ТРОМБОЦИТЫ - раздел Образование, ФИЗИОЛОГИЯ ЭРИТРОЦИТОВ
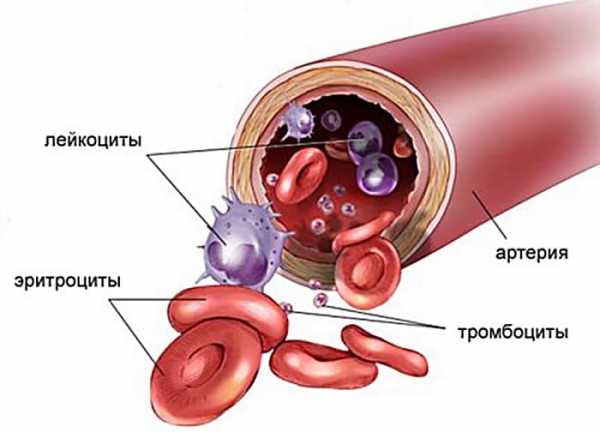
Тромбоциты - это мелкие, безъядерные кровяные пластинки неправильной формы диаметром 2-5 мкм. У здоровых людей в 1мм3 крови содержится 140-450 тысяч. В течение суток количество тромбоцитов меняется: днем их больше, ночью - меньше. Увеличивается их количество при эмоциях, физической нагрузке. После тяжелой мышечной работы содержание тромбоцитов возрастает в 3-5 раз.
Продолжительность жизни тромбоцитов - 8-11 суток. Поэтому через такой срок их количество полностью обновляется. Кровяные пластинки образуются из гигантских клеток красного мозга - мегакариоцитов. Разрушаются в МФС (мононуклеарно-фагоцитарной системе): печени, селезенке, костном мозге. Основные регуляторы образования тромбоцитов - тромбоцитопоэтины.
На наружной поверхности мембраны тромбоцитов находятся плазменные факторы свертывания крови, энзимы, рецепторы. Мембрана богата арахидоновой кислотой, из которой образуются простагландины, в частности, тромбоксан А2.
В липидный бислой мембраны тромбоцитов встроены различные гликопротеины, состоящие из разных субъединиц, которые играют роль рецепторов, ответственных за агрегацию тромбоцитов, для факторов свертывания крови и других белков. Снижение содержания различных субъединиц гликопротеинов в мембране тромбоцитов вызывает повышенную кровоточивость.
К нижнему слою мембраны прилегает зона гиалоплазмы (гиаломер), отделяющая грануломер - зону внутриклеточных органелл, в которой располагаются гранулы. В гиаломере вдоль края клетки находится краевое кольцо микротрубочек, контактирующее с миофиламентом, представляющим сократительный аппарат тромбоцита. При стимуляции тромбоцита кольцо микротрубочек, сокращаясь, смещает гранулы в центру клетки, сжимает их, вызывая секрецию содержимого наружу через систему открытых канальцев. Сокращение кольца микротрубочек позволяет тромбоциту также образовывать псевдоподии, что увеличивает его способность к агрегации. Гранулы зоны органелл содержат тромбоцитарные факторы свертывания крови, а также различные лизосомальные ферменты.
Тромбоциты имеют ряд специфических особенностей.
1 - клейкость - адгезивность - способность прилипать к чужеродной поверхности. Этому способствуют образующиеся псевдоподии.
2 - аггрегационная способность - свойство образовывать конгломераты - агрегаты тромбоцитов.
3 - выделение ферментов и факторов свертывания крови.
В тромбоцитах содержится полный набор клеточных ферментов /липазы, глюкозидазы, пептидазы и т.д./ - всего до 90 ферментов. В них много серотонина, гистамина, адреналина, норадреналина. Лизоцим тромбоцитов, подавляя гепарин, ускоряет свертывание крови.
При разрушении тромбоцитов из них выделяются факторы свертывания крови. Обнаружено до 20 факторов.
ЖИДКИЕ СРЕДЫ ОРГАНИЗМА Жидкости, находящиеся в организме, подразделяются на внутриклеточные и внеклеточные. Вн
ВНУТРЕННЯЯ СРЕДА ОРГАНИЗМА Среда – это совокупность условии обитания живых существ. Выделяют 2 среды: 1 – внешнюю ср
СИСТЕМА КРОВИ Понятие “система крови” в физиологию ввел отечественный клиницист Г.Ф. Ланг в 1939 году. Система крови
Основные функции крови 1. Транспортная функция заключается в переносе необходимых для жизнедеятельности веществ. Из нее вытекают
Количество и состав крови Объем крови у человека составляет 6-8 % массы тела, т.е. 4-5 л при массе 60 кг. Объем крови в норме постоянен: не за
ЭРИТРОЦИТЫ Эритроциты – красные кровяные тельца. Они наиболее часто имеют двояковогнутую форму. Диаметр эритроци
Функции эритроцитов 1. Основная функция эритроцитов – транспортная, и прежде всего вытекающая из неё - дыхательная - это
Гемоглобин, его строение, количество и соединения Гемоглобин – основная составная часть эритроцита. Гемоглобин (Hb) – дыхательный пигмент из группы хром
Жизненный цикл эритроцитов В норме число эритроцитов у человека постоянно. В крови мужчин 4–5,1×1012/л, женщин – 3,7-4,7 ×1012/
Эритропоэз Вся масса эритроцитов, находящихся в циркулирующей крови, в кровяных депо и костном мозге, составляет эри
Некоторых показателей крови и эритроцитов Объем крови относительно массы тела с возрастом снижается. У новорожденных он составляет 15 %, в конце 1-го го
Работа 1. Техника взятия крови Исследование крови является одним из важнейших диагностических методов в клинической практике. С это
Работа 2. Подсчет эритроцитов пробирочным методом Ход работы. Берут кровь из вены или из капилляра пальца. К счетной камере притирают покровное стек
Работа 3. Определение гемоглобина по Сали Ход работы. В среднюю пробирку гемометра наливают 0,1 н р-р HCl до нижней метки. Пипеткой берут 20 мм3
Работа 5. Определение гематокритного числа Ход работы. Гематокритные капилляры – это стеклянные трубочки, разделенные на 100 равных частей. Их
ЛЕЙКОЦИТЫ Лейкоциты, или белые кровяные тельца, – истинные клетки крови. Они содержат ядро и органеллы.
Лейкоцитозы и лейкопении Лейкоциты – одна из самых реактивных систем организма, поэтому их количество и качество изменяются при сам
Различают несколько видов физиологических лейкоцитозов. Пищеварительный. Он наблюдается после приема пищи и при этом число лейкоцитов увеличивается незначите
НЕЙТРОФИЛЫ Нейтрофилы – основная часть лейкоцитов периферической крови. В норме число сегментоядерных нейтрофилов в
Основные функции нейтрофилов 1 – фагоцитоз; 2 – внутриклеточное переваривание; 3 – цитотоксическое дейст
БАЗОФИЛЫ Базофилы – это самая малочисленная группа лейкоцитов. В периферической крови их содержится 0,5-1% (22
ЭОЗИНОФИЛЫ Эозинофилыкрупнее нейтрофилов. Их количество в периферической крови колеблется в пределах 1-5% (45-70
Основные функции эозинофилов 1. Уменьшение аллергической реакции гиперчувствительности немедленного типа.При этой реакции
Некоторые физиологические свойства клеток МФС 1. Клетки МФС, как и нейтрофилы, обладают спонтанной (ненаправленной) и хемотаксической двигательной а
Функции моноцитов и макрофагов I. Секреторная функция заключается в высвобождении активных веществ, участвующих во многих процес
ЛИМФОЦИТЫ Лимфоциты – центральное звено иммунной системы. Их количество в периферической крови в норме сос
Другие разновидности лимфоцитов Кроме двух видов лимфоцитов (Т- и В-) существуют и другие лимфоциты. Третья группа лимфоцитов – ни Т-, ни
Плазматические клетки В крови человека в норме плазматические клетки отсутствуют. Они находятся в костном мозге, лимфатических у
РЕГУЛЯЦИЯ ЛИМФОПОЭЗА Продукция лимфоцитов регулируется на 3-х различных уровнях. Межклеточный уровень регуляции осущес
МЕХАНИЗМЫ ЗАЩИТЫ КЛЕТОЧНОГО ГОМЕОСТАЗА В процессе жизнедеятельности организма во внутреннюю среду могут попадать из внешней среды молекулы и мик
Неспецифическая резистентность организма Она является по своему происхождению врожденной. К ней относятся барьеры между внешней и внутренней средой
Специфические механизмы защиты клеточного гомеостаза Иммунитет – комплекс реакций, направленных на поддержание гомеостаза при встрече с антигенами, н
Реакции, осуществляющие иммунологический надзор 1. Спонтанная клеточная цитотоксичность (СКЦ) – основная реакция, осуществляемая макрофагами, нейтрофи
Регуляция иммунитета На интенсивность иммунного ответа влияют нервный и гуморальный механизмы регуляции. Так, раздражение разл
ИММУННАЯ РЕГУЛЯТОРНАЯ СИСТЕМА В последние годы установлено, что иммунокомпетентные клетки участвуют не только в иммуногенезе, но и являю
ВОЗРАСТНЫЕ ИЗМЕНЕНИЯ ЛЕЙКОЦИТОВ Лейкоциты появляются в конце 3-го месяца внутриутробного развития, а на последней неделе беременности и
Особенности неспецифической резистентности Микрофагальная функция нейтрофилов формируется уже внутриутробно (по одним данным, на 20-23 неделе, а по д
Особенности иммунной системы Дифференцировка на Т- и В-лимфоциты начинается еще внутриутробно. В-лимфоциты определяются на 10-12-й
Работа 1. Подсчет лейкоцитов пробирочным способом Ход работы. В центрифужную пробирку набирают 0,4 мл 5% р-ра уксусной кислоты, подкрашенной метиленов
Работа 2. Определение лейкоцитарной формулы Ход работы. Мазок крови помещают под микроскоп и считают лейкоциты в иммерсионной системе. Необхо
СОСТАВ ПЛАЗМЫ КРОВИ Плазма– это жидкая часть крови. Она имеет желтоватый цвет, слегка опалесцирует. В ее состав входят
ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА КРОВИ Функции крови во многом определяются ее физико-химическими свойствами, к которым относятся: цвет, относите
Некоторых показателей физико-химических свойств крови В ходе развития детей физико-химические свойства крови претерпевают некоторые изменения. У новорожден
ГРУППЫ КРОВИ Часто для сохранения жизни при кровопотерях и травмах приходится возмещать кровь. Первое упоминание о пере
Серологический состав основных групп крови системы АВО Группы крови Эритроцит Антигены П Л А З М А Агглютинины и гемо
Система агглютиногенов резус Среди агглютиногенов, не входящих в систему АВ0, особое значение имеет агглютиноген резус. Этот аг
Антигенов, антител и правил переливания крови Групповые антигены.В онтогенезе человека антигены системы АВО появляются у эмбриона на 5-6 неделе
Самостоятельная работа студентов Для самостоятельной работы необходимы:водяная баня, клиническая центрифуга, стерильные скарификат
Работа 1. Определение осмотической резистентности эритроцитов Эритроциты характеризуются различной устойчивостью к повреждающим факторам. Более высокую осмотическую с
Работа 3. Химический гемолиз Гемолиз - один из способов разрушения эритроцитов, сопровождающийся выходом гемоглобина в плазму крови или
Работа 4. Определение группы крови по системе АВО перекрестным методом Под группами крови по системе АВО подразумевают различные сочетания антигенных свойств эритроцитов, назыв
Ход работы. 1. Приготавливают 5% взвесь однократно отмытых в 0,9% р-ре NaCl стандартных эритроцитов. 2. В 2 маркированные
Работа 5. Определение резус-принадлежности Применяют набор двух видов цоликлонов: анти-D Супер (выявляет полные антитела при определении резус-принадл
Реакция агглютинации на плоскости с помощью цоликлона анти-D Супер (содержащего полные IgМ антитела) Ход работы. Определение проводят в помещении с хорошим освещением. Планшетку подогреваютпри темп
Реакция агглютинации в присутствии высокомолекулярных субстанций с помощью цоликлона анти-Д А. Реакция агглютинации на плоскости при участии цоликлона анти-Д Эта реакция проводится также как в пу
Функции системы гемостаза 1. Содержание крови в жидком состоянии, что обеспечивает процессы нормального кровообращения и тк
Плазменнные факторы свертывания крови Плазменные факторы обозначаются римскими цифрами в порядке их хронологического открытия (соглас
Тромбоцитарные факторы Эти факторы обозначают арабскими цифрами. ПФ 1 - идентичен фактору V плазмы. Это Ас-глобулин пл
Функции тромбоцитов 1. Ангиотрофическая функция. Тромбоциты - постоянные поставщики питательных веществ в эндотелий с
Участие эритроцитов в свертывании крови Механическая роль эритроцитов заключается в том, что они являются плацдармом для прикрепления нитей фи
Эритроцитарные факторы 1 - тромбопластичеcкий фактор /эритроцитин/. Этот фактор очень активный. Он является основой для образования
Лейкоцитарные факторы 1 - тромбопластический фактор - фосфолипиды мембраны лейкоцитов. 2 - антигепариновый фактор - стимулятор
Тканевые факторы 1 - тромопластический фактор. Он содержится во всех тканях и органах. Это соединение очень активное: 1 г ткани
СОСУДИСТО-ТРОМБОЦИТАРНЫЙ ГЕМОСТАЗ Сосудисто-тромбоцитарный гемостаз обеспечивает остановку кровотечения в микроциркуляторных сосудах, диа
I фаза-образование протромбиназ Различают 4 вида протромбиназ: тканевую, эритроцитарную, тромбоцитарную и лейкоцитарную. Причем 3 последние
III фаза–превращение фибриногена в фибрин Эта фаза протекает в 3 этапа. На первом этапе фибриноген под влиянием тромбина расщепляется на фибрин-моном
ФИБРИНОЛИЗ Спонтанный /естественный, самопризвольный/ фибринолиз - сложная реакция между плазминовой системой органи
Кровяной проактиватор плазминогена
Плазминоген к
ПРИЧИНЫ ПОДДЕРЖАНИЯ ЖИДКОГО СОСТОЯНИЯ КРОВИ Главная задача организма в условиях нормальной жизнедеятельности – поддержание крови в жидком состоянии.
ЛАТЕНТНОЕ МИКРОСВЕРТЫВАНИЕ КРОВИ Латентное, или скрытое, микросвертывание в циркуляции крови происходит в небольших масштабах непрерывно. В
ПРИЧИНЫ ВНУТРИСОСУДИСТОГО ТРОМБООБРАЗОВАНИЯ Существует множество механизмов, поддерживающих жидкое состояние крови. Однако внутри этих механизмов мог
РЕГУЛЯЦИЯ СВЕРТЫВАНИЯ КРОВИ Регуляция свертывания крови осуществляется на трех уровнях. На молекулярном уровне обеспечиваетс
СИСТЕМА ГЕМОСТАЗА И ИММУННАЯ СИСТЕМА Система гемостаза взаимодействует с иммунной системой, что особенно заметно при патологии. Так, на гемоста
СИСТЕМА ГЕМОСТАЗА И ПОТЕНЦИАЛЫ ВОЗБУДИМЫХ ТКАНЕЙ Известно, что при возбуждении цитоплазма переходит из состояния золя в гель. В паузах между потенциалами де
СИСТЕМА РЕГУЛЯЦИИ АГРЕГАТНОГО СОСТОЯНИЯ КРОВИ И ТРОМБОГЕМОРРАГИЧЕСКИЙ СИНДРОМ Система свёртывания крови входит в состав функциональной системы - системы регуляции агрегатного сост
Основные компоненты системы РАСК Центральные органы. К ним относятся костный мозг, печень и селезенка. Костный мозг продуцирует кле
ВОЗРАСТНЫЕ ИЗМЕНЕНИЯ ГЕМОСТАЗА Система свёртывания крови формируется в процессе внутриутробного развития. Появление коагуляционно-а
Самостоятельная работа студентов Для самостоятельной работы необходимы: клиническая центрифуга, торсионные весы, водяная баня, секу
Работа 1. Определение времени свёртывания крови по Ли-Уайту Время свёртывания венозной крови оценивает общую коагуляционную активность цельной крови по скорости
Работа 3. Определение времени рекальцификации плазмы Время рекальцификации плазмы – это время свертывания цитратной плазмы при t 37оС после добавления
Работа 4. Определение протромбинового времени Протромбиновое время по Квику - тест, характеризующий внешний механизм свёртывания крови. В основе этог
Работа 5. Определение тромбинового времени Тромбиновое время - это время свёртывания цитратной плазмы после добавления к ней раствора тромбина сл
Работа 6. Определение уровня фибриногена по Рутберг Его можно определить путём взвешивания: цитратная плазма рекальцифицируется, полученный сгусток высушива
Работа 7. Определение длительности кровотечения по Дьюку Метод основан на определении времени кровотечения из нанесённой на поверхности кожных покровов ранки стан
Работа 8. Исследование ретракции кровяного сгустка по Матиссу Оценка ретракции используется как один из наиболее важных показателей функциональной активности тромбоци
Работа 9. Определение свёртывания крови по Сухареву Принцип метода заключается в определении времени спонтанного свёртывания цельной капиллярной крови и поз
Работа 10. Определение спонтанного фибринолиза и ретракции по Кузнику В клинической практике необходимо знать результирующую взаимодействия компонентов фибринолитической сис
СРЕДСТВА ИНФУЗИННО-ТРАНСФУЗИОННОЙ ТЕРАПИИ В настоящее время инфузионно-трансфузионная терапия прочно заняла самостоятельное место в лечении различ
КРИСТАЛЛОИДНЫЕ И КОЛЛОИДНЫЕ РАСТВОРЫ Они обладают специфическими свойствами и получили в современной хирургии и интенсивной терапии особое зна
Фотогемометрия Это более точный метод определения содержания гемоглобина. Так цианметгемоглобиновый фотометрический мет
Цитофотометрия Этот метод основан на фотометрическом измерении степени поглощения света определённых длин волн, например
Электронно-автоматический метод Он основан на разных принципах, но наиболее часто применяется импульсный принцип - разница электропроводно
Тромбоэластография Это метод записи процесса свёртывания крови и образования сгустка на тромбоэластографе. Принцип рабо
ЗАНЯТИЕ 1 1. Верны ли утверждения: а) гомеостаз - это способность сохранять постоянство внутренней среды организ
Педиатрического отделения ЗАНЯТИЕ 1 1. Укажите последовательность этапов гемопоэза в период внутриутробного развития.
ЗАНЯТИЕ 2 1. Укажите отличия в составе белой крови у новорожденных после рождения. 1. Количество лейк
ЗАНЯТИЕ 4 1. Верны ли утверждения: а) у новорожденных общее количество крови составляет 15% массы тела, б) у вз
А) нет, б) нет, в) да. СИТУАЦИОННЫЕ ЗАДАЧИ 1. Человек потерял 2л крови. Группа крови не установлена. Какова тактика врача? Что
Ответы на вопросы тестового контроля знаний Занятие 1 Занятие 2 Занятие 3 Занятие 4 1. 1 2. 1,2 3. 3 4. 3 5. 4 6.
Знаний для студентов педиатрического отделения Занятие 1 Занятие 2 Занятие 3 Занятие 4 1. 2,3,1,4 2. 1,2,3,4
allrefers.ru
Количество просмотров публикации Тромбоциты - 78
Эритроциты, число, форма и размеры. Образование, продолжительность жизни и разрушение эритроцитов. Гемолиз и анемия. Скорость оседания эритроцитов (СОЭ). Функции эритроцитов. Гемоглобин.
Количество и состав крови, гематокрит. Плазма и сыворотка крови. Белки плазмы и их функциональное значение. Электролиты плазмы. Осмотическое онкотическое давление крови. Кровезаменяющие растворы.
Кровь и лимфа как внутренняя среда организма. Основные функции крови.
Физиология крови и кровообращения
Лекция 13-14
Кровь - это жидкая соединительная ткань, обеспечивающая жизнедеятельность других тканей и клеток. Основная функция крови — это транспорт различных веществ в организме. Кровь переносит дыхательные газы - кислород и углекислый газ - как в физически растворенном, так и в химически связанном виде — это дыхательная функция. Кровь доставляет питательные вещества от органов, где они всасываются или хранятся, к месту их потребления (трофическая функция), конечные продукты обмена — к органам выделения (экскреторная функция). Кровь осуществляет транспорт гормонов, витаминов и ферментов, образующихся в организме, от органов, где они вырабатываются, к органам-мишеням (регуляторная функция).
Благодаря высокой теплоемкости своей главной составной части – воды, кровь обеспечивает распределение тепла, образующегося в процессе метаболизма и его выделение во внешнюю среду через легкие и поверхность кожи, ᴛ.ᴇ. выполняет терморегуляторную функцию.
Способность организма обезвреживать инородные тела и патогенные микроорганизмы связана с защитной функцией крови, обусловленной, прежде всего, присутствием в ней фагоцитирующих и антителообразующих клеток
Важным свойством крови является ее участие в остановке кровотечения. Эта функция обусловлена способностью крови свертываться.
Довольно важной функцией крови, которую часто выпускают из виду, особенно у беспозвоночных является передача механической силы.
На долю крови приходится примерно 6-8% общего веса тела, или 4-6 л у взрослого человека. Она представляет собой непрозрачную красную жидкость, состоящую из бледно-желтой плазмы и взвешенных в ней клеток - красных кровяных телец (эритроцитов), белых кровяных телец (лейкоцитов) и кровяных пластинок (тромбоцитов). — часть объёма крови приходящаяся на долю эритроцитов принято называть гематокритом. В норме у взрослого мужчины гематокрит равен 44-46 об.%, а у женщины 41-43 об.%. В случае если принять вязкость воды за 1, то средняя относительная вязкость крови у здорового человека составит 4,5 (3,5-5,4).
90-91% веса плазмы крови человека приходится на долю воды; 6,5-8% составляют белки, остальные два процента – низкомолекулярные вещества. Её рН незначительно колеблется от 7,37 до 7,43, составляя в среднем (в артериальной крови) около 7,4.
Около 60% всех белков плазмы составляют альбумины, сравнительно небольшие белки, имеющие М.В. около 69000, обладающие, однако огромной общей поверхностью. Это свойство позволяет молекулам альбумина сорбировать и транспортировать с током крови как эндогенные вещества, такие как соли желчных кислот, жирные кислоты, билирубин, так и некоторые экзогенные лекарственные препараты: пенициллин и другие антибиотики, сульфаниламиды.
Другой группой белков плазмы являются глобулины. По электрофоретической подвижности, а электрофорезом принято называть движение электрически заряженных макромолекул в электрическом поле, глобулины разделяют на альфа1-и альфа2-, бета- и гамма- глобулины, причем минимальной электрофоретической подвижностью обладают γ-глобулины, а максимальной α1-глобулины. В состав фракции α1-глобулинов входят преимущественно гликопротеины, переносящие около 60% всей глюкозы плазмы. Фракция α2-глобулинов включает медьсодержащий белок церулоплазмин, связывающий около 90% всей меди, содержащейся в плазме. При этом с током крови к клеткам медь транспортируется в большей степени альбуминами, тогда как церулоплазмин играет важную роль в антиокислительной защитной системе крови. К другим белкам из данной фракции относятся витамин В12-связывающий глобулин (транскобаламин), билирубин-связывающий глобулин и кортизолсвязывающий глобулин (кортизол, глюкокортикостероид - гормон вырабатываемый корой надпочечников и лекарственный препарат).
К β-глобулинам относятся важнейшие белковые переносчики липидов (липопротеины). Около 75% всех липидов плазмы (фосфолипидов, жиров, холестерола) входят в состав липопротеинов. Кроме переносчиков липидов в состав β-глобулинов входит переносчик ионов железа - трансферрин. Этот белок способен связать и транспортировать два атома трехвалентного железа. К фракции α- и β-глобулинов принадлежат факторы свертывания крови.
Во фракцию γ-глобулинов входят большинство защитных белков - антител, защищающих организм от вторжения вирусов и микроорганизмов. Почти при всех заболеваниях, особенно воспалительных и иммунизации, содержание γ-глобулинов в плазме повышается.
Важным компонентом плазмы крови является фибриноген, занимающий промежуточное положение в ряду электрофоретической подвижности между β- и γ-глобулинами. Этот белок обладает свойством становиться нерастворимым и принимать волокнистую структуру превращаясь в процессе свертывания крови в фибрин. Плазма, лишенная фибриногена, принято называть сывороткой.
Альбумины и фибриноген образуются в печени, а глобулины - в печени, костном мозгу, селезенке, лимфатических узлах.
Суммарная концентрация солей в крови млекопитающих составляет около 0,9%. Соли находятся в диссоциированном состоянии. Основными катионами являются натрий (140-150 мМ) и калий (5 мМ), а анионами - хлор (около 100 мМ) и бикарбонат (НСО3 около 30 мМ). Низкомолекулярные электролиты вносят основной вклад (96%) в осмотическое давление крови, составляющие в норме 7,3 атм (5600 мм рт. ст.). Вследствие относительно низкой молекулярной концентрации белки вносят небольшой вклад в осмотическое давление крови (около 25 мм рт.ст.). Белковый компонент в общем осмотическом давлении крови, принято называть онкотическое давление. Осмотическое давление крови и межклеточной (интерстициальной) жидкости различается именно на величину онкотического давления, поскольку их электролитный состав совпадает, а белок в межклеточной жидкости практически отсутствует, вследствие неспособности макромалекул в проходить через стенки капилляров. Согласно гипотезе Старлинга онкотическое давление крови, способствует переходу воды из тканей в кровяное русло, однако ему противодействует гидростатическое давление крови в капиллярах. В их артериальной части оно достигает приблизительно 35 мм рт.ст. и, следовательно, превышает величину онкотического давления плазмы. По этой причине здесь жидкость переходит из крови в окружающую капилляры ткань. Наоборот, у венозного конца капилляра гидростатическое давление ниже онкотического (около 15 мм. рт.ст.) и вода из тканей переходит обратно в кровь. Благодаря такому механизму кровь находится в непрерывном обмене с тканевой жидкостью. Несмотря на существование этого механизма, далеко не вся профильтровавшаяся жидкость возвращается в кровеносные сосуды. Образуется лимфа или тканевая жидкость, которая по специальным лимфатическим сосудам отводится в венозную систему.
Эритроциты или красные кровяные тельца - самые многочисленные клетки крови. У мужчин в 1 мкл крови содержится в среднем 5,1 млн. эритроцитов, а у женщин - 4,6 млн. Эритроциты человека - это безъядерные клетки, имеющие форму двояковогнутых дисков. Средняя величина их диаметров у взрослого человека равна 7,5 мкм. Благодаря двояковогнутой форме эритроцита его поверхность больше, чем, в случае если бы он имел форму шара. Общая площадь поверхности эритроцитов взрослого человека составляет около 3800 м2. Большая площадь поверхности эритроцитов способствует выполнению ими основной функции - переносу дыхательных газов. Эритроциты обладают большой способностью к обратимой деформации при прохождении через узкие изогнутые капилляры. По мере старения клеток и при некоторых патологиях пластичность эритроцитов уменьшается, что является одной из причин задержки и разрушения таких клеток в ретикулярной ткани селезенки. Эритроциты образуются из предшествующих стволовых недифференцированных клеток. Стволовые клетки имеют ядро и обладают неограниченной способностью делиться, но не выполняют конкретных функций. При делении стволовой клетки образуются две дочерние, одна из которых превращается (созревает) в зрелый эритроцит, а вторая остается недифференцированной и сохранившей способность к делению стволовой клеткой. Этот процесс происходит в красном костном мозге плоских костей и принято называть эритропоэз. Андрогены усиливают эритропоэз, а эстрогены угнетают. Созревшие эритроциты циркулируют в крови в течение 100-120 дней, после чего они фагоцитируются клетками ретикулоэндотелиальной системы печени, селезенки и костного мозга. При этом в определенной степени и любая другая ткань способна разрушать кровяные тельца, о чем свидетельствует постепенное исчезновение "синяков" (подкожных кровоизлияний).
Снижение способности крови переносить кислород принято называть анемия. Анемия дословно означает "бескровие". В клинике этим термином обозначают, прежде всего, снижение способности крови переносить кислород в связи с недостатком гемоглобина. При анемии должна быть уменьшено число эритроцитов, либо содержание в них гемоглобина, либо то и другое. Чаще всего встречается железодефицитная анемия. Она должна быть следствием недостатка железа в пище, нарушения всасывания железа в пищеварительном тракте или хронической кровопотери (к примеру, при язвенной болезни). При некоторых патологических состояниях вследствие повышенной хрупкости эритроцитов возрастает скорость гемолиза (разрушения эритроцитов). В случае если образование эритроцитов не компенсирует ускоренного их разрушения, возникает гемолитическая анемия.
Удельный вес эритроцитов (1,096) выше, чем плазмы (1,027), и в связи с этим в пробирке с кровью, лишенной возможности свертываться, они медленно оседают на дно. Скорость оседания эритроцитов (СОЭ) у здорового мужчины составляет 3-6 мм, а у женщин 8-10 мм за первый час. Главным фактором, определяющим СОЭ, является белковый состав плазмы. СОЭ снижается при увеличении в плазме содержания альбуминов и повышается при увеличении количества глобулинов.
В эритроцитах на долю белка гемоглобина приходится 34% общего и 90% сухого веса. Содержание гемоглобина в крови человека составляет в среднем 158 г/л у мужчин и 140 г/л у женщин. Гемоглобин относится к классу хромопротеинов. Его молекула состоит из четырех полипептидных цепей, в состав каждой из которых входит гем - протопорфирин, в центре которого находится ион двухвалентного железа. М.В. гемоглобина около 64500. В процессе переноса кислорода его молекула образует обратимую связь с гемом, причем валентность железа при этом не меняется, ᴛ.ᴇ. железо не окисляется. Гемоглобин (Hb), присоединивший кислород, становится оксигемоглобином (HbO2). Поскольку присоединение кислорода к гемоглобину в данном соединении не сопровождается изменением валентности железа, реакцию связывания кислорода гемоглобином называют не окислением, а оксигенацией; обратный процесс принято называть деоксигенацией. Гем может подвергаться и истинному окислению. При этом железо становится из двух валентного трехвалентным. Окисленный гем носит название метгема, а вся белковая молекула - метгемоглобина. В крови человека метгемоглобин находится в незначительных количествах, но при некоторых заболеваниях и отравлениях, к примеру, нитритами его содержание увеличивается. Опасность таких состояний состоит по сути в том, что окисленный гемоглобин не способен отдавать кислород. При отравлении "угарным газом" (СО), образуется карбоксигемоглобин (железо при этом остается двухвалентным). Окись углерода активнее взаимодействует с гемом, чем кислород, в связи с этим гемоглобин утрачивает способность связываться с кислородом и переносить его, что может оказаться смертельным. Слабое отравлении СО обратимый процесс, поскольку карбоксигемоглобин хоть и медленно, но разлагается. Образующийся в процессе метаболизма СО2 также может связываться с гемоглобином, взаимодействуя не с гемом, а аминогруппами этого белка. Образование карбаминовых соединений (Белок-Nh3 + CO2→Белок-NHCOO- + H+) происходит быстро и не нуждается в присутствии ферментов. Подобным образом кровью транспортируется до 30% углекислоты. Остальные 70% СО2 переносится в виде иона бикарбоната͵ образование которого контролируется содержащейся в эритроцитах ферментом карбоангидразой.
Кислородная емкость крови. Для расчета количества кислорода способного связаться с гемоглобином крови, следует учесть, что молекула последнего состоит из четырех субъединиц, каждая из которых связывает молекулу кислорода. Следовательно, реакцию оксигенации можно записать следующим образом:
Hb + 4O2 → Hb(O2)4
Из приведенного уравнения реакции следует, что один моль гемоглобина может связать до 4 молей кислорода; 64500 г гемоглобина связывают 89,6 л, а 1 г - 1,39 мл кислорода. При анализе газового состава крови получают несколько меньшее значение 1,34-1,36 мл О2 на 1г Hb, ᴛ.ᴇ. насыщение гемоглобина кислородом составляет около 97%. При прохождении крови через тканевые капилляры используется только 25% общей кислородной емкости, хотя при интенсивной физической нагрузке из оксигемоглобина должна быть извлечено до 50% кислорода. ТО кислородная емкость 1 л крови составит в среднем 150 г х 1,34=200 мл.
Лейкоциты, классификация и функции. Защитная функция крови, фагоцитоз. Тромбоциты и их функции. Свертывание крови. Фибринолиз. Группы крови человека.
Лейкоциты формируют в организме человека мощный кровяной и тканевой барьеры против микробной, вирусной и паразитарной инфекции.
Лейкоциты, или белые кровяные тельца - это клетки с ядрами, не содержащие гемоглобина. В 1 мкл крови здорового человека содержится 4000-10000 лейкоцитов (4-9 109/л). Большая часть лейкоцитов (более 50%) находится за пределами сосудистого русла, в межклеточном пространстве, более 30% - в костном мозге. Процес образования и созревания лейкоцитов принято называть лейкопоэзом. Увеличение количества лейкоцитов принято называть лейкоцитозом, уменьшение - лейкопенией. Различают физиологический и реактивный лейкоцитоз. Первый чаще всего наблюдается после приема пищи, мышечной работе, беременности, сильных эмоциях. Второй вид лейкоцитоза характерен для воспалительных процессов и инфекционных заболеваний. Лейкопения (наиболее тяжелая форма – агранулоцитоз) приводит к резкому снижению защитных сил организма. При лейкопении прежде всего уменьшается число нейтрофилов, как за счёт подавления лейкопоэза, так и в следствие усиленного удаления лейкоцитов из крови. Лейкопения наблюдается при некоторых инфекционных заболеваний. Выявленная в последние годы неинфекционная лейкопения связана главным образом с повышением радиационного фона, широким применением ряда лекарственных препаратов (цитостатики, анальгин). Злокачественная, неконтролируемая пролиферация лейкоцитов, ведущая к увеличению числа малодифференцированных клеток, не способных выполнять свою функцию наблюдается при лейкозах.
Учитывая зависимость оттого, содержит ли цитоплазма зернистость (ᴛ.ᴇ. многочисленные лизосомы и секреторные гранулы) или она однородна, лейкоциты делятся на две группы: гранулоциты и агранулоциты.
ГРАНУЛОЦИТЫ - образуются в костном мозге и составляют 60% всех лейкоцитов крови. Срок жизни 2 суток. Гранулоциты в свою очередь, подразделяются на три вида: эозинофилы, гранулы которых окрашиваются в розовый цвет кислым красителем эозином; базофилы - гранулы окрашиваются в синий цвет основными красителями; нейтрофилы, гранулы воспринимают те и другие краски.
1. Нейтрофилы или нейтрофильные лейкоциты составляют 93-96% от всех гранулоцитов. Так как с возростом клеток изменяется форма их ядра, их еще называют полиморфноядерные лейкоциты. Часть гранул нейтрофилов представлена лизосомами, содержащими многочисленные энзимы: лизоцим, повреждающий стенку бактерий; катионные белки, нарушающие дыхание и рост микроорганизмов; протеазы и кислые гидролазы, позволяющие переваривать захваченные объекты. Другие гранулы содержат энзимы, участвующие в процессах регенерации тканей.
Нейтрофилы являются самыми важными функциональными элементами неспецифической защитной системы крови. Οʜᴎ способны быстро мигрировать и накапливаться в инфицированном или поврежденном участке тела, где они фагоцитируют, а затем убивают и переваривают бактерии. Выбор направления движения обусловлен появлением в участке повреждения хемотаксических факторов. Под влиянием этих факторов или хемоаттрактантов клетки поляризуются и начинают ползти по направлению к источнику аттрактанта. В результате большое количество лейкоцитов переходит в пораженную ткань. Самым мощным хемотаксическим эффектом обладают лейкотриены, они секретируются активированными Т-лимфоцитами (хелперами) после воздействия на них бактериальных агентов. Нейтрофилы способны получать энергию путем анаэробного гликолиза, в связи с этим могут существовать в очагах воспаления, бедных кислородом. Бактерицидный эффект нейтрофилов связан с возникающим в них через 30-60 секунд после контакта бактериального агента с мембраной нейтрофила "метаболическим (дыхательным) взрывом", при котором резко увеличивается потребление кислорода и образуются активные формы кислорода (АФК), являющихся сильными окислителями. Этот процесс инициируется НАДФН-оксидазой, катализирующей реакцию между восстановленным пиридиннуклеотидом и молекулярным кислородом благодаря чему образуется перекись водорода и супероксидный анион-радикал. АФК, в частности анион-радикал кислорода, являются эффективными хемоаттрактантами. Нейтрофилы содержат в так называемых азурофильных гранулах большое количество фермента миелопероксидазы (5 % от сухого веса клеток). При фагоцитозе, сопровождающимся обязательной активацией фагоцитирующих клеток и развитием дыхательного взрыва, МПО секретируется вместе с АФК как во внеклеточную среду, так и в фагосому. В обоих компартментах МПО усиливает окислительный потенциал АФК, катализируя образование различных гипогалитов, при этом пероксид водорода используется как ко-субстрат в соответствии со следующим уравнением:
Cl-+ h3O2 + H+HOCl + h3O
Именно с продукцией гипохлорита (HOCl), который является чрезвычайно эффективным бактерицидным агентом, токсичным для бактерий даже в концентрации 50 мкМ, связывают участие МПО в процессах неспецифического иммунитета. Бактерицидный эффект нейтрофилов связан также с разрушением бактерий и продуктов распада тканей лизосомными ферментами. Образующийся в очагах воспаления гной состоит, главным образом, из нейтрофилов и их остатков.
2. Базофильные гранулоциты составляют 0,5-1% всех лейкоцитов крови. Эти клетки способны к фагоцитозу и миграции из кровяного русла в ткани. Могут накапливать в гранулах биологически активные вещества, очищая от них ткани. Базофилы способны выделять гистамин, который участвует в формировании аллергических реакций. Эти реакции развиваются при повторном воздействии нетоксических антигенов. При аллергии через нескольких минут самое большее часов наблюдается покраснения кожи, появляются сыпи, иногда развиваются спазмы легких (к примеру, при воздействии пыльцы - сенная лихорадка) и в крайних случаях аллергия приводит к анафилактическому шоку.
3. Эозинофильные гранулоциты составляют 2-4% всех лейкоцитов крови. Οʜᴎ обладают фагоцитарной и бактерицидной активностью и способны убивать микроорганизмы и более крупных паразитов, к примеру личинок глистов (аскарид, трихинелл), благодаря целому ряду механизмов и в первую очередь - продукции пероксида водорода. Эозинофилы уменьшают концентрацию биологически активных соединений, возникающих при аллергии, ᴛ.ᴇ. являются антагонистами базофилов.
АГРАНУЛОЦИТЫ - в цитоплазме этих лейкоцитов при окрашивании зернистость не выявляется. Οʜᴎ представлены лимфоцитами и моноцитами.
Лимфоциты - составляют 25-40% всех лейкоцитов. Образуются во многих органах: лимфатических узлах, миндалинах, пейеровых бляшках, аппендиксе, селезенке, вилочковой железе (тимусе), но главным образом в костном мозге. Лимфоциты играют ключевую роль в иммунитете (след. лекция).
Моноциты - 4-8% всех лейкоцитов крови. Образуются в костном мозге. Способность к фагоцитозу у них более выражена, чем у других форменных элементов крови. Из крови выходят в окружающие ткани - здесь растут, в них увеличивается содержание лизосом и митохондрий. Достигнув зрелости, превращаются в неподвижные клетки - тканевые макрофаги. Макрофаги способны функционировать в анаэробных условиях. Их цитотоксические свойства связаны с развитием дыхательного взрыва, в ходе которого образуются анион радикал кислорода, Н2О2, гидроксильный радикал (‣‣‣ОН) и другие формы активного кислорода и азота͵ в т.ч. монооксид азота (NO). Вместе с тем, макрофаги секретируют более 100 биологически активных соединений. Вблизи очага воспаления эти клетки могут размножаться делением. Тканевые макрофаги образуют ограничивающий вал вокруг тех инородных тел, которые не бывают удалены с помощью фагоцитоза. Остеокласты (специализированные макрофаги костной ткани) применяют АФК для разрушения кости - обязательного условия ее обновления. Во всех этих случаях клетки-защитники быстро поглощают большое количество О2, используют его для образования АФК при помощи расположенной в плазматической мембране НАДФН-оксидазы дыхательного взрыва. Важное значение АФК для защиты от бактерий доказывается тем, что при инактивирующей мутации этого фермента возникает хронический септический грануломатоз: фагоцитированные микроорганизмы остаются живыми, что приводит к повторным хроническим инфекциям и чревато сепсисом.
При инфекционных заболеваниях наблюдаются характерные изменения в содержании и соотношении различных форм лейкоцитов. При остром инфекционном заболевании на начальном этапе – нейтрофильный лейкоцитоз и снижение числа лимфоцитов и эозинофилов, затем возрастает количество моноцитов и это свидетельствует об успешной борьбе организма с инфекцией и в заключительный период болезни, при окончательном выздоровлении увеличивается содержание лимфоцитов и эозинофилов, очищающих организм от токсинов.
Тромбоциты или кровяные пластинки это плоские, безъядерные клетки с большим диаметром 1-4 мкм и толщиной 0,5-0,75 мкм. В норме один мкл крови содержит от 150 до 300 тыс. тромбоцитов. Образуются в костном мозге из гигантских клеток – мегакариоцитов (а те из стволовых). Из одного мегакариоцита может образоваться до 1000 тромбоцитов. В кровяных пластинках содержатся специфические гранулы, содержащие серотонин и вещества, участвующие в свертывании крови. Кровяные пластинки циркулируют в кровяном русле 5-11 дней и затем разрушаются в печени, легких, селезенке. Обычно тромбоциты пребывают в крови в неактивном состоянии (в норме неповрежденный эндотелий сосудов вырабатывает NO – фактор ингибирующий агрегацию тромбоцитов) и активируются только при крайне важно сти остановки кровотечения и свертывания крови (гемостаз).
referatwork.ru
Количество просмотров публикации Функции тромбоцитов. - 364
Кровяные пластинки образуют главную линию обороны организма против внезапных потерь крови. Οʜᴎ аккумулируются почти тотчас вместе повреждения кровеносных сосудов и закупоривают их вначале временной, а затем постоянной тромбоцитарной пробкой, облегчают превращение фибриногена в фибрин в поврежденном участке.
Структура и функции тромбоцитов.Циркулирующие в крови тромбоциты имеют дисковидную форму, диаметром от 2 до 5 мкм, объёмом 5- 10 мкм3. Тромбоциты оказались весьма сложным клеточным комплексом, представленным системами мембран, микротрубочек, микрофиламентом и органелл. Используя технику, позволяющую разрезать распластанный тромбоцит параллельно поверхности, в клетке выделяют несколько зон: периферическую, золя-гель, внутриклеточных органелл (рис.6.4.). На наружной поверхности периферической зоны располагается покров, толщиной до 50 нм, содержа-
Рис.6.4. Ультраструктурная организация тромбоцита.
Сечение параллельное горизонтальной плоскости.
ЕС — периферическая зона тромбоцита͵ СМ — трехслойная мембрана, SMF — субмембранный филомент, МТ— микротрубочки, Gly — гликоген. Зона органнелл — М — митохондрии, G — гранулы, DB — плотные гранулы, DTS — система плотных трубочек, CS — система открытых канальцев.
21У
щий плазматические факторы свертывания крови, энзимы, рецепторы, необходимые для активации тромбоцитов, их адгезии (приклеивания к субэндотелию) и агрегации (приклеиванию друг к другу). Так, мембрана тромбоцитов содержит "мембранный фосфолипидный фактор 3" — "фосфолипидную матрицу", формирующую активные коагуляционные комплексы с плазменными факторами свертывания крови. Мембрана богата также арахидоновой кислотой, в связи с этим важным ее компонентом является фермент — фосфолипаза А,, способная образовывать свободную арахидоновую кислоту для синтеза простагландинов, из метаболитов которых формируется короткожи-вущий агент — тромбоксан А2, вызывающий мощную агрегацию тромбоцитов. Активация фосфолипазы А2 в мембране тромбоцита осуществляется при ее контакте с коллагеном и фактором Вилле-бранда — адгезивными белками субэндотелия, обнажающимися при повреждении эндотелия сосудов.
В липидный бислой мембраны тромбоцитов встроены гликопро-теины I, II, III, IV, V. Гликопротеин I состоит из субединиц — Iа, Iв, Iс. Iа — рецептор, ответственный за адгезию тромбоцитов к коллагену субэндотелия. Комплекс "Iв — фактор свертывания крови IX" на поверхности кровяных пластинок выполняет функцию рецептора для фактора Виллебранда, что также крайне важно для адгезии пластинок на субэндотелии. Iс обеспечивает связывание с еще одним адгезивным белком субэндотелия — фибронектином, а также распластывание пластинки на субэндотелии.
Гликопротеин II состоит из субединиц IIа и IIв, необходимых для всех видов агрегации тромбоцитов. Гликопротеин Ша с гликопро-теином IIв образуют Са-зависимый комплекс, связывающий на тромбоцитах фибриноген, что обеспечивает дальнейшую агрегацию тромбоцитов и ретракцию (сокращение) сгустка. Гликопротеин V гидролизируется тромбином, поддерживает агрегацию тромбоцитов. Снижение в мембране тромбоцитов содержания различных субединиц гликопротеинов I-V вызывает повышенную кровоточивость.
К нижнему слою периферической зоны прилегает зона золя-геля гиалоплазмы, в свою очередь отделяющая зону внутриклеточных ор-ганелл. В указанной зоне вдоль края клетки располагается краевое кольцо микротрубочек, контактирующее с микрофиламентом, представляющим сократительный аппарат тромбоцита. При стимуляции тромбоцита кольцо микротрубочек, сокращаясь, смещает гранулы к центру клетки ("централизация гранул"), сжимает их, вызывая секрецию содержимого наружу через систему открытых канальцев. Сокращение кольца микротрубочек позволяет тромбоциту также образовывать псевдоподии, что увеличивает его способность к агрегации.
Зона органелл тромбоцитов содержит плотные гранулы, альфагра-нулы 1 и II типов. В плотных гранулах находятся АДФ, АТФ, кальций, серотонин, норадреналин и адреналин. Кальций участвует в регуляции адгезии, сокращении, секреции тромбоцита͵ активации его фосфолипаз и, следовательно, продукции эндоперекиси, простагландинов, в ходе дальнейших превращений которых образуется тромбоксан А,. АДФ секретируется в больших количествах при ад-
220
гезии тромбоцитов к стенке сосуда и способствует прикреплению циркулирующих тромбоцитов к адгезированным, тем самым поддерживая рост тромбоцитарного агрегата. Серотонин (5-гидроксит-риптамин) секретируется тромбоцитом во время "реакции освобождения гранул" и обеспечивает вазоконстрикцию в месте повреждения.
Альфа-гранулы I типа содержат антигепариновый фактор пластинок 4, фактор роста тромбоцитов, тромбоспондин (гликопротеин G) и др. Размещено на реф.рфАнтигепариновый фактор тромбоцитов 4 секретируется тромбоцитами под влиянием АДФ, тромбина, адреналина, сопровождая агрегацию тромбоцитов. Тромбоспондин образует комплекс с фибриногеном на поверхности активированных тромбоцитов, необходимый для формирования тромбоцитарных агрегатов. Тромбоцитарный ростковый фактор (ТРФ) — полипептид, стимулирующий рост гладких мышц сосудов и фибробластов, восстановление сосудистой стенки и соединительной ткани. Благодаря его свойствам, кровяные пластинки поддерживают целостность сосудистой стенки. Больные с тромбоцитопенией имеют сниженную устойчивость стенки капилляра, в связи с этим петехии (точечные кровоизлияния в коже) появляются вслед за легкими травмами или изменением давления крови. Петехии вызваны слущиванием эндотелия капилляров. В нормальных условиях возникший дефект устраняется пластинками, секретиру-ющими ТРФ.
Альфа- гранулы II типа содержат лизосомальные энзимы (кислые гидролазы). Большая часть гранул исчезает после адгезии или агрегации тромбоцита. Этот феномен ("реакция освобождения гранул") имеет место после активации тромбоцита различными соединениями — тром-боксаном А2, АДФ, адреналином, тромбином, протеолитическими энзимами, бактериальными эндотоксинами, коллагеном и др.
Тромбоцитопоэз и его регуляция.Под тромбоцитопоэзом понимают процесс образования тромбоцитов в организме. В основном, он протекает в костном мозге и включает следующие этапы: коло-ниеобразующая единица мегакариоцитарная (КОЕ-мег) -> промега-кариобласт -> мегакариобласт -> промегакариоцит -> зрелый мегака-риоцит -> тромбоцитогенный мегакариоцит -> тромбоциты (рис.6.5.).
Истинные митозы, ᴛ.ᴇ. деление клеток, присущи только КОЕ-меᴦ. Для промегакариобластов и мегакариобластов характерен эндомитоз (глава I), ᴛ.ᴇ. удвоение ДНК в клетке без ее деления. После остановки эндомитоза, в основном после 8, 16, 32, 64-кратного удвоения ДНК, мегакариобласт начинает дифференциацию до тромбоцитарного мегакариоцита͵ образующего тромбоциты.
В костном мозге тромбоцитогенные мегакариоциты локализованы преимущественно на поверхности синусного эндотелия и их цито-плазматические отростки проникают в просвет синуса через эндотелий. Одни из них проникают на 1-2 мкм в просвет синуса и фиксируют мегариоцит на эндотелии (функция "якоря"). Второй тип отростков представлен вытянутыми цитоплазматическими лентами (до 120 мкм в длину), входящими в просвет синуса и получившими
221
Рис.6.5. Схема дифференцировки клеток мегакариоцитарной серии.
СКК — стволовая кроветворная клетка; КОЕгммэ — КОЕгрануло-цитарно-мегакариоцитарно-моноцитарно-эритроцитарная; КОЕэрмег — КОЕэритроцитарно-мегакариоцитарная; КОЕмег — КОЕмегакариоцитарная; КОЕмег-1 — менее дифференцированная; КОЕмег-2 — более дифференцированная клетка; 0.07, 0.48, 0.74 — вероятность вовлечения клетки-предшественницы в мегакариоцитарную дифференциацию.
название протромбоцитов. Их количество у одного мегакариоцита может достигать 6-8. В просвете синуса цитоплазма протромбоцита после локальных сокращений разрывается, и он образует около 1000 тромбоцитов. В циркулирующее русло поступают и протромбоциты. Вышедшие в кровь протромбоциты достигают микроциркуляторного русла легких, где из них освобождаются тромбоциты. По этой причине количество тромбоцитов оказывается более высоким в легочных венах, чем в легочной артерии. Количество тромбоцитов, образовавшихся в легких, может достигать 7- 17% от массы тромбоцитов в крови.
У человека время полного созревания мегакариоцитов занимает 4-5 дней. Костный мозг человека содержит около 15.106 мегакари-оцитов/кг веса тела. Дневная продукция тромбоцитов у человека 66000+14600 в 1 мкл крови. В среднем мегакариоцит высвобождает до 3000 тромбоцитов. Количество тромбоцитов в крови взрослого человека достигает 150-375.109л; у детей —150-250- 109/л. Содержание тромбоцитов в крови взрослого ниже 150.10 9/л воспринимается как тромбоцитопения.
Общая популяция тромбоцитов представлена циркулирующими в крови (70%) и находящимися в селезенке (30%). Накопление тром-
222
боцитов в селезенке происходит благодаря более медленному их движению через извилистые селезеночные корды и занимает до 8 минут. Сокращение селезенки (к примеру, вызванное адреналином) освобождает пластинки в общий кровоток. Существование селезеночного депо тромбоцитов объясняет, почему их количество неизменно выше у спленэктомированных (с удаленной селезенкой) людей, чем у нормальных индивидуумов. У пациентов со спленомега-лией (увеличенной селезенкой) значительная часть циркулирующих тромбоцитов медленно продвигается через увеличенную селезенку, количество пластинок в крови оказывается сниженным, возникает тяжелая тромбоцитопения.
Продолжительность жизни тромбоцитов человека колеблется от 6,9 до 9,9 дней. Их разрушение происходит, преимущественно, в костном мозге и, в меньшей степени, в селезенке и печени.
В плазме крови людей обнаружен колониестимулируюший фактор мегакариоцитарный (КСФ-мег), стимулирующий митозы и дифференциацию КОЕ- меᴦ. Стимулом для его образования является истощение содержания мегакариоцитов и их предшественников в костном мозге. Регуляция тромбоцитопоэза в фазу немитотического развития мегакариоцитов осуществляется другим гуморальным фактором — тромбопоэтином. Его количество в плазме повышается при усилении потребления тромбоцитов (воспаление, необратимая агрегация тромбоцитов). Тромбопоэтин необходим для полного созревания цитоплазмы мегакариоцитов, нормального формирования в ней тромбоцитов. Регуляция мегакариоwитопоэза включает и особые вещества — его гуморальные ингибиторы, угнетающие как пролифе-ративную, так и немитотическую стадии развития мегакариоцитов. Ингибитор деления КОЕ-мег выделен из активированных тромбоцитов. Это гликопротеин, массой в 12-17 кд. Источником ингибитора тромбоцитопоэза является, также, селезенка.
6.4. Механизмы гемостаза.
Тромбоцитарный гемостаз.Тромбоцитарная реакция, ᴛ.ᴇ. реакция тромбоцитов на нарушение целостности сосудистой стенки, формируется параллельно реакции самих сосудов на повреждение — их сокращение в месте повреждения, шунтирование крови выше поврежденного участка. Поскольку сосудисто-тромбоцитарная реакция на повреждение первой обеспечивает остановку кровотечения из микрососуда, то ее называют сосудисто- тромбоцитарным или первичным гемостазом, а формирование и закрепление тромбов — вторичным коагуляционным гемостазом. Первая волна агрегации тромбоцитов связана с их адгезией к пластинкам, приклеившимся рецепторами гликопротеинов I и II к фактору Виллебранда, фибро-нектину и коллагену субэндотелия поврежденных тканей. Вторая волна агрегации вызвана высвобождением АДФ из плотных гранул тромбоцитов, образованием тромбоксана А, в их мембране, взаимодействием мембранных гликопротеинов Ив-Шс с фибриногеном, V — с тромбином.
223
Вторую волну агрегации тромбоцитов обеспечивают тромбоксан А2 и тромбин. Мощным стимулом для образования тромбоксана А2 являются коллаген и фактор Виллебранда, активирующие фосфоли-пазу A2. Тромбин, образуемый на мембране кровяной пластинки, благодаря секреции ею фактора V и взаимодействию его с глико-протеином V, начинает быстро действовать как сильный агрегаци-онный агент. Новые порции тромбоцитов соприкасаются с адгези-рованными тромбоцитами, начинается агрегация вновь принесенных кровью клеток, и вновь включаются вышеописанные механизмы, а это, в свою очередь, усиливает агрегацию новой партии тромбоцитов и т.д. Тромбоцитарный тромб уплотняется и сокращается. Его формирование усиливается образованием фибрина (фибринового тромба) вследствие активации системы свертывания крови.
Система свертывания крови.Эта система обеспечивает остановку кровотечения, благодаря образованию фибриновых тромбов. В физиологических условиях большинство факторов свертывания крови содержится в ней в неактивном состоянии, в виде неактивных форм ферментов, обозначаемых римскими цифрами: I, II, VIII, IX, X, XI, XII, XIII(табл.6.3.). При этом повреждение сосуда или клеток крови приводит к активации этих ферментов. При этом переход в активное состояние одного фактора вызывает активацию следующего, образуя каскадную реакцию, заканчивающуюся образованием фибриновых нитей, формирующих сеть тромба (рис.6.6.). В начале этой реакции в крови, в зоне поврежденного сосуда образуется активная протромбо-
Таблица6.3 Факторы свертывания крови
| Фактор | Период | Молекулярная | Нормальная | |
| полужизни | масса | концентрация | ||
| (дальтон) | в плазме | |||
| (мг/мл) | ||||
| 1 _ | фибриноген | 3,7 дня | 340.000 | 1500-4000 |
| II - | - протромбин | 2,8 дня | 72.500 | |
| III - | - тромбопластин | — | — | |
| IV ■ | - ионы кальция | — | — | 0,9-1,2 ммоль/л |
| V - | - проакцелерин | 15-24 ч | 330.000 | |
| VI - | - проконвертин | 1,2-6 ч | 48.000 | менее 1 |
| VII | с — антигемофилический А | 5-12 ч 1 | менее 0,5 | |
| VIII | ФВ — фактор Виллебранда | 24-40 ч 1 | I .UUU.UUU | |
| IX - | - антигемофильный фактор Б, | 20-24 ч | 57.000 | |
| фактор Кристнаса | ||||
| X - | - фактор Стюарта-Прауэра | 32-48 ч | 59.000 | |
| XI - | - антигемофильный фактор В, | 40-48 ч | 160.000 | |
| предшественник плазменного | ||||
| тромбопластина | ||||
| XII | — фактор Хагемана | 48-52 ч | 76.000 | |
| XIII | — фибрин-стабилизирующий | 5-12 дней | 320.000 | |
| фактор | ||||
| Плазминоген | 2,2 дня | 90.000 | ||
| Прекалликреин (фактор Флетчера) | — | 85.000 | ||
| Высокомолекулярный кениноген | — | 150.000 |
224
Рис.6.6. Схема последовательной активации факторов свертывания крови.
а — активированный.
киназа, превращающая неактивный протромбин в тромбин — активный протеолитический фермент, отщепляющий от молекулы фибриногена 4 пептида мономера. Каждый из мономеров имеет 4 свободных связи. Соединяясь ими друг с другом, конец к концу, бок к боку, они в течение нескольких секунд формируют волокна фибрина. Их сеть вначале слаба, однако под влиянием фибрин-стабилизирующего фактора (фактора XIII), также активируемого в крови тромбином в присутствии ионов кальция, в фибрине образуются дополнительные ди-сульфидные связи и сеть фибриновых волокон становится прочной. В этой сети задерживаются тромбоциты, лейкоциты, эритроциты и белки плазмы, формируя фибриновый тромб.
В крови содержатся и неферментные белки — акселераторы, также обозначаемые римскими цифрами (фактор V, VII), обеспечивающие ускорение во много тысяч раз течение реакции свертывания
225
крови при взаимодействии с фосфолипидными поверхностями тромбоцитов (с тромбоцитарным фактором 3), участками мембран (мик-ромебран) других поврежденных клеток.
Превращение в тромбин протромбина происходит под воздействием активной протромбиназы. Различают два пути формирования активной протромбиназы (рис.6.6) — внешний, возникающий при повреждении сосудистой стенки и окружающих тканей, и внутренний — возникающий при контакте крови с субэндотелием, компонентами соединительной ткани сосудистой стенки или при повреждении самих клеток крови.
При внешнем пути из мембран клеток поврежденной ткани в плазму высвобождается комплекс фосфолипидов (тканевой тром-бопластин или фактор III), вместе с фактором свертывания крови VII действующий как протеолититеский энзим на фактор X. Активированный таким образом фактор X в присутствии ионов кальция немедленно соединяется с тканевыми фосфоли-пидами и фактором V. Этот комплекс и составляет активную протромбиназу. Через несколько секунд после ее формирования часть протромбина превращается в тромбин. Образовавшийся тромбин, действуя как протеолитический фермент, активирует фактор V, что дополнительно резко ускоряет превращение протромбина в тромбин.
Внутренний механизм свертывания крови запускается ее травма-тизацией (появление разрушенных тромбоцитов и эритроцитов) или контактом с субэндотелием, что активирует фактор XII. Фактор ХПа (а — активированный) действует энзиматически на фактор XI и при взаимодействии с тромбоцитарным фактором 3 и высоко молекулярным кининогеном плазмы превращает его в фактор ХIа. Эта реакция ускоряется прекалликриином плазмы. ХIа активирует фактор IX (антигемофильный фактор В). После образования фактора IХа формируется комплекс: "фактор IХа + фактор VIII (антигемофильный глобулин А,) + тромбоцитарный фактор 3 + ионы кальция". Этот комплекс активирует фактор X. Фактор Ха образует с фактором V и тромбоцитарным фактором 3 новый комплекс, называемый протромбиназой, который в присутствии ионов кальция в считанные секунды превращает протромбин в тромбин.
Следует подчеркнуть весьма важную роль фосфолипидной матрицы (тромбоцитарного фактора 3) в повышении активности протромбиназы — при отсутствии фосфолипидной матрицы ее активность снижается в 1000 раз!
Нарушенное формирование комплекса, активирующего фактор X, сопровождается нарушениями гемостаза. Так, тромбоцитопения, создавая дефицит тромбоцитарного фактора 3, приводит к геморрагическому диатезу, дефицит фактора IX вызывает гемофилию В, дефицит фактора VIII — гемофилию А.
Активация протромбокиназы по внешнему пути занимает около 15 секунд, а по внутреннему — 2-10 минут. Активированная протром-бокиназа и ионы кальция превращают протромбин в тромбин. Количество образующегося тромбина прямо пропорционально количеству
226
активированной протромбокиназы. Протромбин синтезируется в печени, для его образования необходим витамин К, в связи с этим поражение печени или гиповитаминоз К сопровождаются кровоточивостью.
После образования сгустка через 30-60 минут начинается его сокращение {ретракция). Оно связано с сокращением нитей актина и миозина тромбоцитов, а также сети фибрина под влиянием тромбина и ионов кальция. В результате ретракции сгусток сжимается в плотную массу, тромб уплотняется.
Противосвертывающие механизмы.Физиологические антикоагулянты поддерживают кровь в жидком состоянии и ограничивают процесс тромбообразования. К ним относятся антитромбин III,гепарин, протеины "С" и "S", альфа-2-макроглобулин, нити фибрина. На долю антитромбина III (альфа-2-глобулин) приходится 75% всей антикоагулянтной активности плазмы. Он является основным плазменным кофактором гепарина, ингибирует активность тромбина, факторов Ха, IXa, VIIa, XIIa. Его концентрация в плазме достигает 240 мг/мл.
Гепарин — сульфатированный полисахарид. Образует комплекс с антитромбином III,трансформируя его в антикоагулянт немедленного действия и в 1000 раз усиливая его эффекты, активируя неферментный фибринолиз.
Протеины "С" и "S" синтезируются в печени при участии витамина К. Протеин "С" инактивирует активированные факторы VIII и V. Протеин "S" резко снижает способность тромбина активировать факторы VIIIи V. Нити фибрина обладают антитромбинным действием, благодаря адсорбции на них до 85-90% тромбина крови. Это помогает сконцентрировать тромбин в формирующемся сгустке и предотвратить его распространение по току крови.
Эндотелиальные клетки неповрежденной сосудистой стенки препятствуют адгезии тромбоцитов на ней. Этому же противодействуют гепариноподобные соединения, секретируемые тучными клетками соединительной ткани, а также простациклин, синтезируемый эндо-телиальными и гладкомышечными клетками сосуда, активация протеина "С" на эндотелии сосуда. Гепариноподобные соединения эндотелия (и гепарин крови) усиливают антикоагуляционную активность антитромбина III.Простациклин является мощным ингибитором агрегации тромбоцитов. В физиологических условиях он образуется в венозных и артериальных эндотелиальных клетках из ара-хидоновой кислоты, через этап формирования эндоперекисей про-стагландинов. В результате между образующейся в тромбоцитах про-агрегационной субстанцией — тромбоксаном А2 и антиагрегационной активностью простациклина сосудистой стенки возникает динамическое равновесие, регулирующее агрегацию тромбоцитов. Сниженная или утраченная продукция простациклина участком эндотелия должна быть одной из причин агрегации кровяных пластинок и вести к формированию тромба.
Тромбомодулин — рецептор тромбина на эндотелии сосудов взаимодействуя с тромбином, активирует белок "С", обладающий
227
способностью высвобождать тканевой активатор плазминогена из стенки сосуда. Дефицит белка "С" сочетается с повышенной свертываемостью крови, наклонностью к тромбозам. Свертывание крови в сосуде предупреждает и гладкая поверхность эндотелия, препятствующая включению внутреннего пути формирования активной протромбиназы. Мономолекулярный слой белка, адсорбированный на поверхности эндотелия, отталкивает факторы свертывания и тромбоциты, также предупреждая свертывание крови.
Фибринолиз — это процесс разрушения {лизиса) сгустка крови, связанный с расщеплением фибрина, фибриногена на мелкие фрагменты. Важнейшая функция фибринолиза — восстановление просвета сосудов, закупоренных тромбами. Расщепление сгустка крови осуществляется системой ферментов, активными компонентами которой являются плазмин — протеолитический фермент, расщепляющий нити фибрина, а также фибриноген, факторы свертывания крови V, VII, XII и протромбин.
В плазме крови содержится неактивная форма фермента плазминогена — белок плазминоген. Существует несколько механизмов его активации. Одни из них связан с высвобождением белкового тканевого активатора из эндотелиальных клеток на участке формирующегося кровяного сгустка. Активировать плазминоген могут также активированный фактор свертывания крови XII при взаимодействии с калликриином и высокомолекулярным кининогеном, а также ли-зосомальные ферменты поврежденной ткани. Активатором плазминогена является урокиназа, образующаяся в почках и выделяющаяся с мочой. В кровь попадает ее небольшое количество, и с ней связано лишь около 15% общей фибринолитической активности. Активатором плазминогена является стрептокиназа бактерий. Данным действием стрептококков в инфицированных тканях объясняют растворения плазменного сгустка в лимфе и тканевой жидкости и распространение инфекции.
Активный плазмин блокируется антиплазминами. Наиболее деятельным является а2-антиплазмин (а2-глобулин), способный нейтрализовать 2/3 всего плазмина. Другой ингибитор фибринолиза — а2- макроглобулин. Продукты, образующиеся в ходе фибринолиза, тормозят агрегацию тромбоцитов и формирование волокон фибрина, тормозят фибринолиз. Лизис кровяных сгустков продолжается в течение нескольких дней. Выброс тканевых активаторов фибринолиза имеет место под влиянием физических нагрузок, адреналина, норадреналина.
6.5. Общие закономерности кроветворения.
Кроветворные клетки-предшественницы.Ежечасно у здорового человека в крови погибает 20 миллиардов тромбоцитов, 10 миллиардов эритроцитов и 5 миллиардов лейкоцитов. Эта непрерывная утрата клеток постоянно компенсируется равным ей количеством вновь образующихся форменных элементов крови. Масштаб этого восполнения огромен: примерно каждые два года в организме че-
228
ловека производится масса клеток крови, равная массе его тела. Указанный огромный пролиферативный потенциал кроветворной ткани заключен в стволовых кроветворных клетках (СКК) — предшественницах, способных к самообновлению, ᴛ.ᴇ. производству дочерних СКК на протяжении всей жизни человека. СКК диффрен-цируется: 1) в направлении клетки-предшественницы всех линий миелопоэза, ᴛ.ᴇ. гранулоцитопоэза, моноцитопоэза, мегакариоцито-поэза и эритропоэза; 2) в направлении клетки-предшественницы Т-лимфоцитов; 3) клетки-предшественницы В-лимфоцитов (рис.6.7.)-Эти клетки-предшественницы получили название колониестимули-рующих единиц (КОЕ) или колониеобразующих клеток (КОК), от формируемых ими при культивировании ин витро колоний. Так, КОЕ-ГММЭ — гранулоцитарно-макрофагально-мегакариоцитарно-эритроцитарная колониеобразующая единица получила название от формируемой ею колонии, состоящей из гранулоцитов, макрофагов, мегакариоцитов и эритроцитов; пре-КОЕ-Т — от колонии, состоящей из Т- клеточных субпопуляций; пре-КОЕ-В — от колонии, состоящей из В-лимфоцитов. КОЕ-ГММЭ в ходе дифференциации формируют би- и унипотентные КОЕ, которые также классифицируют на основании произведенного ими потомства при культивировании ин витро. Так, нейтрофильные гранулоциты и макрофаги образуются из одной общей КОЕ. По этой причине возникло ее название — КОЕ-гранулоцитарно-моноцитарная (КОЕ-ГМ). КОЕ, образующая колонии из эритроидных клеток и мегакариоцитов, принято называть эрит-роиитарно-мегакариоцитарной (КОЕ-Эмег) и т.п. Это примеры бипотентных клеток, ᴛ.ᴇ. клеток, дифференцирующихся в направлении каких-либо двух линий гемопоэза. Формирующиеся из них унипотентные КОЕ образуют колонии из клеток только одной линии. В связи с этим возникли их названия — КОЕ-эритроцитар-ные, эозинофильные, базофильные, нейтрофильные, мегакариоци-тарные.
Бипотентные КОЕ обладают значительной способностью к размножению. К примеру, КОЕ-Эмег, будучи стимулированной ростковыми факторами, способна совершать до 13 митозов и образовывать несколько десятком КОЕ-Э, из которых далее формируется несколько тысяч эритроидных клеток. Количество КОЕ-Эмег достигает 30-40 на 105 клеток костного мозга, а содержание КОЕ-Э в нем почти в 10 раз выше, чем КОЕ-Эмеᴦ. Из каждой КОЕ-Э образуется до 50 эритроцитов. КОЕ-ГМ способна совершать 5-6 делений, образующиеся из нее КОЕ-Г и КОЕ-М —еще 5-6 делений каждая. Это позволяет одной КОЕ-ГМ формировать тысячи зрелых клеток-потомков — гранулоцитов и моноцитов.
Дифференциация клеток-предшественниц КОЕ-ГММЭ до унипо-тентных КОЕ сопровождается формированием рецепторов к гемопо-этическим гормонам (интерлейкину-3, КСФ-ГМ, эритропоэтину, тромбоцитопоэтину), к нейромедиаторам, катехоламинам, тиреотроп-ному гормону, производным тестостерона, в связи с этим указанные гормоны регулируют пролиферацию и дифференциацию клеток крови.
229
Рис.6.7. Схема развития гемопоэтических клеток и регулирующие их колониестимулирующие факторы.
СКК — стволовая кроветворная клетка;
КОЕ-ГММЭ — колониеобразующая единица гранулоцитарно-моноци-
тарно-мегакариоцитарно-эритроцитарная;
КОЕ нейтр.эоз. — КОЕ нейтрофильно-эозинофильная;
КОЕ-гм — КОЕ гранулоцитарно-моноцитарная;
КОЕ эр.эоз. — КОЕ эритроцитарно-эозинофильная;
КОЕ эр.меᴦ. — КОЕ эритроцитарно-мегакариоцитарная;
КОЕ ᴦ.эрит. — КОЕ гранупоцитарно-эритроцитарная;
КОЕ меᴦ. — КОЕ мегакариоцитарная;
КОЕ баз. — КОЕ базофильная; КОЕ нейтр. Размещено на реф.рф— КОЕ нейтрофильная;
КОЕ эоз. — КОЕ эозинофильная; КОЕ-мс — КОЕ моноцитарная;
КСФ — колониестимупирующий фактор;
КСФ-мег — КСФ мегакариоцитов; КСФ-г — КСФ гранулоцитов;
КСФ-баз. — КСФ базофилов; КСФ-м — КСФ моноцитов;
КСФ-эоз. — КСФ эозинофилов.
230
Регуляция пролиферации и дифференциации КОЕ (КОК).Рост различных КОЕ в культурах удается получить, добавляя стимулирующие его факторы. Отсюда факторы, стимулирующие образование гра-нулоцитарных колоний, получили название "колониестимулирующие факторы гранулоцитарные" — КСФ- Г, макрофагальных — КСФ- М, гранулоиитарно- макрофагальных — КСФ- ГМ, эозинофильных — КСФэоз, мегакариоиитарных — КСФмег, стимулирующие развитие колоний из КОЕ-ГММЭ — мульти-КСФ (позже обозначенные как интерлейкин 3 — ИЛ-3). Установлено, что ИЛ-3 и КСФ-ГМ стимулируют пролиферацию и дифференциацию не только КОЕ-ГММЭ и КОЕ-ГМ, но и других бипотентных КОЕ. В связи с этим, ИЛ-3 и КСФ-ГМ рассматривают как факторы неспецифические, поддерживающие самообновление и пролиферацию КОЕ-ГММЭ и бипотенциаль-ных. Все КСФ-ы гликопротеины, относимые ныне к семье полипептидных гормонов, регулирующих гемопоэз. Источниками ИЛ-3 и КСФэоз являются Т-лимфоциты, спленоциты; КСФ-ГМ — Т-лимфоциты, моноциты, эндотелиальные клетки и фибробласты; КСФ- Г, КСФ- М — моноциты и фибробласты; эритропоэтина — перитубуляр-ные клетки почек, купферовские клетки печени.
Гены, контролирующие синтез ИЛ-3, КСФ-Г и КСФ-М у человека, находятся на дистальной части длинного плеча 5 хромосомы, КСФ-Г — локализованы у человека на 17 хромосоме, эритропоэтина — на 7 хромосоме. Нарушение участков генома, ответственных за синтез молекулярных регуляторов гемопоэза, вызывает у человека тяжелые нарушения в системе крови. Продукцию КСФ усиливают различные стимулы: гипоксия —эритропоэтина, тромбо-цитопения — тромбоцитопоэтина, микробная инфекция — КСФ-ГМ, КСФ-Г, КСФ-М, гельминтная инфекция — КСФэоз. Но каждый из перечисленных раздражителей одновременно стимулирует в организме и продукцию неспецифических ростковых факторов — ИЛ-3, КСФ-ГМ и других. При этом ИЛ-3 и КСФ-ГМ стимулируют как пролиферацию КОЕ-ГММ и КОЕ бипотентных, так и формирование на их мембране рецепторов к КСФ-ам, действие которых направлено на унипотентные КОЕ. Высокие же концентрации КСФ- Г, КСФ- М и других стимулируют пролиферацию и дальнейшую дифференциацию унипотентных клеток-предшественниц гранулоцитарного, моноцитарного и др. Размещено на реф.рфрядов.
На интенсивность продукции КСФ-ов оказывает регулирующее влияние вегетативная нервная система. Так, введение животным а-и В-адреноблокаторов значительно уменьшает уровень КСФ-ов в крови. Стимулируют гемопоэз Т-лимфоииты. Так, действие на организм возбуждающих гемопоэз факторов (кровопотеря, высотная гипоксия и др.) вызывает миграцию лимфоцитов в костный мозг и активацию ими КОЕ. КСФ-ы регулируют функции не только КОЕ, но и зрелых клеток крови. Так, КСФ- м усиливает фагоцитарную активность, метаболизм, миграцию в ткани зрелых нейтрофилов и моноцитов-макрофагов. КСФ-Г стимулирует бактерицидную, фагоцитарную и цитотоксическую активность этих клеток. ИЛ-3 также усиливает цитотоксическую активность макрофагов, увеличивает
231
эозинофильную фагоцитарную активность, а КСФ- М — бактерицидную и тумороцидную (разрушающую опухолевые клетки) функции моноцитов и макрофагов.
referatwork.ru