Казахский Государственный Университет
им. Аль-Фараби
Географический факультет
Механизмы саморегуляции численности популяции .
Выполнила: Першикова О.
Проверила: Аскарова М.А.
Алматы, 1999
План
1. Понятие популяции.
1.1. Экотипы.
1.2. Эдафотипы.
2. Плотность популяции.
2.1. Механизмы саморегуляции.
Популяция(от. Лат. Populus – народ, население) – это исторически сложившаяся совокупность особей одного вида, более или менее длительно время занимающая определенное пространство и воспроизводящая себя в течение большого числа поколений. Популяция является объектом изучения одновременно экологии и биологии.
Биологический вид обычно представлен множеством популяций, находящихся в той или иной форме изоляции, которая ограничивает или нарушает свободное скрещивание особей (панмиксия). Популяция занимает ту часть ареала, в которой отсутствуют изоляционные барьеры, препятствующие панмиксии. В результате естественного отбора, происходившего на протяжении тысяч поколений, каждая популяция приспосабливалась(адаптировалась) к среде своего обитания со свойственной ей совокупностью природных условий.
Характерные черты этого приспособления передаются из поколения в поколение. Популяции, адаптированные к определенным экологическим условиям окружающей среды, называются экотипами.
Экотипы, населяющие территории физико-географического ранга, т.е. со сравнительно однородным комплексом природных условий, являются географическими расами. В их выделения положено физико-географическое районирование суши и акваторий.
Морфологические, физиографические и другие приспособления особей этих географических популяций к единому ритму жизненных явлений и особенностям среды отличают их от особей соседних популяций, находящихся в иных природных условиях. Синонимом географической расы часто выступает подвид.
Популяция в пространстве не всегда экологически и географически изолирована, и ареалы подвидов могут перекрываться.
В приделах географических экотипов могут быть выделены климатические, или климатипы. Климат формирует совокупность особей с наследственно закрепленными особенностями вида, возникшими г.о. под влиянием климатических условий.
В приделах климатических экотипов могут быть выделены почвенные (эдофические) экотипы – эдафотипы. Они образуются под влиянием почвенных условий. Для сосны обыкновенной выделены и описаны эдафотипы сосны меловой, растущей на меловых отложениях, и сосны болотной, характерной для моховых болот.
В рамках определенного биоценоза совокупность особей, как правило, одного вида растений образует ценопопуляцию, т.е. популяцию, приуроченную только к этому биоценозу. Кроме того, в экосистеме находятся также полиценотические
(приуроченные к нескольким биоценозам) популяции, например, свободно передвигающихся животных. Характеризуя экотип как единое целое, экология рассматривает особей вида, его образующих, как статистические единицы, подлежащие учету и анализу пространственного размещения и взаимодействия. К экологическим особенностям популяции относятся: её место обитания и экологическая ниша, численность и плотность особей, их пространственное размещение и расселение, а также отношение между особями в популяции и между популяциями в биоценозе. Экологическая ниша представляет собой положение популяции в сообществе, включая участие в трофических цепях.
Внутрипопуляционная регуляция численности популяции.
Плотность популяции обычно имеет определенный оптимум. При любом отклонении от этого оптимума начинают срабатывать механизмы ее внутрипопуляционной регуляции. Одним из основных механизмов, способствующих установлению в популяции устойчивой стабильности, служит действие зависимых от плотности факторов. Абиотические факторы так же влияют на смертность популяции, но самостоятельно не создают ей устойчивой стабильности.
Регуляция численности популяции у различных видов животных и растений осуществляется по-разному. Тем не менее, в каждой из них определенным путем устанавливается оптимум плотности.
Рост плотности популяции сопровождается уменьшением размеров особей, снижением их плодовитости, повышением смертности личинок и куколок изменением скорости развития и соотношения полов, а также увеличением количества диапаузирующих особей, что резко снижает активную часть популяции.
Нередко при чрезмерном возрастании плотности популяции стимулируется каннибализм. Ярким примером может служить явления поедания своих же яиц мучными хрущаками. Каннибализм наблюдается у некоторых видов рыб, у земноводных и других животных.
Одним из важнейших механизмов внутрипопуляционной регуляции численности выступает эмиграция, интенсивность которой стимулируется повышением плотности популяции. Это довольно типично для многих насекомых, у которых при определенной величены плотности популяции отмечается выселение части особей, иногда значительной, в менее предпочитаемые ими места обитания того же ареала. У некоторых видов тлей повышение плотности популяции сопровождается появлением крылатых особей, способных расселяться. При переуплотнении популяции эмиграции происходят у ряда млекопитающих (особенно у мышевидных грызунов и птиц) и птиц.
Достаточно изучена регулирующая роль внутривидовой конкуренции за ограниченные ресурсы. У падальных мух из огромного количества откладываемых на труп яиц выходит так много личинок, что пищи на всех не хватает. В результате катастрофически растет смертность в ранних возрастах. Сходное явление обнаружено у короедов (Scolytidae), муравьев-лазиусов (Lasius), у некоторых стрекоз (Odonata) и других насекомых.
В наиболее простых случаях внутрипопуляционные регуляторные механизмы численности проявляются в виде непосредственной конкуренции за необходимые для жизни ресурсы, количество которых недостаточно для удовлетворения потребностей всех особей. Известно, что плотность популяции яблонной плодожорки и капустной моли регулируется конкуренцией за пищу и места для окукливания. Внутривидовая конкуренция у некоторых мух в случае возрастания плотности популяции до определенного уровня приводит к падению массы куколок, что сопровождается повышенной смертностью.
Важной является проблема «минимальной жизнеспособности популяции», суть которой состоит в определении минимальной численности популяции, которая гарантировала бы ее существование в течение какого-то длительного периода. В тоже время падение плотности популяции ниже уровня оптимума, например при усиленном истреблении крыс, вызывает повышение плодовитости и стимулирует их более раннее половое созревание.
Некоторые механизмы регуляции численности популяции одновременно могут выступать и как механизмы, предотвращающие внутривидовую конкуренцию. Так, если птица отмечает свой гнездовой участок пением, то другая пара этого же вида гнездится за приделами этого участка. Метки оставляемые многими видами млекопитающими, ограничивают их охотничий участок и предупреждают вселение других особей. Все это снижает внутривидовую конкуренцию и препятствует чрезмерному уплотнению популяции.
Изменение плотности популяции может иметь рефлекторное, или сигнальное действие. В случаи высокой плотности такая сигнализация регулярным путем приводит к сокращению численности популяции, а при малой плотности — ее усиленному размножению и росту. Как было установлено С.С. Шварцем, у земноводных и крыс такая сигнализация имеет химическую природу. Приведем пример, подтверждающий этот вывод. Если воздух из клетки с переуплотненной популяции крыс подавать в клетку с разреженной популяцией, физиология особей последней изменяется в направлении, характерном для популяции с повышенной плотностью.
У растений регуляторными механизмами численности популяций служит, прежде всего, внутривидовая конкуренция. Она обычно связана с повышенной густотой произрастания. В переуплотненных посевах, например, происходит уменьшение количества семенной продукции, что имеет большое значение для сельского и лесного хозяйства. Чаще всего растения одного вида конкурируют за свет и влагу. В густых посевах они затеняют друг друга, при ограниченном количестве воды испытывают ее недостаток. В результате часть их погибает. Такое явление наиболее характерно для многих огородных культур и лесных растений. В лесу всегда значительно больше молодых растений, чем старых. Внутривидовой конкуренцией за влагу объясняется не редко встречающаяся правильное распределение пустынных растений. Создается впечатление, словно их кто-то рассадил на строго определенном расстоянии друг от друга. В понижениях местности, в оазисах это равномерная разреженность популяции растений сразу же исчезает. Подобным же образом распределяется в африканских саваннах светолюбивые и относительно влаголюбивые баобабы.
Однако следует учитывать, что популяция обычно входит в состав сообщества и что устойчивое существование биоценозов возможно только при определенных количественных соотношениях всех компонентов. Этим и вызвано необходимость регуляции численности, обеспечивающей устойчивое состояние, как отдельных популяций, так и биоценозов в целом. Механизмы регуляции численности популяций, основаны на сложных межвидовых взаимоотношениях.
Список использованной литературы.
1. Радкевич В.А. “Экология”. Минск-1997, стр. 81.
2. Киселев В.Н. “Основы экологии “. Минск-1998, стр.256.
www.ronl.ru
УДК 582. 734(47. 53)К ИЗУЧЕНИЮ ЭКОТИПОВ Fragaria vesca L. В ПЕРМСКОМ КРАЕТ. П. Ларькина, канд. биол. наук,ФГБОУ ВПО Пермская ГСХА,ул. Петропавловская, 23, г. Пермь, Россия, 614 990,E-mail: vlarkin2@gmail. comАннотация. Земляника лесная на территории Пермского края представлена тремя экотипами: лесным, луговым и скальным. Под экотипом мы подразумеваем совокупность близкородственных ценопопуляций одного вида, сложившихся под воздействием определенных экологических факторов и естественного отбора- способных к самовоспроизведению не только при постоянстве условий существования, но и длительно сохраняющих свои морфологоанатомические особенности при перенесении в иную среду. Лесной и луговой экотипы являются ценотическими, связанными с условиями определенных растительных сообществ.Скальный ксерофитный экотип следует классифицировать как эдафический экотип, поскольку ведущая роль в его формировании принадлежит почвенному субстрату.Лесной экотип произрастает под пологом осветленных лиственных, смешанных и хвойных лесов. Растения высокие, плоды крупные, листья имеют типичное мезоморфное строение.Луговой экотип земляники лесной приурочен к обширным лесным полянам, опушкам, лугам и открытым пологим склонам. Размеры листьев, плодов у растений этого экотипа мельче, чем у представителей лесного экотипа. Анатомическое строение мезоморфное, но иногда (на южных склонах и опушках) проявляются признаки ксероморфизма.Скальный ксерофитный экотип земляники лесной встречается на южных склонах с выходами известняка. Его представители низкорослые, плоды в два раза мельче, чем у растений лесного экотипа. Листья мелкие, прижатые к земле, густоопушенные, их анатомическое строение ксероморфное. В легкодоступных для населения местообитаниях состояние ценопопуляций земляники лесной постоянно ухудшается, что особенно характерно для ценопопуляций, представляющих луговой экотип. Для ценопопуляций лесного экотипа, приуроченных к сосновым борам, губительны низовые лесные пожары, часто возникающие по вине населения. На горель-никах ценопопуляции земляники лесной не восстанавливаются десятилетиями, что связано с биологичнскими особенностями данного вида.Ключевые слова: земляника лесная, ценопопуляция, ценотический экотип, эдафический экотип, мезоморфизм, ксероморфизм.Введение. Род Fragaria на территории Пермского края представлен тремя видами: земляника лесная (Fragaria vesca L.), земляника зеленая (Fragaria viridis Duch. j и земляника мускусная (Fragaria moschata Duch.) [1]. Наиболее хорошо изучена земляника лесная, поскольку она издавна известна во всех частях ее ареала как ценное пищевое и лекарственное растение научной и народной медицины [2,3,4,5].Границы ареала земляники лесной на севере и юге выходят за пределы Пермского края, проходя, соответственно, через 61 градус и 53 градуса СШ. Южнее она встречается очень редко, удерживаясь под пологом леса и на северных склонах. На севере Пермского края для земляники лесной более благоприятны склоны южной экспозиции, на склонах северной экспозиции она практически не плодоносит. В центральных и южных районах она распространена под пологом осветленных лиственных, хвойных и смешанных лесов, на луговых и лесных полянах, опушках и вырубках, где в отдалении от крупных населенных пунктов она хорошо растет и обильно плодоносит.В Пермском крае произрастает три экотипа земляники лесной: лесной (мезофитный), луговой (мезофитный) и скальный (ксерофит-ный). Под экотипами (экологическими типами земляники лесной) подразумеваются совокупности близкородственных ценопопуляций, сложившихся под воздействием определенных экологических факторов и естественного отбора- способных к самовоспроизведению не только при постоянстве условий существования, но и длительно сохраняющих свои наследственные особенности при перенесении в иную среду (6). Внутривидовая дифференциация морфологических, фенологических, физиологических особенностей земляники лесной, связанная с условиями ее произрастания, отмечена и другими исследователями [7,8].Цель и задачи исследований. Цель исследования — выявить основные тенденции состояния ценопопуляций земляники лесной на территории Пермского края. Задачами исследований явилось изучение основных морфологических параметров растений и фитоце-нотических характеристик ценопопуляций, принадлежащих к разным экотипам.Материалы и методы исследований.Мониторинг трех ценопопуляций земляники лесной, принадлежащих к разным экотипам, на территории Пермского края начат в 1990 году, первые итоги подведены в 2010 году. Работа велась на трех площадках: 1) ценопо-пуляция под пологом осветленного березового леса (Краснокамский район), площадь 180 квадратных метров- 2) ценопопуляция на юговосточной опушке того же леса площадью 160 квадратных метров 3) ценопопуляция на южном склоне с выходами известняка площадью 90 квадратных метров в Кунгурском районе. Описание фитоценозов, изучение плотности и состава ценопопуляций, биометрический анализ растений и математическая обработка полученных данных проводились по общепринятым методикам. Биометрический анализ растений делали на свежих и гербаризированных растениях, анатомические исследования -на фиксированных растениях. Коэффициент встречаемости устанавливался по 50 учетным площадкам [9,10,11].Результаты и их обсуждение. Лесной и луговой экотипы являются типичными цено-тическими, скальный ксерофитный следует отнести к категории эдафических экотипов [6]. Лесной экотип земляники лесной распространен под пологом всех типов осветленных лесов. Он представлен растениями с крупными листьями (средняя площадь тройчатого листа до 2700 квадратных миллиметров), плоды достигают веса 580 миллиграммов, высота цветоносов может превышать 25 сантиметров. Листья имеют типичное мезоморфное строение: число устьиц на одном квадратном миллиметре не превышает 200, палисадная паренхима всегда однослойная, отношение толщины палисадной паренхимы к губчатой 1:1.Луговой экотип земляники лесной приурочен к лесным опушкам, лугам, пологим открытым склонам. Его плоды и листья мельче, чем у представителей лесного экотипа: средняя площадь листьев не превышает 2000 квадратных миллиметров, а вес плодов не более 520 миллиграммов. Анатомическое строение листьев лугового экотипа тоже типично мезоморфное, но иногда проявляются признаки ксероморфизма: увеличение числа устьиц на единицу поверхности листа, большая общая мелкоклетность и плотность тканей, увеличение слоев клеток палисадной паренхимыдо двух, отношение толщины палисадной паренхимы к губчатой 1,2:1.Скальный экотип земляники лесной был обнаружен на территории Кунгурской лесостепи на склонах южной экспозиции с выходами известняка крутизной 40−60 градусов. Земляника лесная здесь имеет очень своеобразный фенотип с четко выраженной ксеро-морфностью строения: высота цветоноса не превышает три с половиной сантиметра, листья и плоды мелкие: средняя площадь листа не более 500 квадратных миллиметров, максимальный вес плодов 280−290 миллиграммов. Листья густо опушены волосками, количество устьиц на одном квадратном миллиметре достигает 500, палисадная паренхима двухслойная и даже трехслойная, отношение толщины палисадной паренхимы к губчатой 2:1. Фитоценоз с земляникой лесной в этом местообитании изобилует степными видами — ксерофитами: Veronica spicata L., Stipa pennata L., Aster alpinus L. и др.Изучение ценопопуляций земляники лесной трех экотипов за период с 1990 по 2009 годы (таблица) показал следующее: поднаибольшим пагубным антропогенным воздействием находится луговой экотип. Так, за 19 лет наблюдений плотность ценопопуляции на юго-восточной опушке березняка сократилась почти в три раза. Этот показатель на расположенной рядом луговой поляне пологого склона юго-восточной экспозиции уменьшился в 2,3 раза. Подобные местообитания — излюбленные места сбора земляники лесной. Здесь она имеет высокий коэффициент встречаемости (табл.), ее плоды при достаточнокрупных размерах созревают на 10−12 дней раньше, чем под пологом леса.Состояние ценопопуляции, представляющей лесной экотип, за период наблюдений почти не ухудшилось: среднее количество особей земляники лесной под пологом осветленного березняка сократилось только на 8%, а коэффициент встречаемости даже увеличился с 25 до 28. Площадь листьев, вес плодов тоже колебались незначительно (табл. 1), что предположительно связано с количеством выпадающих осадков. Известно [6,12], что цено-популяции земляники лесной страдают не столько от сбора плодов, сколько от вырывания растений с корневищем и усами в период массового цветения, так как в этот период она содержит максимальное количество биологически активных веществ. Заготовителей лекарственного сырья останавливает обилие ик-содовых клещей в лесах Пермского края, наибольшая активность которых по времени совпадает с периодом начала цветения земляники лесной.Лесной экотип широко распространен на территории Пермского края в осветленных сосновых борах. Здесь его ценопопуляции постоянно подстерегает летальный экологический фактор — низовые лесные пожары, возникающие по вине туристов и сборщиков грибов и ягод. Полученные ранее результаты совпадают с мнением других исследователей: низовые пожары губительны для земляники лесной в силу ее биологических особенностей, ценопопуляции не восстанавливаются полностью даже через десять лет [13, 14, 1].Состояние ценопопуляции земляники лесной в Пермском краеТаблица 1Ценопопуляция Под пологом разреженного березняка На юго-восточной опушке березняка На южном склоне с выходами известнякаГод 1990 2009 1990 2009 1990 2009Площадь ценопопуляции, кв.м. 180 200 130 100 90 100Количество особей на 1 кв.м., шт. 54,0± 1,9 47,5± 1,8 60,0± 1,9 23,1± 1,3 23,0±0,8 24,0± 0,7Количество цветущих особей от общего, % 12,0 12,6 19,0 16,5 19,8 24,0Площадь самого крупного листа кв. мм. 2751± 24 2704± 21 1920± 11 1806± 14 481± 5,0 486± 14Вес первого на растении плода, мг. 589± 17 570± 12 528± 16 501± 14 290± 9Коэффициент встречаемости з. лесной, %. 25 28 45 22 10 12Эдификатор F. vesca F. vesca F. vesca Alchemilla vulgaris F. vesca F. vescaНазвание ассоциации Злаково-бобово-разнотравная Злаково-разнотравная Разнотравно-злаковаяКоличество особей на одном квадратном экотипами: лесным, луговым и скальным.метре, процент цветущих растений от общего 2. Состояние популяций лугового экотипачисла, коэффициент встречаемости у ценопо- катастрофически ухудшается, особенно околопуляции скального ксерофитного экотипа за крупных населенных пунктов. период наблюдений практически не изменил- 3. В труднодоступных и непопулярныхся (табл.). Причины этого очевидны — трудно- местах сбора земляники лесной как лекар-доступность местообитаний, мелколистность ственного и пищевого растения, ее ценопопу-и мелкоплодность особей- такой низкопродук- ляции вполне стабильны.тивный объект сбора не интересен заготови- 4. Необходимо продолжить мониторингтелям ни в качестве пищевого растения, ни в ценопопуляций земляники лесной и заложитькачестве лекарственного сырья. новые учетные площадки в целях поиска вВыводы. 1. На территории Пермского природе новых экотипов для более полного края земляника лесная представлена тремя познания природы данного вида.Литература1. Овеснов С. А. Конспект флоры Пермской области. Пермь: Изд-во Пермского университета, 1997. 252с.2. Гаммерман А. Ф., Кадаев Г. Н., Яценко-Хмелевский А. А. Лекарственные растения. М.: Высшая школа, 1983.399 с.3. Мацку Я., Крейча И. Атлас лекарственных растений. Братислава: Словацкая Академия Наук, 1989. 464 с.4. Чиков П. С. Лекарственные растения. М.: Лесная промышленность, 1982. 383 с.5. Zorika, Popovic Wild flora and its usage in traditional phytotherapy (Deliblato Sands, Serbia, South East Europe)// Indian Journal of Traditional Knowledge Vol. 13(1). January 2014. p 9−35.6. Ларькина Т. П., Ковязин Н. Я., Жебелева Т. И.: Земляника лесная в Среднем Предуралье // Растительные ресурсы. 1982. т. 18. вып. 1. С. 27−29.7. Olga M. Heide and Anita Sonsteby Interactions of temperature and photoperiod in the control of flowering of latitudinal and altitudinal populations of wild strawberry (Fragaria vesca) // Physiologia Plantarum. 2007. Vol. 130. Р. 280−289.8. Erika Malinkova Altitudinal variation of plant traits: morphological characteristics in Fragaria vesca L. (Rosaceae) // Annals of Forest Research. 2013. Vol. 56. Issue 1. Р. 79−89.9. Зайцев Г. Н. Математическая статистика в экспериментальной ботанике. М.: Наука, 1984. 424 с.10. Прозина М. Н. Ботаническая микротехника. М.: Высшая школа, 1960. 206 с.11. Шенников, А. П. Введение в геоботанику. Л.: Изд-во ЛГУ, 1964. 447с.12. Ларькина Т. П. Состояние естественных зарослей земляники лесной в Пермском крае // Сб. науч. тр. ПГСХА (Пермский аграрный вестник LXVIII Всероссийской науч.- практическая конф. «Инновации и технологии — эффективному агропроизводству»). Пермь: ФГОУ ВПО «Пермская ГСХА», 2008. Ч. 1. С. 174−176.13. G. Geoff Wang and Kevin J. Kemball. Effects of fire severity on early development of understory vegetation. // Canadian Journal of Forest Research. 2005. Vol. 35(2). Р. 254−262.14. Francisco Moreira et al. Post-Fire Management and Restoration of Southern European Forests: Springer. 2011.329 p.15. Ларькина, Т. П. Влияние низового пожара на популяцию земляники лесной. // Материалы Всероссийской заоч. науч. -практической конф. (Актуальные проблемы аграрной науки в ХХ1 в). Пермь: ФГБОУ ВПО Пермская ГСХА, 2013. С. 28−30.ON THE INVESTIGATION OF ECOTYPES OF FRAGARIA VESCA IN PERMSKII KRAIT.P. Larkina, Cand. Bio. Sci., Assoc. Prof.Perm State Agricultural Academy 23 Petropavlovskaia st., Perm 614 990 Russia E-mail: v1a. rkin2@gmail. comABSTRACTFragaria vesca in the Permskii krai is represented by three ecotypes: forest, meadow and rock. Under ecotype we mean the set of closely cenopopulations of one species formed under the influence of certain environmental factors and natural selection- able to reproduce not only in constant condi-tions of existence, but also protractedly saving the morphological and anatomical features during transference in another environment. Forest and meadow ecotypes are cenotic type related to the terms of certain plant communities. Rocky xerophytic ecotype should be classified as edaphic ecotype, as the leading role in its formation belongs to the soil substrate. Forest ecotype grows under the canopy of bleached hardwood, mixed and coniferous forests. Plants are tall, large-fruit, the leaves are of typical mesomorphic structure. The meadow ecotype of Fragaria vesca grows at the extensive forest clearings, forest edges, open meadows and gentle slopes. Size of leaves, fruits, plants of this ecotype are smaller than those of the forest ecotype. It has mesomorphic anatomical structure, but sometimes (on the southern slopes and edges) is showing signs of xeromorphism. Rocky xerophytic ecotype is found on the southern slopes of the outputs of limestone. The samples of rocky ecotypes undersized fruit are twice smaller than in the forest ecotype plants. Leaves are small, pinned to the ground, with dense pubescence- their anatomical structure is xeromorphic. In easily accessible to the public habitats state, cenopopulations of strawberry timber is deteriorating, which is especially characteristic for ce-nopopulations representing meadow ecotype. The devastating forest fires, often caused by population, are fatal for the cenopopulation of forest ecotypes confined to pine forest. The cenopopulations of forest ecotype on burnt forest would not reverse during decades.Key words: Fragaria vesca, cenopopulation, cenotic ecotype, edaphic ecotype, mesomorphism, xeromorphism.References1. Ovesnov S.A. Konspekt flory Permskoi oblasti (Flora conspectus of Permskaia oblast), Perm: Perm university press, 1997, 252 p.2. Gammerman A.F., Kadaev G.N., Yatsenko-Khmelevskii A.A. Lekarstvennye rasteniya (medical plants), M.: Vysshaya shkola, 1983, 399 p.3. Matsku Ya., Kreicha I. Atlas lekarstvennykh rastenii (Atlas of medical plants), Bratislava: Slovatskaya Akademiya Nauk, 1989, 464 p.4. Chikov P. S. Lekarstvennye rasteniya (Medical plants), M.: Lesnaya promyshlennost'-, 1982, 383 p.5. Popovic Z. Wild flora and its usage in traditional phytotherapy (Deliblato Sands, Serbia, South East Europe), Indian Journal of Traditional Knowledge Vol. 13(1), January 2014, P. 9−35.6. Larkina T.P., Kovyazin N. Ya., Zhebeleva T.I. Zemlyanika lesnaya v Srednem Preduralie (Wood strawberry in the Middle Preduralie), Plant resources, 1982, Vol. 18, Issue 1, P. 27−29.7. Olga M. Heide and Anita Sonsteby Interactions of temperature and photoperiod in the control of flowering of latitudinal and altitudinal populations of wild strawberry (Fragaria vesca), Physiologia Plantarum, 2007, V ol. 130, P. 280−289.8. Erika Malinkova Altitudinal variation of plant traits: morphological characteristics in Fragaria vesca L. (Rosaceae), Annals of Forest Research, 2013, Vol. 56, Issue 1, P. 79−89.9. Zaitsev G.N. Matematicheskaia statistika v eksperimentalnoi botanike (Mathematical statistics in experimental botany), M.: Nauka, 1984, 424 p.10. Prozina M.N. Botanicheskaia mikrotekhnika (Botany microtechnigue), M.: Vysshaya shkola, 1960, 206 p.11. Shennikov, A.P. Vvedenie v geobotaniku (Introduction into geobotany), L.: Izd-vo LGU, 1964, 447 p.12. Larkina T.P. Sostoianie estestvennykh zaroslei zemlianiki lesnoi v Permskom krae (State of natural strawberry beds in Permskii krai), Permskii agrarnyi vestnik LXVIII Vserossiiskoi nauch.- prakticheskoi konf. «Innovatsii i tekhnologii -effektivnomu agroproizvodstvu»). Perm: FGOU VPO «Permskaya GSKhA», 2008, Part 1, P. 174−176.13. G. Geoff Wang and Kevin J. Kemball. Effects of fire severity on early development of understory vegetation, C anadian Journal of Forest Research, 2005, Vol. 35(2), P. 254−262.14. Francisco Moreira [et al.] Post-Fire Management and Restoration of Southern European Forests: Springer, 2011,329 p.15. Larkina, T.P. Vliianie nizovogo pozhara na populiatsiiu zemlianiki lesnoi (Influence of ground fire on wood strawberry population) Materialy Vserossiiskoi zaoch. nauch. -prakticheskoi konf. (Aktual'-nye problemy agrarnoi nauki), Perm: FGBOU VPO Permskaya GSKhA, 2013, P. 28−30.
Показать Свернутьsinp.com.ua
Казахский Государственный Университет
им. Аль-Фараби
Географический факультет
Механизмы саморегуляции численности популяции.
Выполнила: Першикова О.
Проверила: Аскарова М.А.
Алматы, 1999
План
1. Понятие популяции.
1.1. Экотипы.
1.2. Эдафотипы.
2. Плотность популяции.
2.1. Механизмы саморегуляции.Популяция (от. Лат. Populus – народ, население) – это исторически сложившаяся совокупность особей одного вида, более или менее длительно время занимающая определенное пространство и воспроизводящая себя в течение большого числа поколений. Популяция является объектом изучения одновременно экологии и биологии.
Биологический вид обычно представлен множеством популяций, находящихся в той или иной форме изоляции, которая ограничивает или нарушает свободное скрещивание особей (панмиксия). Популяция занимает ту часть ареала, в которой отсутствуют изоляционные барьеры, препятствующие панмиксии. В результате естественного отбора, происходившего на протяжении тысяч поколений, каждая популяция приспосабливалась (адаптировалась) к среде своего обитания со свойственной ей совокупностью природных условий.
Характерные черты этого приспособления передаются из поколения в поколение. Популяции, адаптированные к определенным экологическим условиям окружающей среды, называются экотипами.
Экотипы, населяющие территории физико-географического ранга, т.е. со сравнительно однородным комплексом природных условий, являются географическими расами. В их выделения положено физико-географическое районирование суши и акваторий.
Морфологические, физиографические и другие приспособления особей этих географических популяций к единому ритму жизненных явлений и особенностям среды отличают их от особей соседних популяций, находящихся в иных природных условиях. Синонимом географической расы часто выступает подвид.
Популяция в пространстве не всегда экологически и географически изолирована, и ареалы подвидов могут перекрываться.
В приделах географических экотипов могут быть выделены климатические, или климатипы. Климат формирует совокупность особей с наследственно закрепленными особенностями вида, возникшими г.о. под влиянием климатических условий.
В приделах климатических экотипов могут быть выделены почвенные (эдофические) экотипы – эдафотипы. Они образуются под влиянием почвенных условий. Для сосны обыкновенной выделены и описаны эдафотипы сосны меловой, растущей на меловых отложениях, и сосны болотной, характерной для моховых болот.
В рамках определенного биоценоза совокупность особей, как правило, одного вида растений образует ценопопуляцию, т.е. популяцию, приуроченную только к этому биоценозу. Кроме того, в экосистеме находятся также полиценотические
(приуроченные к нескольким биоценозам) популяции, например, свободно передвигающихся животных. Характеризуя экотип как единое целое, экология рассматривает особей вида, его образующих, как статистические единицы, подлежащие учету и анализу пространственного размещения и взаимодействия. К экологическим особенностям популяции относятся: её место обитания и экологическая ниша, численность и плотность особей, их пространственное размещение и расселение, а также отношение между особями в популяции и между популяциями в биоценозе. Экологическая ниша представляет собой положение популяции в сообществе, включая участие в трофических цепях.
Внутрипопуляционная регуляция численности популяции.
Плотность популяции обычно имеет определенный оптимум. При любом отклонении от этого оптимума начинают срабатывать механизмы ее внутрипопуляционной регуляции. Одним из основных механизмов, способствующих установлению в популяции устойчивой стабильности, служит действие зависимых от плотности факторов. Абиотические факторы так же влияют на смертность популяции, но самостоятельно не создают ей устойчивой стабильности.
Регуляция численности популяции у различных видов животных и растений осуществляется по-разному. Тем не менее, в каждой из них определенным путем устанавливается оптимум плотности.
Рост плотности популяции сопровождается уменьшением размеров особей, снижением их плодовитости, повышением смертности личинок и куколок изменением скорости развития и соотношения полов, а также увеличением количества диапаузирующих особей, что резко снижает активную часть популяции.
Нередко при чрезмерном возрастании плотности популяции стимулируется каннибализм. Ярким примером может служить явления поедания своих же яиц мучными хрущаками. Каннибализм наблюдается у некоторых видов рыб, у земноводных и других животных.
Одним из важнейших механизмов внутрипопуляционной регуляции численности выступает эмиграция, интенсивность которой стимулируется повышением плотности популяции. Это довольно типично для многих насекомых, у которых при определенной величены плотности популяции отмечается выселение части особей, иногда значительной, в менее предпочитаемые ими места обитания того же ареала. У некоторых видов тлей повышение плотности популяции сопровождается появлением крылатых особей, способных расселяться. При переуплотнении популяции эмиграции происходят у ряда млекопитающих (особенно у мышевидных грызунов и птиц) и птиц.
Достаточно изучена регулирующая роль внутривидовой конкуренции за ограниченные ресурсы. У падальных мух из огромного количества откладываемых на труп яиц выходит так много личинок, что пищи на всех не хватает. В результате катастрофически растет смертность в ранних возрастах. Сходное явление обнаружено у короедов (Scolytidae), муравьев-лазиусов (Lasius), у некоторых стрекоз (Odonata) и других насекомых.
В наиболее простых случаях внутрипопуляционные регуляторные механизмы численности проявляются в виде непосредственной конкуренции за необходимые для жизни ресурсы, количество которых недостаточно для удовлетворения потребностей всех особей. Известно, что плотность популяции яблонной плодожорки и капустной моли регулируется конкуренцией за пищу и места для окукливания. Внутривидовая конкуренция у некоторых мух в случае возрастания плотности популяции до определенного уровня приводит к падению массы куколок, что сопровождается повышенной смертностью.
Важной является проблема «минимальной жизнеспособности популяции», суть которой состоит в определении минимальной численности популяции, которая гарантировала бы ее существование в течение какого-то длительного периода. В тоже время падение плотности популяции ниже уровня оптимума, например при усиленном истреблении крыс, вызывает повышение плодовитости и стимулирует их более раннее половое созревание.
Некоторые механизмы регуляции численности популяции одновременно могут выступать и как механизмы, предотвращающие внутривидовую конкуренцию. Так, если птица отмечает свой гнездовой участок пением, то другая пара этого же вида гнездится за приделами этого участка. Метки оставляемые многими видами млекопитающими, ограничивают их охотничий участок и предупреждают вселение других особей. Все это снижает внутривидовую конкуренцию и препятствует чрезмерному уплотнению популяции.
Изменение плотности популяции может иметь рефлекторное, или сигнальное действие. В случаи высокой плотности такая сигнализация регулярным путем приводит к сокращению численности популяции, а при малой плотности - ее усиленному размножению и росту. Как было установлено С.С. Шварцем, у земноводных и крыс такая сигнализация имеет химическую природу. Приведем пример, подтверждающий этот вывод. Если воздух из клетки с переуплотненной популяции крыс подавать в клетку с разреженной популяцией, физиология особей последней изменяется в направлении, характерном для популяции с повышенной плотностью.
У растений регуляторными механизмами численности популяций служит, прежде всего, внутривидовая конкуренция. Она обычно связана с повышенной густотой произрастания. В переуплотненных посевах, например, происходит уменьшение количества семенной продукции, что имеет большое значение для сельского и лесного хозяйства. Чаще всего растения одного вида конкурируют за свет и влагу. В густых посевах они затеняют друг друга, при ограниченном количестве воды испытывают ее недостаток. В результате часть их погибает. Такое явление наиболее характерно для многих огородных культур и лесных растений. В лесу всегда значительно больше молодых растений, чем старых. Внутривидовой конкуренцией за влагу объясняется не редко встречающаяся правильное распределение пустынных растений. Создается впечатление, словно их кто-то рассадил на строго определенном расстоянии друг от друга. В понижениях местности, в оазисах это равномерная разреженность популяции растений сразу же исчезает. Подобным же образом распределяется в африканских саваннах светолюбивые и относительно влаголюбивые баобабы.
Однако следует учитывать, что популяция обычно входит в состав сообщества и что устойчивое существование биоценозов возможно только при определенных количественных соотношениях всех компонентов. Этим и вызвано необходимость регуляции численности, обеспечивающей устойчивое состояние, как отдельных популяций, так и биоценозов в целом. Механизмы регуляции численности популяций, основаны на сложных межвидовых взаимоотношениях.Список использованной литературы.
1. Радкевич В.А. “Экология”. Минск-1997, стр. 81.
2. Киселев В.Н. “Основы экологии “. Минск-1998, стр.256.
bukvasha.ru
Человечество возникло в тропической зоне Африки, и следует полагать, что поэтому наиболее древним и исходным для остальных экологических типов человека является тропический тип. Об этом свидетельствует и наибольший полиморфизм морфофункциональных признаков человека в рамках этого типа на территории Центральной и Северо-Восточной Африки. В связи с этим уместно обратиться к учению Н.И. Вавилова о центрах происхождения и многообразия культурных растений, в соответствии с которым именно в зонах первоначального вовлечения биологических видов в социальную среду наблюдается наиболее выраженный наследственный полиморфизм местных популяций по большому количеству признаков.
Об этом свидетельствует и то, что именно в тропической Африке в непосредственной близости друг от друга, т.е. под действием сходных экологических факторов, обитают наиболее низкорослые и наиболее высокорослые племена, например пигмеи, готтентоты и бушмены, с одной стороны, и массаи — с другой. Пределы изменчивости африканских популяций, например, по росту и массе тела таковы, что в них укладываются все известные на Земле человеческие популяции. Основные же черты тропического типа в Африке, отмеченные выше, остаются неизменными. Они же характеризуют монголоидные популяции Индокитая, Малайского архипелага и некоторые группы индейских племен зоны влажных тропических лесов Центральной и Южной Америки.
Это указывает на то, что тропический адаптивный тип развивается в результате асинхронного параллелизма в эволюции человека, причем вначале на африканском континенте, а потом и в других областях (см. раздел 3.13). Отсюда следует и еще один вывод: адаптивный тип формируется на фоне расогенеза и вне зависимости от него. В процессе адаптогенеза в популяциях человека можно проследить и проявление закона гомологических рядов (см. разд. 13.3.5). Это выражается, например, в том, что в бассейне Меконга в Индокитае и на острове Суматра имеются пигмеоидные популяции, по антропометрическим признакам соответствующие африканским пигмеям.
Экологический тип умеренного пояса сформировался на базе исходного генетического и фенотипического полиморфизма тропического типа при расселении популяций человека в умеренных зонах Евразии и позже — Северной Америки. Он оформился в рамках двух больших рас: европеоидной и монголоидной.
При заселении человеком арктической зоны в Евразии и Северной Америке произошло формирование арктического типа. Независимо от него среди индейцев Южной Патагонии и Огненной Земли в приантарктической зоне Южной Америки возникли популяции индейцев, по основному комплексу признаков соответствующие арктическому типу. Это еще один убедительный пример параллелизма эволюции человеческих популяций и реализации закона гомологических рядов, а также доказательство вторичности адаптивных типов по отношению к большим расам человечества.
Формирование горного адаптивного типа иллюстрирует общие закономерности адаптогенеза, отмеченные выше. Этот тип также развился независимо от расовой и этнической принадлежности популяций — среди европеоидов Альп, Кавказа, Памира и Гималаев, а также в монголоидных популяциях Тибета, Тянь-Шаня и Анд. В связи с тем что высокогорья заселялись человеком в последнюю очередь, горный экологический тип является по происхождению самым молодым. Интересно, что, несмотря на особенно выраженную расовую и этническую разнородность этого типа, комплекс основных признаков его является монолитным. Однако в отличие от других типов, вероятно, именно горный проявляется в основном только на фенотипическом уровне и не имеет наследственной природы. Об этом свидетельствует то, что количество эритроцитов в крови и объем грудной клетки людей, переселяющихся в условия высокогорья и обратно, могут меняться на протяжении жизни одного поколения.
Следовательно, адаптивные типы человека не только отражают его прошлое, но формируются и в настоящем, а разные типы имеют и разную по длительности историю.
В настоящее время пока еще на фенотипическом уровне идет формирование адаптивного типа человека городской среды, который характеризуется широкой лабильностью психических реакций, обеспечивающих способность переживать состояния постоянного стресса, и рядом морфофизиологических особенностей, оптимальных для жизни в специфических условиях города (см. разд. 17.3.2).
Формирование экологических типов человека в значительной степени обеспечило всесветное расселение людей. Меняющаяся среда обитания под действием антропогенных факторов ставит перед популяциями человека новые задачи, решаемые и сегодня за счет не только социальных, но и биологических адаптации. Схему эволюционных взаимоотношений больших рас и адаптивных типов человека см. на рис. 15.10.
Рис. 15.10. Адаптивные типы человека и большие расы
www.ronl.ru
Экотипы, переносящие повышенные концентрации тяжелых металлов.[ ...]
Экотип — совокупность организмов того или иного вида, обладающая свойствами приспособления к тому или иному месту обитания.[ ...]
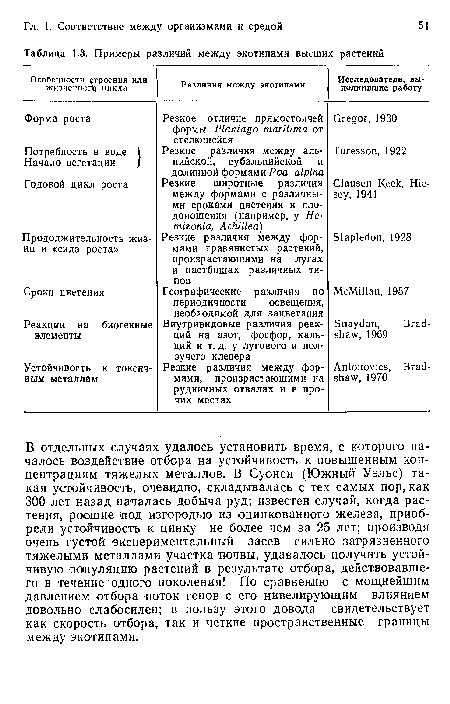
Термин «экотип» был впервые использован применительно к некоторым растениям — для описания внутривидовых генетически предопределенных локальных соответствий между организмами и средой. Выращивая растения из разнообразных природных местообитаний в одном общем в течение одного или более вегетационных сезонов, исследователи выявили значительные внутривидовые различия между этими растениями. При постановке подобных исследований существенно, чтобы растения, происходящие из разных мест, выращивались и сравнивались в одних и тех же условиях, потому что некоторые из различий между природными популяциями могут оказаться фенотипическими реакциями на условия существования, не связанными с генотипическими различиями. К примеру, на интенсивно используемом пастбище ползучие столоны клевера разветвлены очень сильно, а на сенокосном лугу — в гораздо меньшей степени, но различия эти могут возникнуть между частями одного и того же растения — стоит только поместить их в различные условия (рис. 1.14).[ ...]
По изучению экотипов сельскохозяйственных животных работы Д. Н. Кашкарова считаются классическими. Им выделено четыре экотипа овец.[ ...]
Переходя от экотипов к еще более детальному анализу популяционных структур, биологи приобретают все больший навык выявления связанной с отбором изменчивости внутри небольших локальных популяций. Такая изменчивость известна как полиморфизм. Точнее говоря, генетический полиморфизм — это «сосуществование в пределах одного и того же местообитания двух или более отчетливо различимых внутривидовых форм, причем в таких соотношениях, что постоянное присутствие редчайшей из этих форм не может быть отнесено только на счет непрерывного мутагенеза и иммиграции» (Рог[ ...]
Использование экотипов растений и животных может сыграть важную роль в развитии растениеводства и животноводства, особенно при экологическом обосновании районирования сортов и пород в регионах с разнообразными природно-климатическими условиями.[ ...]
Среди указанных экотипов встречаются группы с более или менее выраженными особенностями (подэкотипы).[ ...]
Экосистемы 11,38,118 Экотип 27 Экотон 103 Экоцентризм 540 Экоцид 391,388 Электромагнитные поля и излучения 384, 496 Энергия экосистемы 126 Эпидемии чумы, малярии и др.[ ...]
Хотя климатические экотипы (расы) еще недостаточно изучены, но и то, что сейчас известно о них, имеет определенный лесо-водственный интерес.[ ...]
Способность людей разных экотипов к акклиматизации (длительной адаптации) к высоким и низким температурам различна. В среднем акклиматизация к теплу происходит быстрее, чем к холоду, так как связана со скоростью изменения основного обмена. Акклиматизация к холоду зависит от питания, в частности, от содержания в диете белков и жиров. При этом у людей, как и у животных, образуется подкожный жировой теплоизолирующий слой.[ ...]
| 1.3 | |
Заготовка семян с учетом особенностей экотипов дуба еще не ¡налажена. Поэтому довольно часто встречаются культуры, состоящие из обеих форм дуба. В результате нередко дуб поздний оказывается на сухих местоположениях, а ранний — на морозобойных. Надо иметь в виду, что в насаждениях смешанного формового состава нет условий для ограничения роста численности вредителей. Поэтому там, где можно, надо отдавать предпочтение дубу позднему.[ ...]
Важнейшим условием существования популяции или ее экотипа является их толерантность к факторам (условиям) среды. Толерантность у разных особей и к разным частям спектра разная, поэтому толерантность популяции значительно шире, чем у отдельных особей (см. рис. 2.1 с. 37). Но из этого правила могут быть исключения, зависящие от того, какую стадию жизненного цикла проходит особь: толерантность наибольшая у покоящейся особи.[ ...]
Эвритопные виды могут быть представлены несколькими экотипами — разновидностями, приспособленными к выживанию в средах, различающихся по некоторым факторам. Например, сложноцветное растение тысячелистник образует равнинные и горные экотипы.[ ...]
Типы географо-биологической среды обитания, определяющие экотипы человека, соответствуют основным климатическим зонам. Для каждого типа среды обитания, как правило, характерен определенный тип хозяйственного уклада людей. Наибольшая населенность, развитие городов, современный уклад жизни и хозяйства более всего характерны для зон смешанных лесов умеренного климатического пояса, а также для зоны тропических лесов и степей. В этих зонах сосредоточено 79% населения Земли. Еще 12% людей обитают в горных районах, а остальные климатические зоны заселены слабо. Возникновению древнейших цивилизаций на Ближнем Востоке способствовал комплекс благоприятных экологических условий, а именно: обилие воды, плодородные почвы, речные транспортные пути, благоприятный климат в долинах рек Египта и Месопотамии.[ ...]
Примеры различий между внутривидовыми «расами», признанными экотипами, перечислены в табл. 1.3.[ ...]
Большой интерес представляют работы по разведению двух различных экотипов яка в Якутии, выявлена способность их к обитанию в условиях низких температур и потреблению небольшого количества корма. В процессе акклиматизации в новых экологических условиях у яка произошли некоторые сдвиги физиологических параметров, что нашло выражение в более высокой скорости роста молодняка. Живая масса телят в месячном возрасте увеличивается почти вдвое. Начата работа по акклиматизации яков па Кавказе, завезенных из Киргизии.[ ...]
В опытах с географическими посевами сосны выявлено несколько географических экотипов.[ ...]
Наряду с изучением наиболее обильных на Урале и в Екатеринбурге кровососущих комаров Aedes это поможет наметить пути того, как адаптационное разнообразие гемипопуляций, видовых форм, видов и сообществ комаров содействует их немалой жизнеспособности в меняющихся условиях города.[ ...]
Гетерогенность П. может быть связана с присутствием в ней генетических вариантов — экотипов, которые различаются по требованиям к условиям среды и ритмике развития (у растений — с разным временем зацветания, разной устойчивостью к морозам и засухе и т. д.). Такая гетерогенность повышает устойчивость П. к действию неблагоприятных условий. Наглядный пример генетически неоднородной П. — П. тысячелистника, в которой есть растения с белыми и розовыми цветками.[ ...]
При ведении хозяйства на лесотипологической основе нельзя не учитывать особенностей экотипов дуба и практиковать смешанную заготовку и использование семян.[ ...]
Случайный дрейф и миграции генов среди животных обычное явление, приводящее к образованию экотипов. Что же касается человека, то в настоящее время, в условиях высокой подвижности населения, географические расстояния уже не играют роли. В этих условиях генный дрейф теряет значение как фактор популяционной динамики, в то время как еще в конце XIX в. можно было говорить о существовании неких групп населения, которые называли «изолятами».[ ...]
Сукачев), которые различаются в отношении своих морфологических и экологических особенностей. До настоящего времени эти экотипы остаются совершенно недостаточно изученными, несмотря на то, что они имеют определенный лесовод-ственный интерес, поскольку отличаются рядом чрезвычайно важных для выращивания особенностей.[ ...]
В настоящее время американские генетики получили устойчивые к паразиту экотипы вяза. Выведены и ослабленные формы гриба, поражающего каштан. Этот гриб вызывает лишь локальные поражения, которые нейтрализуются образованием наплывов тканей. Разработана специальная методика «лечения» больных деревьев. Очевидно, что невозможно восстановить каштановые леса, но каштан может снова стать украшением парков.[ ...]
Известно, что восприимчивость к заболеванию одного и того же вида растений меняется в зависимости от географического происхождения растения. Одни экотипы страдают от болезни сильнее, другие слабее. В ГДР, Дании, Швеции при скрещивании европейской и американской осины получены гетерозисные гибриды, отличающиеся высокой устойчивостью к заболеванию ржавчиной и физиклядиумом.[ ...]
Имеется большое число работ, посвященных генетическому анализу озимости у растений. Всего же у A. thaliana обнаружено до 20 генов, связанных с яровизацией.[ ...]
Любая экосистема, существующая в непосредственной близости к земной поверхности, представляет собой биогеоценоз. Биогеоценоз — это реально существующее природное явление, состоящее из биоценоза и экотипа (условий среды) и характеризующееся постоянным и непрерынным течением двух противоречивых процессов — построением органического вещества с консервацией солнечной энергии и разрушением органики с высвобождением энергии. В результате этих процессов совершается обмен веществом и энергией между отдельными составными частями биогеоценоза, между ними и окружающей средой, происходит перераспределение веществ и энергии в пространстве. Схема взаимосвязей компонентов биогеоценоза приведена на рис. 1.[ ...]
Иркутская область не входит в число регионов в которых возделывают озимую пшеницу, так как этому не благоприятствуют климатические условия - ограниченные ресурсы тепла, суровые морозы, часто повторяющаяся малоснежность зим. Полевые испытания экотипов озимой пшеницы, районированных в Европейской части России и обладающих повышенной зимостойкостью, проводившиеся ранее в Иркутской области, не давали положительных результатов и обычно оканчивались гибелью растений от вымерзания (Писарев, 1963; Юдин, 1963).[ ...]
Действие необычных условий на популяцию в течение многих лет приводит к соответствующим изменениям ее генофонда. В таком случае приспособительный комплекс признаков складывается в процессе индивидуального развития и характеризует определенный экотип организмов данного вида. Это находит отражение в у экологических правилах К. Бергмана (1847) и Д. Аллена (1906).[ ...]
Пределы толерантности и экологической валентности видов во многом зависят от адаптации организмов к среде обитания. Совокупность организмов любого вида, обладающая более или менее выраженными свойствами адаптации к месту обитания, получила название экотипа. Термин «экотип» сначала применяли к растениям. Под экотипом понимали более или менее характерные формы растений одного вида, объединяемые некоторыми общими признаками и специально приспособленные к условиям определенного местообитания. Многие ботаники и экологи рассматривали вид как систему экотипов. Физиологически и морфологически экотипы могут выделяться как подвиды.[ ...]
Современный систематик, предпочитающий рассматривать классификацию популяций растений на основе экспериментальных доказательств, особенно генетических и экологических, считает необходимым вводить такие новые понятия, как компариум, ценовид и эковид. Далее эковид подразделяется на один или более экотипов. Таким образом термин экотип является заменой традиционного подвида, разновидности или формы. Экотип вполне способен скрещиваться с другими экотипами внутри эковида, но их разделяют различные требования к условиям внешней среды и, следовательно, ареал. Часто можно ожидать появления экотипической изменчивости внутри вида с широкой географической распространенностью. Ярким примером этому служит сосна обыкновенная, обычно используемая как рождественская елка. Если у всех членов эковида проявляется последовательное постепенное изменение признаков, такой вид представляет собой клинальную или экокли-нальную вариацию.[ ...]
Вообще говоря, возможность успешного развития семени во взрослое растение может реализоваться только в условиях, соответствующих его наследственной основе, исторически сложившейся приспособленности к развитию в определенных условиях внешней среды. Так, бук не может успешно развиваться на светлокаштановых почвах. Однако даже в пределах естественного ареала данного вида, например сосны обыкновенной, могут быть различные экотипы, приспособленные к более или менее определенным климатическим или почвенным условиям. Эти экотипы представляют собой наследственные формы, образовавшиеся под влиянием конкретных лесорастительных условий и приспособленные к существованию в них. Так как по внешним признакам (фенотипически) формы эти могут не различаться или плохо различаться между собой, то при использовании семян в лесоводстве весьма важно знать условия их происхождения, т. е. их местопроисхожде-ния в почвенном и, особенно, в климатическом отношении.[ ...]
Как уже много раз подчеркивалось, организмы не являются рабами физических условий среды; они приспосабливаются сами и изменяют условия среды так, чтобы ослабить лимитирующее влияние температуры, света, воды и других физических факторов. Такая компенсация факторов особенно эффективна на уровне сообщества, но возможна и на уровне вида. Виды с широким географическим распространением почти всегда образуют адаптированные к местным условиям популяции, называемые экотипами. Их оптимумы и пределы толерантности соответствуют местным условиям. Компенсация в отношении разных участков градиента температуры, света и других факторов может сопровождаться появлением генетических рас (с морфологическими проявлениями или без них) или может быть просто физиологической акклимацией. Удобным методом для проверки того, сопровождается ли появление экотипов генетическим закреплением, служит метод реципрокных пересадок. МакМиллан (1956), например, обнаружил, что злаки, принадлежащие к одному виду и идентичные по всем внешним признакам, при пересадке на экспериментальные участки из разных районов географического ареала по-разному реагировали на свет. В каждом случае они сохраняли приспособленную к исходному району сезонную цикличность (время роста и размножения). В прикладной экологии часто оставляли без внимания возможность генетического закрепления особенностей местных линий, в результате чего интродукция животных и растений часто оканчивалась неудачей, так как вместо приспособленных к местным условиям линий использовались особи из отдаленных областей. Компенсация факторов в местном или сезонном градиенте также может сопровождаться появлением генетических рас, но часто осуществляется за счет физиологической адаптации органов или сдвигов взаимоотношений «фермент—субстрат» на клеточном уровне. Сомеро (1969), например, указывает, что кратковременная температурная компенсация основана на обратной зависимости сродства фермента к субстрату от температуры, а длительная эволюционная адаптация скорее изменяет само это сродство. У животных, особенно у крупных, с хорошо развитой локомоторной способностью, компенсация факторов возможна благодаря адаптивному поведению — они избегают крайностей местного градиента условий.[ ...]
На примыкающих друг к другу морских террасах с одинаковой материнской породой (горизонт С) — пляжевые отложения и песчаники — растут лес из высоких секвой и карликовый хвойный лес. Хвойный лес чахлый и низкорослый, потому что сцементированный окислами железа горизонт В залегает примерно на глубине 45 см от поверхности. Почва над непроницаемой подстилающей породой очень кислая (pH 2,8—3,9) и бедиа Са, Мй, К, Р и другими веществами. По крайней мере одна из доминирующих карликовых сосеи является экотипом (см. стр. 143), специально адаптированным к этим крайним почвенным условиям.[ ...]
Наиболее крупными очагами адвентизации являются города с их промышленными комплексами, куда свозится масса грузов из-за границы, с которыми поступают диаспоры различных растений. Многие из занесенных семязачатков погибают, а выжившие часто становятся серьезными конкурентами естественной флоры и вытесняют ее. Парки, кладбища, железные и автомобильные дороги, вокзалы, речные порты, склады и т. п. местообитания являются «лабораториями природы», где в измененных условиях могут возникнуть новые и хорошо адаптированные экотипы и даже образоваться новые виды натурализующихся заносных растений (Зу-копп и др., 1981).[ ...]
Западным пунктом его ареала являются верховья р. Вычегды, восточным — верховья р. Олекмы за Байкалом. Общая площадь, занятая древостоями с преобладанием, в составе кедра сибирского («кедровники», «кедрачи»), исчисляется примерно в 26 млн. га. На протяжении всего своего ареала кедр находит самые различные условия существования, поднимаясь высоко в горы или встречаясь на торфяных болотах долин. До настоящего времени не найдено-сколько-нибудь значительных отличий между кедрами, чтобы установить характерные признаки их форм. Однако наличие географических и климатических экотипов кедра вполне вероятно, хотя в этом отношении кедр еще недостаточно исследован. В нормальных условиях роста кедр — дерево первой величины и благодаря густоте охвоения, цилиндрической форме крон поражает своей величественностью. Лишь под влиянием неблагоприятных условий роста внешний вид кедра резко меняется, и он выглядит чахлым невысоким деревом на сфагновых болотах Сибири или встречается в виде стелющегося по камням кустарника высоко в; горах, на границе древесной растительности.[ ...]
ru-ecology.info
