейроны.
Хотя послойная типография серого вещества была первоначально выявлена в спинном мозге кошки, она оказалась достаточно универсальной и вполне применима к спинному мозгу, как других позвоночных, так и человека.
Согласно этим данным, все серое вещество можно разделить на 10 пластин. Самая первая дорсальная пластина содержит главным образом, так называемые краевые нейроны. Их аксоны проецируются рострально, давая начало спиноталамическому тракту. На краевых нейронах оканчиваются волокна тракта Лиссауэра, который образован смесью первичных афферентных волокон и аксонами проприоспинальных нейронов.
Вторая и третья пластины образуют желатинозную субстанцию. Здесь локализуются два основных типа нейронов: более мелкие и относительно крупные нейроны. Хотя тела нейронов второй пластины имеют небольшой диаметр, их дендритные разветвления весьма многочисленны. Аксоны нейронов второй пластины проецируются на тракт Лиссауэра и собственный дорсолатеральный пучок спинного мозга, но многие остаются в пределах желатинозной субстанции. На клетках второй и третьей пластин оканчиваются волокна первичных афферентных нейронов, преимущественно кожной и болевой чувствительности.
Четвертая пластина занимает примерно центр дорсального рога. Дендрита нейронов IV слоя проникают в желатинозную субстанцию, а их аксоны проецируются в таламус и боковое цервикальное ядро. Синаптические входы они получают от нейронов желатинозной субстанции, а их аксоны проецируются в таламус и боковое цервикальное ядро. Синаптические входы они получают от нейронов желатинозной субстанции и первичных афферентных нейронов.
В целом нервные клетки первой-четвертой пластин захватывают всю вершину дорсального рога и образуют первичную сенсорную область спинного мозга. Сюда проецируются волокна большей частя дорсально-корешковых афферентов от экстерорецепторов, включая кожную и болевую чувствительность. В этой же зоне локализованы нервные клетки, дающие начало нескольким восходящим трактам.
В пятой и шестой пластинах локализуются многочисленные типы вставочных нейронов, получающие синаптические входы от волокон заднего корешка и нисходящих путей, в особенности кортико-спинального и руброспинального тракта.
В седьмой и восьмой пластинах локализуются проприоспинальные вставочные нейроны, дающие начало длинным аксонам, достигающим нейронов отдаленных сегментов. Здесь заканчиваются афферентные волокна от проприорецепторов, волокна вестибулоспинального и ретикулоспинального трактов, аксоны проприоспинальных нейронов.
В девятой пластине располагаются тела a- и g-мотонейронов. Этой области достигают также пресинаптические окончания первичных афферентных волокон от мышечных рецепторов растяжения, окончания волокон нисходящих трактов, кортико-спинальных волокон, терминали аксонов возбуждающих и тормозящих вставочных нейронов.
Десятая пластина окружает спинномозговой канал и содержит наряду с нейронами значительное количество глиальных клеток и комиссуральных волокон.
Клетки нейроглии спинного мозга на значительном протяжении покрывают поверхность нейронов, причем отростки глиальной клетки направлены, с одной стороны, к телам нейронов, а с другой, часто контактируют с кровеносными капиллярами, являясь посредниками между нервными элементами и источниками их питания.
Спинной мозг по восходящим путям передает сигналы в надсегментарные уровни головного мозга, а по нисходящим получает оттуда команды к действию. Восходящие пути передают импульсы от проприоцепторов по волокнам спинобульбарных пучков Голля и Бурдаха и спинномозжечковых путей Говерса и Флексиго, от болевых и температурных рецепторов по латеральному спиноталамическому тракту, от тактильных рецепторов по вентральному спиноталамическому пути и частично по пучкам Голля и Бурдаха.
Нисходящие пути проходят в составе кортикоспинальных, или пирамидных, трактов и экстракортикоспинальных, или экстрапирамидных.
Глава 2. Рефлекторная деятельность
В спинном мозге замыкается огромное количество рефлекторных дуг, с помощью которых регулируются как соматические, так и вегетативные функции организма. К числу наиболее простых рефлекторных реакций относятся сухожильные рефлексы и рефлексы растяжения, вызываемые раздражением рецепторов растяжения той же мышцы, которая развивает рефлекторное сокращение. Центральные окончания афферентных волокон от рецепторов растяжения образуют синапсы непосредственно на мотонейронах без дополнительного переключения на вставочных нейронах. Таким образом, дуга этих рефлексов может иметь моносинаптический характер. Указанное обстоятельство, а также высокая скорость проведения по афферентным волокнам, идущим от мышечных рецепторов и по аксонам a-мотонейронов, обеспечивают короткое время рефлекса (что особенно демонстративно в случае сухожильных рефлексов).
Сухожильные рефлексы легко вызываются с помощью короткого удара по сухожилию и имеют важное диагностическое значение в неврологической практике. Рефлекторная реакция проявляется в виде резкого сокращения мышцы. Особенно выражены сухожильные рефлексы в мышцах разгибателей ноги, таких, как четырехглавая мышца бедра (коленный рефлекс) или трехглавая мышца голени (ахиллов рефлекс). Однако сухожильные рефлексы вызываются и в мышцах-сгибателях. На руке они четко проявляются на двуглавой и трехглавой мышцах, на лице – на мышцах нижней челюсти.
Быстрота мышечного сокращения и отсутствие последействия обусловлены способом вызывания сухожильного рефлекса. Адекватным раздражителем для соответствующих рецепторов является растяжение мышцы. Постукивание по сухожилию растягивает мышцу только на очень краткий срок. Зато при этом чувствительные к растяжению рецепторы активируются с высокой степенью синхронности.
Поскольку афферентные волокна, идущие в спинной мозг от рецепторов растяжения, представляют собой довольно гомогенную группу по диаметру и скоростям проведения, афферентные импульсы поступают к мотонейронам в виде синхронной волны. В результате мотонейроны отвечают с незначительной временной дисперсией, посылая в двигательный нерв синхронный разряд, вызывающий короткое мышечное подергивание, сходное с ответом мышцы на одиночное электрическое раздражение двигательного нерва.
Совсем иначе характеризуется рефлекс растяжения, возникающий при адекватном раздражении тех же самых мышечных рецепторов. Естественные растяжения обычно прикладываются к мышцам под действием силы тяжести. Так, при стоянии четырехглавая мышца бедра подвергается растяжению из-за тенденции колена сгибаться под влиянием гравитационных сил. Возникающая в ответ на это растяжение афферентная импульсация характеризуется значительной асинхронностью, так как многочисленные рецепторы растяжения под влиянием постоянной нагрузки генерируют ритмические импульсы, частота которых определяется индивидуальным порогом каждого рецептора. Мотонейроны получают длительные асинхронные импульсы и сами разряжаются асинхронно. В результате этого мышца отвечает плавным длительным сокращением, автоматически противодействующим силе тяжести. Это определяет большое физиологическое значение рефлекса растяжения как механизма поддержания выпрямленной позы или стояния.
Более сложно организованы рефлекторные ответы, выражающиеся в координированном сгибании или разгибании мышц конечности. Сгибательные рефлексы направлены на избежание различных повреждающих воздействий. Поэтому рецептивное поле сгибательного рефлекса достаточно сложно и включает различные рецепторные образования и различные по скорости проведения афферентные пути. Сгибательный рефлекс возникает при раздражении болевых рецепторов кожи, мышц и внутренних органов. Вовлекаемые при этих раздражениях афферентные волокна имеют широкий спектр скоростей проведения – от миелинизированных волокон группы А до немиелинизированных волокон группы С. Все разнообразные афферентные волокна, импульсация по которым приводит к развитию сгибательного рефлекса, объединяют под названием афферентов сгибательного рефлекса.
Центральное время сгибательного рефлекса довольно продолжительно, что обусловлено его полисинаптическим характером. Сгибательные рефлексы отличаются от собственных рефлексов мышц – миостатических и сухожильных – не только большим числом синаптических переключений на пути к мотонейронам, но и вовлечением ряда мышц, координированное сокращение которых обусловливает движение целой конечности. Одновременно с возбуждением мотонейронов, иннервирующих мышцы-сгибатели, происходит реципрокное торможение мотонейронов мышц-разгибателей.
При достаточно интенсивном раздражении рецепторов задней конечности происходят иррадиация возбуждения и вовлечение в реакцию мышц передней конечности и туловища. При активации мотонейронов противоположной стороны тела наблюдается не сгибание, а разгибание мышц задней конечности – перекрестный разгибательный рефлекс.
К разгибательным рефлексам, кроме уже рассмотренных выше миостатических рефлексов и перекрестного разгибательного рефлекса, относится ряд полисинаптических рефлексов, возникающих при раздражении рецепторов стопы.
Еще более сложный характер имеют ритмические и позные рефлексы, или рефлексы положения. К ритмическим рефлексам у млекопитающих относится чесательный рефлекс. Его аналогом у земноводных является потиральный рефлекс. Ритмические рефлексы характеризуются координированной работой мышц конечностей и туловища, правильным чередованием сгибания и разгибания конечностей наряду с тоническим сокращением приводящих мышц, устанавливающих конечность в определенное положение к кожной поверхности.
Позные рефлексы представляют собой большую группу рефлексов, направленных на поддержание определенной позы, что возможно при наличии определенного мышечного тонуса. Примером позного рефлекса является сгибательный тонический рефлекс, который у лягушки определяет основную позу – сидение. Даже у декапитированной лягушки, подвешенной к штативу, задние конечности всегда несколько согнуты, и сгибательный тонус исчезает только после разрушения спинного мозга или перерезки дорсальных корешков. Различия в тонусе сгибательных мышц особенно отчетливы, если перерезку дорсальных корешков осуществить на одной стороне, оставив другую интактной.
Сгибательный тонический рефлекс наблюдается и у млекопитающих, для которых характерно подогнутое положение конечностей (кролик). В то же время для большинства млекопитающих главное значение для поддержания положения тела играет не сгибательный, а разгибательный рефлекторный тонус.
Ввиду того что особенно важную роль в рефлекторной регуляции разгибательного тонуса играют шейные сегменты спинного мозга, специально выделяют шейные тонические рефлексы положения. Эти рефлексы были впервые описаны голландским физиологом Р. Магнусом (1924).
Рецептивным полем шейных тонических рефлексов являются проприорецепторы мышц шеи и фасций, покрывающих шейный участок позвоночника. Центральная часть рефлекторной дуги имеет полисинаптический характер, т.е. включает вставочные нейроны.
Рефлекторная реакция вовлекает мышцы туловища и конечностей. Кроме спинного мозга, в ней участвуют и моторные ядра мозгового ствола, иннервирующие мышцы глазных яблок.
Шейные тонические рефлексы возникают при поворотах и наклонах головы, что вызывает растяжение мышц шеи и активирует рецептивное поле рефлекса.
Кроме рассмотренных выше рефлексов, которые относятся к категории соматических, так как выражаются в активации скелетных мышц, спинной мозг играет важную роль в рефлекторной регуляции внутренних органов, являясь центром многих висцеральных рефлексов. Эти рефлексы осуществляются при участии расположенных в боковых и вентральных рогах серого вещества преганглионарных нейронов вегетативной нервной системы. Аксоны этих нервных клеток покидают спинной мозг через передние корешки и заканчиваются на клетках симпатических или парасимпатических вегетативных ганглиев. Ганглионарные нейроны в свою очередь посылают аксоны к клеткам различных внутренних органов, включая гладкие мышцы кишечника, сосудов, мочевого пузыря, к железистым клеткам, сердечной мышце.
Заключение
Таким образом, спинной мозг, являясь главным исполнительным отделом центральной нервной системы, в то же время участвует в первичной обработке проприо-, висцеро- и экстероцептивных кожных сигналов, осуществляет координационно-интегративную рефлекторную функцию на сегментарном уровне и обеспечивает обратную афферентацию от проприоцептивного аппарата к управляющим структурам головного мозга.
Список использованной литературы:
1. Алейникова Т.В. и др. Физиология центральной нервной системы. Ростов н/Д.: Феникс, 2000. – 384 с.
2. Гистофизиология кровообращения в спинном мозге. / Под ред. П.А. Мотавкина и др. – М.: Наука, 1994. – 233 с.
3. Принципы настройки в деятельности центральной нервной системы: Место и значение в рефлекторной деятельности и поведении. / Под ред. Р.Н.Порукаева. – М.: Наука, 1991. – 63 с.
4. Физиология центральной нервной системы и сенсорных систем. Хрестоматия. / Под ред. Т.Е. Россолимо и др. – М.: Моск. псих.-соц. ин-т., 1999. – 57 с.
www.ronl.ru
Введение 3
1. Внешнее строение спинного мозга 5
2. Внутреннее строение спинного мозга 8
3. Проводящие пути спинного мозга 10
4. Функции спинного мозга 13
Вывод 16
Список использованной литературы 17
Содержание
В этом заключается рефлекторная функция спинного мозга. Примером рефлекса, что осуществляется спинным мозгом, у человека может быть коленный рефлекс. Спинной мозг участвует и в сложных двигательных реакциях организма. Двигательные нейроны спинного мозга обеспечивают иннервацию всей скелетной мускулатуры, за исключением мышц головы, которые иннервируются черепно-мозговыми нервами. Итак, в спинном мозгу содержатся рефлекторные центры мышц шеи, туловища, конечностей. Поэтому рефлексы спинного мозга играют важное значение в регулировании движений туловища и конечностей. Фактически все двигательные акты человека происходят с участием спинного мозга. Кроме двигательных центров скелетных мышц, в спинном мозге находится ряд симпатических и парасимпатических вегетативных центров. Рефлексы мочеиспускания, дефекации, деятельность половых органов и многие другие тоже связаны с функциями спинного мозга[3].
Однако, так как спинной мозг способен осуществлять различные рефлексы, он не является автономной системой. Его деятельность проходит под постоянным регулирующим влиянием головного мозга. Головной мозг может затормозить, усилить или изменить характер рефлекторных реакций спинного мозга. Так, например, происходит произвольная задержка мочеиспускания, дефекации и др. В целом же спинной мозг является частью сложного аппарата рефлекторной деятельности всей нервной системы организма. Спинной мозг осуществляет и важную проводниковую функцию с участием восходящих и нисходящих путей, расположенных в его белом веществе. Длинными восходящими и нисходящими путями спинной мозг соединяет двусторонними связями периферию с головным мозгом. Афферентные импульсы по проводящим путям спинного мозга проводятся в головной мозг и несут ему информацию обо всех изменениях во внешнем и внутреннем средах организма. По нисходящих путях импульсы от головного мозга передаются кэффекторнымнейронам спинного мозга, которые вызывают или регулируют деятельность соответствующих органов. Спинной мозг покрывают три оболочки, защищающие его от механических и других повреждений.
От оболочек к внутренней стенке позвоночного канала идут связки, фиксирующие спинной мозг. Твердая мозговая оболочка (внешняя) в виде длинного плотного мешка окружает спинной мозг со всех сторон. Сверху она прочно срастается с краями большого затылочного отверстия затылочной кости, а внизу слепо заканчивается на уровне второго крестцового позвонка. Между надкостницей позвоночного канала и этой оболочкой размещен пространство заполнено жировой клетчаткой, которая играет роль эластичной прокладки. Паутинная оболочка находится под твердой. Между этой оболочкой и мягкой расположено крупнейшее щелевидноеподпаутинное (субарахноидальное) пространство, которое, в отличие от других межоболочечных пространств, заполненно мозговой жидкостью. Мягкая, или сосудистая, оболочка (внутренняя) плотно прилегает к внешней поверхности спинного мозга и включает в себя большое количество кровеносных сосудов[5].
Спинномозговые нервы (nn.Spinales) отходят от спинного мозга. Согласно сегментам спинного мозга различают
3. пару спинномозговых нервов: 8 пар шейных,
1. грудных, 5 поясничных, 5 крестцовых и одну пару копчиковых. Все они смешанные и содержат чувствительные, двигательные и вегетативные нервные волокна. Из позвоночного канала спинномозговые нервы выходят через межпозвонковые отверстия. Каждый из них около выхода из межпозвонкового отверстия разветвляется на четыре ветви. Две из них длинные — задняя и передняя, две короткие — оболочечная и соединительная. Оболочковые ветви сразу же возвращаются в позвоночный канал и иннервирующих оболочки спинного мозга. Соединительные ветви соединяются с нервными узлами симпатических стволов. Задние ветви значительно тоньше передних направляются назад между поперечными отростками позвонков, идут самостоятельно до определенных мышц и участков кожи. Они иннервируют глубокие мышцы, надкостницу и кожу задних участков головы, шеи, туловища. Передние ветви спинномозговых нервов значительно толще, иннервируют шею, грудь, живот и конечности. В отличие от задних ветвей, они в большинстве объединяются между собой образуя сплетения. Исключение составляют только передние ветви грудных спинномозговых нервов, большая часть которых не участвует в образовании сплетений.
Различают четыре пары спинномозговых нервных сплетений: шейное, плечевое, поясничное, крестцово-копчиковое.Непосредственно от нервных сплетений отходят отдельные короткие и длинные периферические нервы. Короткие нервы иннервируют мышцы, среди которых размещены сплетение, и мышцы и кожу, расположенные недалеко от них. Длинные нервы иннервируют отдаленные части тела, как, например, верхние и нижние конечности. ВыводОрганы центральной нервной системы состоят из очень нежной ткани. Нервные клетки головного и спинного мозга относят к самым нежным структурам в организме человека, а поэтому они находятся под надежной защитой костей. Никакая другая система органов не имеет такой надежной защиты как мозг. Мозговые оболочки также выполняют защитную функцию. Спинной мозг окружен тремя оболочками и закреплен связями в позвоночном канале, а головной мозг, покрытый такими же оболочками, расположен в костной коробке черепа. Связь центральной нервной системы с органами осуществляется с помощью нервов, отходящих от головного и спинного мозга. От головного мозга отходят
1. пар нервов, которые называются черепно-мозговыми.От спинного мозга отходит
3. пара спинномозговых нервов. Совокупность этих нервных стволов и их многочисленные разветвления, сплетение и узлы составляют периферическую часть нервной системы. Список использованной литературыАнатомия человека.(под ред. В.И. Козлова).
1. Анатомия человека. (под ред. В.И. Козлова).
2. Гранит P. Основы регуляции движении, пер. с англ. М., 1973.
3. Костюк П. Г. Структура и функция нисходящих систем спинного мозга. Л., 1973.
4. Привес М.Г., Лысенков Н.К., Бушкович В.И. Анатомия человека. СПб., Гиппократ, 2000.
5. Яковлев В.А., Феник С.И. Анатомия и эволюция нервной системы, 2003
список литературы
referatbooks.ru
Анатомия и физиология человека
Проводящие пути с точки зрения направления проведения импульса могут быть разделены на две группы - афферентные и эфферентные. Афферентные проводящие пути составляют среднее звено - кондуктор того или иного анализатора...
Вспомогательные органы глаза
Средний мозг состоит из дорсального отдела крыши среднего мозга и вентрального - ножек мозга, которые разграничиваются полостью - водопроводом мозга. Нижней границей среднего мозга на его вентральной поверхности является передний край моста...
Головной мозг
Двигательные функции Корковый отдел двигательного анализатора расположен главным образом в передней центральной извилине, кпереди от центральной борозды. В этой области находятся нервные клетки...
Головной мозг
головной мозг нервная система Выделяют две основные функции спинного мозга: собственную сегментарно-рефлекторную и проводниковую, обеспечивающую связь между головным мозгом, туловищем, конечностями, внутренними органами и др...
Заболевания, обусловленные поражением поясничного отдела головного мозга
Рефлекторные синдромы в области поясничного отдела спинного мозга сопровождают течение многих заболеваний, в частности остеохондроза. Они могут сочетаться также и компрессионными синдромами...
Заболевания, обусловленные поражением поясничного отдела головного мозга
Данный тип заболеваний встречается достаточно часто. Хронические ишемические поражения спинного мозга описываются под названием дискциркуляторной (сосудистой) миелопатии, или хронической спинальной сосудистой недостаточности...
Заболевания, обусловленные поражением поясничного отдела головного мозга
Опухоли пояснично-крестцового отдела спинного мозга можно разделить на опухоли верхнепояоничных сегментов (Li-Lin), эпиконуса (Liv-Sn) и конуса (SIH-SV)...
Принципы передачи информации и структурная организация мозга
Анализ взаимодействия нейронов в сложных путях, вовлекающих в буквальном смысле миллионы нейронов, существенно более труден, чем анализ простых рефлексов. Пере- дача информации в мозг при восприятии звука, прикосновения...
Развитие мозга. Психические свойства человека
Головной мозг, encephalon (cerebrum), с окружающими его оболочками находится в полости мозгового черепа. Выпуклая верхнелатеральная поверхность головного мозга по форме соответствует внутренней вогнутой поверхности свода черепа...
Развитие центральной нервной системы в эмбриогенезе
Медуллярная (нервная) трубка в области будущего спинного мозга представляет собой сначала сравнительно тонкостенную трубку. В краниальных отделах она имеет приблизительно одинаковый диаметр, причем...
Серое и белое вещество головного и спинного мозга
Волокна, составляющие массу белого вещества, имеют разное происхождение. Одни из них представлены отростками нервных клеток серого вещества спинного мозга, другие идут от клеток чувствительных ганглиев, которые лежат вне спинного мозга...
Серое и белое вещество головного и спинного мозга
Серое вещество образовано огромным числом нейронов, сгруппированных в ядра. В нем различают мультиполярные нейроны трех типов: 1...
Серое и белое вещество головного и спинного мозга
Белое вещество спинного мозга выполняет проводниковую функцию, осуществляет передачу нервных импульсов. Она включает три системы проводящих путей - восходящие, нисходящие и собственные пути спинного мозга...
Строение головного мозга
...
Строение спинного мозга
Нервная клетка имеет большое количество отростков. Отростки, удаленные от тела клетки, называются нервными волокнами. Нервные волокна, не выходящие за пределы центральной нервной системы, образуют проводники головного и спинного мозга...
bio.bobrodobro.ru
Он расположен в позвоночном канале и представляет собой слегка уплощенный в переднезаднем направлении белый тяж длиной 40—45 см и толщиной около 1 см. В верхней своей части он переходит в продолговатый мозг, а в нижней оканчивается на уровне 2-го поясничного позвонка.
Делится на отделы: шейный, грудной, поясничный, крестцовый, копчиковый.
Спинной мозг продольными бороздками разделяется на зеркально симметричные правую и левую половины. В центре имеется полость — спинномозговой канал, заполненный жидкостью. Спинной мозг покрыт тремя оболочками: наружной —твердой, средней —паутинной, и внутренней — сосудистой. Твердая оболочка — плотная и прочная соединительнотканная оболочка мозга, состоящая из двух слоев. Наружный слой выстилает кости черепа и позвоночный канал, а внутренний, гладкий и блестящий, обращен к мозгу. Функция твердой оболочки — защитная. Паутинная оболочка представляет собой тонкую мембрану, отделяющую твердую оболочку от сосудистой. Внутренняя сосудистая оболочка богата кровеносными сосудами, проникающими внутрь мозгового вещества. Она плотно прилегает к мозгу, заходя в борозды на его поверхности. Между паутинной и сосудистой оболочками имеется пространство, заполненное спинномозговой жидкостью. Ее назначение — смягчать толчки и ушибы спинного мозга.
Наружный слой спинного мозга представлен белым веществам, состоящим из отростков нейронов. Одни отростки тянутся вдоль спинного мозга и частично проходят в головной мозг, образуя проводящие пути, связывающие нервные центры разных сегментов спинного мозга между собой и с нервными центрами головного мозга. Участок спины, от которого отходит пара нервов, называется сегментом спинного мозга. Проводящие пути делятся на восходящие (чувствительные), передающие возбуждение в головной мозг, и нисходящие (двигательные), проводящие нервные импульсы от головного мозга к рабочим органам. Другие отростки нейронов выходят за пределы спинного мозга, где формируют передние и задние корешки. Передние корешки образованы отростками двигательных нейронов, а задние — чувствительных. Утолщения — ганглии — на задних корешках сформированы скоплениями тел чувствительных нейронов. Выйдя из позвоночного канала через межпозвоночные отверстия, передние и задние корешки объединяются друг с другом и образуют пару смешанных спинномозговых нервов. Их общее число составляет 31 пару. Каждая пара иннервирует определенную группу скелетных мышц и ограниченный участок кожи. В местах выхода спинномозговых нервов к верхним и нижним конечностям спинной мозг имеет два утолщения — шейное и поясничное.
Функции спинного мозга —рефлекторная и проводниковая. В спинном мозге находятся нервные центры (двигательные центры скелетной мускулатуры, сосудодвигательные центры, центры потоотделения, мочеиспускания, дефекации, половой деятельности и др.), которые непосредственно связаны с рецепторами и исполнительными (рабочими) органами. Благодаря этим центрам осуществляются многие простые, не затрагивающие головного мозга рефлексы. Примером такого рефлекса может служить коленный: при легком ударе по сухожилию под коленной чашечкой возникает резкое разгибание согнутой ноги. Все спинномозговые рефлексы являются врожденными, безусловными. Они передаются по наследству и сохраняются в течение всей жизни
Проводниковая функция спинного мозга заключается в проведении центростремительных импульсов к головному мозгу и центробежных импульсов от головного мозга ко всем частям тела. Деятельность спинного мозга контролируется головным мозгом, оказывающим регулирующее влияние на спинномозговые рефлексы.
Возрастные особенности спинного мозга касаются как его топографии, так и строения. Во 2-й половине внутриутробного периода рост спинного мозга отстает от роста позвоночного столба. В детском возрасте рост спинного мозга продолжает отставать. Длина спинного мозга за весь период роста увеличивается в 2.7 раза, преимущественно за счет грудных сегментов. Масса спинного мозга возрастает в 6-7 раз. Серое и белое вещество спинного мозга растут неравномерно, объем серого вещества увеличивается в 5 раз, а объем белого — в 14 раз.
Законченность строения наблюдается уже у плода до рождения. Это объясняется усиленной целенаправленной активностью конечностей новорожденного. Увеличение размеров нейронов спинного мозга наблюдается у детей в школьные годы. Следовательно, двигательная способность младшего школьника в значительной степени зависит от спинного мозга.
www.ronl.ru
