Известно около 5000 видов этого типа, из которых лишь небольшое количество обитает в пресных водах, а остальные — морские животные.
Наиболее примитивные представители типа — одиночные губки — имеют бокаловидую форму. Стенки их тела состоят из двух слоев клеток — наружного и внутреннего, разделенных студневидной мезоглеей. В состав наружного слоя входят в основном эпителиальные клетки, выполняющие роль защитного покрова. Во внутреннем слое имеются очень характерные для губок воротничковые клетки, или хоаноциты, названные так потому, что их передний край вытянут в прозрачный воротничок, окружающий основания жгутика). В мезоглее находятся неспециализированные клетки. Соединение клеток в теле губок, рыхлое и они могут до известной степени менять свое положение.
Стенки тела губок пронизаны множеством мелких пор, которые ведут в полость, окруженную внутренним слоем клеток и открывающуюся наружу более крупным отверстием оскулюмом. Жгутики воротничковых клеток создают ток воды, которая входит через поры в упомянутую полость и выходит наружу через оскулюм. Эту полость не следует считать кишечной, она служит только для прохождения воды и в отличие от настоящей кишечной, или гастральной, полости называется парагастральной. В клетках мезоглеи развиваются скелетные образования разного химического состава: известковые, кремневые, спонгиновые (спонгин — вещество, близкое по химическому строению к шелку). Благодаря скелету губки могут расти вверх, а не растекаться по субстрату, что облегчает движение воды через их тело.
Губки медленно и слабо отвечают на разные раздражения, так как в их теле нет нервных клеток. В теле губок нет мышечных клеток, за исключением тех, которые иногда развиваются вокруг оскулюмов. Поэтому тело губок почти не сократимо, а движение воды через их тело осуществляется, как было сказано выше, только благодаря биению жгутиков воротничковых клеток. Кроме того, некоторые клетки могут передвигаться внутри тела, как амебы, т. е. выпуская ложноножки.
Перенос веществ обеспечивается, во-первых, постоянным движением воды через парагастральную полость, доставляющей внутреннему слою кислород и уносящей продукты диссимиляции, во-вторых, диффузным способом (проникновение переваренной пищи, кислорода и других веществ от слоя к слою).
Получение кислорода совершается через внутреннюю и наружную поверхности тела.
Выделение продуктов диссимиляции тоже происходит через поверхность тела. Кроме того, эти продукты могут скапливаться в некоторых клетках (главным образом в мезоглее), переходя из растворимого состояния в нерастворимое и поэтому переставая оказывать вредное влияние на организм.
Пища в виде мелких организмов или их остатков попадает с водой в парагастральную полость и захватывается воротничковыми клетками, которые способны выпускать ложноножки. Захватывать пищу могут и некоторые клетки наружного слоя, а также мезоглеи. Пищеварение, как и у простейших, внутриклеточное. Разложенные в результате пищеварения вещества частично диффундируют в другие клетки и там усваиваются, а частично ассимилируются на месте.
У губок очень распространено бесполое размножение, которое совершается путем разных видов почкования. Образование колоний происходит в результате того, что почки остаются на материнском организме. Кроме того, образуются особые почки, которые выпадают из материнского организма после его гибели и дают начало новым организмам. Губки размножаются и половым путем. Большинство представителей этого типа — гермафродиты, меньшинство — раздельнополы. Половые клетки развиваются из недифференцированных клеток мезоглеи, а микрогаметы, по новым данным, могут образоваться также из воротничковых клеток. Сперматозоиды выходят в воду и проникают в тело других губок, где оплодотворяют яйцеклетки.
Развитие зиготы происходит через стадии — морула, бластула, паренхимула и у морских губок заканчивается образованием планктонных личинок, плавающих при помощи жгутиков наружных клеток. Личинки, плавая в воде и переносимые течениями, способствуют расселению этих сидячих животных. У пресноводных губок планктонных личинок нет и они распространяются другими способами. При превращении личинок во взрослые сидячие формы у губок происходит извращение зародышевых слоев: наружные жгутиковые клетки мигрируют внутрь, а клетки внутреннего слоя перемещаются наружу. Благодаря такому процессу образуется слой воротничковых клеток, окружающих парагастральную полость, создающий столь важный для описываемых животных ток воды, приносящий пищу и кислород и удаляющий продукты распада обмена веществ.
www.ronl.ru
works.tarefer.ru
Реферат на тему:
Гу́бки (лат. Porifera) — парафилетическая группа организмов; многоклеточные водные, преимущественно морские, модульные беспозвоночные животные, не достигшие уровня тканевой организации. Часто образуют колонии. В настоящее время описано около 8000 видов[1]. Губки известны с докембрия[2]. Настоящие зародышевые листки отсутствуют; тело губки образовано тремя основными группами клеток: пинакодермой — покровным слоем, хоанодермой — выстилкой жгутиковых камер и мезохилом — межклеточным веществом со скелетными элементами. Мышечная и нервная системы отсутствуют. Отличительной чертой губок является наличие водоносной системы, с помощью которой осуществляется дыхание, питание, выделение и размножение. Губки — фильтраторы. Размножение бесполое и половое. У большинства — развитие с метаморфозом: образуются типичные личинки — паренхимула и амфибластула.
В самом простом случае губка имеет бокаловидную форму и обладают гетерополярной осевой симметрией. К субстрату губка крепится подошвой; на верхнем полюсе расположено устье — оскулюм, через которое из тела губки выводится вода. У колониальных осевая симметрия нарушается и возникает множество оскулюмов. Стенки губки пронизаны многочисленными порами, ведущими в парагастральную полость. Тело состоит из двух слоёв клеток: наружного — дермального и внутреннего, выстилающего внутреннюю полость — гастрального. Между ними формируется мезохил — бесструктурное вещество с отдельными разбросанными в нём клетками.
Гистология известковой губки — пинакоциты — хоаноциты — археоциты и другие типы клеток в мезохиле — мезохил — спикулы — карбонатные элементы — субстрат — направление тока воды
Дермальный слой губок — пинакодерма — имеет вид погруженного эпителия и образован тонким (1 мкм) слоем пинакоцитов. Пинакодерма выстилает наружную стенку тела всех губок, а также стенки крупных каналов известковых и обыкновенных губок. В зависимости от месторасположения пинакоциты делятся на несколько групп:
От настоящего эпителия пинакодерма большинства губок отличается отсутствием десмосом и плотных контактов.
Гастральный слой тела губок — хоанодерма — выстилает жгутиковые камеры или парагастральную полость и состоит из хоаноцитов — воротничковых клеток цилиндрической формы. Из центра обращённого в парагастральную полость конца хоаноцитов выходит длинный жгутик, окружённый плазматическим воротничком. Хоаноциты делятся на 2 типа:
| — хоаноцит — пинакоцит — лофоцит — ооцит — археоцит | — склероцит — пороцит — мезохил — спикулы — ток воды |
Мезоглея — бесструктурное вещество, заполняющее пространство между жгутиковыми камерами. Имеет тонковолокнистую структуру и состоит из различных типов клеток:
В состав неклеточного компонента мезохила входят органические молекулы (гликопротеины), свободный коллаген, пищевые частицы и продукты секреционной деятельности клеток.
Основные типы строения водоносной системы губок: сикон, аскон и лейкон (слева направо)
Строение аскона1 — оскулюм2 — спонгиоцель3 — хоанодерма4 — остиум5 — пинакодерма
Строение сикона| 1 — оскулюм2 — спонгиоцель3 — приводящий канал4 — отводящий канал5 — хоанодерма | 6 — прозопиль7 — остиум8 — хоанодерма9 — мезохил10 — апопиль |
У ряда Известковых губок произошли увеличение размера жгутиковых камер и редукция системы приводящих и отводящих каналов. Такие губки состоят из трубки или разветвлённой системы трубок, изнутри выстланной хоаноцитами; при этом поры (остии) открываются непосредственно в жгутиковые камеры. Водоносная система характерна только для двух родов — Leucosolenia и Clathrina.
Формирование сикона вызвано разрастанием мезохила и впячиванием в него участков парагастральной полости, образующих радиальные трубки. Хоаноциты выстилают только жгутиковые трубки и исчезают с других участков атриальной полости. Стенки тела губки утолщаются, и между поверхностью тела и жгутиковыми трубками образуются приводящие каналы. Строение сиконального типа характерно лишь для немногих видов (например, для Scypha и Grantia).
Наиболее сложный тип строения. Это колониальные губки с многочисленными оскулюмами. В мощном слое мезоглей много скелетных элементов. Стенка тела пронизана сетью каналов, связывающих многочисленные жгутиковые камеры.
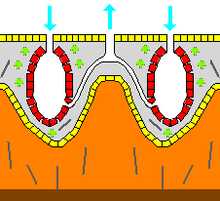
В среднем каждые 5 секунд губка прокачивает количество воды, равное объёму своего тела. Поскольку вода несжимаема, её объём на входе должен быть в каждый момент равен объёму на выходе. С наибольшей скоростью вода течёт в области оскулюма, а с наименьшей — через жгутиковые камеры, так как эти два участка водоносной системы губки имеют соответственно наименьшую и наибольшую общую площадь сечения. Многие губки при необходимости могут снижать общую скорость движения воды или полностью останавливать поток, чтобы избежать попадания ила внутрь организма. Это достигается за счёт регуляции диаметра оскулюма с помощью сократимых миоцитов, закрывания остий (иногда посредством трубковидных пороцитов) или же регуляции биения жгутиков хоаноцитов. Последнее характерно, например, для стеклянных губок, у которых нет ни миоцитов, ни пороцитов.
Ток воды создаётся биением жгутиков хоаноцитов. Каждый жгутик совершает ундулирующие движения в одной плоскости. Расположенные на нижнем участке жгутика в области воротничка лопасти из мастигонем, возможно, помогают «выкачивать» из него воду. По меньшей мере у одного вида (Trochospongilla pennsylvanicus) плоскость биения жгутика хоаноцитов каждые несколько секунд немного смещается, так что в конечном счёте она поворачивается на 360°. Жгутики и воротнички хоаноцитов направлены в противоположную от остий сторону у асконоидных губок или от прозопилей — у сиконоидных и лейконоидных губок. Волна проходит по жгутику от его основания к вершине, создавая ток воды в сторону отводящих каналов и оскулюма.
У многих губок оскулюмы расположены на вершине специальных трубчатых выростов, сильно возвышающихся над основной массой тела, поверхность которого несёт остии. Скорость течения омывающей губку воды в зоне, где находятся оскулюмы, обычно выше, чем в придонном слое, у основания губки. Интенсивное движение воды над трубчатыми выростами создаёт область пониженного давления непосредственно в оскулюмах по сравнению с давлением в остиях. В результате возникает ток воды из области высокого давления в область низкого давления, то есть от остий к оскулюмам. Многие губки обитают в местах, где течения весьма значительны, поэтому дополнительно возникающий градиент давления в водоносной системе усиливает ток, создаваемый работой жгутиков. В свою очередь это позволяет губкам экономить энергию.
Хотя губки в своём абсолютном большинстве сидячие животные, ведущие прикреплённый образ жизни, некоторые виды способны к ограниченной локомоции. И пресноводные (Ephydatia), и морские (Chondrilla, Hymeniacidon, Tethya) губки могут перемещаться по субстрату со скоростью от 1 до 4 мм в день. Это достигается в результате амебоидного движения пинакоцитов и клеток других типов, которое осуществляется одновременно и более или менее согласованно. Губки могут также сокращаться (Clathrina coriacea) и изменять с помощью миоцитов диаметр оскулюма. Благодаря этим проявлениям сократимости останавливается или ограничивается поток воды через водоносную систему в ответ на неблагоприятные воздействия окружающей среды, например при внезапном увеличении количества взвеси ила в воде.
Отличительный признак губок — способность значительной части клеток, образующих тело, свободно перемещаться. Клетки мезохила (все они амебоидные) передвигаются практически постоянно. Эндопинакоциты и хоаноциты тоже могут менять свою локализацию в процессе перестройки водоносной системы. Реорганизация последней проявляется в образовании новых и объединении уже существующих жгутиковых камер, слиянии и образовании новых ветвей каналов. В результате ток воды оптимизируется по мере роста губки или перемен в окружающей среде. Такие независимые и частые перемещения клеток возможны потому, что в тканях губок отсутствуют клеточные контакты.
Губки отфильтровывают пищевые частицы из воды, протекающей через их тело. Размеры частиц обычно варьируют от 50 до 1 мкм, но могут быть и ещё меньше. В эти пределы попадают одноклеточные планктонные организмы, такие, как динофлагелляты и бактерии, вирусы, мелкий органический детрит и, возможно, даже растворённые в воде органические вещества. В тропических морях, где губки очень многочисленны, мелкие фракции пищи примерно в 7 раз более доступны, чем крупные. Все клетки губок способны поглощать частицы путём фагоцитоза.
Фильтры, улавливающие пищевые частицы, — это приводящие каналы, постепенно уменьшающиеся в диаметре по мере погружения в глубь тела, и хоаноциты. Пищевые и иные частицы в зависимости от их диаметра отфильтровываются разными участками системы. Наиболее крупные частицы, диаметр которых превышает примерно 50 мкм, не проходят через остии, но их могут фагоцитировать клетки экзопинакодермы. Частицы в пределах от 5 до 50 осаждаются в приводящих каналах, их поглощают также путём фагоцитоза эндопинакоциты или археоциты. Последние проникают в просвет каналов через их выстилку между пинакоцитами или через пороциты. Мелкие частицы размерного класса бактерий поступают в жгутиковые камеры, где хоаноциты поглощают их путём фагоцитоза или пиноцитоза. Самые мелкие частицы могут улавливаться «ситом», образуемым микроворсинками воротничков хоаноцитов и окружающим их внеклеточным матриксом.
И хоаноциты, и археоциты поглощают и переваривают пищу в пищеварительных вакуолях, однако часто хоаноциты только захватывают частицы, а затем «передают» их археоцитам, в которых и осуществляется процесс пищеварения. Археоциты, по-видимому, также запасают питательные вещества (гликоген и липиды).
К семейству Cladorhizidae (Demospongiae) относятся хищные губки. Они ловят ракообразных и других мелких животных, которые прилипают к раскинутым губкой клейким, состоящим из клеток нитям. Когда добыча попалась, нить укорачивается и подтягивает жертву к поверхности тела. Губка медленно обволакивает пойманное животное и переваривает его, вероятно, с помощью археоцитов. У этих необычных губок нет ни хоаноцитов, ни водоносной системы.
Два основных типа частиц, подлежащих удалению из тела губки, — это непереваренные в процессе внутриклеточного пищеварения остатки пищи и неорганические частицы, попавшие в губку с током воды. Минеральные частицы необходимо удалять из приводящих каналов, иначе они закупорят их и нарушат их деятельность. Застрявшую в приводящем канале частицу фагоцитирует археоцит, который переносит её ближе к выходу из водоносной системы и путём экзоцитоза выводит в отводящий канал. У губок, использующих инородный материал для постройки скелета, археоцит может транспортировать захваченную частицу к месту сборки скелетных элементов.
Многие губки, как морские, так и пресноводные, содержат в тканях фотосинтезирующих эндосимбионтов и используют продукты фотосинтеза для питания. В археоцитах и других клетках пресноводных губок обычно поселяются зелёные водоросли (зоохлореллы). Морские губки (известковые и обыкновенные) могут выступать в роли хозяев динофлагеллят (зооксантелл) или, что бывает чаще, цианобактерий. В спонгиновых волокнах скелета губки Mycale laxissima из Белиза поселяются как зелёные, так и красные водоросли. В некоторых случаях симбиотические цианобактерии составляют до одной трети массы губки (например, у Verongia). Такие губки обитают на мелководных, хорошо освещённых участках дна, и их симбиотические бактерии часто локализуются в поверхностных участках тела. Губка использует избыточные продукты фотосинтеза в виде глицерола и фосфорилированных соединений. Некоторые губки с Большого Барьерного рифа за счёт симбиотических цианобактерий покрывают от 48 до 80 % своих энергетических потребностей. Помимо цианобактерий и других мутуалистических симбионтов в губках часто встречаются внутри- и внеклеточные бактерии, роль которых неясна.
Расстояние между каналами водоносной системы, с помощью которой вентилируется тело губки, и любыми входящими в его состав клетками не превышает 1 мм. Соответственно поступление газов и продуктов обмена (в основном, аммиака) из тела в воду, протекающую по водоносной системе, обеспечивается простой диффузией. Возможно, и питательные вещества из многочисленных участков, где осуществляется внутриклеточное пищеварение, также распространяются по телу губки за счёт диффузии, хотя за их доставку к развивающимся гаметам и тканям ответственны способные к амебоидному движению археоциты. По крайней мере у одного вида Aplysina (Verongia) есть специализированные внутренние волокна, служащие своего рода «рельсами», по которым движутся нагруженные питательными веществами археоциты.
Транспорт питательных веществ у стеклянных губок осуществляется в пределах синцития. После захвата пищевых частиц воротничковыми телами вакуоли с частицами транспортируются по всему синцитию губки вдоль пучков микротрубочек динеиновыми молекулярными моторами. Этот способ транспортировки идентичен транспорту везикул в псевдоподиях фораминифер и в аксонах нервных клеток высших животных.
Практически полное отсутствие межклеточных контактов в пинакодерме и хоанодерме заставляет предполагать, что эти слои образуют довольно ненадёжный регуляторный барьер между мезохилом и водой, омывающей губку снаружи. Вода извне свободно поступает в их тело. Соответственно интерстициальная по своему составу, то есть находящаяся между клетками губки, жидкость должна мало отличаться от воды, в которой губки живут. Это относится и к пресноводным видам. В клетках пресноводных губок, как правило, имеются сократительные вакуоли, но с их помощью осморегуляция осуществляется только в каждой отдельной клетке, а не во всём теле животного.
У губок нет ни нервных клеток, ни нервной ткани, хотя у некоторых видов в ограниченной мере проявляется способность к передаче импульсов. Как правило, эта проводимость представляет собой медленное, «эпителиальное» распространение электрического сигнала на расстояние нескольких миллиметров и приводит к местному сокращению миоцитов в ответ на локальный стимул. Малая скорость передачи сигнала связана с отсутствием специализированных межклеточных контактов (щелевых контактов), улучшающих проводимость эпителиев. Поэтому клеточные мембраны скорее играют роль изолирующих барьеров, нежели обеспечивают прохождение «волны» деполяризации. Стеклянные губки представляют собой исключение из этого общего правила. У Rhabdocalyptus dawsoni электрические импульсы (потенциалы действия) быстро распространяются по синцитиальным тяжам от точки стимуляции по всему телу губки. В результате биение жгутиков прекращается и ток воды через губку останавливается.
Очень часто продукты обмена губок препятствуют поселению других организмов на их поверхности или отпугивают хищников. Было показано, что 9 из 16 антарктических губок и 27 из 36 карибских губок токсичны для рыб. Однако далеко не всегда пагубные для рыб токсины отпугивают и других животных, да и среди рыб встречаются специализированные «губкоядные» виды, например в семействах Pomacanthidae (рыбы-ангелы) и Monacanthidae (единороговые), а также мавританский идол (Zanclus cornutus). Морские черепахи, особенно бисса (Eretmochelys), охотно питаются губками; кремнезёмные спикулы губок могут составлять до 95 % их фекалий. Представители одной из групп голожаберных моллюсков (Doridacea) демонстрируют узкую специализацию, питаясь лишь определёнными видами губок, почти как гусеницы ряда бабочек, которые тесно связаны с конкретными кормовыми растениями. Некоторые губки используют метаболиты в конкурентной борьбе с другими организмами за пространство. Например, карибская губка Chondrilla nucula выделяет вещества, которые убивают находящиеся поблизости кораллы, а губка после этого обрастает их прочные скелеты. Многие губки обладают сильным запахом, например «чесночная» губка Lissodendoryx isodictyalis. Встречаются среди губок и такие, прикосновение к которым вызывает сильное раздражение кожи (например, карибская «огненная» губка Tedania ignis). Выделяемые губками биохимические соединения активно изучаются для определения их медицинского и промышленного значения.
Многие губки становятся убежищами для эндосимбионтов — более мелких организмов, которые живут в их водоносной системе как в многоквартирном доме и используют токи воды и защиту, предоставляемую хозяином. Крупные лейкопоидные «дома» заселены мелкими ракообразными и офиурами. Один исследователь извлёк более 16000 раков-щелкунов (Alpheidae) из каналов водоносной системы одной-единственной крупной бочонковидной губки (Spheciospongia). Некоторые многощетинковые черви из семейства Spionidae поселяются в губках, питаются ими и благодаря этому приобретают цвет приютившей их губки. Острорылый краб (Oregonia gracilis) прикрепляет губок, водоросли и некоторых сидячих животных к своему панцирю, так что на нём образуется своего рода микросообщество. Разрастаясь на этом подвижном субстрате, организмы, формирующие сообщество, надёжно маскируют краба. Другие крабы (Dromiidae) отрезают и прикрепляют к себе на спину, как покрышку, верхнюю часть губки или используют для этой цели лишь её небольшой фрагмент, которым постепенно обрастает весь панцирь, обеспечивая крабу защиту.
Губки из семейства Clionidae (Demospongiae) играют важную роль в разрушении известковых раковин и коралловых рифов в морях. Cliona celata, например, сверлит раковины моллюсков, проделывая в их толще сеть туннелей. Периодически в образовавшиеся отверстия губка выдвигает небольшие участки тела, имеющие вид сосочков (папиллы). Одни папиллы несут остии, другие — оскулюмы. В конце концов губка полностью разрушает всю раковину.
«Сверление» осуществляют специализированные археоциты, которые носят название «травильных» клеток. Каждая такая клетка воздействует на раковину химическими веществами, после чего отделяет от неё маленький кусочек. Образовавшийся «осколок» транспортируется в водоносную систему и выводится наружу через оскулюм. Скрытая внутри раковины или скалы губка надёжно защищена от хищников, поэтому такой способ существования может повышать вероятность выживания как молодых, так и взрослых губок.
Бесполое размножение губок осуществляется путём фрагментации, почкования или образования зимующих пропагул (геммул). Фрагментация, как правило, происходит в результате повреждения губки течениями, прибоем или хищниками. Оторвавшиеся куски выживают благодаря способности губки к регенерации. Фрагмент прикрепляется к субстрату и, перестраиваясь, превращается в нормально функционирующую губку. Крайнее проявление фрагментации — диссоциация губки на отдельные клетки или группы клеток. Это может быть достигнуто путём протирания кусочка губки через мелкоячеистую ткань. Подобный эксперимент был впервые поставлен зоологом Г. В. Уилсоном в начале XX в. и с тех пор часто воспроизводился для изучения механизмов клеточного распознавания и моделирования морфогенеза.
Почкование у губок встречается относительно редко, но всё же имеет место у небольшого числа видов. Например, есть данные, что у Clathrina, имеющей асконоидный тип строения, свободные концы трубок вздуваются и образуют почки, которые затем отделяются, прикрепляются к субстрату и дают начало новой губке. У некоторых видов Tethya почки образуются на особых стебельках. Губки, относящиеся к родам Oscarella и Aplysilla, по некоторым данным, формируют папиллы, затем папиллы отделяются и дают начало новым организмам.
Многие пресноводные и некоторые морские губки образуют сотни, а то и тысячи напоминающих споры геммул. Как правило, это происходит осенью. Осенние геммулы пресноводных видов могут входить в состояние диапаузы, при котором практически полностью подавляется их метаболическая активность. Для активизации таких геммул необходимо, чтобы они в течение определённого периода подвергались воздействию очень низкой температуры. Затем геммулы прорастают и дают начало новым губкам. Обычно это происходит весной. Пока геммула находится в диапаузе, она устойчива к экстремальным воздействиям окружающей среды, таким, как очень низкая температура, повышенная солёность и высыхание.
Геммулы образуются в мезохиле погибающей губки вокруг группы богатых питательными веществами археоцитов. Спонгоциты формируют спонгиновую оболочку вокруг этой клеточной массы. Оболочка может быть инкрустирована спикулами, которые образуются в склероцитах. Она полностью покрывает клеточную массу; лишь на одном полюсе остаётся небольшое отверстие — микропиле. Сформированная геммула состоит из оболочки и находящихся внутри археоцитов, каждый из которых вскоре округляется и становится похож на эмбриональную клетку. Такие модифицированные археоциты называются тезоцитами.
Весной геммулы «прорастают»: периферические тезоциты дифференцируются в пинакодерму, которая, быстро разрастаясь, выступает наружу через микропиле, как надувающийся пузырь. Этот «пузырь» пинакодермы соприкасается с субстратом и прикрепляется к нему. Затем через микропиле внутрь пузыря перемещаются остальные тезоциты, которые составляли центральную часть геммулы. После дифференциации они образуют внутренние структуры молодой губки.
Наблюдения за развитием молодых губок из геммул позволили получить данные, ставящие под сомнение возможность использования по отношению к губкам понятия «индивидуальность». В процессе прорастания тезоциты из разных геммул одной и той же губки или из геммул, принадлежащих разным экземплярам, но обязательно одного вида, могут объединяться. В этом случае из них развивается одна новая губка, обладающая своей «индивидуальностью».
Губки, за редким исключением, — гермафродиты. Когда приходит время, одна губка выбрасывает наружу сперматозоиды, которые переносятся токами воды к другой губке, внутри которой происходит оплодотворение яиц. У яйцекладущих видов (например, Cliona) зиготы выводятся во внешнюю среду, где и протекает их развитие. Многим губкам свойственно живорождение: зиготы остаются внутри материнского организма, который покидают уже сформированные личинки (иногда такие формы называют «личинкородящими»). Зародыши и личинки губок лецитотрофны.
Считается, что у губок нет половых органов (гонад). Половые клетки залегают в мезохиле либо небольшими группами (сперматозоиды), либо поодиночке (яйца). Сперматозоиды образуются из отдельных хоаноцитов или целых жгутиковых камер, которые погружаются в мезохил и окружаются тонкой клеточной стенкой, давая начало сперматоцисте. Яйца образуются из археоцитов или, как у некоторых известковых губок, — из дедифференцированных хоаноцитов. Обычно яйцо накапливает желток, фагоцитируя окружающие их питающие клетки, или клетки-«няньки». Яйцо и клетки-«няньки» иногда заключены в особый фолликул, имеющий клеточные стенки. (По определению, орган состоит из двух и более тканей. Если будет доказано, что клетки стенки цисты или фолликула и половые клетки имеют разное происхождение и формируются из разных тканей, то можно будет сперматоцисты и яйцевые фолликулы считать органами — гонадами.) Поскольку водоносная система одинаково снабжает все части тела, половые клетки довольно равномерно распределены по мезохилу, но при этом всегда находятся недалеко от канала или камеры, в пределах расстояния, на котором действует диффузия.
В период размножения стенка сперматоцисты разрушается, сперматозоиды попадают в отводящие каналы (или в атриум) и выбрасываются наружу через оскулюмы. Некоторые тропические виды внезапно извергают из оскулюмов молочные облачка спермы. Возможно, одновременное высвобождение большого количества сперматозоидов характерно для большинства губок.
Когда выброшенные сперматозоиды оказываются рядом с другой губкой, токи воды заносят их в её водоносную систему. Внутри губки сперматозоиды доставляются к поверхности хоанодермы или в жгутиковые камеры, где их фагоцитируют (но не переваривают) хоаноциты. Хоаноцит, захвативший сперматозоид, теряет жгутик и воротничок и трансформируется в амебоидную клетку, которая называется клеткой-носительницей. Она переносит головку сперматозоида (яйцо) к яйцу. Когда клетка-носительница достигает ближайшего расположенного в мезохиле яйца, она либо «передаёт» ему ядро спермия, либо яйцо фагоцитирует эту клетку вместе с содержащимся в ней ядром спермия. В любом случае оплодотворение происходит внутри «яичника» губки.
Как правило, сперматозоиды губок лишены акросомы — структуры, которая у большинства других животных отвечает за проникновение спермия через мембрану яйцеклетки при оплодотворении. Вероятно акросома не нужна в тех случаях, когда ядро спермия попадает в яйцо путём фагоцитоза. У губки Oscarella lobularis спермии всё же имеют акросому, что может свидетельствовать о существовании у этого вида обычного способа оплодотворения яйцеклетки.
Зигота претерпевает полное и равномерное дробление. Однако взаимное расположение образующихся бластомеров варьирует у разных видов губок. Разнообразны и развивающиеся из зародыша личинки. Они носят названия целобластула, амфибластула, паренхимула и трихимелла.
Личинка целобластула характерна для известковых губок, например видов рода Clathrina (Calcinea). Целобластула представляет собой полый шар, стенка которого состоит из одного слоя жгутиковых клеток. Пока личинка ведёт планктонный образ жизни, некоторые её клетки теряют жгутики, становятся амебоидными и мигрируют в бластоцель, так что он, в конце концов, полностью заполняется клетками. Полая целобластула превращается в лишённую полости стерробластулу.
Личинка амфибластула встречается у других известковых губок, например Grantia, Sycon и Leucosolenia (Calcaronea). Амфибластула представляет собой полый шар, состоящий из клеток двух типов: жгутиковых клеток на переднем полюсе личинки и безжгутиковых клеток с зернистой цитоплазмой на заднем полюсе. Когда личинка ещё находится в мезохиле материнского организма, все жгутики обращены в бластоцель, однако между «зернистыми» клетками на заднем полюсе личинки вскоре образуется отверстие, через которое она выворачивается наизнанку. После инверсии жгутики оказываются на наружной поверхности личинки, которая таким образом приобретает способность плавать с их помощью. На этой стадии личинка покидает материнский организм. Инверсия имеет место у тех губок, яйца которых образуются из хоаноцитов: после оплодотворения клетки делятся так, словно они должны образовать новые жгутиковые камеры, внутрь которых направлены жгутики. Обыкновенные губки Oscarella и Plakina тоже имеют в развитии личинку типа амфибластулы, но она образуется вторично, уже после стадии паренхимы. В своём развитии они проходят стадию плотной морулы, которая позднее в результате выселения клеток из центральной части в наружный клеточный слой (мультиполярная эмиграция) превращается в личинку особого типа — цинктобластулу, которая по своему строению принципиально отличается от типичной амфибластулы, свойственной известковым губкам.
Личинка паренхимула характерна для большинства обыкновенных губок. В этом случае зародыш развивается непосредственно в стерробластулу — клеточную массу без внутренней полости. Наружный слой в основном состоит из жгутиковых клеток, между которыми изредка попадаются клетки, лишённые жгутиков и содержащие в цитоплазме множество везикул. Внутри личинки находятся дифференцированные клетки многих типов: склероциты, колленциты, пинакоциты, даже жгутиковые камеры, а также археоциты. Таким образом, паренхимулу в какой-то степени можно считать «почти готовой» молодой губкой, специализированной для плавания.
Личинка трихимелла — типичная стадия развития стеклянных губок. Трихимелла представляет собой стерробластулу с пояском жгутиковых клеток по экватору. Внутри она заполнена клетками, содержащими желток, склероцитами (со спикулами), клетками других типов и даже жгутиковыми камерами.
Все личинки губок лецитотрофны и, следовательно, живут относительно недолго. Как правило, выход личинок стимулируется светом и обычно происходит на рассвете. По истечении определённого периода, продолжительность которого варьирует от нескольких часов до нескольких дней, личинки оседают и начинают ползать по дну в поисках походящего места для прикрепления. Обнаружив такое место, личинка прикрепляется, претерпевает метаморфоз и превращается в молодую губку. Её строение несколько отличается у разных губок и определяется типом личинки. Поскольку метаморфоз сопровождается перегруппировкой клеток с образованием более или менее чётко выраженных пластов, его часто сравнивают с гаструляцией у других многоклеточных. Однако и погружение клеток в полость личинки, что имеет место у многих губок при формировании стерробластул, также можно считать своего рода гаструляцией.
Попытки найти гаструляцию у губок сопровождают практически всю историю изучения этих организмов. В своё время даже выдвигалась идея об «извращении положения зародышевых листков» у губок. Считалось, что поверхностно расположенные жгутиковые клетки личинок представляют собой эктодерму, которая позднее погружается внутрь тела и даёт начало хоанодерме, а крупные безжгутиковые клетки — это энтодерма, которая остаётся снаружи и превращается в пинакодерму. Отсутствие у губок жёстко детерминированных тканевых дифференцировок позволяет некоторым исследователям говорить о том, что у этих животных не может быть и настоящей гаструляции, в процессе которой осуществляется становление двух первых зародышевых листков — экто- и энтодермы. Процессы обособления нескольких клеточных совокупностей, сопровождающие эмбриональное развитие губок, чисто внешне могут напоминать разные типы гаструляции настоящих многоклеточных животных, но не гомологичны им, точно так же, как не гомологичны ткани и органы Eumetazoa пинакодерме, хоанодерме и мезохилу губок.
Непосредственно перед метаморфозом целобластулы, которая к этому моменту уже превратилась в стерробластулу, её клетки дифференцируются и образуют сплошную массу тотипотентных клеток. После прикрепления клеточная масса распластывается по субстрату. Клетки, расположенные на её поверхности, дают начало пинакодерме, а залегающие более глубоко дифференцируются и дают начало всем остальным типам клеток, характерным для губок. В процессе перераспределения внутренних клеток щелевидные просветы между ними постепенно объединяются, образуя атриум.
Амфибластула оседает на субстрат и прикрепляется к нему передним, несущим жгутики концом. После прикрепления жгутиковые клетки теряют жгутики, мигрируют внутрь и образуют внутренние части губки. «Зернистые» клетки дают начало пинакодерме. Когда молодой организм начинает активно функционировать и питаться, он представляет собой миниатюрную асконоидную трубку. Эта стадия называется олинтус.
Метаморфоз личинок, относимых к парнехимулам, протекает немного по-разному у разных видов. Обычно после прикрепления личинки её внутренние клетки дифференцируются и перераспределяются. Из них образуется большая часть тела губки, если не вся она целиком.
У вида Mycale contarenii жгутиковые клетки превращаются в хоаноциты. У других видов (например, у некоторых пресноводных губок, а также у Microciona prolifera) жгутиковые клетки фагоцитируются археоцитами и соответсвенно не входят в число клеток, принимающих участие в формировании тела молодой губки. В любом случае, прежде чем приобрести лейконоидную организацию, молодая, только что претерпевшая метаморфоз губка часто характеризуется аксоноидным или сиконоидным строением, но имеет при этом толстые стенки. Ювенильные губки, находящиеся на этой стадии развития носят название рагон.
В умеренных широтах губки живут от одного года до нескольких лет. Представители многих тропических видов (а также, возможно, глубоководные губки) могут жить очень долго, до 200 лет и более. Некоторые губки начинают размножаться половым путём только по достижении ими возраста в несколько лет, в то время как другие приступают к размножению в возрасте 2 или 3 недель. Некоторые коралловые губки (Sclerospongia) растут очень медленно, давая прирост около 0,2 мм в год. Если эта скорость роста постоянна, то возраст коралловых губок диаметром 1 м может достигать 5000 лет.
Шестилучевая губка Scolymastra joubini живёт до 10 тысяч лет, это максимальная величина в животном мире.
Первые свидетельства о существовании Hexactinellida относятся к протерозою. В позднем протерозое обнаружены и остатки обыкновенных губок, в том числе и «роговых». Известковых губок и Homoscleromorpha пока удаётся обнаруживать только начиная с палеозоя (кембрий).
Ископаемые «группы» Sphinctozoa и Stromatoporata в настоящее время не рассматриваются как самостоятельные таксоны. По мнению ряда исследователей, это не более чем своего рода морфологические типы, отражающие разные варианты организации тела губок. Это в полной мере относится и к Sclerospongae. Вымершие Archaeocyatha, которых и сейчас рассматривают как самостоятельный класс, по-видимому, ближе всего стояли к обыкновенным губкам. Представители таксона Archaeocyatha имели пористый известковый скелет в форме перевёрнутого полого конуса с радиальными септами и двойной стенкой. Представители Stromatoporata напоминали современных коралловых губок (Sclerospongiae). Они тоже обладали массивным базальным известковым скелетом с внутренними трубками, но в отличие от Sclerospongiae у них не было кремнезёмных спикул. Губки, входящие в группу Sphinctozoa (в основном от ордовика до триаса), имели пористый известковый скелет с кольчатыми перетяжками, напоминающий нитку бус. Изучение современной реликтовой сфинктозойной губки Vaceletia crypta, относящейся к этому же таксону, позволяет сделать вывод о том, что мягкие ткани вымерших Sphinctozoa находились внутри скелета. Черты сходства в строении тканей Vaceletia crypta и обыкновенных губок дают основание относить эту губку (а возможно, и некоторых других Sphinctozoa) к Demospongiae.
Представители Archaeocyatha, Sphinctozoa, Stromatoporata, а также древние Demospongiae в кембрийских и мезозойских морях играли большую роль в образовании рифов. Это заставляет некоторых спонгиологов полагать, что основными тенденциями в эволюции губок были редукция присущих этим древним формам массивных скелетов, из которых формировались рифы, и замена их скелетами из спикул. Низкая скорость роста ныне живущих губок с массивными скелетами по сравнению со скоростью роста кораллоа могла быть причиной эволюционного заката рифообразующих губок. Этой гипотезе противоречит тот факт, что хоанофлагелляты, возможно, имеющие с губками общего предка, строят свой скелет из спикул, состоящих из двуокиси кремния.
Систематика губок весьма противоречива даже в тех случаях, когда ресь идёт о таксонах самого высокого уровня. В последнее время коралловых губок (ранее самостоятельный таксон Sclerospongiae) включают в группу Demospongiae. Недавно было также предложено выделить два подтаксона — Symplasma (Hexactinellida) и Cellularia, поскольку это формально отражает существующую разницу между синцитиальной организацией Hexactinellida и клеточным строением Calcarea и Demospongiae.
wreferat.baza-referat.ru
Губки - многоклеточные водные, главным образом морские, неподвижно прикрепленные ко дну и подводным предметам животные.
В современной фауне насчитывается около 5000 видов губок.
Описано около 5 тыс. видов губок, большинство их обитает в морях (рис. 16). Тип делится на четыре класса: известковые губки (Calcarea), кремнероговые, или обыкновенные, губки (Demospon-gia), стеклянные, или шестилучевые, губки (Hexactinellida, или Hyalospongia) и коралловые губки (Sclerospongia). Последний класс включает небольшое число видов, которые обитают в гротах и туннелях среди коралловых рифов и имеют скелет, состоящий из массивного известкового основания из карбоната кальция и кремневых одноосных игл.
Симметрия отсутствует или имеет место неотчетливая радиальная симметрия. Органы и ткани не выражены, хотя тело построено из разнообразных клеток, выполняющих многие функции, и межклеточного вещества. Внутренние полости выстланы хоаноцитами - особыми жгутиковыми воротничковыми клетками. Нервная система отсутствует. Тело пронизано многочисленными порами и идущими от них каналами, сообщающимися с полостями, выстланными хоаноцитами. Через тело губки осуществляется непрерывный ток воды. Почти все обладают сложным минеральным (СаСО3, SiO2) или органическим скелетом.
Среда обитания
Существует более трех тысяч видов губок, которые живут, прикрепившись к морскому дну, как в холодных, так и в теплых морях; лишь немногие корневые, то есть без спикул, губки обитают в пресных водах. Некоторые морские губки встречаются на небольшой глубине, другие живут на глубине от 100 до 1000 м.
Строение.
Губка имеет бокаловидную форму и обладают гетерополярной осевой симметрией.
К субстрату губка крепится подошвой; на верхнем полюсе расположено устье — оскулюм, через которое из тела губки выводится вода. У колониальных осевая симметрия нарушается и возникает множество оскулюмов.
Стенки губки пронизаны многочисленными порами, ведущими в парагастральную полость.
Тело состоит из двух слоев клеток: наружного - дермального (эктодерма) и внутреннего, выстилающего внутреннюю полость, - гастрального (энтодерма). Между ними выделяется мезоглея - слой особого бесструктурного вещества с отдельными разбросанными в нем клетками. У большинства губок мезоглея сильно утолщается. В мезоглее формируется также скелет.
Наружный слой тела губок образован плоскими покровными клетками – пинакоцитами, среди которых имеются крупные цилиндрические клетки, пронизывающие тело насквозь, – пороциты. Каналы пороцитов соединяют внешнюю среду с внутренней полостью. Внутренний слой тела образуют жгутиковые клетки – хоаноциты. Они имеют цилиндрическую форму, а из центра свободного, торчащего в парагастральную полость конца клетки выдается длинный жгутик, основание которого окружено цитоплазматическим воротничком. Клетки, свободно рассеянные в мезоглее, подразделяются на неподвижные звездчатые, выполняющие опорную функцию (колленциты), скелетные подвижные (склеробласты), занятые перевариванием пищи (амебоциты), резервные амебоидные, которые могут превращаться в любой из названных типов, и половые. Способность клеточных элементов переходить друг в друга говорит об отсутствии дифференцированных тканей.
По строению стенки тела и канальной системы, а также месту расположения участков жгутиконосного слоя различают три типа губок:
Аксон
В наиболее простом случае тело губки имеет вид небольшого тонкостенного бокала или мешочка, основанием прикрепленного к субстрату, а отверстием, которое называется устьем или оскулумом, обращенного кверху
Сикон
Формирование сикона вызвано разрастанием мезохила и впячиванием в него участков парагастральной полости, образующих радиальные трубки. Хоаноциты выстилают только жгутиковые трубки и исчезают с других участков атриальной полости. Стенки тела губки утолщаются, и между поверхностью тела и жгутиковыми трубками образуются приводящие каналы. Строение сиконального типа характерно лишь для немногих видов (например, для Scypha и Grantia).
Лейкон
Наиболее сложный тип губок – лейкон. Для губок этого типа характерен мощный слой мезоглеи с множеством скелетных элементов. Внутренние впячивания погружаются вглубь мезоглеи и приобретают вид жгутиковых камер, соединяющихся выносящими каналами сатриальной полостью. Атриальная полость у лейконоидных губок, так же как у сиконоидных выстлана пинакоцитами. Лейконоидные губки обычно образуют колонии с множеством устьев на поверхности: в виде корок, пластинок, комьев, кустов. Ток воды в лейконоидной губке осуществляется по следующему пути: поры → поровые каналы → жгутиковые камеры → выносящие каналы → атриальная полость → оскулум.
Скелет.
У немногих губок тело остается совершенно мягким у большинства скелет твердый и служит для опоры тела и стенок канальной системы.
Скелет губок образуется в мезоглее.
Скелет состоит либо из минерального вещества: углекислой извести или кремнезема, либо из органического вещества спонгина, либо из сочетания кремнезема и спонгина.
Минеральный скелет состоит из отдельных или спаянных между собой игл (спикул), формирующихся внутри клетоксклеробластов.
В цитоплазме склеробласта появляется маленькое зернышко, которое увеличивается, разрастается и образует правильной формы скелетную иглу. Во время роста игла окружена цитоплазмой склеробласта, которая одевает иглу тончайшим слоем. Рост происходит путем отложения на поверхности иглы новых слоев минерального вещества. Когда игла достигает предельных размеров, рост ее прекращается, склеробласт отмирает и игла остается свободно лежать в мезоглее.
Роговой, или спонгиновый, скелет состоит из сильно разветвляющейся внутри мезоглеи сети роговых волокон желтоватого цвета. Химический состав спонгина близок к шелку, притом с некоторым, иногда довольно значительным (до 14%) содержанием йода. Он формируется несколько иначе‚ чем минеральный. Растущие волокна скелета окружены сплошным футляром из мелких клеток-
скелетообразовательниц, так что образование волокон идет не внутриклеточно (как в случае игл), а межклеточно. Электронно-микроскопические исследования показали‚ что тяжи спонгина слагаются из тончайших субмикроскопических фибриллей, обладающих поперечной исчерченностью (как коллагеновые волокна в соединительной ткани высших животных).
Имеются губки‚ совершенно лишенные скелета. Бесскелетные губки очень мелкие—свидетельство опорного значения скелета, без которого губки не могут разрастаться
Движение.
Как уже отмечалось, губки — неподвижные животные и не способны к каким-либо изменениям формы тела. Лишь при довольно сильном раздражении у некоторых губок наблюдается очень медленное сужение отверстий (устий и пор) и просветов каналов, которое достигается сокращением миоцитов или протоплазмы других клеток.
Большинство клеток в теле губок способно выпускать и втягивать ложноножки, или псевдоподии, или даже с их помощью передвигаться в толще мезоглеи. Особенной подвижностью отличаются амебоциты, перемещающиеся иногда со скоростью до 20 мк в минуту.
Дыхание.
Как и большинство животных, обитающих в водной среде, губки используют для дыхания растворенный в воде кислород. Ток воды, проникающий во все полости и каналы губки, снабжает близлежащие клетки и мезоглею кислородом и уносит выделяемую ими углекислоту. Таким образом, газовый обмен с наружной средой осуществляется у губок непосредственно каждой клеткой или через мезоглею.
Питание.
Губки питаются главным образом взвешенными в воде остатками отмерших животных и растений, а также мелкими одноклеточными организмами, они не обладают избирательной способностью, а поглощают все взвешенное в воде.
Размер пищевых частиц обычно не превышает 10 мк. Частицы пищи приносятся с током воды к жгутиковым камерам, где они захватываются хоаноцитами и затем поступают в мезоглею. Здесь пища попадает к амебоцитам, которые разносят ее по всем частям тела губки. Внутри этих клеток, в пищеварительных вакуолях, образующихся вокруг захваченных частиц, происходит переваривание пищи. Захваченная частица растворяется, а на поверхности вакуоли появляются зерна жироподобного вещества. Так происходит переваривание и усвоение пищевого материала клетками губок.
Размножение и развитие.
Большая часть губок (в том числе все известковые) гермафродиты, часть видов
раздельнополы. Половые клетки их происходят из амебойдных клеток
(археоцитов)‚ ползающих в мезоглее. Они залегают в мезоглее под энтодермой
жгутиковых камер.
При половом размножении из оплодотворенной яйцеклетки развивается бластула, состоящая из одного слоя клеток, имеющих жгутики (рис. 3). Затем часть клеток мигрирует внутрь и превращается в амебоидные клетки. После того, как личинка оседает на дно, происходит перемещение жгутиковых клеток внутрь, они становятся хоаноцитами, а амебоидные клетки выходят на поверхность и превращаются в пинакоциты.
Далее личинка превращается в молодую губку. То есть первичная эктодерма (мелкие жгутиковые клетки) становится на место энтодермы, а энтодерма – на место эктодермы: зародышевые пласты меняются местами. На этом основании зоологи называют губок животными, вывернутыми наизнанку (Enantiozoa).
Губки в высокой степени способны к регуляции. При удалении отдельных участков
тела происходит их восстановление. Если губку растереть или даже протереть
через сито‚ то образовавшаяся кашица‚ состоящая из отдельных клеток и групп
клеток‚ оказывается способной к восстановлению целого организма. При этом
клетки‚ активно двигаясь‚ собираются вместе‚ а затем из клеточного скопления
формируется маленькая губка. Процесс формирования целого организма из
скопления соматических клеток называют соматическим эмбриогенезом. Продолжительность жизни, или возраст, которого достигают губки, колеблется у разных видов от нескольких недель и месяцев до многих лет. Известковые губки обычно живут в среднем до одного года. Некоторые из них (Sycon coronatum, Grantia compressa) отмирают сразу же по достижении половой зрелости, как только сформировавшиеся личинки нового поколения покидают их организм. Большинство мелких четырехлучевых и кремнероговых губок живет в пределах 1—2 лет. Более крупные стеклянные и обыкновенные губки относятся к долгоживущим организмам. Особой долговечностью отличаются те из них, которые достигают величины 0, 5 м и более. Так, экземпляры конской губки (Hippospongia communis) около 1 м в диаметре, по мнению специалистов, достигают возраста не менее 50 лет.
myunivercity.ru
Губки настолько не похожи на других многоклеточных животных, что долгое время их считали представителями особой группы «зоофитов», то есть животно-растений. Действительно, они ведут прикрепленный образ жизни, неспособны совершать активные движения, у них отсутствует нервная система и органы чувств. К тому же отдельные их представители могут иметь зеленую окраску, поскольку в их клетках поселяются водоросли.
Известно приблизительно 9 тыс. видов этих удивительных существ, распространенных в морях и пресных водоемах.
Впервые детально строение и процессы жизнедеятельности губок исследовал Р. Е. Грант, который и предложил научное название этой группы животных.
Особенности строения губок. Среди губок имеются одиночные формы, но большинство видов образует колонии, размеры которых могут достигать 2 м. Колонии губок своей формой могут напоминать кустики, корковые наросты, комочки и т.д., обрастающие различные поверхности. Окраска также разнообразна — желтая, коричневая, белая, красная, фиолетовая или зеленая.
Имеются данные, что на поверхности контейнеров с отработанным ядерным топливом, захороненных на морском дне, обитают гигантские губки.
В пресных водоемах встречаются различные виды бадяг. Их колонии часто образуются вокруг погруженных в воду предметов. В стоячих водоемах они имеют форму кустика, в проточных — похожи на корковое обрастание. Окраска колонии — серая или грязно-зелёная.
Тело губки бокаловидной формы (рис. 58, 1). Своей нижней частью животные прикрепляются к подводным предметам. С помощью специальной видеосъемки установлено, что некоторые губки могут передвигаться за счет амебоидных клеток. Но даже самые быстрые из них за сутки не преодолевают расстояние более 1 мм.
На противоположном — верхнем — конце тела губки находится отверстие. Но это не рот. Если в аквариум с губками насыпать растертую сухую тушь, то ее частички сначала направятся к телу губки, потом через канальцы в стенках тела попадут внутрь и, в конце концов, выведутся через отверстие на верхнем конце тела.
Таким образом, это отверстие служит не для поглощения пищи, а для выведения из тела воды с ее непереваренными остатками.
Тело губок состоит из клеток разных типов. Но тканей они не образуют. Каждая клетка функционирует самостоятельно.
| Рис. 58. Строение губки (1) и клетки со жгутиками (2) |
Наружный слой тела губок образуют клетки, напоминающие клетки покровного эпителия других многоклеточных животных. Среди клеток наружного слоя есть и такие, которые имеют пору. Этими порами начинается система канальцев, пронизывающая стенки тела. Отверстия этих канальцев окружены клетками, способными сокращаться и закрывать их. Канальцы проводят воду с частицами пищи к внутренней полости. Эта полость обычно выстелена особыми клетками со жгутиками, основа которых окружена пленчатым воротничком (рис. 58, 2). Такие клетки образуют внутренний слой. У многих губок они расположены внутри стенок тела, образуя жгутиковые камеры. Работа жгутиков обеспечивает движение воды по системе канальцев и внутренней полости.
Между наружным и внутренним слоями клеток находится межклеточное вещество, в котором расположены разные типы клеток. Некоторые из них образуют внутренний скелет губок.
Еще один тип клеток — амебоидные. Эти клетки с помощью ложноножек захватывают частицы пищи, которые перевариваются в их пищеварительных вакуолях. Передвигаясь по телу губки, амебоидные клетки распределяют питательные вещества. Материал с сайта http://worldofschool.ru
В крупных вакуолях особых клеток многих видов губок, обитающих на небольших глубинах при достаточном освещении, поселяются особые виды цианобактерий. Эти прокариоты могут составлять до 50% массы клеток самой губки. Они поставляют кислород и синтезированные органические вещества, а получают от животных необходимый для фотосинтеза углекислый газ и защиту от врагов.
Строению губок характерны такие признаки:
worldofschool.ru
Реферат на тему:
Гу́бки (лат. Porifera) — парафилетическая группа организмов; многоклеточные водные, преимущественно морские, модульные беспозвоночные животные, не достигшие уровня тканевой организации. Часто образуют колонии. В настоящее время описано около 8000 видов[1]. Губки известны с докембрия[2]. Настоящие зародышевые листки отсутствуют; тело губки образовано тремя основными группами клеток: пинакодермой — покровным слоем, хоанодермой — выстилкой жгутиковых камер и мезохилом — межклеточным веществом со скелетными элементами. Мышечная и нервная системы отсутствуют. Отличительной чертой губок является наличие водоносной системы, с помощью которой осуществляется дыхание, питание, выделение и размножение. Губки — фильтраторы. Размножение бесполое и половое. У большинства — развитие с метаморфозом: образуются типичные личинки — паренхимула и амфибластула.
В самом простом случае губка имеет бокаловидную форму и обладают гетерополярной осевой симметрией. К субстрату губка крепится подошвой; на верхнем полюсе расположено устье — оскулюм, через которое из тела губки выводится вода. У колониальных осевая симметрия нарушается и возникает множество оскулюмов. Стенки губки пронизаны многочисленными порами, ведущими в парагастральную полость. Тело состоит из двух слоёв клеток: наружного — дермального и внутреннего, выстилающего внутреннюю полость — гастрального. Между ними формируется мезохил — бесструктурное вещество с отдельными разбросанными в нём клетками.
Гистология известковой губки — пинакоциты — хоаноциты — археоциты и другие типы клеток в мезохиле — мезохил — спикулы — карбонатные элементы — субстрат — направление тока воды
Дермальный слой губок — пинакодерма — имеет вид погруженного эпителия и образован тонким (1 мкм) слоем пинакоцитов. Пинакодерма выстилает наружную стенку тела всех губок, а также стенки крупных каналов известковых и обыкновенных губок. В зависимости от месторасположения пинакоциты делятся на несколько групп:
От настоящего эпителия пинакодерма большинства губок отличается отсутствием десмосом и плотных контактов.
Гастральный слой тела губок — хоанодерма — выстилает жгутиковые камеры или парагастральную полость и состоит из хоаноцитов — воротничковых клеток цилиндрической формы. Из центра обращённого в парагастральную полость конца хоаноцитов выходит длинный жгутик, окружённый плазматическим воротничком. Хоаноциты делятся на 2 типа:
| — хоаноцит — пинакоцит — лофоцит — ооцит — археоцит | — склероцит — пороцит — мезохил — спикулы — ток воды |
Мезоглея — бесструктурное вещество, заполняющее пространство между жгутиковыми камерами. Имеет тонковолокнистую структуру и состоит из различных типов клеток:
В состав неклеточного компонента мезохила входят органические молекулы (гликопротеины), свободный коллаген, пищевые частицы и продукты секреционной деятельности клеток.
Основные типы строения водоносной системы губок: сикон, аскон и лейкон (слева направо)
Строение аскона1 — оскулюм2 — спонгиоцель3 — хоанодерма4 — остиум5 — пинакодерма
Строение сикона| 1 — оскулюм2 — спонгиоцель3 — приводящий канал4 — отводящий канал5 — хоанодерма | 6 — прозопиль7 — остиум8 — хоанодерма9 — мезохил10 — апопиль |
У ряда Известковых губок произошли увеличение размера жгутиковых камер и редукция системы приводящих и отводящих каналов. Такие губки состоят из трубки или разветвлённой системы трубок, изнутри выстланной хоаноцитами; при этом поры (остии) открываются непосредственно в жгутиковые камеры. Водоносная система характерна только для двух родов — Leucosolenia и Clathrina.
Формирование сикона вызвано разрастанием мезохила и впячиванием в него участков парагастральной полости, образующих радиальные трубки. Хоаноциты выстилают только жгутиковые трубки и исчезают с других участков атриальной полости. Стенки тела губки утолщаются, и между поверхностью тела и жгутиковыми трубками образуются приводящие каналы. Строение сиконального типа характерно лишь для немногих видов (например, для Scypha и Grantia).
Наиболее сложный тип строения. Это колониальные губки с многочисленными оскулюмами. В мощном слое мезоглей много скелетных элементов. Стенка тела пронизана сетью каналов, связывающих многочисленные жгутиковые камеры.
В среднем каждые 5 секунд губка прокачивает количество воды, равное объёму своего тела. Поскольку вода несжимаема, её объём на входе должен быть в каждый момент равен объёму на выходе. С наибольшей скоростью вода течёт в области оскулюма, а с наименьшей — через жгутиковые камеры, так как эти два участка водоносной системы губки имеют соответственно наименьшую и наибольшую общую площадь сечения. Многие губки при необходимости могут снижать общую скорость движения воды или полностью останавливать поток, чтобы избежать попадания ила внутрь организма. Это достигается за счёт регуляции диаметра оскулюма с помощью сократимых миоцитов, закрывания остий (иногда посредством трубковидных пороцитов) или же регуляции биения жгутиков хоаноцитов. Последнее характерно, например, для стеклянных губок, у которых нет ни миоцитов, ни пороцитов.
Ток воды создаётся биением жгутиков хоаноцитов. Каждый жгутик совершает ундулирующие движения в одной плоскости. Расположенные на нижнем участке жгутика в области воротничка лопасти из мастигонем, возможно, помогают «выкачивать» из него воду. По меньшей мере у одного вида (Trochospongilla pennsylvanicus) плоскость биения жгутика хоаноцитов каждые несколько секунд немного смещается, так что в конечном счёте она поворачивается на 360°. Жгутики и воротнички хоаноцитов направлены в противоположную от остий сторону у асконоидных губок или от прозопилей — у сиконоидных и лейконоидных губок. Волна проходит по жгутику от его основания к вершине, создавая ток воды в сторону отводящих каналов и оскулюма.
У многих губок оскулюмы расположены на вершине специальных трубчатых выростов, сильно возвышающихся над основной массой тела, поверхность которого несёт остии. Скорость течения омывающей губку воды в зоне, где находятся оскулюмы, обычно выше, чем в придонном слое, у основания губки. Интенсивное движение воды над трубчатыми выростами создаёт область пониженного давления непосредственно в оскулюмах по сравнению с давлением в остиях. В результате возникает ток воды из области высокого давления в область низкого давления, то есть от остий к оскулюмам. Многие губки обитают в местах, где течения весьма значительны, поэтому дополнительно возникающий градиент давления в водоносной системе усиливает ток, создаваемый работой жгутиков. В свою очередь это позволяет губкам экономить энергию.
Хотя губки в своём абсолютном большинстве сидячие животные, ведущие прикреплённый образ жизни, некоторые виды способны к ограниченной локомоции. И пресноводные (Ephydatia), и морские (Chondrilla, Hymeniacidon, Tethya) губки могут перемещаться по субстрату со скоростью от 1 до 4 мм в день. Это достигается в результате амебоидного движения пинакоцитов и клеток других типов, которое осуществляется одновременно и более или менее согласованно. Губки могут также сокращаться (Clathrina coriacea) и изменять с помощью миоцитов диаметр оскулюма. Благодаря этим проявлениям сократимости останавливается или ограничивается поток воды через водоносную систему в ответ на неблагоприятные воздействия окружающей среды, например при внезапном увеличении количества взвеси ила в воде.
Отличительный признак губок — способность значительной части клеток, образующих тело, свободно перемещаться. Клетки мезохила (все они амебоидные) передвигаются практически постоянно. Эндопинакоциты и хоаноциты тоже могут менять свою локализацию в процессе перестройки водоносной системы. Реорганизация последней проявляется в образовании новых и объединении уже существующих жгутиковых камер, слиянии и образовании новых ветвей каналов. В результате ток воды оптимизируется по мере роста губки или перемен в окружающей среде. Такие независимые и частые перемещения клеток возможны потому, что в тканях губок отсутствуют клеточные контакты.
Губки отфильтровывают пищевые частицы из воды, протекающей через их тело. Размеры частиц обычно варьируют от 50 до 1 мкм, но могут быть и ещё меньше. В эти пределы попадают одноклеточные планктонные организмы, такие, как динофлагелляты и бактерии, вирусы, мелкий органический детрит и, возможно, даже растворённые в воде органические вещества. В тропических морях, где губки очень многочисленны, мелкие фракции пищи примерно в 7 раз более доступны, чем крупные. Все клетки губок способны поглощать частицы путём фагоцитоза.
Фильтры, улавливающие пищевые частицы, — это приводящие каналы, постепенно уменьшающиеся в диаметре по мере погружения в глубь тела, и хоаноциты. Пищевые и иные частицы в зависимости от их диаметра отфильтровываются разными участками системы. Наиболее крупные частицы, диаметр которых превышает примерно 50 мкм, не проходят через остии, но их могут фагоцитировать клетки экзопинакодермы. Частицы в пределах от 5 до 50 осаждаются в приводящих каналах, их поглощают также путём фагоцитоза эндопинакоциты или археоциты. Последние проникают в просвет каналов через их выстилку между пинакоцитами или через пороциты. Мелкие частицы размерного класса бактерий поступают в жгутиковые камеры, где хоаноциты поглощают их путём фагоцитоза или пиноцитоза. Самые мелкие частицы могут улавливаться «ситом», образуемым микроворсинками воротничков хоаноцитов и окружающим их внеклеточным матриксом.
И хоаноциты, и археоциты поглощают и переваривают пищу в пищеварительных вакуолях, однако часто хоаноциты только захватывают частицы, а затем «передают» их археоцитам, в которых и осуществляется процесс пищеварения. Археоциты, по-видимому, также запасают питательные вещества (гликоген и липиды).
К семейству Cladorhizidae (Demospongiae) относятся хищные губки. Они ловят ракообразных и других мелких животных, которые прилипают к раскинутым губкой клейким, состоящим из клеток нитям. Когда добыча попалась, нить укорачивается и подтягивает жертву к поверхности тела. Губка медленно обволакивает пойманное животное и переваривает его, вероятно, с помощью археоцитов. У этих необычных губок нет ни хоаноцитов, ни водоносной системы.
Два основных типа частиц, подлежащих удалению из тела губки, — это непереваренные в процессе внутриклеточного пищеварения остатки пищи и неорганические частицы, попавшие в губку с током воды. Минеральные частицы необходимо удалять из приводящих каналов, иначе они закупорят их и нарушат их деятельность. Застрявшую в приводящем канале частицу фагоцитирует археоцит, который переносит её ближе к выходу из водоносной системы и путём экзоцитоза выводит в отводящий канал. У губок, использующих инородный материал для постройки скелета, археоцит может транспортировать захваченную частицу к месту сборки скелетных элементов.
Многие губки, как морские, так и пресноводные, содержат в тканях фотосинтезирующих эндосимбионтов и используют продукты фотосинтеза для питания. В археоцитах и других клетках пресноводных губок обычно поселяются зелёные водоросли (зоохлореллы). Морские губки (известковые и обыкновенные) могут выступать в роли хозяев динофлагеллят (зооксантелл) или, что бывает чаще, цианобактерий. В спонгиновых волокнах скелета губки Mycale laxissima из Белиза поселяются как зелёные, так и красные водоросли. В некоторых случаях симбиотические цианобактерии составляют до одной трети массы губки (например, у Verongia). Такие губки обитают на мелководных, хорошо освещённых участках дна, и их симбиотические бактерии часто локализуются в поверхностных участках тела. Губка использует избыточные продукты фотосинтеза в виде глицерола и фосфорилированных соединений. Некоторые губки с Большого Барьерного рифа за счёт симбиотических цианобактерий покрывают от 48 до 80 % своих энергетических потребностей. Помимо цианобактерий и других мутуалистических симбионтов в губках часто встречаются внутри- и внеклеточные бактерии, роль которых неясна.
Расстояние между каналами водоносной системы, с помощью которой вентилируется тело губки, и любыми входящими в его состав клетками не превышает 1 мм. Соответственно поступление газов и продуктов обмена (в основном, аммиака) из тела в воду, протекающую по водоносной системе, обеспечивается простой диффузией. Возможно, и питательные вещества из многочисленных участков, где осуществляется внутриклеточное пищеварение, также распространяются по телу губки за счёт диффузии, хотя за их доставку к развивающимся гаметам и тканям ответственны способные к амебоидному движению археоциты. По крайней мере у одного вида Aplysina (Verongia) есть специализированные внутренние волокна, служащие своего рода «рельсами», по которым движутся нагруженные питательными веществами археоциты.
Транспорт питательных веществ у стеклянных губок осуществляется в пределах синцития. После захвата пищевых частиц воротничковыми телами вакуоли с частицами транспортируются по всему синцитию губки вдоль пучков микротрубочек динеиновыми молекулярными моторами. Этот способ транспортировки идентичен транспорту везикул в псевдоподиях фораминифер и в аксонах нервных клеток высших животных.
Практически полное отсутствие межклеточных контактов в пинакодерме и хоанодерме заставляет предполагать, что эти слои образуют довольно ненадёжный регуляторный барьер между мезохилом и водой, омывающей губку снаружи. Вода извне свободно поступает в их тело. Соответственно интерстициальная по своему составу, то есть находящаяся между клетками губки, жидкость должна мало отличаться от воды, в которой губки живут. Это относится и к пресноводным видам. В клетках пресноводных губок, как правило, имеются сократительные вакуоли, но с их помощью осморегуляция осуществляется только в каждой отдельной клетке, а не во всём теле животного.
У губок нет ни нервных клеток, ни нервной ткани, хотя у некоторых видов в ограниченной мере проявляется способность к передаче импульсов. Как правило, эта проводимость представляет собой медленное, «эпителиальное» распространение электрического сигнала на расстояние нескольких миллиметров и приводит к местному сокращению миоцитов в ответ на локальный стимул. Малая скорость передачи сигнала связана с отсутствием специализированных межклеточных контактов (щелевых контактов), улучшающих проводимость эпителиев. Поэтому клеточные мембраны скорее играют роль изолирующих барьеров, нежели обеспечивают прохождение «волны» деполяризации. Стеклянные губки представляют собой исключение из этого общего правила. У Rhabdocalyptus dawsoni электрические импульсы (потенциалы действия) быстро распространяются по синцитиальным тяжам от точки стимуляции по всему телу губки. В результате биение жгутиков прекращается и ток воды через губку останавливается.
Очень часто продукты обмена губок препятствуют поселению других организмов на их поверхности или отпугивают хищников. Было показано, что 9 из 16 антарктических губок и 27 из 36 карибских губок токсичны для рыб. Однако далеко не всегда пагубные для рыб токсины отпугивают и других животных, да и среди рыб встречаются специализированные «губкоядные» виды, например в семействах Pomacanthidae (рыбы-ангелы) и Monacanthidae (единороговые), а также мавританский идол (Zanclus cornutus). Морские черепахи, особенно бисса (Eretmochelys), охотно питаются губками; кремнезёмные спикулы губок могут составлять до 95 % их фекалий. Представители одной из групп голожаберных моллюсков (Doridacea) демонстрируют узкую специализацию, питаясь лишь определёнными видами губок, почти как гусеницы ряда бабочек, которые тесно связаны с конкретными кормовыми растениями. Некоторые губки используют метаболиты в конкурентной борьбе с другими организмами за пространство. Например, карибская губка Chondrilla nucula выделяет вещества, которые убивают находящиеся поблизости кораллы, а губка после этого обрастает их прочные скелеты. Многие губки обладают сильным запахом, например «чесночная» губка Lissodendoryx isodictyalis. Встречаются среди губок и такие, прикосновение к которым вызывает сильное раздражение кожи (например, карибская «огненная» губка Tedania ignis). Выделяемые губками биохимические соединения активно изучаются для определения их медицинского и промышленного значения.
Многие губки становятся убежищами для эндосимбионтов — более мелких организмов, которые живут в их водоносной системе как в многоквартирном доме и используют токи воды и защиту, предоставляемую хозяином. Крупные лейкопоидные «дома» заселены мелкими ракообразными и офиурами. Один исследователь извлёк более 16000 раков-щелкунов (Alpheidae) из каналов водоносной системы одной-единственной крупной бочонковидной губки (Spheciospongia). Некоторые многощетинковые черви из семейства Spionidae поселяются в губках, питаются ими и благодаря этому приобретают цвет приютившей их губки. Острорылый краб (Oregonia gracilis) прикрепляет губок, водоросли и некоторых сидячих животных к своему панцирю, так что на нём образуется своего рода микросообщество. Разрастаясь на этом подвижном субстрате, организмы, формирующие сообщество, надёжно маскируют краба. Другие крабы (Dromiidae) отрезают и прикрепляют к себе на спину, как покрышку, верхнюю часть губки или используют для этой цели лишь её небольшой фрагмент, которым постепенно обрастает весь панцирь, обеспечивая крабу защиту.
Губки из семейства Clionidae (Demospongiae) играют важную роль в разрушении известковых раковин и коралловых рифов в морях. Cliona celata, например, сверлит раковины моллюсков, проделывая в их толще сеть туннелей. Периодически в образовавшиеся отверстия губка выдвигает небольшие участки тела, имеющие вид сосочков (папиллы). Одни папиллы несут остии, другие — оскулюмы. В конце концов губка полностью разрушает всю раковину.
«Сверление» осуществляют специализированные археоциты, которые носят название «травильных» клеток. Каждая такая клетка воздействует на раковину химическими веществами, после чего отделяет от неё маленький кусочек. Образовавшийся «осколок» транспортируется в водоносную систему и выводится наружу через оскулюм. Скрытая внутри раковины или скалы губка надёжно защищена от хищников, поэтому такой способ существования может повышать вероятность выживания как молодых, так и взрослых губок.
Бесполое размножение губок осуществляется путём фрагментации, почкования или образования зимующих пропагул (геммул). Фрагментация, как правило, происходит в результате повреждения губки течениями, прибоем или хищниками. Оторвавшиеся куски выживают благодаря способности губки к регенерации. Фрагмент прикрепляется к субстрату и, перестраиваясь, превращается в нормально функционирующую губку. Крайнее проявление фрагментации — диссоциация губки на отдельные клетки или группы клеток. Это может быть достигнуто путём протирания кусочка губки через мелкоячеистую ткань. Подобный эксперимент был впервые поставлен зоологом Г. В. Уилсоном в начале XX в. и с тех пор часто воспроизводился для изучения механизмов клеточного распознавания и моделирования морфогенеза.
Почкование у губок встречается относительно редко, но всё же имеет место у небольшого числа видов. Например, есть данные, что у Clathrina, имеющей асконоидный тип строения, свободные концы трубок вздуваются и образуют почки, которые затем отделяются, прикрепляются к субстрату и дают начало новой губке. У некоторых видов Tethya почки образуются на особых стебельках. Губки, относящиеся к родам Oscarella и Aplysilla, по некоторым данным, формируют папиллы, затем папиллы отделяются и дают начало новым организмам.
Многие пресноводные и некоторые морские губки образуют сотни, а то и тысячи напоминающих споры геммул. Как правило, это происходит осенью. Осенние геммулы пресноводных видов могут входить в состояние диапаузы, при котором практически полностью подавляется их метаболическая активность. Для активизации таких геммул необходимо, чтобы они в течение определённого периода подвергались воздействию очень низкой температуры. Затем геммулы прорастают и дают начало новым губкам. Обычно это происходит весной. Пока геммула находится в диапаузе, она устойчива к экстремальным воздействиям окружающей среды, таким, как очень низкая температура, повышенная солёность и высыхание.
Геммулы образуются в мезохиле погибающей губки вокруг группы богатых питательными веществами археоцитов. Спонгоциты формируют спонгиновую оболочку вокруг этой клеточной массы. Оболочка может быть инкрустирована спикулами, которые образуются в склероцитах. Она полностью покрывает клеточную массу; лишь на одном полюсе остаётся небольшое отверстие — микропиле. Сформированная геммула состоит из оболочки и находящихся внутри археоцитов, каждый из которых вскоре округляется и становится похож на эмбриональную клетку. Такие модифицированные археоциты называются тезоцитами.
Весной геммулы «прорастают»: периферические тезоциты дифференцируются в пинакодерму, которая, быстро разрастаясь, выступает наружу через микропиле, как надувающийся пузырь. Этот «пузырь» пинакодермы соприкасается с субстратом и прикрепляется к нему. Затем через микропиле внутрь пузыря перемещаются остальные тезоциты, которые составляли центральную часть геммулы. После дифференциации они образуют внутренние структуры молодой губки.
Наблюдения за развитием молодых губок из геммул позволили получить данные, ставящие под сомнение возможность использования по отношению к губкам понятия «индивидуальность». В процессе прорастания тезоциты из разных геммул одной и той же губки или из геммул, принадлежащих разным экземплярам, но обязательно одного вида, могут объединяться. В этом случае из них развивается одна новая губка, обладающая своей «индивидуальностью».
Губки, за редким исключением, — гермафродиты. Когда приходит время, одна губка выбрасывает наружу сперматозоиды, которые переносятся токами воды к другой губке, внутри которой происходит оплодотворение яиц. У яйцекладущих видов (например, Cliona) зиготы выводятся во внешнюю среду, где и протекает их развитие. Многим губкам свойственно живорождение: зиготы остаются внутри материнского организма, который покидают уже сформированные личинки (иногда такие формы называют «личинкородящими»). Зародыши и личинки губок лецитотрофны.
Считается, что у губок нет половых органов (гонад). Половые клетки залегают в мезохиле либо небольшими группами (сперматозоиды), либо поодиночке (яйца). Сперматозоиды образуются из отдельных хоаноцитов или целых жгутиковых камер, которые погружаются в мезохил и окружаются тонкой клеточной стенкой, давая начало сперматоцисте. Яйца образуются из археоцитов или, как у некоторых известковых губок, — из дедифференцированных хоаноцитов. Обычно яйцо накапливает желток, фагоцитируя окружающие их питающие клетки, или клетки-«няньки». Яйцо и клетки-«няньки» иногда заключены в особый фолликул, имеющий клеточные стенки. (По определению, орган состоит из двух и более тканей. Если будет доказано, что клетки стенки цисты или фолликула и половые клетки имеют разное происхождение и формируются из разных тканей, то можно будет сперматоцисты и яйцевые фолликулы считать органами — гонадами.) Поскольку водоносная система одинаково снабжает все части тела, половые клетки довольно равномерно распределены по мезохилу, но при этом всегда находятся недалеко от канала или камеры, в пределах расстояния, на котором действует диффузия.
В период размножения стенка сперматоцисты разрушается, сперматозоиды попадают в отводящие каналы (или в атриум) и выбрасываются наружу через оскулюмы. Некоторые тропические виды внезапно извергают из оскулюмов молочные облачка спермы. Возможно, одновременное высвобождение большого количества сперматозоидов характерно для большинства губок.
Когда выброшенные сперматозоиды оказываются рядом с другой губкой, токи воды заносят их в её водоносную систему. Внутри губки сперматозоиды доставляются к поверхности хоанодермы или в жгутиковые камеры, где их фагоцитируют (но не переваривают) хоаноциты. Хоаноцит, захвативший сперматозоид, теряет жгутик и воротничок и трансформируется в амебоидную клетку, которая называется клеткой-носительницей. Она переносит головку сперматозоида (яйцо) к яйцу. Когда клетка-носительница достигает ближайшего расположенного в мезохиле яйца, она либо «передаёт» ему ядро спермия, либо яйцо фагоцитирует эту клетку вместе с содержащимся в ней ядром спермия. В любом случае оплодотворение происходит внутри «яичника» губки.
Как правило, сперматозоиды губок лишены акросомы — структуры, которая у большинства других животных отвечает за проникновение спермия через мембрану яйцеклетки при оплодотворении. Вероятно акросома не нужна в тех случаях, когда ядро спермия попадает в яйцо путём фагоцитоза. У губки Oscarella lobularis спермии всё же имеют акросому, что может свидетельствовать о существовании у этого вида обычного способа оплодотворения яйцеклетки.
Зигота претерпевает полное и равномерное дробление. Однако взаимное расположение образующихся бластомеров варьирует у разных видов губок. Разнообразны и развивающиеся из зародыша личинки. Они носят названия целобластула, амфибластула, паренхимула и трихимелла.
Личинка целобластула характерна для известковых губок, например видов рода Clathrina (Calcinea). Целобластула представляет собой полый шар, стенка которого состоит из одного слоя жгутиковых клеток. Пока личинка ведёт планктонный образ жизни, некоторые её клетки теряют жгутики, становятся амебоидными и мигрируют в бластоцель, так что он, в конце концов, полностью заполняется клетками. Полая целобластула превращается в лишённую полости стерробластулу.
Личинка амфибластула встречается у других известковых губок, например Grantia, Sycon и Leucosolenia (Calcaronea). Амфибластула представляет собой полый шар, состоящий из клеток двух типов: жгутиковых клеток на переднем полюсе личинки и безжгутиковых клеток с зернистой цитоплазмой на заднем полюсе. Когда личинка ещё находится в мезохиле материнского организма, все жгутики обращены в бластоцель, однако между «зернистыми» клетками на заднем полюсе личинки вскоре образуется отверстие, через которое она выворачивается наизнанку. После инверсии жгутики оказываются на наружной поверхности личинки, которая таким образом приобретает способность плавать с их помощью. На этой стадии личинка покидает материнский организм. Инверсия имеет место у тех губок, яйца которых образуются из хоаноцитов: после оплодотворения клетки делятся так, словно они должны образовать новые жгутиковые камеры, внутрь которых направлены жгутики. Обыкновенные губки Oscarella и Plakina тоже имеют в развитии личинку типа амфибластулы, но она образуется вторично, уже после стадии паренхимы. В своём развитии они проходят стадию плотной морулы, которая позднее в результате выселения клеток из центральной части в наружный клеточный слой (мультиполярная эмиграция) превращается в личинку особого типа — цинктобластулу, которая по своему строению принципиально отличается от типичной амфибластулы, свойственной известковым губкам.
Личинка паренхимула характерна для большинства обыкновенных губок. В этом случае зародыш развивается непосредственно в стерробластулу — клеточную массу без внутренней полости. Наружный слой в основном состоит из жгутиковых клеток, между которыми изредка попадаются клетки, лишённые жгутиков и содержащие в цитоплазме множество везикул. Внутри личинки находятся дифференцированные клетки многих типов: склероциты, колленциты, пинакоциты, даже жгутиковые камеры, а также археоциты. Таким образом, паренхимулу в какой-то степени можно считать «почти готовой» молодой губкой, специализированной для плавания.
Личинка трихимелла — типичная стадия развития стеклянных губок. Трихимелла представляет собой стерробластулу с пояском жгутиковых клеток по экватору. Внутри она заполнена клетками, содержащими желток, склероцитами (со спикулами), клетками других типов и даже жгутиковыми камерами.
Все личинки губок лецитотрофны и, следовательно, живут относительно недолго. Как правило, выход личинок стимулируется светом и обычно происходит на рассвете. По истечении определённого периода, продолжительность которого варьирует от нескольких часов до нескольких дней, личинки оседают и начинают ползать по дну в поисках походящего места для прикрепления. Обнаружив такое место, личинка прикрепляется, претерпевает метаморфоз и превращается в молодую губку. Её строение несколько отличается у разных губок и определяется типом личинки. Поскольку метаморфоз сопровождается перегруппировкой клеток с образованием более или менее чётко выраженных пластов, его часто сравнивают с гаструляцией у других многоклеточных. Однако и погружение клеток в полость личинки, что имеет место у многих губок при формировании стерробластул, также можно считать своего рода гаструляцией.
Попытки найти гаструляцию у губок сопровождают практически всю историю изучения этих организмов. В своё время даже выдвигалась идея об «извращении положения зародышевых листков» у губок. Считалось, что поверхностно расположенные жгутиковые клетки личинок представляют собой эктодерму, которая позднее погружается внутрь тела и даёт начало хоанодерме, а крупные безжгутиковые клетки — это энтодерма, которая остаётся снаружи и превращается в пинакодерму. Отсутствие у губок жёстко детерминированных тканевых дифференцировок позволяет некоторым исследователям говорить о том, что у этих животных не может быть и настоящей гаструляции, в процессе которой осуществляется становление двух первых зародышевых листков — экто- и энтодермы. Процессы обособления нескольких клеточных совокупностей, сопровождающие эмбриональное развитие губок, чисто внешне могут напоминать разные типы гаструляции настоящих многоклеточных животных, но не гомологичны им, точно так же, как не гомологичны ткани и органы Eumetazoa пинакодерме, хоанодерме и мезохилу губок.
Непосредственно перед метаморфозом целобластулы, которая к этому моменту уже превратилась в стерробластулу, её клетки дифференцируются и образуют сплошную массу тотипотентных клеток. После прикрепления клеточная масса распластывается по субстрату. Клетки, расположенные на её поверхности, дают начало пинакодерме, а залегающие более глубоко дифференцируются и дают начало всем остальным типам клеток, характерным для губок. В процессе перераспределения внутренних клеток щелевидные просветы между ними постепенно объединяются, образуя атриум.
Амфибластула оседает на субстрат и прикрепляется к нему передним, несущим жгутики концом. После прикрепления жгутиковые клетки теряют жгутики, мигрируют внутрь и образуют внутренние части губки. «Зернистые» клетки дают начало пинакодерме. Когда молодой организм начинает активно функционировать и питаться, он представляет собой миниатюрную асконоидную трубку. Эта стадия называется олинтус.
Метаморфоз личинок, относимых к парнехимулам, протекает немного по-разному у разных видов. Обычно после прикрепления личинки её внутренние клетки дифференцируются и перераспределяются. Из них образуется большая часть тела губки, если не вся она целиком.
У вида Mycale contarenii жгутиковые клетки превращаются в хоаноциты. У других видов (например, у некоторых пресноводных губок, а также у Microciona prolifera) жгутиковые клетки фагоцитируются археоцитами и соответсвенно не входят в число клеток, принимающих участие в формировании тела молодой губки. В любом случае, прежде чем приобрести лейконоидную организацию, молодая, только что претерпевшая метаморфоз губка часто характеризуется аксоноидным или сиконоидным строением, но имеет при этом толстые стенки. Ювенильные губки, находящиеся на этой стадии развития носят название рагон.
В умеренных широтах губки живут от одного года до нескольких лет. Представители многих тропических видов (а также, возможно, глубоководные губки) могут жить очень долго, до 200 лет и более. Некоторые губки начинают размножаться половым путём только по достижении ими возраста в несколько лет, в то время как другие приступают к размножению в возрасте 2 или 3 недель. Некоторые коралловые губки (Sclerospongia) растут очень медленно, давая прирост около 0,2 мм в год. Если эта скорость роста постоянна, то возраст коралловых губок диаметром 1 м может достигать 5000 лет.
Шестилучевая губка Scolymastra joubini живёт до 10 тысяч лет, это максимальная величина в животном мире.
Первые свидетельства о существовании Hexactinellida относятся к протерозою. В позднем протерозое обнаружены и остатки обыкновенных губок, в том числе и «роговых». Известковых губок и Homoscleromorpha пока удаётся обнаруживать только начиная с палеозоя (кембрий).
Ископаемые «группы» Sphinctozoa и Stromatoporata в настоящее время не рассматриваются как самостоятельные таксоны. По мнению ряда исследователей, это не более чем своего рода морфологические типы, отражающие разные варианты организации тела губок. Это в полной мере относится и к Sclerospongae. Вымершие Archaeocyatha, которых и сейчас рассматривают как самостоятельный класс, по-видимому, ближе всего стояли к обыкновенным губкам. Представители таксона Archaeocyatha имели пористый известковый скелет в форме перевёрнутого полого конуса с радиальными септами и двойной стенкой. Представители Stromatoporata напоминали современных коралловых губок (Sclerospongiae). Они тоже обладали массивным базальным известковым скелетом с внутренними трубками, но в отличие от Sclerospongiae у них не было кремнезёмных спикул. Губки, входящие в группу Sphinctozoa (в основном от ордовика до триаса), имели пористый известковый скелет с кольчатыми перетяжками, напоминающий нитку бус. Изучение современной реликтовой сфинктозойной губки Vaceletia crypta, относящейся к этому же таксону, позволяет сделать вывод о том, что мягкие ткани вымерших Sphinctozoa находились внутри скелета. Черты сходства в строении тканей Vaceletia crypta и обыкновенных губок дают основание относить эту губку (а возможно, и некоторых других Sphinctozoa) к Demospongiae.
Представители Archaeocyatha, Sphinctozoa, Stromatoporata, а также древние Demospongiae в кембрийских и мезозойских морях играли большую роль в образовании рифов. Это заставляет некоторых спонгиологов полагать, что основными тенденциями в эволюции губок были редукция присущих этим древним формам массивных скелетов, из которых формировались рифы, и замена их скелетами из спикул. Низкая скорость роста ныне живущих губок с массивными скелетами по сравнению со скоростью роста кораллоа могла быть причиной эволюционного заката рифообразующих губок. Этой гипотезе противоречит тот факт, что хоанофлагелляты, возможно, имеющие с губками общего предка, строят свой скелет из спикул, состоящих из двуокиси кремния.
Систематика губок весьма противоречива даже в тех случаях, когда ресь идёт о таксонах самого высокого уровня. В последнее время коралловых губок (ранее самостоятельный таксон Sclerospongiae) включают в группу Demospongiae. Недавно было также предложено выделить два подтаксона — Symplasma (Hexactinellida) и Cellularia, поскольку это формально отражает существующую разницу между синцитиальной организацией Hexactinellida и клеточным строением Calcarea и Demospongiae.
wreferat.baza-referat.ru
Известно около 5000 видов этого типа, из которых лишь небольшое количество обитает в пресных водах, а остальные — морские животные.
Наиболее примитивные представители типа — одиночные губки — имеют бокаловидую форму. Стенки их тела состоят из двух слоев клеток — наружного и внутреннего, разделенных студневидной мезоглеей. В состав наружного слоя входят в основном эпителиальные клетки, выполняющие роль защитного покрова. Во внутреннем слое имеются очень характерные для губок воротничковые клетки, или хоаноциты, названные так потому, что их передний край вытянут в прозрачный воротничок, окружающий основания жгутика). В мезоглее находятся неспециализированные клетки. Соединение клеток в теле губок, рыхлое и они могут до известной степени менять свое положение.
Стенки тела губок пронизаны множеством мелких пор, которые ведут в полость, окруженную внутренним слоем клеток и открывающуюся наружу более крупным отверстием оскулюмом. Жгутики воротничковых клеток создают ток воды, которая входит через поры в упомянутую полость и выходит наружу через оскулюм. Эту полость не следует считать кишечной, она служит только для прохождения воды и в отличие от настоящей кишечной, или гастральной, полости называется парагастральной. В клетках мезоглеи развиваются скелетные образования разного химического состава: известковые, кремневые, спонгиновые (спонгин — вещество, близкое по химическому строению к шелку). Благодаря скелету губки могут расти вверх, а не растекаться по субстрату, что облегчает движение воды через их тело.
Губки медленно и слабо отвечают на разные раздражения, так как в их теле нет нервных клеток. В теле губок нет мышечных клеток, за исключением тех, которые иногда развиваются вокруг оскулюмов. Поэтому тело губок почти не сократимо, а движение воды через их тело осуществляется, как было сказано выше, только благодаря биению жгутиков воротничковых клеток. Кроме того, некоторые клетки могут передвигаться внутри тела, как амебы, т. е. выпуская ложноножки.
Перенос веществ обеспечивается, во-первых, постоянным движением воды через парагастральную полость, доставляющей внутреннему слою кислород и уносящей продукты диссимиляции, во-вторых, диффузным способом (проникновение переваренной пищи, кислорода и других веществ от слоя к слою).
Получение кислорода совершается через внутреннюю и наружную поверхности тела.
Выделение продуктов диссимиляции тоже происходит через поверхность тела. Кроме того, эти продукты могут скапливаться в некоторых клетках (главным образом в мезоглее), переходя из растворимого состояния в нерастворимое и поэтому переставая оказывать вредное влияние на организм.
Пища в виде мелких организмов или их остатков попадает с водой в парагастральную полость и захватывается воротничковыми клетками, которые способны выпускать ложноножки. Захватывать пищу могут и некоторые клетки наружного слоя, а также мезоглеи. Пищеварение, как и у простейших, внутриклеточное. Разложенные в результате пищеварения вещества частично диффундируют в другие клетки и там усваиваются, а частично ассимилируются на месте.
У губок очень распространено бесполое размножение, которое совершается путем разных видов почкования. Образование колоний происходит в результате того, что почки остаются на материнском организме. Кроме того, образуются особые почки, которые выпадают из материнского организма после его гибели и дают начало новым организмам. Губки размножаются и половым путем. Большинство представителей этого типа — гермафродиты, меньшинство — раздельнополы. Половые клетки развиваются из недифференцированных клеток мезоглеи, а микрогаметы, по новым данным, могут образоваться также из воротничковых клеток. Сперматозоиды выходят в воду и проникают в тело других губок, где оплодотворяют яйцеклетки.
Развитие зиготы происходит через стадии — морула, бластула, паренхимула и у морских губок заканчивается образованием планктонных личинок, плавающих при помощи жгутиков наружных клеток. Личинки, плавая в воде и переносимые течениями, способствуют расселению этих сидячих животных. У пресноводных губок планктонных личинок нет и они распространяются другими способами. При превращении личинок во взрослые сидячие формы у губок происходит извращение зародышевых слоев: наружные жгутиковые клетки мигрируют внутрь, а клетки внутреннего слоя перемещаются наружу. Благодаря такому процессу образуется слой воротничковых клеток, окружающих парагастральную полость, создающий столь важный для описываемых животных ток воды, приносящий пищу и кислород и удаляющий продукты распада обмена веществ.
www.ronl.ru





