 Здесь можно сделать предварительный вывод актуальности фотосинтеза: источником энергии практически всех метаболических процессов на Земле, служит Солнце, а сам процесс фотосинтеза нужен для поддержания всевозможных форм жизни на планете.
Здесь можно сделать предварительный вывод актуальности фотосинтеза: источником энергии практически всех метаболических процессов на Земле, служит Солнце, а сам процесс фотосинтеза нужен для поддержания всевозможных форм жизни на планете.Растения превращают солнечный свет в запасенную химическую энергию в два этапа: сначала они улавливают энергию солнечного света, а затем используют ее для связывания углерода с образованием органических молекул.
Зеленые растения — биологи называют их автотрофами — основа жизни на планете. С растений начинаются практически все пищевые цепи. Они превращают энергию, падающую на них в форме солнечного света, в энергию, запасенную в углеводах, из которых важнее всего шестиуглеродный сахар глюкоза. Этот процесс преобразования энергии называется фотосинтезом. Другие живые организмы получают доступ к этой энергии, поедая растения. Так создается пищевая цепь, поддерживающая планетарную экосистему.
Кроме того, воздух, которым мы дышим, благодаря фотосинтезу насыщается кислородом. Суммарное уравнение фотосинтеза выглядит так:
вода + углекислый газ + свет —> углеводы + кислород
Растения поглощают углекислый газ, образовавшийся при дыхании, и выделяют кислород — продукт жизнедеятельности растений. К тому же, фотосинтез играет важнейшую роль в круговороте углерода в природе.
В начале XVII в. фламандский врач Ван Гельмонт вырастил в кадке с землей дерево, которое он поливал только дождевой водой. Он заметил, что спустя пять лет, дерево выросло до больших размеров, хотя количество земли в кадке практически не уменьшилось. Ван Гельмонт, естественно, сделал вывод, что материал, из которого образовалось дерево произошел из воды, использованной для полива. В 1777 английский ботаник Стивен Хейлс опубликовал книгу, в которой сообщалось, что в качестве питательного вещества, необходимого для роста, растения используют главным образом воздух. В тот же период знаменитый английский химик Джозеф Пристли (он был одним из первооткрывателей кислорода) провел серию опытов по горению и дыханию и пришел к выводу о том, что зелёные растения способны совершать все те дыхательные процессы, которые были обнаружены в тканях животных. Пристли сжигал свечу в замкнутом объеме воздуха, и обнаруживал, что получавшийся при этом воздух уже не может поддерживать горение. Мышь, помещенная в такой сосуд, умирала. Однако веточка мяты продолжала жить в воздухе неделями. В заключение Пристли обнаружил, что в воздухе, восстановленном веточкой мяты, вновь стала гореть свеча, могла дышать мышь. Теперь мы знаем, что свеча, сгорая, потребляла кислород из замкнутого объема воздуха, но затем воздух снова насыщался кислородом благодаря фотосинтезу, происходившему в оставленной веточке мяты. Спустя несколько лет голландский врач Ингенхауз обнаружил, что растения окисляют кислород лишь на солнечном свету и что только их зеленые части обеспечивают выделение кислорода. В 1817 г. два французских химика, Пельтье и Каванту, выделили из листьев зеленое вещество и назвали его хлорофиллом. Следующей важной вехой в истории изучения фотосинтеза было сделанное в 1845 г. немецким физиком Робертом Майером утверждение о том, что зеленые растения преобразуют энергию, солнечного света в химическую энергию.
В течение тысячелетий люди считали, что питается растение исключительно благодаря корням, поглощая с их помощью все необходимые вещества из почвы. Проверить эту точку зрения взялся в начале девятнадцатого века голландский натуралист Ян Ван Гельмонт. Он взвесил землю в горшке и посадил туда побег ивы. В течение пяти лет он поливал деревце, а затем высушил землю и взвесил её и растение. Ива весила семьдесят пять килограмм, а вес земли изменился всего на несколько сот граммов. Вывод учёного был таков – растения получают питательные вещества, прежде всего, не из почвы, а из воды.
На два столетия в науке утвердилась теория водного питания растений. Листья, по этой теории, лишь помогали растению испарять излишнюю влагу.
К самому неожиданному, но правильному предположению о воздушном питании растений ученые пришли лишь к началу девятнадцатого века. Важную роль в понимании этого процесса сыграло открытие, совершенное английским химиком Джозефом Пристли в 1771 году. Он поставил опыт, в результате которого он сделал вывод: растения очищают воздух и делают его пригодным для дыхания. Позднее выяснилось: для того, чтобы растение очищало воздух, необходим свет.
Десять лет спустя, учёные поняли, что растение не просто превращает углекислый газ в кислород. Углекислый газ необходим растениям для жизни, он служит для них настоящей пищей (вместе с водой и минеральными солями).
Воздушное питание растений называется фотосинтезом. Кислород в процессе фотосинтеза выделяется в качестве необычного продукта.
Миллиарды лет назад на земле не было свободного кислорода. Весь кислород, которым дышат почти все живые существа нашей планеты, выделен растениями в процессе фотосинтеза. Фотосинтез сумел изменить весь облик нашей планеты!
Начиная с семидесятых годов прошлого столетия, крупные успехи в области фотосинтеза были получены в России. Работами русских учёных Пуриевича, Ивановского, Риктера, Иванова, Костычева были изучены многие стороны этого процесса .
Значение фотосинтеза не осознавалось до сравнительно недавнего времени. Аристотель и другие учёные Греции, наблюдая, что жизненные процессы животных зависят от потребления пищи, полагали, что растения добывают свою «пищу» из почвы.
Немногим более трехсот лет назад в одном из первых тщательно продуманных биологических экспериментов голландский врач Ян Ван Гельмонт представил доказательства того, что не одна почва кормит растение. Ван Гельмонт выращивал маленькое дерево ивы в глиняном горшке, добавляя в него только воду.
Через пять лет масса игл увеличилась на 74,4 кг, в то время, как масса почвы уменьшилась только на 57 гр.
В конце XVIII века английский ученый Джозеф Пристли сообщил, что он «случайно обнаружил метод исправления воздуха, который был испорчен горением свечей». 17 августа 1771 г. Пристли «… поместил живую веточку мяты в закрытый сосуд, в котором горела восковая свеча», а 21 числа того же месяца обнаружил, что «… другая свеча снова могла гореть в этом же сосуде». «Исправляющим началом, которым для этих целей пользуется природа, – полагал Пристли, – было растение». Он расширил свои наблюдения и скоро показал, что воздух, «исправляемый» растением, не был «совсем не подходящим для мыши».
Опыты Пристли впервые позволили объяснить, почему воздух на Земле остается «чистым» и может поддерживать жизнь, несмотря на горение бесчисленных огней и дыхание множества живых организмов. Он говорил: «Благодаря этим открытиям мы уверены, что растения произрастают не напрасно, а очищают и облагораживают нашу атмосферу».
Позднее голландский врач Ян Ингенхауз (1730-1799) подтвердил работу Пристли и показал, что воздух «исправляется» только на солнечном свету и только зелеными частями растения. В 1796 году Ингенхауз предположил, что углекислота разлагается при фотосинтезе на С и О2, а О2 выделяется в виде газа. В последствие было обнаружено, что соотношение атомов углерода, водорода и кислорода в сахарах и крахмале таково, что один атом углерода приходится на одну молекулу воды, на что и указывает слово «углеводы». Считалось общепринятым, что углеводы образуются из С и Н2О, а О2 выделяется из углекислоты. Это вполне разумная гипотеза была широко признана, но, как позднее выяснилось, она была совершенно неверной.
Исследователем, который опроверг эту общепринятую теорию, был Корнелиус ван Ниль из Стамфордского университета, когда он, будучи еще студентом – дипломником, исследовал метаболизм различных фотосинтезирующих бактерий. Одна группа таких бактерий, а именно пурпурные серные бактерии, восстанавливает С до углеводов, но не выделяет О2. Пурпурным серным бактериям для фотосинтеза необходим сероводород. В результате фотосинтеза внутри бактериальных клеток накапливаются частицы серы. Ван Ниль обнаружил, что для этих бактерий уравнение фотосинтеза может быть записано как:
свет
С О2 + 2Н2S (Ch3O) + Н2О + 2S
Этот факт не привлекал внимания исследователей до тех пор, пока ван Ниль не сделал смелого сообщения и не предложил следующего суммарного уравнения фотосинтеза:
свет
С О2 + 2Н2А (Ch3O) + Н2О + 2А
В этом уравнении Н2А представляет собой либо воду, либо другое окисляемое вещество, например, сероводород или свободный Н2. У зеленых растений и водорослей Н2А = Н2О. То есть ван Ниль предположил, что Н2О, а не углекислота, разлагается при фотосинтезе. Эта блестящая идея, выдвинутая в тридцатые годы, экспериментально была доказана позднее, когда исследователи, использую тяжелый изотоп О2(18О2), проследили путь кислорода от воды до газообразного состояния:
свет
С О2 + 2Н218О2 (Ch3O) + Н2О + 18О2
Таким образом, для водорослей или зеленых растений, у которых вода служит донором электронов, суммарное уравнение фотосинтеза записывается следующим образом:
свет
6СО2 + 12Н2О C6h22O6 + 6О2 + 6Н2О
Хлоропласты в клетках листа
Фотосинтез растений осуществляется в хлоропластах: обособленных двухмембранных органеллах клетки. Хлоропласты могут быть в клеткахплодов, стеблей, однако основным органом фотосинтеза, анатомически приспособленным к его ведению, является лист. В листе наиболее богата хлоропластами ткань палисадной паренхимы. У некоторых суккулентов с вырожденными листьями (например, кактусы) основная фотосинтетическая активность связана со стеблем.
Свет для фотосинтеза захватывается более полно благодаря плоской форме листа, обеспечивающей большое отношение поверхности к объёму. Вода доставляется из корня по развитой сети сосудов (жилок листа). Углекислый газ поступает отчасти посредством диффузии через кутикулу иэпидермис, однако большая его часть диффундирует в лист через устьица и по листу по межклеточному пространству. Растения, осуществляющие С4 и CAM фотосинтез, сформировали особые механизмы для активной ассимиляции углекислого газа.
Внутреннее пространство хлоропласта заполнено бесцветным содержимым (стромой) и пронизано мембранами (ламеллами), которые соединяясь друг с другом образуют тилакоиды, которые в свою очередь группируются в стопки, называемые граны. Внутритилакоидное пространство отделено и не сообщается с остальной стромой, предполагается также что внутреннее пространство всех тилакоидов сообщается между собой. Световые стадии фотосинтеза приурочены к мембранам, автотрофная фиксация CO2 происходит в строме.
В хлоропластах имеются свои ДНК, РНК, рибосомы (70s типа), идёт синтез белка (хотя этот процесс и контролируется из ядра). Они не синтезируются вновь, а образуются путём деления предшествующих. Всё это позволило считать их потомками свободных цианобактерий, вошедших в состав эукариотической клетки в процессе симбиогенеза.
Цианобактерии, таким образом, как бы сами являются хлоропластом и в их клетке фотосинтетический аппарат не вынесен в особую органеллу. Их тилакоиды, однако, не образуют стопок, а формируют различные складчатые структуры (у единственной цианобактерии Gloeobacter violaceus тилакоиды отсутствуют вовсе, а весь фотосинтетический аппарат находится в цитоплазматической мембране, не образующей впячиваний). У них и растений также есть различия в светособирающем комплексе (см. ниже) и пигментном составе.
Процессы, происходящие в листе
Лист осуществляет три важных процесса – фотосинтез, испарение воды и газообмен. В процессе фотосинтеза в листьях из воды и двуокиси углерода под действием солнечных лучей синтезируются органические вещества. Днем, в результате фотосинтеза и дыхания, растение выделяет кислород и двуокись углерода, а ночью – только двуокись углерода, образующуюся при дыхании.
Большинство растений способно синтезировать хлорофилл при слабом освещении. При прямом солнечном освещении хлорофилл синтезируется быстрее.
Необходимая для фотосинтеза световая энергия в известных пределах поглощается тем больше, чем меньше затемнен лист. Потому у растений в процессе эволюции выработалась способность поворачивать пластину листа к свету так, чтобы на нее падало больше солнечных лучей. Листья на растении располагаются так, чтобы не притеснять друг друга.
Тимирязев доказал, что источником энергии для фотосинтеза служат преимущественно красные лучи спектра. На это указывает спектр поглощения хлорофилла, где наиболее интенсивная полоса поглощения наблюдается в красной, и менее интенсивное – в сине-фиолетовой части.
В хлоропластах вместе с хлорофиллом имеются пигменты каротин и ксантофилл. Оба этих пигмента поглощают синие и, отчасти, зеленые лучи и пропускают красные и желтые. Некоторые ученые приписываю каротину и ксантофиллу роль экранов, защищающих хлорофилл от разрушительного действия синих лучей.
Процесс фотосинтеза слагается из целого ряда последовательных реакций, часть которых протекает с поглощением световой энергии, а часть – в темноте. Устойчивыми окончательными продуктами фотосинтеза являются углеводы (сахара, а затем крахмал), органические кислоты, аминокислоты, белки.
Фотосинтез при различных условиях протекает с разной интенсивностью.
Интенсивность фотосинтеза также зависит от фазы развития растения. Максимальная интенсивность фотосинтеза наблюдается в фазе цветения.
Обычное содержание углекислоты в воздухе составляет 0,03% по объему. Уменьшение содержания углекислоты в воздухе снижает интенсивность фотосинтеза. Повышение содержания углекислоты до 0,5% увеличивает интенсивность фотосинтеза почти пропорционально. Однако при дальнейшем повышении содержания углекислоты, интенсивность фотосинтеза не возрастает, а при 1% – растение страдает.
Растения испаряют или трансперируют очень большое количество воды. Испарение воды является одной из причин восходящего тока. Вследствие испарения воды растением в нем накапливаются минеральные вещества, и происходит полезное для растения понижение температуры во время солнечного нагрева. Иногда трансперация снижает температуру растения на 6о.
Растение регулирует процесс испарения воды посредством работы устьиц. Отложение кутикулы или воскового налета на эпидерме, образование его волосков и другие приспособления направлены к сокращению нерегулируемой трансперации.
Процесс фотосинтеза и постоянное протекающее дыхание живых клеток листа требуют газообмена между внутренними тканями листа и атмосферой. В процессе фотосинтеза из атмосферы поглощается ассимилируемый углекислый газ и возвращается в атмосферу кислородом.
Применение изотопного метода анализа показало, что кислород, возвращаемый в атмосферу (16О) принадлежит воде, а не углекислому газу воздуха, в котором преобладает другой его изотоп – 15О. При дыхании живых клеток (окисление свободным кислородом органических веществ внутри клетки до углекислого газа и воды) необходимо поступление из атмосферы кислорода и возвращение углекислоты. Этот газообмен также в основном осуществляется через устьичный аппарат.
Еще в 1905 г. английский физиолог растений Ф. Ф. Блекмэн, интерпретируя форму кривой светового насыщения фотосинтеза, высказал предположение, что фотосинтез представляет собой двухстадийный процесс, включающий фотохимическую, т.е. светочувствительную реакцию и нефотохимическую, т. е. темновую, реакцию. Темновая реакция, будучи ферментативной, протекает медленнее, чем световая реакция, и поэтому при высоких интенсивностях света скорость фотосинтеза полностью определяется скоростью темновой реакции. Световая реакция либо вообще не зависит от температуры, либо зависимость эта выражена очень слабо, тогда темновая реакция, как и все ферментативные процессы, зависит от температуры в довольно значительно и степени. Следует ясно представлять себе, что реакция, называемая темновой, может протекать как в темноте, так и на свету. Световую и темновую реакции можно разделить, используя вспышки света, длящиеся краткие доли секунды. Вспышки света длительностью меньше одной миллисекунды (10-3 с) можно получить либо с помощью механического приспособления, поставив на пути пучка постоянного света вращающийся диск со щелью, либо электрически, заряжая конденсатор и разряжая его через вакуумную или газоразрядную лампу. В качестве источников света пользуются также рубиновыми лазерами с длиной волны излучения 694 нм. В 1932 г. Эмерсон и Арнольд освещали суспензию клеток вспышками света от газоразрядной лампы с длительностью около 10-3с. Они измеряли скорость выделения кислорода в зависимости от энергии вспышек, длительности темнового промежутка между вспышками и температуры суспензии клеток. При увеличении интенсивности вспышек насыщение фотосинтеза в нормальных клетках наступало, когда выделялась одна молекула 02 на 2500 молекул хлорофилла. Эмерсон и Арнольд сделали вывод, что максимальный выход фотосинтеза определяется не числом молекул хлорофилла, поглощающих свет, а числом молекул фермента, катализирующего темновую реакцию. Они также обнаружили, что при увеличении темновых интервалов между последовательными вспышками за пределы 0,06 с выход кислорода в расчете на одну вспышку уже не зависел от длительности темнового интервала, тогда как при более коротких промежутках он возрастал с увеличением длительности темнового интервала (от 0 до 0,06 с). Таким образом, темновая реакция, которая определяет уровень насыщения фотосинтеза, завершается примерно за 0,06 с. На основе этих данных было рассчитано, что среднее время, характеризующее скорость реакции, составило около 0,02 с при 25°С.
Лимитирующие факторы
Интенсивность, или скорость процесса фотосинтеза в растении зависит от ряда внутренних и внешних факторов. Из внутренних факторов наиболее важное значение имеют структура листа и содержание в нем хлорофилла, скорость накопления продуктов фотосинтеза в хлоропластах, влияние ферментов, а также наличие малых концентраций необходимых неорганических веществ. Внешние параметры – это количество и качество света, попадающего на листья, температура окружающей среды, концентрация углекислоты и кислорода в атмосфере вблизи растения.
Скорость фотосинтеза возрастает линейно, или прямо пропорционально увеличению интенсивности света. По мере дальнейшего увеличения интенсивности света нарастание фотосинтеза становится все менее и менее выраженным, и, наконец, прекращается, когда освещенность достигает определенного уровня 10000 люкс. Дальнейшее увеличение интенсивности света уже не влияет на скорость фотосинтеза. Область стабильной скорости фотосинтеза называется областью светонасыщения. Если нужно увеличить скорость фотосинтеза в этой области, следует изменять не интенсивность света, а какие-либо другие факторы. Интенсивность солнечного света, попадающего в ясный летний день на поверхность земли, во многих местах нашей планеты составляет примерно 100000 люкс. Следовательно, растениям, за исключением тех, которые растут в густых лесах и в тени, падающего солнечного света бывает достаточно для насыщения их фотосинтетической активности (энергия квантов, соответствующих крайним участкам видимого диапазона – фиолетового и красного, различается всего лишь в два раза, и все фотоны этого диапазона в принципе способны осуществить запуск фотосинтеза).
В случае низких интенсивностей света скорость фотосинтеза при 15 и 25°С одинакова. Реакции, протекающие при таких интенсивностях света, которые соответствуют области лимитирования света, подобно истинным фотохимическим реакциям, не чувствительны к температурам. Однако при более высоких интенсивностях скорость фотосинтеза при 25°С гораздо выше, чем при 15°С. Следовательно, в области светового насыщения уровень фотосинтеза зависит не только от поглощения фотонов, но и от других факторов. Большинство растений в умеренном климате хорошо функционируют в интервале температур от 10 до 35°С, наиболее благоприятные условия – это температура около 25°С.
В области лимитирования светом скорость фотосинтеза не изменяется при уменьшении концентрации СО2 . Отсюда можно сделать вывод, что С02 участвует непосредственно в фотохимической реакции. В то же время при более высоких интенсивностях освещения, лежащих за пределами области лимитирования, фотосинтез существенно возрастает при увеличении концентрации СО2. У некоторых зерновых культур фотосинтез линейно возрастал при увеличении концентрации СО2 до 0,5%. (эти измерения проводили в кратковременных опытах, поскольку длительное воздействие высоких концентраций СО2 повреждает листы). Высоких значений скорость фотосинтеза достигает при содержании С02 около 0,1%. Средняя концентрация углекислоты в атмосфере составляет от 0,03%. Поэтому в обычных условиях растениям не хватает СО2 для того, чтобы с максимальной эффективностью использовать попадающий на них солнечный свет. Если помещенное в замкнутый объем растение освещать светом насыщающей интенсивности, то концентрация СО2 в объеме воздуха будет постепенно уменьшаться и достигнет постоянного уровня, известного под названием «С02 компенсационного пункта». В этой точке появление СО2 при фотосинтезе уравновешивается выделением О2 в результате дыхания (темнового и светового). У растений разных видов положения компенсационных пунктов различны.
Наряду с фотосинтезом высших растений и водорослей, сопровождаемым выделением O2, в природе осуществляется бактериальный фотосинтез, в котором окисляемым субстратом является не вода, а другие соединения, обладающие более выраженными восстановительными свойствами, например h3S, SO2. Кислород при бактериальном фотосинтезе не выделяется, например:
Фотосинтезирующие бактерии способны использовать не только видимое, но и ближнее ИК излучение (до 1000 нм) в соответствии со спектрами поглощения преобладающих в них пигментов – бактериохлорофиллов. Бактериальный фотосинтез не имеет существенного значения в глобальном запасании солнечной энергии, но важен для понимания общих механизмов фотосинтеза Кроме того, локально бескислородный фотосинтез может вносить существенный вклад в суммарную продуктивность планктона. Так, в Черном море количество хлорофилла и бактериохлорофилла в столбеводы в ряде мест приблизительно одинаково.
Единственный известный в природе нехлорофилльный способ запасания энергии света осуществляют бактерииHalobacterium halobium. Ha ярком свету при пониженной концентрации O2 они образуют в своих мембранах пурпурный белок бактериородопсин. В результате индуцированной светом цис-транс-изомеризации ретиналя (хромофора этого пигмента) происходит поглощение H+ и синтез АТФ. Последний используется для частичного обеспечения энергетических потребностей клетки.
В настоящее время известно, что фотосинтез проходит две стадии, но только одна из них – на свету. Доказательства двухстадийности процесса впервые были получены в 1905 году английским физиологом растений Ф.Ф. Блэклином, который исследовал влияние освещенности и температуры на объем фотосинтеза.
На основании экспериментов, Блэклин сделал следующие выводы.
1. Имеется одна группа светозависимых реакций, которые не зависят от температуры. Объем этих реакций в диапазоне низких освещенностей мог возрастать с увеличением освещенности, но не с увеличением температуры.
2. Имеется вторая группа реакций, зависимых от температуры, а не от света. Оказалось, что обе группы реакций необходимы для осуществления фотосинтеза. Увеличение объема только одной группы реакций увеличивает объем всего процесса, но только до того момента, пока вторая группа реакций не начнет удерживать первую. После этого необходимо ускорить вторую группу реакций, чтобы первые могли проходить без ограничений.
Таким образом, было показано, что обе стадии светозависимы: «световая и темновая». Важно помнить, что темновые реакции нормально проходят на свету и нуждаются в продуктах световой стадии. Выражение «темновые реакции» просто означает, что свет как таковой в них не участвует.
Объем темновых реакций возрастает с увеличением температуры, но только до 30о, а затем начинает падать. На основании этого факта предположили, что темновые реакции катализируются ферментами, поскольку обмен ферментативных реакций, таким образом, зависит от температуры. В последствие оказалось, что данный вывод был сделан неправильно.
На первой стадии фотосинтеза (световые реакции) энергия света используется для образования АТР (молекула аденозин-трифосфата) и высокоэнергетических переносчиков электронов. На второй стадии фотосинтеза (темновые реакции) энергетические продукты, образовавшиеся в световых реакциях, используются для восстановления СО2 до простого сахара (глюкозы).
Процесс фотосинтеза все больше и больше привлекает к себе внимание ученых. Наука близка к разрешению важнейшего вопроса – искусственного создания при помощи световой энергии ценных органических веществ из широко распространенных неорганических веществ. Проблема фотосинтеза усиленно разрабатывается ботаниками, химиками, физиками и другими специалистами.
В последнее время уже удалось искусственно получить синтез формальдегида и сахаристых веществ из водных растворов карбонатной кислоты; при этом роль поглотителя световой энергии играли вместо хлорофилла карбонаты кобальта и никеля. Недавно синтезирована молекула хлорофилла.
Успехи науки в области синтеза органических веществ наносят сокрушительный удар по идеалистическому учению – витализму, который доказывал, что для образования органических веществ из неорганических необходима особая «жизненная сила» и что человек не сможет синтезировать сложные органические вещества.
Фотосинтез в растениях осуществляется в хлоропластах. Он включает преобразования энергии (световой процесс), превращение вещества (темновой процесс). Световой процесс происходит в гилакоидах, темновой – в строме хлоропластов. Обобщенное циркулирование фотосинтеза выглядит следующим образом:
свет
6СО2 + 12Н2О C6h22O6 + 6Н2О + 6О2
Два процесса фотосинтеза выражаются отдельными уравнениями
свет
12Н2О 12h3 + 6О2 + энергия АТР
(световой процесс)
свет
12h3 + 6О2 + энергия АТР С6Н12О6 + Н2О
(темновой процесс)
Фотосинтез – единственный процесс в биосфере, ведущий к увеличению ее свободной энергии за счет внешнего источника. Запасенная в продуктах фотосинтеза энергия – основной источник энергии для человечества.
Ежегодно в результате фотосинтеза на Земле образуется 150 млрд. тонн органического вещества и выделяется около 200 млн. тонн свободного кислорода.
Круговорот кислорода, углерода и других элементов, вовлекаемых в фотосинтез, поддерживает современный состав атмосферы, необходимый для жизни на Земле. Фотосинтез препятствует увеличению концентрации СО2, предотвращая перегрев Земли вследствие так называемого «парникового эффекта».
Поскольку зеленые растения представляют собой непосредственную или опосредованную базу питания всех других гетеротрофных организмов, фотосинтез удовлетворяет потребность в пище всего живого на нашей планете. Он – важнейшая основа сельского и лесного хозяйства. Хотя возможности воздействия на него еще не велики, но все же и они, в какой то мере используются. При повышении концентрации углекислого газа в воздухе до 0,1% (против 0,3% в естественной атмосфере) удалось, например, повысить урожайность огурцов и томатов втрое.
Квадратный метр поверхности листьев в течение одного часа продуцирует около одного грамма сахара; это значит, что все растения, по приблизительной оценке, изымают из атмосферы от 100 до 200 млрд. тонн С в год. Около 60% этого количества поглощают леса, занимающие 30% непокрытой льдами поверхности суши, 32% – окультуренные земли, а оставшиеся 8% – растения степей и пустынных мест, а также городов и поселков.
Зеленое растение способно не только использовать углекислый газ и создавать сахар, но и превращать азотные соединения, и соединения серы в вещества, слагающие его тело. Через корневую систему растение получает растворенные в почвенной воде ионы нитратов и перерабатывает их в своих клетках в аминокислоты – основные компоненты всех белковых соединений. Компоненты жиров также возникают из соединений, образующихся в процессах обмена веществ и энергии. Из жирных кислот и глицерина возникают жиры и масла, которые служат для растения, главным образом, запасными веществами. В семенах приблизительно 80% всех растений, в качестве богатого энергией запасного вещества, содержатся жиры. Получение семян, жиров и масел играет важную роль в сельскохозяйственной и пищевой промышленности.
Прочие факты
Заключение
Во второй половине XIX столетия было установлено, что энергия солнечного света усваивается и трансформируется при помощи зеленого пигмента хлорофилла.
На основе проведенных опытов можно сказать что, зеленая окраска хлорофилла определяется наличием в нем атома металла вне зависимости от того, будет ли это магний, медь или цинк.
Современная наука подтвердила правильность взглядов К. А. Тимирязева относительно исключительной важности для фотосинтеза именно красных лучей солнечного спектра. Оказалось, что коэффициент использования красного света в ходе фотосинтеза выше, чем синих лучей, которые также поглощаются хлорофиллом. Красные лучи, по представлениям К. А. Тимирязева, играют основополагающую роль в процессе мироздания и созидания жизни.
Как известно растения поглащают углекислый газ, который присоединяется к пятиуглеродному веществу под названием рибулезодифосфат, где потом он в дальнешем участвует во многих других реакциях.
Изучение особенностей фотосинтеза у разных растений, безусловно, будет способствовать расширению возможностей человека в управлении их фотосинтетической деятельностью, продуктивностью и урожаем. В целом фотосинтез это один из основополагающих процессов жизни, на котором основана большая часть современной растительной фауны на поверхности земли.
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
kursak.net
Фотосинтез — это процесс, при котором световая энергия поглощается и используется на синтез восстановленных углеродсодержащих соединений из двуокиси углерода и воды. Этот процесс происходит только в освещенной зеленой ткани, потому что хлорофилл играет существенную роль в превращении световой энергии в химическую. Термин фотосинтез был предложен Чарльзом Рейдом Бансом из Чикагского университета в начале этого столетия. В Европе этот процесс часто называют ассимиляцией или ассимиляцией углерода. Большинство американских физиологов растений предпочитают употреблять термин ассимиляция, когда речь идет об образовании новых тканей из углеводов и азотистых соединений.
Значение фотосинтеза нельзя переоценить, потому что вся энергия, содержащаяся в нашей пище, накапливается прямо или косвенно благодаря процессу фотосинтеза, а источником большей части используемой энергии, на которой работают заводы, являются горючие ископаемые, где она была запасена посредством фотосинтеза в далеком прошлом. Лит (1972, 1975) подсчитал, что наземные растения образуют за год 100-109 т сухого вещества, из которых более 2/3 вырабатывают деревья. Общая энергия, ежегодно накапливаемая в растениях при фотосинтезе, приблизительно в 100 раз превышает энергию, содержащуюся в угле, добываемом в течение года во всех шахтах мира. Хотя леса и занимают только 1/3 поверхности суши земного шара, они вырабатывают около 2/3 сухого вещества, в то время как возделываемые земли занимают приблизительно 9% и дают только 9% сухого вещества.
Пока человек не стал широко использовать уголь, большая часть потребляемой им тепловой энергии получалась от сжигания древесины. В настоящее время древесина шире используется как строительный материал и источник получения бумаги, чем в качестве топлива, хотя интерес к последнему опять возрастает. Независимо от того, как используется древесина, необходимо помнить, что содержащиеся в ней энергия и сухое вещество накапливаются благодаря процессу фотосинтеза. В связи с этим рациональное ведение лесного хозяйства должно быть направлено на повышение количества продуктов фотосинтеза на единицу земной поверхности и эффективности их превращения в растительный материал.
Из общего количества солнечного излучения, попадающего на нашу планету, лишь половина доходит до поверхности Земли, только 1/8 имеет длину волны, подходящую для фотосинтеза, и лишь 0,4 % таких лучей (около 1 % от общего объёма энергии) используется растениями. Именно от этого одного процента зависит вся жизнь на Земле.
В процессе фотосинтеза углекислый газ в присутствии хлорофилла реагирует с водой; при этом образуется глюкоза и выделяется кислород:
6CO2 + 6h3 O → C6 h22 O6 + 6O2.
Более грамотной будет запись
CO2 + 2h3 O → [Ch3 O] + O2 + h3 O,
Которая показывает, что выделяющийся кислород образуется из воды. Похожим уравнением описывается и хемосинтез серобактерий:
CO2 + 2h3 S → [Ch3 O] + 2S + h3 O,
www.ebio.ru/images/10020201.gif
Общая схема фотосинтеза
Таким образом, процесс фотосинтеза включает в себя две стадии:
— получение водорода (фотолиз) – при этом кислород выделяется как побочный продукт реакции;
— получение глюкозы (восстановление).
www.ebio.ru/images/10020202.jpg
Газообмен в листе происходит через устьица
www.ebio.ru/images/10020203.gif
Интенсивность фотосинтеза на различных длинах волн
Первая стадия фотосинтеза протекает на свету. Световые кванты дают электронам энергию, необходимую для переноса их от хлорофилла или другого фотосинтезирующего пигмента. В ходе первой стадии из АДФ (аденозиндифосфата) и фосфата синтезируется АТФ (аденозинтрифосфат), а НАДФ (никотинамидадениндинуклеотидфосфат) восстанавливается до НАДФ∙h3. Синтез АТФ за счёт энергии световых квантов называется фотофосфорилированием. Этот процесс может быть циклическим (в реакции «работают» одни и те же электроны) и нециклическим (электроны в конце концов доходят до НАДФ и, взаимодействуя с ионами водорода, образуют НАДФ∙h3 ). Кислород как побочный продукт реакции выделяется только во втором случае.
Для реакций второй стадии свет не нужен. Восстановление CO2 происходит за счет энергии АТФ и накопленного НАДФ∙h3. Углекислый газ связывается с пятиуглеродным сахаром рибулозобисфосфатом, образуя две молекулы трёхуглеродной фосфоглицериновой кислоты (ФГК). Такой процесс получил название C3 -фотосинтеза. Последующий цикл реакций (цикл Кальвина) приводит к образованию из ФГК сахара (например, глюкозы), а также ресинтезу рибулозобисфосфата. У некоторых растений (например, сахарного тростника, сои) наблюдается так называемый C4 -фотосинтез, в реакциях которого CO2, восстанавливаясь, включается в состав органических кислот, имеющих четыре атома углерода (например, яблочной). При этом поглощение углекислоты идёт гораздо эффективнее, повышается и продуктивность растений.
На скорость фотосинтеза влияют многие факторы. Основными из них являются интенсивность света, концентрация кислорода и углекислого газа, температура окружающей среды. Состояние, когда скорость выделения кислорода растением равна скорости его дыхания, называется точкой компенсации.
Кислород в процессе фотосинтеза может действовать как конкурентный ингибитор, взаимодействуя с рибулозодисфосфатом вместо углекислого газа. При этом образуется одна молекула ФГК и фосфогликолат, сразу расщепляющийся до гликолата. Чтобы вернуть хотя бы часть углерода, связанного в бесполезном гликолате, у растения имеется процесс, называемый фотодыханием. Это зависимое от света потребление кислорода с выделением углекислого газа, заметное лишь у C3 -растений, не имеет ничего общего с обычным дыханием. Фотодыхание, в целом, идёт с поглощением энергии; в результате образуется фосфоглицерат, а 25 % углерода теряется в виде CO2. В фотодыхании участвуют хлоропласты, пероксисомы и митохондрии. У C4 -растений фотодыхания практически нет, что и является причиной их большей продуктивности.
В связи с энергетической проблемой учёные пытаются провести фотосинтетические процессы искусственно, особенно их первые этапы, когда вода под действием солнечной радиации расщепляется на кислород и водород. Сжигание водорода (с образованием воды) – экологически чистый процесс, который мог бы стать неплохой заменой современным источникам энергии
ПИГМЕНТЫ ХЛОРОПЛАСТОВ
Хлорофилл заслуживает особого внимания, потому что в процессе фотосинтеза он является светопоглощающим пигментом, а также и потому, что создает доминирующую окраску земной поверхности. У некоторых декоративных деревьев и кустарников иногда бывают видны желтые пигменты-каротиноиды. Эти пигменты обнаруживаются и вследствие того, что условия становятся неблагоприятными для синтеза хлорофилла или его сохранения. Листья некоторых разновидностей древесных растений, например, темно-пунцовой формы бука европейского, клена дланевидного, имеют красную или пурпурную окраску из-за присутствия в клеточном соке (а не в пластидах) антоцианов. Многие другие деревья вырабатывают антоцианы осенью. (Образование осенней окраски листьев будет рассмотрено в главе 7). Хлорофилл и каротиноиды встречаются, кроме листовых пластинок, во многих тканях, в том числе в черешках, почках, семядолях, в коровой паренхиме молодых побегов и в феллодерме более старых стеблей некоторых видов. Обычно хлоропласты редко встречаются в эпидермальных клетках, за исключением замыкающих клеток устьиц. Изредка они развиваются в корнях, выставленных на свет.
Интенсивность фотосинтеза листьев древесных растений, как правило, ниже интенсивности фотосинтеза травянистых растений.
ФАКТОРЫ, ВЛИЯЮЩИЕ НА ОБРАЗОВАНИЕ ХЛОРОФИЛЛА
Пожелтение, или хлороз листьев — результат их неспособности увеличивать или сохранять содержание хлорофилла. Всесторонним изучением установлено, что это явление зависит от ряда внутренних и внешних факторов
ВНУТРЕННИЕ ФАКТОРЫ
Наиболее важный фактор — генетический потенциал растения, так как иногда в результате мутаций, приводящих к полной потере способности образовывать хлорофилл, возникают ко-роткоживущие альбиносные сеянцы. Чаще процесс синтеза хлорофилла нарушается лишь частично, проявляясь в отсутствии хлорофилла в некоторых участках листьев (пестролистность) или в общем низком содержании хлорофилла. Вследствие этого листья принимают желтоватый оттенок золотистых разновидностей деревьев и кустарников, часто используемых в качестве декоративных растений. Иногда в результате почковых мутаций возникают альбиносные или пестролистные ветви на нормальных в других отношениях деревьях и кустарниках. В синтезе хлорофилла и сборке его в фотосинтетические единицы участвуют многие гены, поэтому в образовании хлорофилла распространены аномалии. Развитие хлоропластов зависит как от ядерной, так и от плас-тидной ДНК, а также от цитоплазматических и хлоропластных рибосом.
Достаточное снабжение углеводами необходимо, по-видимому, для образования хлорофилла, и листья, испытывающие недостаток растворимых углеводов, могут не зеленеть, даже если все прочие условия благоприятные. Такие листья, опущенные в раствор сахара, обычно начинают образовывать хлорофилл. Часто образованию хлорофилла препятствуют вирусы, вызывая «желтуху», характеризующуюся общим хлорозом или бронзовой окраской листовых пластинок, а также просветлением жилок. Крапчатые листья некоторых декоративных растений — результат вирусной инфекции.
ФАКТОРЫ ВНЕШНЕЙ СРЕДЫ
Главными внешними факторами, влияющими на образование и сохранение хлорофилла, являются: свет, температура, минеральное питание, вода и кислород. Синтез хлорофилла очень чувствителен почти к любому фактору, нарушающему метаболические процессы.
Свет. Свет необходим для образования хлорофилла, хотя сеянцы некоторых хвойных и небольшое число других видов растений образуют хлорофилл в темноте. Относительно низкая освещенность эффективна для инициации или ускорения образования хлорофилла. Выращенные в темноте желтые сеянцы содержат протохлорофилл — предшественник хлорофилла а, для восстановления которого до хлорофилла а требуется свет. Очень яркий свет вызывает разложение хлорофилла. Следовательно, хлорофилл всегда одновременно синтезируется и разрушается. На ярком свету равновесие устанавливается при более низкой концентрации хлорофилла, чем при свете малой интенсивности. Теневые листья обычно имеют более высокую концентрацию хлорофилла, чем световые.
Температура. Синтез хлорофилла происходит, по-видимому, в широком интервале температур. Вечнозеленые растения умеренной зоны синтезируют хлорофилл от температур близких к температурам замерзания до самых высоких температур в середине лета. Многие хвойные становятся зимой до некоторой степени хлоротичными, вероятно, вследствие того, что распад хлорофилла превышает его синтез при очень низких температурах.
Минеральное питание. Одна из наиболее частых причин хлороза — недостаток какого-либо необходимого элемента. Недостаток азота — обычная причина хлороза древесных растений, особенно у старых листьев. Другая распространенная причина хлороза заключается в недостатке железа, преимущественно у молодых листьев. Достаточное снабжение железом необходимо, очевидно, для синтеза хлорофилла. В состав хлорофилла железо не входит, но оно служит кофактором для предшественника хлорофилла. Магний является составной частью хлорофилла, поэтому его недостаток, естественно, вызывает хлороз. Недостаток большинства макроэлементов и некоторых микроэлементов может быть причиной хлороза. Можно предположить, что почти любое нарушение нормального метаболизма препятствует синтезу хлорофилла. (Хлороз, вызываемый недостатком минеральных элементов, обсуждается более подробно в главе 10).
Вода. Умеренный водный стресс замедляет образование хлорофилла, а сильное обезвоживание растительных тканей не только нарушает синтез хлорофилла, но и вызывает распад уже имеющихся молекул. В результате листья растений, подвергшихся воздействию засухи, имеют тенденцию к пожелтению. Листья деревьев и кустарников могут также пожелтеть при насыщении водой почвы вокруг их корней. Действие засухи и плохой аэрации почвы является до некоторой степени косвенным: синтез хлорофилла задерживается вследствие общего нарушения метаболизма (см. главы 16 и 17).
Кислород. При отсутствии кислорода сеянцы не образуют хлорофилла даже на свету. Это указывает на то, что аэробное дыхание необходимо для некоторых процессов образования промежуточных соединений, что для синтеза хлорофилла необходим приток метаболической энергии.
МЕХАНИЗМ ФОТОСИНТЕЗА
Фотосинтез заключается в восстановлении атмосферной С02 до углеводов с использованием световой энергии, сопровождаемой освобождением кислорода из воды. Фотосинтез, подобно многим другим физиологическим процессам, состоит из нескольких последовательных этапов. Вследствие их сложности подробное обсуждение этих этапов не входит в задачу данной книги.
СВЕТОВЫЕ И ТЕМНОВЫЕ РЕАКЦИИ
Фотосинтез включает две фазы: фотохимическую, для которой требуется свет, и химическую, протекающую в темноте.
Фотохимическая фаза протекает очень быстро (за 0,00001 с). Первичная фотохимическая реакция не зависит от температуры, хотя транспорт электронов — процесс, зависящий от температуры. Темновая фаза происходит более медленно, приблизительно за 0,04 с при 25°С, замедляясь при низких температурах. Интенсивность фотосинтеза ограничивается реакцией, протекающей наиболее медленно. В связи с этим на ярком свету, когда ограничивающей является химическая фаза, процесс фотосинтеза чувствителен к температуре, а при слабой освещенности более вероятно, что ограничивающей становится фотохимическая фаза. При нормальных условиях освещения концентрация С02 чаще бывает ограничивающим фактором, чем температура, за исключением, возможно, вечнозеленых растений зимой. Большой интерес представляет поиск методов повышения эффективности фотосинтеза. Естественный отбор, действовавший в течение тысячелетий, создал согласованность биохимии и биофизики фотосинтеза, которую будет очень трудно улучшить в полевых условиях.
ПОГЛОЩЕНИЕ ДВУОКИСИ УГЛЕРОДА ФОТОСИНТЕЗИРУЮЩИМИ ТКАНЯМИ
С02 поступает преимущественно через устьица, хотя небольшое количество, по-видимому, диффундирует через кутикулу и эпидермальные клетки.
Таким образом, устьица, закрываясь, увеличивают сопротивление передвижению воды гораздо сильнее, чем сопротивление диффузии СО2, и, очевидно, понижают транспирацию в большей степени, чем фотосинтез. Это создает обоснование для использования метаболических антитранспирантов, которые вызывают закрывание устьиц, для уменьшения транспирации (см. главу 13)
ТКАНИ, ОСУЩЕСТВЛЯЮЩИЕ ФОТОСИНТЕЗ
Фотосинтез, происходящий главным образом в листьях, отмечается в некоторой степени и в других зеленых тканях, в том числе в семядолях (см. главу 15), почках, стеблях, цветках и плодах. У большинства видов фотосинтетическая деятельность тканей, не входящих в состав семядолей и листьев, незначительна. Например, фотосинтез зеленых плодов белой акации, лимона, апельсина, авокадо, винограда и сливы обычно слишком слаб для того, чтобы внести заметный вклад в их рост. Фотосинтез быстро разрастающихся зеленых шишек сосны смолистой недостаточен для того, чтобы уравновесить наблюдающееся у них интенсивное выделение СО2 в процессе дыхания. Большая часть необходимых для роста шишек углеводов мобилизуется из других источников.
Фотосинтез коры у большинства видов невелик, составляет 5% общего у тополя дельтовидного, но фотосинтез ветвей и коры очень важен для некоторых растений засушливой зоны. Ряд пустынных кустарников, сбрасывающих листья в сухое время года, имеет устойчивые фотосинтетически активные стволы и ветви. Например, Cercidium microphyllum находится в безлистном состоянии большую часть года, и его зеленые ветви могут вырабатывать до 40 % всех ежегодных продуктов фотосинтеза.
ИЗМЕНЕНИЯ ИНТЕНСИВНОСТИ ФОТОСИНТЕЗА
Интенсивность фотосинтеза древесных растений широко варьирует в зависимости от взаимодействия многих внешних и внутренних факторов, причем эти взаимодействия изменяются во времени и различны у разных видов.
Фотосинтетическую способность иногда оценивают по чистому приросту сухой массы. Такие данные имеют особое значение, потому что прирост представляет собой среднее истинное увеличение массы за большой промежуток времени в условиях внешней среды, включающих обычные периодически наступающие стрессы.
Некоторые виды покрытосеменных эффективно осуществляют фотосинтез как при низкой, так и при высокой интенсивности света. Многие голосеменные гораздо более продуктивны при высокой освещенности. Сравнение этих двух групп при низкой и высокой интенсивности света часто дает различное представление о фотосинтетической способности с точки зрения накопления питательных веществ. Кроме того, голосеменные часто накапливают некоторое количество сухой массы в период покоя, тогда как листопадные покрытосеменные теряют ее вследствие дыхания. Поэтому голосеменное растение с несколько более низкой интенсивностью фотосинтеза, чем листопадное покрытосеменное во время периода роста, может накапливать в течение года столько же или даже больше общей сухой массы благодаря гораздо большей продолжительности периода фотосинтетической активности.
ВИДОВЫЕ И ГЕНЕТИЧЕСКИЕ РАЗЛИЧИЯ
Фотосинтетическая способность часто значительно различается у разных видов, а также среди разновидностей, клонов и особей одного вида, но разного происхождения. Такое разнообразие обычно связано с основными различиями в метаболизме или в анатомической структуре листьев. Кроме того, разные виды и разновидности одного вида заметно различаются по развитию кроны, поэтому образование большего количества листьев или более продолжительный вегетационный период часто компенсируют низкую интенсивность фотосинтеза на единицу листовой поверхности или сухой массы.
Видовые различия. Некоторые исследователи сообщали о различиях в фотосинтетической активности разных видов. Например, среди лесных деревьев особенно высокая интенсивность фотосинтеза наблюдалась у некоторых покрытосеменных (у тополя, яблони, ясеня и эвкалипта) и у голосеменных (таких, как дугласия, лиственница и метасеквойя). Обнаружено, что поглощение СО2 на единицу листовой поверхности значительно выше у дуба красного и белого, чем у дерена флоридского или сосны ладанной. Наблюдалась высокая интенсивность фотосинтеза у дугласии, средняя у сосны веймутовой и низкая у ели европейской. Определено, что метасеквойя, имеющая очень высокую интенсивность фотосинтеза и слабое дыхание, является продуктивнее бука, дуба, сосны, если или лиственницы. Часто интенсивность фотосинтеза заметно различается у видов одного рода. Например, Eucalyptus globulus имеет гораздо более высокую интенсивность, чем Е. marginata. Имеются также сообщения о различиях среди плодовых деревьев. Поглощение двуокиси углерода на единицу поверхности листьев груши и яблони в 3 раза больше, чем у листьев таких вечнозеленых многолетних растений, как цитрусовые. В других исследованиях фотосинтез изменялся в следующем порядке: яблоня > груша > вишня > слива. У листьев яблони фотосинтез был не только наиболее интенсивным, но и дольше сохранялся осенью, а слива была первой, у которой прекращалось поглощение СО2 .
Генетические различия. Имеется много примеров генетических различий в интенсивности фотосинтеза. Например, обнаружены, большие различия в фото синтетической способности двух клонов лиственницы европейской. Установлено, что сеянцы дугласии по происхождению с острова Ванкувер имели при определенной температуре более высокую интенсивность фотосинтеза, чем сеянцы, происходящие из Монтаны. Однако иногда варьирование среди сеянцев одного и того же географического происхождения было большим, чем различия между сеянцами из двух источников Люк-канен и Козловский (1972) обнаружили большие различия между клонами тополя по интенсивности фотосинтеза, фотодыхания и химического (темнового) дыхания на единицу листовой поверхности и по компенсационной точке СО2, Представители секции Aigeiros имели более низкую интенсивность фотосинтеза и более высокие компенсационные точки, чем представители секции Tacamahaca.
Различия фотосинтетической способности, наблюдаемые среди генетического материала, могут быть результатом анатомических или биохимических особенностей или тех и других одновременно. Часто изменения признаков, характеризующих устьица, являются причиной различий в поглощении СО2.
Большой интерес лесных генетиков вызывает использование интенсивности фотосинтеза в качестве показателя потенциального роста деревьев. Однако между фотосинтетической способностью и ростом деревьев установлена как высокая, так и низкая и даже отрицательная корреляция. Имеются сообщения о положительной корреляции между интенсивностью фотосинтеза и ростом у тополя, у гибрида осины и тополя, у сеянцев дугласии. Некоторые исследователи обнаружили даже обратные отношения между фотосинтезом и потенциальным ростом. Например, установлено, что интенсивность фотосинтеза и рост сеянцев сосны обыкновенной зависят от происхождения семян. Однако отмечено, что у наиболее хорошо росших сеянцев была самая низкая интенсивность фотосинтеза в расчете на сухую массу листьев. У быстрорастущих сеянцев фотосинтетически активной была меньшая часть общей массы хвои. Это обусловлено более сильным взаимным затенением хвои у быстрорастущих растений или, что более вероятно, анатомическими или биохимическими различиями хвои, связанными с происхождением семян.
По мнению некоторых исследователей, кратковременные измерения фотосинтетической способности не всегда являются надежными для оценки потенциального роста, потому что его детерминируют наряду с интенсивностью фотосинтеза, по меньшей мере, еще три важные физиологические обстоятельства. Сюда входят продолжительность роста или характер сезонных изменений фотосинтеза, отношение фотосинтеза к дыханию и распределение продуктов фотосинтеза внутри дерева.
Так, у горного экотипа сосны обыкновенной с апреля по август выявлена более высокая интенсивность фотосинтеза, чем у двух долинных экотипов. Затем интенсивность фотосинтеза горного экотипа быстро уменьшалась и осенью была ниже, чем у обоих долинных экотипов. Таким образом, прогноз прироста сухого вещества по результатам измерения фотосинтеза должен основываться как на интенсивности, так и на продолжительности фотосинтеза. Важный вывод из этого исследования заключается в том, что географические расы с очень высокой интенсивностью фотосинтеза в октябре и ноябре имели и наибольшую скорость роста.
ДНЕВНОЙ ХОД ИЗМЕНЕНИЙ ФОТОСИНТЕЗА
Ранним утром светлого безоблачного теплого дня фотосинтез идет слабо вследствие небольшой интенсивности света и низкой температуры, несмотря на высокую влажность листьев и высокую концентрацию двуокиси углерода в межклетниках листьев. При повышении интенсивности света, нагревании воздуха устьица открываются, нетто-фотосинтез начинает быстро увеличиваться и может достичь максимума до полудня. Часто вслед за максимумом наступает полуденное понижение, которое может быть небольшим или резким. Полуденная депрессия нередко сменяется новым повышением фотосинтеза в более поздние послеполуденные часы, а затем окончательно снижается. Наблюдается это, как правило, ранним вечером вслед за уменьшением интенсивности света и температуры. Вследствие изменений условий внешней среды в разные дни и в пределах одного дня дневной ход фотосинтеза часто значительно отклоняется от описанного выше. Большинство дневных изменений фотосинтеза, за исключением полуденного понижения, хорошо взаимодействуют с изменениями интенсивности света.
Например, на открытом пространстве максимум интенсивности фотосинтеза трех видов покрытосеменных наблюдался в полдень и соответствовал максимуму интенсивности общей солнечной радиации. Под пологом леса интенсивность фотосинтеза значительно изменялась в течение дня. При этом наивысшая интенсивность наблюдалась в солнечных бликах. Дневной ход фотосинтеза голосеменных был очень различным в облачные и в солнечные дни. Без затенения в пасмурные или облачные дни типичная картина была такой: интенсивность нетто-фотосинтеза возрастала, достигая максимума около полудня, затем или уменьшалась, или оставалась более или менее постоянной в течение 1-2 ч и попом понижалась. В ясные солнечные дни фотосинтез обычно быстро возрастал, достигал максимума между 9 и 12 ч утра, затем, до поздних послеполуденных часов, понижался. Позднее он вновь усиливался и достигал второго максимума, но значительно более низкого.
Причины дневных изменений фотосинтеза. Регуляция дневного хода фотосинтеза под влиянием разнообразных внешних и внутренних факторов очень сложна. Главными из внешних факторов являются, по-видимому, свет, температура, наличие воды, содержание СО2 в воздухе и различные взаимодействия между ними. Внутреннюю регуляцию поглощения СО2 относили за счет разных причин: водного стресса, закрывания устьиц, чрезмерного дыхания, накопления конечных продуктов фотосинтеза и фотоокисления ферментов. Значение отдельных факторов, влияющих на поглощение СО2, нередко изменяется. Например, утром, когда клетки тургесцентны, повышение интенсивности фотосинтеза взаимосвязано с нагреванием воздуха и увеличением интенсивности света. Однако полуденный спад при наивысшей интенсивности света часто происходит, особенно в жаркие дни, вследствие слишком большой потери воды, сопровождаемой закрыванием устьиц.
СЕЗОННЫЕ ИЗМЕНЕНИЯ
Необходимо различать сезонные изменения фотосинтетической способности деревьев, вызываемые развитием листьев и состоянием метаболизма, и наблюдаемую в полевых условиях фактическую интенсивность, которую определяют как фото синтетическую способность, так и накладывающиеся друг на друга факторы внешней среды. Это разграничение важно в связи с тем, что при исследовании сезонных изменений фотосинтеза растения часто периодически переносили из открытого грунта в лабораторию и измеряли фотосинтез при стандартных и благоприятных внешних условиях. Фактическая интенсивность фотосинтеза в полевых условиях гораздо сильнее изменяется в разные дни вследствие изменений факторов внешней среды, чем интенсивность, измеренная при стандартных условиях.
Сезонные изменения фотосинтетической способности у голосеменных происходят более постепенно, чем у листопадных покрытосеменных. Когда температура весной повышается и ночные заморозки становятся менее частыми, фотосинтетическая способность голосеменных постепенно увеличивается. Осенью интенсивность фотосинтеза также постепенно понижается. У листопадных покрытосеменных фотосинтез быстро ускоряется весной, когда деревья вновь покрываются листвой, остается высоким в течение лета, быстро уменьшается в конце лета, когда листья стареют, и окончательно падает до нуля, когда они опадают. Сезонные изменения фотосинтетической способности различаются у видов с разным характером развития листьев. У видов, побеги которых полностью формируются в зимующих почках, листовая поверхность достигает максимума в начале вегетационного периода. Гетерофильные и периодически отрастающие виды продолжают увеличивать количество листьев постепенно или путем периодического образования новых побегов в течение всего сезона (см. главу 3). Следовательно, сезонные изменения фотосинтетической способности происходят при изменении площади листьев. У голосеменных фотосинтетическая способность также изменяется при появлении новой листвы, а осенью голосеменные дольше, чем покрытосеменные, сохраняют свою способность к заметному фотосинтезу. В районах с теплыми зимами фотосинтез у вечнозеленых голосеменных может происходить в течение всех месяцев года.
Общий фотосинтез дерева и характер его сезонных изменений часто заметно варьируют в разные годы вследствие различий в размерах листовой поверхности и климатических изменений.
Сезонные изменения фотосинтетической способности сеянцев сосны ладанной и сосны веймутовой в Северной Каролине исследовали Грегор и Крамер (1963). Сеянцы выдерживали на открытом воздухе. Периодически их переносили в лабораторию и измеряли поглощение СО2 при 25°С и освещенности 43000 лк. Начиная с февраля интенсивность фотосинтеза в расчете на один сеянец медленно увеличивалась у обоих видов до апреля, затем быстро возрастала и потом снижалась в течение осени и зимы. Максимальная интенсивность у сосны ладанной достигалась в середине сентября, после чего происходило быстрое осеннее понижение. Максимум фотосинтеза у сосны веймутовой наблюдался между 15 июля и 15 сентября. Осеннее снижение было более постепенным. Более высокий и оолее поздний максимум фотосинтеза сосны ладанной был обусловлен в значительной степени тем, что у сеянцев трижды отрастали побеги, прибавляя новую хвою до конца лета. В то же время сеянцы сосны веймутовой давали новые побеги только один раз, в начале вегетационного периода.
Повышение интенсивности фотосинтеза после 9 апреля отчасти могло быть отнесено за счет увеличения ассимилирующей поверхности у обоих видов. Однако значительное увеличение с 14 февраля до 9 апреля нельзя объяснить этой причиной, так как к 9 апреля новая листва еще не распустилась. Это увеличение должно было происходить в результате восстановления фотосинтетической активности уже имевшейся хвои. Уменьшение фотосинтеза после максимума, наблюдавшегося в середине сезона, было вызвано у обоих видов не потерей хвои, а скорее пониженной фотосинтетической способностью существовавшей хвои.
Сезонные изменения интенсивности фотосинтеза на единицу листвы значительно отличались от сезонного хода интенсивности фотосинтеза в пересчете на один сеянец. У сосны ладанной интенсивность фотосинтеза на единицу длины пучка хвои заметно повышалась с февраля по март, несмотря на то, что в этот период новая хвоя не появлялась. Максимальная интенсивность на единицу длины пучка была достигнута в мае, за 4 месяца до максимума интенсивности на один сеянец. Высокая интенсивность сохранялась до сентября, а затем интенсивность фотосинтеза на единицу длины хвои и на один сеянец начала понижаться. У хвои сосны веймутовой фотосинтез также заметно повышался с февраля по март, но максимум не был достигнут до июля. Высокая интенсивность сохранялась в течение сентября, несколько понизилась в ноябре и достигла минимума в январе.
Зимний фотосинтез вечнозеленых растений. В районах с мягкими зимами у вечнозеленых растений фотосинтез осуществляется в течение всего года. В местах с суровыми зимами фотосинтез может быть незначительным несколько недель или месяцев.
Наблюдаемое поглощение СО2 или прирост сухой массы в течение зимы были установлены для сосны ежовой и сосны ладанной на юге Соединенных Штатов, для сосны замечательной и сосны обыкновенной в Аберистуите (Уэльс) и для широколистных вечнозеленых растений в Северной Италии. Деревья дугласии накапливали значительные количества продуктов фотосинтеза зимой в мягких климатических условиях прибрежного района северо-запада Соединенных Штатов. В засушливый год чистый прирост продуктов фотосинтеза в течение зимы приближался к 1/4 общего прироста за весь год. Вдоль всей прибрежной зоны Норвегии у сосен и елей уменьшалась сухая масса в течение коротких периодов зимой, но за всю зиму у них обнаружился чистый прирост, доказавший превышение фотосинтеза над дыханием. Сухая масса сеянцев ели ситхинской в Южной Шотландии удваивалась за период с конца сентября и до середины апреля. Большая часть прироста приходилась на конец марта — начало апреля, но некоторое количество сухой массы накапливалось в середине зимы.
В районах с холодными зимами интенсивность фотосинтеза у вечнозеленых растений наблюдалась вплоть до точки замерзания или даже немного ниже. Например, около Мюнхена поглощение СО2 елью европейской происходило в дни, когда температура воздуха была немного ниже нуля. В долине около Инсбрука (Австрия) фотосинтез ели европейской продолжался и зимой до наступления сильных морозов. Однако на границе древесной растительности около горы Патчеркофель морозная погода в ноябре (от -10° до -15°) вызвала прекращение фотосинтеза. Пизек и Випклер (1958) показали, что интенсивность нетто-фотосинтеза ели европейской и сосны кедровой европейской была заметной до поздней осени. Последующие изменения температуры на несколько градусов ниже и выше нуля вызывали колебания поглощения СО2. Как только температура падала ниже -4° или -5°С, фотосинтез прекращался. Если же после этого морозы повторялись в течение нескольких ночей, то днем фотосинтез был ингибирован, даже если температура поднималась выше нуля. После мороза от -6° до -8°С нетто-фотосинтез прекращался, и требовалось несколько дней с мягкой погодой для восстановления фотосинтетической способности. Полностью фотосинтез возобновлялся с повышением температуры весной. Особенно это касалось частей кроны, в которых происходил распад хлорофилла в течение зимы. Весной вместе с колебаниями температуры колебалась и интенсивность фотосинтеза. Следовательно, фотосинтетический аппарат оставался функционально активным только до тех пор, пока зимой не наступали морозы, У верхней границы распространения леса температуры были в течение 4-5 месяцев такими низкими, что фотосинтез по существу был невозможен.
www.ronl.ru
Основной источник энергии
Слово «фотосинтез» означает буквально создание или сборку чего-то под действием света. Обычно, говоря о фотосинтезе, имеют в виду процесс, посредством которого растения на солнечном свету синтезируют органические соединения из неорганического сырья. Все формы жизни во Вселенной нуждаются в энергии для роста и поддержания жизни. Водоросли, высшие растения и некоторые типы бактерий улавливают непосредственно энергию солнечного излучения и используют ее для синтеза основных пищевых веществ. Животные не умеют использовать солнечный свет непосредственно в качестве источника энергии, они получают энергию, поедая растения или других животных, питающихся растениями. Итак, в конечном счете источником энергии для всех метаболических процессов на нашей планете, служит Солнце, а процесс фотосинтеза необходим для поддержания всех форм жизни на Земле.
Мы пользуемся ископаемым топливом — углем, природным газом, нефтью и т. д. Все эти виды топлива — не что иное, как продукты разложения наземных и морских растений или животных, и запасенная в них энергия была миллионы лет назад получена из солнечного света. Ветер и дождь тоже обязаны своим возникновением солнечной энергии, а следовательно, энергия ветряных мельниц и гидроэлектростанций в конечном счете также обусловлена солнечным излучением.
Важнейший путь химических реакций при фотосинтезе — это превращение углекислоты и воды в углероды и кислород. Суммарную реакцию можно описать уравнением СО2+Н20? [СН20]+02
Углеводы, образующиеся в этой реакции, содержат больше энергии, чем исходные вещества, т. е. СО2 и Н20. Таким образом, за счет энергии Солнца энергетические вещества (СО2 и Н20) превращаются в богатые энергией продукты — углеводы и кислород. Энергетические уровни различных реакций, описанных суммарным уравнением, можно охарактеризовать величинами окислительно-восстановительных потенциалов, измеряемых в вольтах. Значения потенциалов показывают, сколько энергии запасается или растрачивается в каждой реакции. Итак, фотосинтез можно рассматривать как процесс образования лучистой энергии Солнца в химическую энергию растительных тканей.
Содержание СО2 в атмосфере остается почти полным, несмотря на то, что углекислый газ расходуется в процессе фотосинтеза. Дело в том, что все растения и животные дышат. В процессе дыхания в митохондриях кислород, поглощаемый из атмосферы живыми тканями, используется для окисления углеводов и других компонентов тканей с образованием в конечном счете двуокиси углерода и воды и с сопутствующим выделением энергии. Высвобождающаяся энергия запасается в высокоэнергетические соединения — аденозинтрифосфат (АТФ), который и используется организмом для выполнения всех жизненных функций. Таким образом дыхание приводит к расходованию органических веществ и кислорода и увеличивает содержание СО2 на н планете. На процессы дыхания во всех живых организмах и на сжигание всех видов топлива, содержащих углерод, в совокупности расходуется в масштабах Земли в среднем около 10000 тонн 02 в секунду. При такой скорости.потребления весь кислород в атмосфере должен бы иссякнуть примерно, через 3000 лет. К счастью для нас, расход органических веществ и атомного кислорода уравновешивается созданием углеводов и кислорода в результате фотосинтеза. В идеальных условиях скорость фотосинтеза в зеленых тканях растений примерно в 30 раз превышает скорость дыхания в тех же тканях, таким образом, фотосинтез служит важным фактором, регулирующим содержание 02 на Земле.
История открытия фотосинтеза
В начале XVII в. фламандский врач Ван Гельмонт вырастил в кадке с землей дерево, которое он поливал только дождевой водой. Он заметил, что спустя пять лет, дерево выросло до больших размеров, хотя количество земли в кадке практически не уменьшилось. Ван Гельмонт, естественно, сделал вывод, что материал, из которого образовалось дерево произошел из воды, использованной для полива. В 1777 английский ботаник Стивен Хейлс опубликовал книгу, в которой сообщалось, что в качестве питательного вещества, необходимого для роста, растения используют главным образом воздух. В тот же период знаменитый английский химик Джозеф Пристли (он был одним из первооткрывателей кислорода) провел серию опытов по горению и дыханию и пришел к выводу о том, что зелёные растения способны совершать все те дыхательные процессы, которые были обнаружены в тканях животных. Пристли сжигал свечу в замкнутом объеме воздуха, и обнаруживал, что получавшийся при этом воздух уже не может поддерживать горение. Мышь, помещенная в такой сосуд, умирала. Однако веточка мяты продолжала жить в воздухе неделями. В заключение Пристли обнаружил, что в воздухе, восстановленном веточкой мяты, вновь стала гореть свеча, могла дышать мышь. Теперь мы знаем, что свеча, сгорая, потребляла кислород из замкнутого объема воздуха, но затем воздух снова насыщался кислородом благодаря фотосинтезу, происходившему в оставленной веточке мяты. Спустя несколько лет голландский врач Ингенхауз обнаружил, что растения окисляют кислород лишь на солнечном свету и что только их зеленые части обеспечивают выделение кислорода. Жан Сенебье, занимавший пост министра, подтвердил данные Ингенхауза и продолжил исследование, показав, что в качестве питательного вещества растения используют двуокись углерода, растворенную в воде. В начале XIX века другой швейцарский исследователь де Соседи изучал количественные взаимосвязи между поглощенной растением углекислотой, с одной стороны, и синтезированными органическими веществами и кислородом — с другой. В результате своих опытов он пришел к выводу, что вода также потребляется растением при ассимиляции СО2. В 1817 г. два французских химика, Пельтье и Каванту, выделили из листьев зеленое вещество и назвали его хлорофиллом. Следующей важной вехой в истории изучения фотосинтеза было сделанное в 1845 г. немецким физиком Робертом Майером утверждение о том, что зеленые растения преобразуют энергию, солнечного света в химическую энергию. Представления о фотосинтезе, сложившиеся к середине прошлого века, можно выразить следующим соотношением:
Зеленое растение
СО2+ Н2 О + Свет? О2 + орг. вещества +химическая энергия
Отношение количества С02, поглощенного при фотосинтезе, к количеству выделенного 02, точно измерил французский физиолог растений Бусэнго. В 1864 г. он обнаружил, что фотосинтетическое отношение, т.е. отношение объема выделенного 02 к объему поглощенного С02, почти равно единице. В том же году немецкий ботаник Закс (открывший также у растений дыхание) продемонстрировал образование зерен крахмала при фотосинтезе. Закс помещал зеленые листья на несколько часов в темноту для того, чтобы они израсходовали накопленный в них крахмал. Затем он выносил листья на свет, но при этом освещал лишь половину каждого листа, оставляя другую половину листа в темноте. Спустя некоторое время весь лист целиком обрабатывали парами йода. В результате освещенная часть листа становилась темно-фиолетовой, что свидетельствовало об образовании комплекса крахмала с йодом, тогда как цвет другой половины листа не изменялся. Прямую связь между выделением кислорода и хлоропластами в зеленых листьях, а также соответствие спектра действия фотосинтеза спектру поглощеных хлоропластами установил в 1880 г. Энгельман. Он поместил нитевидную зеленую водоросль имеющую спирально извитые хлоропласты, на предметное стекло, освещая его узким и широким пучком белого света. Вместе с водорослью на предметное стекло наносилась суспензия клеток подвижных бактерий, чувствительных к концентрации кислорода. Предметное стекло помещали в камеру без воздуха и освещали. В этих условиях подвижные бактерии должны были перемещаться в ту часть, где концентрация 02 была выше. После прошествия некоторого времени образец рассматривали под микроскопом и подсчитывали распределение бактериопопуляции. Оказалось, что бактерии концентрировались вокруг зеленых полосок в нитевидной водоросли. В другой серии опытов Энгельман освещал водоросли лучами разного спектрального состава, установив призму между источником света и предметным столиком микроскопа. Наибольшее число бактерий в этом случае скапливалось вокруг тех участков водоросли, которые освещались синим и красным областями спектра. Находящиеся в водорослях хлорофиллы поглощали синий и красный свет. Поскольку к тому времени было уже известно, что для фотосинтеза необходимо поглощение света, Энгельман заключил, что хлорофиллы участвуют в синтезе в качестве пигментов, являющихся активными фоторецепторами. Уровень знаний о фотосинтезе в начале нашего века можно представить следующим образом.
СО2 + Н2О + Свет –О2 + Крахмал + Химическая энергия
Итак, к началу нашего века суммарная реакция фотосинтеза была уже известна. Однако биохимия находилась не на таком высоком уровне, чтобы полно раскрыть механизмы восстановления двуокиси углерода до углеводов. К сожалению, следует признать, что и теперь еще некоторые аспекты фотосинтеза изучены довольно плохо. Издавна делались попытки исследовать влияние интенсивности света, температуры, концентрации углекислоты и т… п. на общий выход фотосинтеза. И хотя в этих работах исследовались растения самых разных видов, большинство измерений было выполнено на одноклеточных зеленых водорослях и на одноклеточной жгутиковой водоросли Эвглена. Одноклеточные организмы удобнее для качественного исследования, поскольку их можно выращивать во всех лабораториях при вполне стандартных условиях. Они могут быть равномерно суспензированы, т. е. взвешены в водных буферных растворах, и нужный объем такой суспензии, или взвеси, можно брать такой дозировки, точно так же, как при работе с обычными растениями. Хлоропласты для опытов лучше всего выделять из листьев высших растений. Чаще всего используют шпинат, потому что его легко выращивать и свежие листья удобны для проведения исследований; иногда используются листья гороха и салата-латука.
Поскольку СО2 хорошо растворяется в воде, а О2 относительно нерастворим в воде, то при фотосинтезе в замкнутой системе давление газа в этой системе может изменяться. Поэтому влияние света на фотосинтетические системы часто исследуют с помощью респиратора Варбурга, позволяющего регистрировать пороговые изменения объема 02 в системе. Впервые респиратор Варбурга был использован применительно к фотосинтезу в 1920г. Для измерения потребления или выделения кислорода в ходе реакции удобнее пользоваться другим прибором — кислородным электродом. В основе этого устройства лежит использование полярографического метода. Кислородный электрод обладает достаточной чувствительностью для того, чтобы обнаружить в таких низких концентрациях как 0,01 ммоль в 1 л. Прибор состоит из катода достаточно тонкой платиновой проволоки, герметично впрессованной в пластину анода, представляющего собой кольцо из серебряной проволоки, погруженной в насыщенный раствор. Электроды отделены от смеси, в которой протекает реакция, мембраной, проницаемой для 02. Реакционная система находится в пластмассовом или стеклянном сосуде и постоянно перемешивается вращающимся стержневым магнитом. Когда к электродам приложено напряжение, платиновый электрод становится отрицательным по отношению к стандартному электроду, кислород в растворе электролитически восстанавливается. При напряжении от 0,5 до 0,8 В величина электрического тока линейно зависит от парциального давления кислорода в растворе. Обычно с кислородным электродом работают при напряжении около 0,6 В. Электрический ток измеряют, присоединив электрод к подходящей регистрирующей системе. Электрод вместе с реакционной смесью орошают потоком воды от термостата. С помощью кислородного электрода измеряют действие света и различных химических веществ на фотосинтез. Преимущество кислородного электрода перед аппаратом Варбурга состоит в том, что кислородный электрод позволяет быстро и непрерывно регистрировать изменения содержания О2 в системе. С другой стороны, в приборе Варбурга можно одновременно исследовать до 20 образцов с различными реакционными смесями, тогда как при работе с кислородным электродом образцы приходится анализировать поочередно.
Примерно до начала 1930-х годов многие исследователи в этой области полагали, что первичная реакция фотосинтеза заключается в расщеплении двуокиси углерода под действием света на углерод и кислород с последующим восстановлением углерода до углеводов с участием воды в ходе нескольких последовательных реакций. Точка зрения изменилась в 1930-х годах в результате двух важных открытий. Во-первых, были описаны разновидности бактерий, способных ассимилировать и синтезировать углеводы, не используя для этого энергию света. Затем, голландский микробиолог Ван Нил сравнил процессы фотосинтеза у бактерий и показал, что некоторые бактерии могут ассимилировать С02 на свету, не выделяя при этом кислорода. Такие бактерии способны к фотосинтезу лишь при наличии подходящего субстрата-донора водорода. Ван Нил предполагал, что фотосинтез зеленых растений и водорослей является частным случаем, когда кислород в фотосинтезе происходит из воды, а не из углекислоты.
Второе важное открытие сделал в 1937 г. Р. Хил в Кембриджском университете. С помощью дифференциального центрифугирования гомогената тканей листа он отделил фотосинтезирующие частицы (хлоропласты) от дыхательных частиц. Полученные Xиллом хлоропласты при освещении сами по себе не выделяли кислорода (возможно, из-за того, что они были повреждены при разделении). Однако они начинали выделять кислород на свету, если в суспензию вносили подходящие акцепторы электрона (окислители), например ферриоксалат калия или феррицианид калия. При выделении одной молекулы 02 фотохимически восстанавливались четыре эквивалента окислителя. Позднее было обнаружено, что многие хиноны и красители восстанавливаются хлоропластами на свету. Однако хлоропласты не могли восстановить СО2, природный акцептор электронов при фотосинтезе. Это явление, известное теперь как реакция Хилла, представляет собой индуцируемый светом перенос электронов от воды к нефизиологическим окислителям (реагентам Хилла) против градиента химического потенциала. Значение реакции Хилла состоит в том, что она продемонстрировала возможность разделения двух процессов — фотохимического выделения кислорода и восстановления углекислоты при фотосинтезе.
Разложение воды, приводящее к выделению свободного кислорода при фотосинтезе, было установлено Рубеном и Каменом, в Калифорнии в 1941 г. Они поместили фотосинтезирующие клетки в воду, обогащенную изотопом кислорода, имеющим массу 18 атомных единиц 180. Изотопный состав кислорода, выделенного клетками, соответствовал составу воды, но не С02. Кроме того, Камен и Рубен открыли радиоактивный изотоп 18О, который впоследствии успешно использовали Бассэт, Бенсон Вин, изучавшие путь превращения углекислоты при фотосинтезе. Кальвин и его сотрудник установили, что восстановление углекислоты до сахаров происходит в результате темновых ферментативных процессов, причем для восстановления одной молекулы углекислоты требуются две молекулы восстановленного АДФ и три молекулы АТФ. К тому времени роль АТФ и пиридиннуклеотидов в дыхании тканей была установлена. Возможность фотосинтетического восстановления АДФ до АТФ выделенными хлорофиллами была доказана в 1951 г. в трех разных лабораториях. В 1954 г. Арнон, Аллен продемонстрировали фотосинтез — они наблюдали ассимиляцию С02 и 02 выделенными хлоропластами шпината. В течение следующего десятилетия из хлоропластов удалось выделить белки, участвующие в переносе электронов в синтезе -ферредоксин, пластоцианин, ферроАТФ-редуктазу, цитохромы и т. д.
Таким образом, в здоровых зеленых листьях, под действием света образуются АДФ и АТФ и энергия гидросвязей используются для восстановления С02 до углеводов в присутствии ферментов, причем активность ферментов регулируется светом.
Лимитирующие факторы
Интенсивность, или скорость процесса фотосинтеза в растении зависит от ряда внутренних и внешних факторов. Из внутренних факторов наиболее важное значение имеют структура листа и содержание в нем хлорофилла, скорость накопления продуктов фотосинтеза в хлоропластах, влияние ферментов, а также наличие малых концентраций необходимых неорганических веществ. Внешние параметры — это количество и качество света, попадающего на листья, температура окружающей среды, концентрация углекислоты и кислорода в атмосфере вблизи растения.
Скорость фотосинтеза возрастает линейно, или прямо пропорционально увеличению интенсивности света. По мере дальнейшего увеличения интенсивности света нарастание фотосинтеза становится все менее и менее выраженным, и, наконец, прекращается, когда освещенность достигает определенного уровня 10000 люкс. Дальнейшее увеличение интенсивности света уже не влияет на скорость фотосинтеза. Область стабильной скорости фотосинтеза называется областью светонасыщения. Если нужно увеличить скорость фотосинтеза в этой области, следует изменять не интенсивность света, а какие-либо другие факторы. Интенсивность солнечного света, попадающего в ясный летний день на поверхность земли, во многих местах нашей планеты составляет примерно 100000 люкс. Следовательно, растениям, за исключением тех, которые растут в густых лесах и в тени, падающего солнечного света бывает достаточно для насыщения их фотосинтетической активности (энергия квантов, соответствующих крайним участкам видимого диапазона – фиолетового и красного, различается всего лишь в два раза, и все фотоны этого диапазона в принципе способны осуществить запуск фотосинтеза).
В случае низких интенсивностей света скорость фотосинтеза при 15 и 25°С одинакова. Реакции, протекающие при таких интенсивностях света, которые соответствуют области лимитирования света, подобно истинным фотохимическим реакциям, не чувствительны к температурам. Однако при более высоких интенсивностях скорость фотосинтеза при 25°С гораздо выше, чем при 15°С. Следовательно, в области светового насыщения уровень фотосинтеза зависит не только от поглощения фотонов, но и от других факторов. Большинство растений в умеренном климате хорошо функционируют в интервале температур от 10 до 35°С, наиболее благоприятные условия — это температура около 25°С.
В области лимитирования светом скорость фотосинтеза не изменяется при уменьшении концентрации СО2. Отсюда можно сделать вывод, что С02 участвует непосредственно в фотохимической реакции. В то же время при более высоких интенсивностях освещения, лежащих за пределами области лимитирования, фотосинтез существенно возрастает при увеличении концентрации СО2. У некоторых зерновых культур фотосинтез линейно возрастал при увеличении концентрации СО2 до 0,5%. (эти измерения проводили в кратковременных опытах, поскольку длительное воздействие высоких концентраций СО2 повреждает листы). Высоких значений скорость фотосинтеза достигает при содержании С02 около 0,1%. Средняя концентрация углекислоты в атмосфере составляет от 0,03%. Поэтому в обычных условиях растениям не хватает СО2 для того, чтобы с максимальной эффективностью использовать попадающий на них солнечный свет. Если помещенное в замкнутый объем растение освещать светом насыщающей интенсивности, то концентрация СО2 в объеме воздуха будет постепенно уменьшаться и достигнет постоянного уровня, известного под названием «С02 компенсационного пункта». В этой точке появление СО2 при фотосинтезе уравновешивается выделением О2 в результате дыхания (темнового и светового). У растений разных видов положения компенсационных пунктов различны.
--PAGE_BREAK--Световые и темновые реакции.
Еще в 1905 г. английский физиолог растений Ф. Ф. Блекмэн, интерпретируя форму кривой светового насыщения фотосинтеза, высказал предположение, что фотосинтез представляет собой двухстадийный процесс, включающий фотохимическую, т.е. светочувствительную реакцию и нефотохимическую, т. е. темновую, реакцию. Темновая реакция, будучи ферментативной, протекает медленнее, чем световая реакция, и поэтому при высоких интенсивностях света скорость фотосинтеза полностью определяется скоростью темновой реакции. Световая реакция либо вообще не зависит от температуры, либо зависимость эта выражена очень слабо, тогда темновая реакция, как и все ферментативные процессы, зависит от температуры в довольно значительно и степени. Следует ясно представлять себе, что реакция, называемая темновой, может протекать как в темноте, так и на свету. Световую и темновую реакции можно разделить, используя вспышки света, длящиеся краткие доли секунды. Вспышки света длительностью меньше одной миллисекунды (10-3 с) можно получить либо с помощью механического приспособления, поставив на пути пучка постоянного света вращающийся диск со щелью, либо электрически, заряжая конденсатор и разряжая его через вакуумную или газоразрядную лампу. В качестве источников света пользуются также рубиновыми лазерами с длиной волны излучения 694 нм. В 1932 г. Эмерсон и Арнольд освещали суспензию клеток вспышками света от газоразрядной лампы с длительностью около 10-3с. Они измеряли скорость выделения кислорода в зависимости от энергии вспышек, длительности темнового промежутка между вспышками и температуры суспензии клеток. При увеличении интенсивности вспышек насыщение фотосинтеза в нормальных клетках наступало, когда выделялась одна молекула 02 на 2500 молекул хлорофилла. Эмерсон и Арнольд сделали вывод, что максимальный выход фотосинтеза определяется не числом молекул хлорофилла, поглощающих свет, а числом молекул фермента, катализирующего темновую реакцию. Они также обнаружили, что при увеличении темновых интервалов между последовательными вспышками за пределы 0,06 с выход кислорода в расчете на одну вспышку уже не зависел от длительности темнового интервала, тогда как при более коротких промежутках он возрастал с увеличением длительности темнового интервала (от 0 до 0,06 с). Таким образом, темновая реакция, которая определяет уровень насыщения фотосинтеза, завершается примерно за 0,06 с. На основе этих данных было рассчитано, что среднее время, характеризующее скорость реакции, составило около 0,02 с при 25°С.
Структурная и биохимическая организация аппарата фотосинтеза
Современные представления о структурной и функциональной организации фотосинтетического аппарата включают широкий круг вопросов, связанных с характеристикой химического состава пластид, спецификой их структурной организации, физиолого-генетическими закономерностями биогенеза этих органоидов и их взаимоотношениями с другими функциональными структурами клетки. У наземных растений специальным органом фотосинтетической деятельности служит лист, где локализованы специализированные структуры клетки — хлоропласты, содержащие пигменты и другие компоненты, необходимые для процессов поглощения и преобразования энергии света в химический потенциал. Кроме листа функционально активные хлоропласты присутствуют в стеблях растений, черешках, остях и чешуях колоса и даже в освещаемых корнях ряда растений. Однако именно лист был сформирован в ходе длительной эволюции как специальный орган для выполнения основной функции зеленого растения – фотосинтеза, поэтому анатомия листа, расположение хлорофиллсодержащих клеток и тканей, их соотношение с другими элементами морфемной структуры листа подчинены наиболее эффективному течению процесса фотосинтеза, и они в наибольшей степени подвергаются интенсивным изменениям в условиях экологического стресса.
В связи с этим проблему структурно — функциональной организации фотосинтетического аппарата целесообразно рассмотреть в двух основных уровнях — на уровне листа как органа фотосинтеза и хлоропластов, где целиком сосредоточен весь механизм фотосинтеза.
Организация фотосинтетического аппарата на уровне листа может быть рассмотрена на основе анализа его мезострктуры. Понятие «мезоструктура» было предложено в 1975 году. По представлениям о структурной и функциональной особенностях фотоситнетического аппарата с характеристикой химического состава, структурной организации, физиолого-генетическими особенностями биогенеза этих органоидов и их взаимоотношениями с другими функциональными структурами специальным органом фотосинтетического процесса является лист, где локализованы специализированные образования — хлоропласты, содержащие пигменты, необходимые для процессов поглощения и преобразования света в химический потенциал. Кроме того, активные хлоропласты присутствуют в стеблях, остях и чешуях колоса и даже в освещенных частях корней некоторых растений. Однако именно лист был сформирован всем ходом эволюции как специальный орган для выполнения основной функции зеленого растения — фотосинтеза.
Мезоструктура включает систему морфофизиологических характеристик фотосинтетического аппарата листа, хлоренхимы и клезофилла. Основные показатели мезоструктуры фотосинте-
тического аппарата (по А. Т. Мокроносову) включают: площадь, число клеток, хлорофилл, белок, объем клетки, количество хлоропластов в клетке, объем хлоропласта, площадь сечения хлоропласта и его поверхность. Анализ мезоструктуры и функциональной активности фотосинтетического аппарата у многих видов растений помогают определить наиболее часто встречающиеся значения исследуемых показателей и пределы варьирования отдельных характеристик. Согласно этим данным, основные показатели мезоструктуры фотосинтетического аппарата (Мокроносов, 19В1):
I — площадь листа;
II — число клеток на 1 см2,
III — хлорофилл на 1 дм2, ключевые ферменты на 1 дм2, объем клетки, тыс. мкм2, число хлоропластов в клетке;
IV — объем хлоропластов, площадь проекции хлоропласта, мкм2, поверхность хлоропласта, мкм2.
Среднее число хлоропластов у закончившего рост листа обычно достигает 10-30, у некоторых видов оно превышает 400. Это соответствует млн хлоропластов в расчете на 1 см2площади листа. Хлоропласты сосредоточены в клетках различных тканей в количестве 15 – 80 штук на клетку. Средний объем хлоропласта — один мкм2. У большинства растений суммарный объем всех хлоропластов составляет 10-20%, у древесных растений — до 35% объема клетки. Отношение общей поверхности хлоропластов к площади листа находится в пределах 3-8. В одном хлоропласте содержится разное количество молекул хлорофилла, у тенелюбивых видов их число возрастает. Приведенные выше показатели могут значительно варьировать в зависимости от физиологического состояния и экологических условий роста растений. По данным А. Т. Мокроносова, в молодом листе активизация фотосинтеза при удалении 50-80% листа обеспечивается увеличением числа хлоропластов в клетке без изменения их индивидуальной активности, в то время как в листе, окончившем рост, усиление фотосинтеза после дефолиации происходит за счет повышения активности каждого хлоропласта без изменения их числа. Анализ мезоструктуры показал, что адаптация к условиям освещения вызывает перестройку, которая оптимизирует светопоглошающие свойства листа.
Хлоропласты имеют наиболее высокую степень организации внутренних мембранных структур по сравнению с другими органоидами клетки. По степени упорядоченности структур хлоропласты можно сравнить только с рецепторными клетками сетчатки глаза, также выполняющими функцию преобразования световой энергии. Высокая степень организации внутренней структуры хлоропласта определяется рядом моментов:
1) необходимостью пространственного разделения восстановленных и окисленных фотопродуктов, возникающих в результате первичных актов разделения заряда в реакционном центре;
2) необходимостью строгой упорядоченности компонентов реакционного центра, где сопряжёны быстропротекающие фотофизиологические и более медленные энзиматические реакции: преобразование энергии фотовозбужденной молекулы пигмента требует ее определенной ориентации по отношению к химическому акцептору энергии, что предполагает наличие определенных структур, где пигмент и акцептор жестко ориентированы друг относительно друга;
3) пространственная организация электронно-транспортной цепи требует последовательной и строго ориентированной организации переносчиков в мембране, обеспечивающей возможность быстрого и регулируемого транспорта электронов и протонов;
4) для сопряжения транспорта электронов и синтеза АТФ требуется определенным образом организованная система хлоропластов.
Липопротеидные мембраны как структурная основа энергетических процессов возникают на самых ранних этапах эволюции, предполагают, что основные липидные компоненты мембран — фосфолипиды — образовались в определенных биологических условиях. Формирование липидных комплексов обусловило возможность включения в них различных соединений, что по-видимому, явилось основой первичных каталитических функций данных структур.
Проведенные в последние годы электронномикроскопические исследования обнаружили организованные мембранные структуры у организмов, стоящих на самой низкой ступени эволюции. У некоторых бактерий мембранные фотесинтезирующие структуры клеток тесно упакованных органелл расположены по периферии клетки и связаны с цитоплазматическими мембранами; кроме того, в клетках зеленых водорослей процесс фотосинтеза связан с системой двойных замкнутых мембран — тилакоидов, локализованных в периферической части клетки. У данной группы фотосинтетических организмов впервые появляется хлорофилл, а образование специализированных органелл – хлоропластов встречается у криптофитовых водорослей. В них находятся по два хлоропласта, содержащих от одного до нескольких тилакоидов. Сходное строение фотосинтетического аппарата имеет место и у других групп водорослей: красных, бурых, и др. В процессе эволюции мембранная структура фотосинтетического процесса усложняется.
Микроскопические исследования хлоропласта, техника криоскопии позволили сформулировать пространственную модель объемной организация хлоропластов. Наиболее известна гранулярно-решетчатая модель Дж. Хеслоп-Харрисона (1964).
Таким образом, фотосинтез – это сложный процесс преобразования световой энергии в энергию химических связей органических веществ, необходимых для жизнедеятельности как самих фотосинтезирующих организмов, так и других организмов, не способных к самостоятельному синтезу органических веществ.
Изучение проблем фотосинтеза, помимо общебиологических, имеет и прикладное значение. В частности, проблемы питания, создания систем жизнеобеспечения при космических исследованиях, использования фотосинтезирующих организмов для создания различных биотехнических устройств непосредственно связаны с фотосинтезом.
Список литературы
1. Д.Халл, К.Рао «Фотосинтез». М.,1983
2. Мокроносов А.Г. «Фотосинтетическая реакция и целостность растительного организма». М.,1983
3. Мокроносов А.Г., Гавриленко В.Ф. «Фотосинтез: физиолого – экологические и биохимические аспекты» М.,1992
4. «Физиология фотосинтеза» под ред. Ничипоровича А.А., М.,1982
5. Вечер А.С. «Пластиды растнеий»
6. Виноградов А.П. «Изотопы кислорода и фотосинтез»
7. Годнев Т.Н. «Хлорофилл и его строение».
8. Гуринович Г.П., Севченко А.Н., Соловьев К.Н. «Спектроскопия хлорофилла»
9. Красновский А.А. «Преобразование энергии света при фотосинтезе»
Для подготовки данной работы были использованы материалы с сайта www.ronl.ru/
www.ronl.ru
Скачать реферат по биологии на тему «Фотосинтез» в формате doc.
Скачать презентацию на тему «Фотосинтез» в формате ppt.
В данной статье Школьного портала представим Вашему вниманию краткое описание явления фотосинтеза и процессов тесно с ним переплетающихся. Слово «фотосинтез» в буквальном смысле подразумевает создание, сборку чего-либо под воздействием света. Когда говорят про фотосинтез, обычно имеют в виду процесс, благодаря которому растения синтезируют на солнечном свету органические соединения из веществ неорганического происхождения.
Известно, что все живые организмы нуждаются в энергии для поддержания жизни и роста. Водоросли, высшие растения и даже некоторые бактерии используют естественным путем энергию солнечного излучения для синтеза необходимых им пищевых веществ. А вот животные не способны использовать солнечный свет в качестве естественного источника энергии, получая ее лишь поеданием растений или иных животных, о чем и упоминает данный реферат по биологии.
Здесь можно сделать предварительный вывод актуальности фотосинтеза: источником энергии практически всех метаболических процессов на Земле, служит Солнце, а сам процесс фотосинтеза нужен для поддержания всевозможных форм жизни на планете.
Данный реферат по биологии (ссылка для скачивания в начале статьи) после вводной информации освещает тему «Фотосинтез», начиная с определения этого процесса в качестве основного источника энергии. Во второй главе подробно описана история открытия фотосинтеза и хронологическое описание событий, тесно связанных с ней. Затем в работе подробно описаны процессы, происходящие в листе, что наглядно иллюстрируется также слайдами презентации «Фотосинтез» (также ссылка для скачивания в начале статьи).
 На следующем этапе рассматриваются световые и темновые стадии фотосинтеза, приводятся реакции, протекающие в каждой из них, и указаны преимущества одной, в сравнении с другой. Интересный материал размещен в пятой главе, где изложена информация о прямой связи процесса фотосинтеза и урожая, в частности культурных растений. Далее описываются гипотезы и аргументы в пользу восклицательного утверждения: «Леса — легкие планеты!». В последней главе реферативной работы рассматривается немного риторический вопрос: «Могут ли существовать существа человекоподобные зеленого цвета?».
На следующем этапе рассматриваются световые и темновые стадии фотосинтеза, приводятся реакции, протекающие в каждой из них, и указаны преимущества одной, в сравнении с другой. Интересный материал размещен в пятой главе, где изложена информация о прямой связи процесса фотосинтеза и урожая, в частности культурных растений. Далее описываются гипотезы и аргументы в пользу восклицательного утверждения: «Леса — легкие планеты!». В последней главе реферативной работы рассматривается немного риторический вопрос: «Могут ли существовать существа человекоподобные зеленого цвета?».
Здесь достаточно логично описана вероятность возникновения человекоподобного организма, в коже которого, как в растении, содержался бы хлорофилл, позволяя синтезировать органические соединения. Однако, по итогам данного рассуждения, автор приход к выводу, что площадь такой кожи давала бы в 20 раз меньше калорий, которые необходимы человеку для нормальной жизнедеятельности.
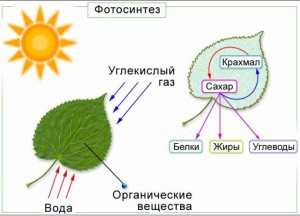 Подводя итог обзора работы и презентации, отметим следующее. Фотосинтез является одним из основополагающих процессов жизни, именно на нем основана наибольшая часть растительной фауны на Земле.
Подводя итог обзора работы и презентации, отметим следующее. Фотосинтез является одним из основополагающих процессов жизни, именно на нем основана наибольшая часть растительной фауны на Земле.
Изучение вопросов, связанных с фотосинтезом, помимо общебиологического значения, имеет и достаточно прикладных направлений. Например, проблемы питания, создание биотехнических устройств на основе деятельности фотосинтезирующих организмов, создание систем жизнеобеспечения для размещения в космических аппаратах и т.д.
Реферат по биологии «Фотосинтез» дополнен интересной презентацией, которую Вы может просмотреть и скачать ниже ↓
Скачать презентацию на тему «Фотосинтез» в формате ppt
nashashcola.ru
Основной источник энергии
Слово «фотосинтез» означает буквально создание или сборку чего-то под действием света. Обычно, говоря о фотосинтезе, имеют в виду процесс, посредством которого растения на солнечном свету синтезируют органические соединения из неорганического сырья. Все формы жизни во Вселенной нуждаются в энергии для роста и поддержания жизни. Водоросли, высшие растения и некоторые типы бактерий улавливают непосредственно энергию солнечного излучения и используют ее для синтеза основных пищевых веществ. Животные не умеют использовать солнечный свет непосредственно в качестве источника энергии, они получают энергию, поедая растения или других животных, питающихся растениями. Итак, в конечном счете источником энергии для всех метаболических процессов на нашей планете, служит Солнце, а процесс фотосинтеза необходим для поддержания всех форм жизни на Земле.
Мы пользуемся ископаемым топливом — углем, природным газом, нефтью и т. д. Все эти виды топлива — не что иное, как продукты разложения наземных и морских растений или животных, и запасенная в них энергия была миллионы лет назад получена из солнечного света. Ветер и дождь тоже обязаны своим возникновением солнечной энергии, а следовательно, энергия ветряных мельниц и гидроэлектростанций в конечном счете также обусловлена солнечным излучением.
Важнейший путь химических реакций при фотосинтезе — это превращение углекислоты и воды в углероды и кислород. Суммарную реакцию можно описать уравнением СО2+Н20? [СН20]+02
Углеводы, образующиеся в этой реакции, содержат больше энергии, чем исходные вещества, т. е. СО2 и Н20. Таким образом, за счет энергии Солнца энергетические вещества (СО2 и Н20) превращаются в богатые энергией продукты — углеводы и кислород. Энергетические уровни различных реакций, описанных суммарным уравнением, можно охарактеризовать величинами окислительно-восстановительных потенциалов, измеряемых в вольтах. Значения потенциалов показывают, сколько энергии запасается или растрачивается в каждой реакции. Итак, фотосинтез можно рассматривать как процесс образования лучистой энергии Солнца в химическую энергию растительных тканей.
Содержание СО2 в атмосфере остается почти полным, несмотря на то, что углекислый газ расходуется в процессе фотосинтеза. Дело в том, что все растения и животные дышат. В процессе дыхания в митохондриях кислород, поглощаемый из атмосферы живыми тканями, используется для окисления углеводов и других компонентов тканей с образованием в конечном счете двуокиси углерода и воды и с сопутствующим выделением энергии. Высвобождающаяся энергия запасается в высокоэнергетические соединения — аденозинтрифосфат (АТФ), который и используется организмом для выполнения всех жизненных функций. Таким образом дыхание приводит к расходованию органических веществ и кислорода и увеличивает содержание СО2 на н планете. На процессы дыхания во всех живых организмах и на сжигание всех видов топлива, содержащих углерод, в совокупности расходуется в масштабах Земли в среднем около 10000 тонн 02 в секунду. При такой скорости.потребления весь кислород в атмосфере должен бы иссякнуть примерно, через 3000 лет. К счастью для нас, расход органических веществ и атомного кислорода уравновешивается созданием углеводов и кислорода в результате фотосинтеза. В идеальных условиях скорость фотосинтеза в зеленых тканях растений примерно в 30 раз превышает скорость дыхания в тех же тканях, таким образом, фотосинтез служит важным фактором, регулирующим содержание 02 на Земле.
История открытия фотосинтеза
В начале XVII в. фламандский врач Ван Гельмонт вырастил в кадке с землей дерево, которое он поливал только дождевой водой. Он заметил, что спустя пять лет, дерево выросло до больших размеров, хотя количество земли в кадке практически не уменьшилось. Ван Гельмонт, естественно, сделал вывод, что материал, из которого образовалось дерево произошел из воды, использованной для полива. В 1777 английский ботаник Стивен Хейлс опубликовал книгу, в которой сообщалось, что в качестве питательного вещества, необходимого для роста, растения используют главным образом воздух. В тот же период знаменитый английский химик Джозеф Пристли (он был одним из первооткрывателей кислорода) провел серию опытов по горению и дыханию и пришел к выводу о том, что зелёные растения способны совершать все те дыхательные процессы, которые были обнаружены в тканях животных. Пристли сжигал свечу в замкнутом объеме воздуха, и обнаруживал, что получавшийся при этом воздух уже не может поддерживать горение. Мышь, помещенная в такой сосуд, умирала. Однако веточка мяты продолжала жить в воздухе неделями. В заключение Пристли обнаружил, что в воздухе, восстановленном веточкой мяты, вновь стала гореть свеча, могла дышать мышь. Теперь мы знаем, что свеча, сгорая, потребляла кислород из замкнутого объема воздуха, но затем воздух снова насыщался кислородом благодаря фотосинтезу, происходившему в оставленной веточке мяты. Спустя несколько лет голландский врач Ингенхауз обнаружил, что растения окисляют кислород лишь на солнечном свету и что только их зеленые части обеспечивают выделение кислорода. Жан Сенебье, занимавший пост министра, подтвердил данные Ингенхауза и продолжил исследование, показав, что в качестве питательного вещества растения используют двуокись углерода, растворенную в воде. В начале XIX века другой швейцарский исследователь де Соседи изучал количественные взаимосвязи между поглощенной растением углекислотой, с одной стороны, и синтезированными органическими веществами и кислородом — с другой. В результате своих опытов он пришел к выводу, что вода также потребляется растением при ассимиляции СО2. В 1817 г. два французских химика, Пельтье и Каванту, выделили из листьев зеленое вещество и назвали его хлорофиллом. Следующей важной вехой в истории изучения фотосинтеза было сделанное в 1845 г. немецким физиком Робертом Майером утверждение о том, что зеленые растения преобразуют энергию, солнечного света в химическую энергию. Представления о фотосинтезе, сложившиеся к середине прошлого века, можно выразить следующим соотношением:
Зеленое растение
СО2+ Н2 О + Свет? О2 + орг. вещества +химическая энергия
Отношение количества С02, поглощенного при фотосинтезе, к количеству выделенного 02, точно измерил французский физиолог растений Бусэнго. В 1864 г. он обнаружил, что фотосинтетическое отношение, т.е. отношение объема выделенного 02 к объему поглощенного С02, почти равно единице. В том же году немецкий ботаник Закс (открывший также у растений дыхание) продемонстрировал образование зерен крахмала при фотосинтезе. Закс помещал зеленые листья на несколько часов в темноту для того, чтобы они израсходовали накопленный в них крахмал. Затем он выносил листья на свет, но при этом освещал лишь половину каждого листа, оставляя другую половину листа в темноте. Спустя некоторое время весь лист целиком обрабатывали парами йода. В результате освещенная часть листа становилась темно-фиолетовой, что свидетельствовало об образовании комплекса крахмала с йодом, тогда как цвет другой половины листа не изменялся. Прямую связь между выделением кислорода и хлоропластами в зеленых листьях, а также соответствие спектра действия фотосинтеза спектру поглощеных хлоропластами установил в 1880 г. Энгельман. Он поместил нитевидную зеленую водоросль имеющую спирально извитые хлоропласты, на предметное стекло, освещая его узким и широким пучком белого света. Вместе с водорослью на предметное стекло наносилась суспензия клеток подвижных бактерий, чувствительных к концентрации кислорода. Предметное стекло помещали в камеру без воздуха и освещали. В этих условиях подвижные бактерии должны были перемещаться в ту часть, где концентрация 02 была выше. После прошествия некоторого времени образец рассматривали под микроскопом и подсчитывали распределение бактериопопуляции. Оказалось, что бактерии концентрировались вокруг зеленых полосок в нитевидной водоросли. В другой серии опытов Энгельман освещал водоросли лучами разного спектрального состава, установив призму между источником света и предметным столиком микроскопа. Наибольшее число бактерий в этом случае скапливалось вокруг тех участков водоросли, которые освещались синим и красным областями спектра. Находящиеся в водорослях хлорофиллы поглощали синий и красный свет. Поскольку к тому времени было уже известно, что для фотосинтеза необходимо поглощение света, Энгельман заключил, что хлорофиллы участвуют в синтезе в качестве пигментов, являющихся активными фоторецепторами. Уровень знаний о фотосинтезе в начале нашего века можно представить следующим образом.
СО2 + Н2О + Свет –О2 + Крахмал + Химическая энергия
Итак, к началу нашего века суммарная реакция фотосинтеза была уже известна. Однако биохимия находилась не на таком высоком уровне, чтобы полно раскрыть механизмы восстановления двуокиси углерода до углеводов. К сожалению, следует признать, что и теперь еще некоторые аспекты фотосинтеза изучены довольно плохо. Издавна делались попытки исследовать влияние интенсивности света, температуры, концентрации углекислоты и т… п. на общий выход фотосинтеза. И хотя в этих работах исследовались растения самых разных видов, большинство измерений было выполнено на одноклеточных зеленых водорослях и на одноклеточной жгутиковой водоросли Эвглена. Одноклеточные организмы удобнее для качественного исследования, поскольку их можно выращивать во всех лабораториях при вполне стандартных условиях. Они могут быть равномерно суспензированы, т. е. взвешены в водных буферных растворах, и нужный объем такой суспензии, или взвеси, можно брать такой дозировки, точно так же, как при работе с обычными растениями. Хлоропласты для опытов лучше всего выделять из листьев высших растений. Чаще всего используют шпинат, потому что его легко выращивать и свежие листья удобны для проведения исследований; иногда используются листья гороха и салата-латука.
Поскольку СО2 хорошо растворяется в воде, а О2 относительно нерастворим в воде, то при фотосинтезе в замкнутой системе давление газа в этой системе может изменяться. Поэтому влияние света на фотосинтетические системы часто исследуют с помощью респиратора Варбурга, позволяющего регистрировать пороговые изменения объема 02 в системе. Впервые респиратор Варбурга был использован применительно к фотосинтезу в 1920г. Для измерения потребления или выделения кислорода в ходе реакции удобнее пользоваться другим прибором — кислородным электродом. В основе этого устройства лежит использование полярографического метода. Кислородный электрод обладает достаточной чувствительностью для того, чтобы обнаружить в таких низких концентрациях как 0,01 ммоль в 1 л. Прибор состоит из катода достаточно тонкой платиновой проволоки, герметично впрессованной в пластину анода, представляющего собой кольцо из серебряной проволоки, погруженной в насыщенный раствор. Электроды отделены от смеси, в которой протекает реакция, мембраной, проницаемой для 02. Реакционная система находится в пластмассовом или стеклянном сосуде и постоянно перемешивается вращающимся стержневым магнитом. Когда к электродам приложено напряжение, платиновый электрод становится отрицательным по отношению к стандартному электроду, кислород в растворе электролитически восстанавливается. При напряжении от 0,5 до 0,8 В величина электрического тока линейно зависит от парциального давления кислорода в растворе. Обычно с кислородным электродом работают при напряжении около 0,6 В. Электрический ток измеряют, присоединив электрод к подходящей регистрирующей системе. Электрод вместе с реакционной смесью орошают потоком воды от термостата. С помощью кислородного электрода измеряют действие света и различных химических веществ на фотосинтез. Преимущество кислородного электрода перед аппаратом Варбурга состоит в том, что кислородный электрод позволяет быстро и непрерывно регистрировать изменения содержания О2 в системе. С другой стороны, в приборе Варбурга можно одновременно исследовать до 20 образцов с различными реакционными смесями, тогда как при работе с кислородным электродом образцы приходится анализировать поочередно.
Примерно до начала 1930-х годов многие исследователи в этой области полагали, что первичная реакция фотосинтеза заключается в расщеплении двуокиси углерода под действием света на углерод и кислород с последующим восстановлением углерода до углеводов с участием воды в ходе нескольких последовательных реакций. Точка зрения изменилась в 1930-х годах в результате двух важных открытий. Во-первых, были описаны разновидности бактерий, способных ассимилировать и синтезировать углеводы, не используя для этого энергию света. Затем, голландский микробиолог Ван Нил сравнил процессы фотосинтеза у бактерий и показал, что некоторые бактерии могут ассимилировать С02 на свету, не выделяя при этом кислорода. Такие бактерии способны к фотосинтезу лишь при наличии подходящего субстрата-донора водорода. Ван Нил предполагал, что фотосинтез зеленых растений и водорослей является частным случаем, когда кислород в фотосинтезе происходит из воды, а не из углекислоты.
Второе важное открытие сделал в 1937 г. Р. Хил в Кембриджском университете. С помощью дифференциального центрифугирования гомогената тканей листа он отделил фотосинтезирующие частицы (хлоропласты) от дыхательных частиц. Полученные Xиллом хлоропласты при освещении сами по себе не выделяли кислорода (возможно, из-за того, что они были повреждены при разделении). Однако они начинали выделять кислород на свету, если в суспензию вносили подходящие акцепторы электрона (окислители), например ферриоксалат калия или феррицианид калия. При выделении одной молекулы 02 фотохимически восстанавливались четыре эквивалента окислителя. Позднее было обнаружено, что многие хиноны и красители восстанавливаются хлоропластами на свету. Однако хлоропласты не могли восстановить СО2, природный акцептор электронов при фотосинтезе. Это явление, известное теперь как реакция Хилла, представляет собой индуцируемый светом перенос электронов от воды к нефизиологическим окислителям (реагентам Хилла) против градиента химического потенциала. Значение реакции Хилла состоит в том, что она продемонстрировала возможность разделения двух процессов — фотохимического выделения кислорода и восстановления углекислоты при фотосинтезе.
Разложение воды, приводящее к выделению свободного кислорода при фотосинтезе, было установлено Рубеном и Каменом, в Калифорнии в 1941 г. Они поместили фотосинтезирующие клетки в воду, обогащенную изотопом кислорода, имеющим массу 18 атомных единиц 180. Изотопный состав кислорода, выделенного клетками, соответствовал составу воды, но не С02. Кроме того, Камен и Рубен открыли радиоактивный изотоп 18О, который впоследствии успешно использовали Бассэт, Бенсон Вин, изучавшие путь превращения углекислоты при фотосинтезе. Кальвин и его сотрудник установили, что восстановление углекислоты до сахаров происходит в результате темновых ферментативных процессов, причем для восстановления одной молекулы углекислоты требуются две молекулы восстановленного АДФ и три молекулы АТФ. К тому времени роль АТФ и пиридиннуклеотидов в дыхании тканей была установлена. Возможность фотосинтетического восстановления АДФ до АТФ выделенными хлорофиллами была доказана в 1951 г. в трех разных лабораториях. В 1954 г. Арнон, Аллен продемонстрировали фотосинтез — они наблюдали ассимиляцию С02 и 02 выделенными хлоропластами шпината. В течение следующего десятилетия из хлоропластов удалось выделить белки, участвующие в переносе электронов в синтезе -ферредоксин, пластоцианин, ферроАТФ-редуктазу, цитохромы и т. д.
Таким образом, в здоровых зеленых листьях, под действием света образуются АДФ и АТФ и энергия гидросвязей используются для восстановления С02 до углеводов в присутствии ферментов, причем активность ферментов регулируется светом.
Лимитирующие факторы
Интенсивность, или скорость процесса фотосинтеза в растении зависит от ряда внутренних и внешних факторов. Из внутренних факторов наиболее важное значение имеют структура листа и содержание в нем хлорофилла, скорость накопления продуктов фотосинтеза в хлоропластах, влияние ферментов, а также наличие малых концентраций необходимых неорганических веществ. Внешние параметры — это количество и качество света, попадающего на листья, температура окружающей среды, концентрация углекислоты и кислорода в атмосфере вблизи растения.
Скорость фотосинтеза возрастает линейно, или прямо пропорционально увеличению интенсивности света. По мере дальнейшего увеличения интенсивности света нарастание фотосинтеза становится все менее и менее выраженным, и, наконец, прекращается, когда освещенность достигает определенного уровня 10000 люкс. Дальнейшее увеличение интенсивности света уже не влияет на скорость фотосинтеза. Область стабильной скорости фотосинтеза называется областью светонасыщения. Если нужно увеличить скорость фотосинтеза в этой области, следует изменять не интенсивность света, а какие-либо другие факторы. Интенсивность солнечного света, попадающего в ясный летний день на поверхность земли, во многих местах нашей планеты составляет примерно 100000 люкс. Следовательно, растениям, за исключением тех, которые растут в густых лесах и в тени, падающего солнечного света бывает достаточно для насыщения их фотосинтетической активности (энергия квантов, соответствующих крайним участкам видимого диапазона – фиолетового и красного, различается всего лишь в два раза, и все фотоны этого диапазона в принципе способны осуществить запуск фотосинтеза).
В случае низких интенсивностей света скорость фотосинтеза при 15 и 25°С одинакова. Реакции, протекающие при таких интенсивностях света, которые соответствуют области лимитирования света, подобно истинным фотохимическим реакциям, не чувствительны к температурам. Однако при более высоких интенсивностях скорость фотосинтеза при 25°С гораздо выше, чем при 15°С. Следовательно, в области светового насыщения уровень фотосинтеза зависит не только от поглощения фотонов, но и от других факторов. Большинство растений в умеренном климате хорошо функционируют в интервале температур от 10 до 35°С, наиболее благоприятные условия — это температура около 25°С.
В области лимитирования светом скорость фотосинтеза не изменяется при уменьшении концентрации СО2. Отсюда можно сделать вывод, что С02 участвует непосредственно в фотохимической реакции. В то же время при более высоких интенсивностях освещения, лежащих за пределами области лимитирования, фотосинтез существенно возрастает при увеличении концентрации СО2. У некоторых зерновых культур фотосинтез линейно возрастал при увеличении концентрации СО2 до 0,5%. (эти измерения проводили в кратковременных опытах, поскольку длительное воздействие высоких концентраций СО2 повреждает листы). Высоких значений скорость фотосинтеза достигает при содержании С02 около 0,1%. Средняя концентрация углекислоты в атмосфере составляет от 0,03%. Поэтому в обычных условиях растениям не хватает СО2 для того, чтобы с максимальной эффективностью использовать попадающий на них солнечный свет. Если помещенное в замкнутый объем растение освещать светом насыщающей интенсивности, то концентрация СО2 в объеме воздуха будет постепенно уменьшаться и достигнет постоянного уровня, известного под названием «С02 компенсационного пункта». В этой точке появление СО2 при фотосинтезе уравновешивается выделением О2 в результате дыхания (темнового и светового). У растений разных видов положения компенсационных пунктов различны.
--PAGE_BREAK--Световые и темновые реакции.
Еще в 1905 г. английский физиолог растений Ф. Ф. Блекмэн, интерпретируя форму кривой светового насыщения фотосинтеза, высказал предположение, что фотосинтез представляет собой двухстадийный процесс, включающий фотохимическую, т.е. светочувствительную реакцию и нефотохимическую, т. е. темновую, реакцию. Темновая реакция, будучи ферментативной, протекает медленнее, чем световая реакция, и поэтому при высоких интенсивностях света скорость фотосинтеза полностью определяется скоростью темновой реакции. Световая реакция либо вообще не зависит от температуры, либо зависимость эта выражена очень слабо, тогда темновая реакция, как и все ферментативные процессы, зависит от температуры в довольно значительно и степени. Следует ясно представлять себе, что реакция, называемая темновой, может протекать как в темноте, так и на свету. Световую и темновую реакции можно разделить, используя вспышки света, длящиеся краткие доли секунды. Вспышки света длительностью меньше одной миллисекунды (10-3 с) можно получить либо с помощью механического приспособления, поставив на пути пучка постоянного света вращающийся диск со щелью, либо электрически, заряжая конденсатор и разряжая его через вакуумную или газоразрядную лампу. В качестве источников света пользуются также рубиновыми лазерами с длиной волны излучения 694 нм. В 1932 г. Эмерсон и Арнольд освещали суспензию клеток вспышками света от газоразрядной лампы с длительностью около 10-3с. Они измеряли скорость выделения кислорода в зависимости от энергии вспышек, длительности темнового промежутка между вспышками и температуры суспензии клеток. При увеличении интенсивности вспышек насыщение фотосинтеза в нормальных клетках наступало, когда выделялась одна молекула 02 на 2500 молекул хлорофилла. Эмерсон и Арнольд сделали вывод, что максимальный выход фотосинтеза определяется не числом молекул хлорофилла, поглощающих свет, а числом молекул фермента, катализирующего темновую реакцию. Они также обнаружили, что при увеличении темновых интервалов между последовательными вспышками за пределы 0,06 с выход кислорода в расчете на одну вспышку уже не зависел от длительности темнового интервала, тогда как при более коротких промежутках он возрастал с увеличением длительности темнового интервала (от 0 до 0,06 с). Таким образом, темновая реакция, которая определяет уровень насыщения фотосинтеза, завершается примерно за 0,06 с. На основе этих данных было рассчитано, что среднее время, характеризующее скорость реакции, составило около 0,02 с при 25°С.
Структурная и биохимическая организация аппарата фотосинтеза
Современные представления о структурной и функциональной организации фотосинтетического аппарата включают широкий круг вопросов, связанных с характеристикой химического состава пластид, спецификой их структурной организации, физиолого-генетическими закономерностями биогенеза этих органоидов и их взаимоотношениями с другими функциональными структурами клетки. У наземных растений специальным органом фотосинтетической деятельности служит лист, где локализованы специализированные структуры клетки — хлоропласты, содержащие пигменты и другие компоненты, необходимые для процессов поглощения и преобразования энергии света в химический потенциал. Кроме листа функционально активные хлоропласты присутствуют в стеблях растений, черешках, остях и чешуях колоса и даже в освещаемых корнях ряда растений. Однако именно лист был сформирован в ходе длительной эволюции как специальный орган для выполнения основной функции зеленого растения – фотосинтеза, поэтому анатомия листа, расположение хлорофиллсодержащих клеток и тканей, их соотношение с другими элементами морфемной структуры листа подчинены наиболее эффективному течению процесса фотосинтеза, и они в наибольшей степени подвергаются интенсивным изменениям в условиях экологического стресса.
В связи с этим проблему структурно — функциональной организации фотосинтетического аппарата целесообразно рассмотреть в двух основных уровнях — на уровне листа как органа фотосинтеза и хлоропластов, где целиком сосредоточен весь механизм фотосинтеза.
Организация фотосинтетического аппарата на уровне листа может быть рассмотрена на основе анализа его мезострктуры. Понятие «мезоструктура» было предложено в 1975 году. По представлениям о структурной и функциональной особенностях фотоситнетического аппарата с характеристикой химического состава, структурной организации, физиолого-генетическими особенностями биогенеза этих органоидов и их взаимоотношениями с другими функциональными структурами специальным органом фотосинтетического процесса является лист, где локализованы специализированные образования — хлоропласты, содержащие пигменты, необходимые для процессов поглощения и преобразования света в химический потенциал. Кроме того, активные хлоропласты присутствуют в стеблях, остях и чешуях колоса и даже в освещенных частях корней некоторых растений. Однако именно лист был сформирован всем ходом эволюции как специальный орган для выполнения основной функции зеленого растения — фотосинтеза.
Мезоструктура включает систему морфофизиологических характеристик фотосинтетического аппарата листа, хлоренхимы и клезофилла. Основные показатели мезоструктуры фотосинте-
тического аппарата (по А. Т. Мокроносову) включают: площадь, число клеток, хлорофилл, белок, объем клетки, количество хлоропластов в клетке, объем хлоропласта, площадь сечения хлоропласта и его поверхность. Анализ мезоструктуры и функциональной активности фотосинтетического аппарата у многих видов растений помогают определить наиболее часто встречающиеся значения исследуемых показателей и пределы варьирования отдельных характеристик. Согласно этим данным, основные показатели мезоструктуры фотосинтетического аппарата (Мокроносов, 19В1):
I — площадь листа;
II — число клеток на 1 см2,
III — хлорофилл на 1 дм2, ключевые ферменты на 1 дм2, объем клетки, тыс. мкм2, число хлоропластов в клетке;
IV — объем хлоропластов, площадь проекции хлоропласта, мкм2, поверхность хлоропласта, мкм2.
Среднее число хлоропластов у закончившего рост листа обычно достигает 10-30, у некоторых видов оно превышает 400. Это соответствует млн хлоропластов в расчете на 1 см2площади листа. Хлоропласты сосредоточены в клетках различных тканей в количестве 15 – 80 штук на клетку. Средний объем хлоропласта — один мкм2. У большинства растений суммарный объем всех хлоропластов составляет 10-20%, у древесных растений — до 35% объема клетки. Отношение общей поверхности хлоропластов к площади листа находится в пределах 3-8. В одном хлоропласте содержится разное количество молекул хлорофилла, у тенелюбивых видов их число возрастает. Приведенные выше показатели могут значительно варьировать в зависимости от физиологического состояния и экологических условий роста растений. По данным А. Т. Мокроносова, в молодом листе активизация фотосинтеза при удалении 50-80% листа обеспечивается увеличением числа хлоропластов в клетке без изменения их индивидуальной активности, в то время как в листе, окончившем рост, усиление фотосинтеза после дефолиации происходит за счет повышения активности каждого хлоропласта без изменения их числа. Анализ мезоструктуры показал, что адаптация к условиям освещения вызывает перестройку, которая оптимизирует светопоглошающие свойства листа.
Хлоропласты имеют наиболее высокую степень организации внутренних мембранных структур по сравнению с другими органоидами клетки. По степени упорядоченности структур хлоропласты можно сравнить только с рецепторными клетками сетчатки глаза, также выполняющими функцию преобразования световой энергии. Высокая степень организации внутренней структуры хлоропласта определяется рядом моментов:
1) необходимостью пространственного разделения восстановленных и окисленных фотопродуктов, возникающих в результате первичных актов разделения заряда в реакционном центре;
2) необходимостью строгой упорядоченности компонентов реакционного центра, где сопряжёны быстропротекающие фотофизиологические и более медленные энзиматические реакции: преобразование энергии фотовозбужденной молекулы пигмента требует ее определенной ориентации по отношению к химическому акцептору энергии, что предполагает наличие определенных структур, где пигмент и акцептор жестко ориентированы друг относительно друга;
3) пространственная организация электронно-транспортной цепи требует последовательной и строго ориентированной организации переносчиков в мембране, обеспечивающей возможность быстрого и регулируемого транспорта электронов и протонов;
4) для сопряжения транспорта электронов и синтеза АТФ требуется определенным образом организованная система хлоропластов.
Липопротеидные мембраны как структурная основа энергетических процессов возникают на самых ранних этапах эволюции, предполагают, что основные липидные компоненты мембран — фосфолипиды — образовались в определенных биологических условиях. Формирование липидных комплексов обусловило возможность включения в них различных соединений, что по-видимому, явилось основой первичных каталитических функций данных структур.
Проведенные в последние годы электронномикроскопические исследования обнаружили организованные мембранные структуры у организмов, стоящих на самой низкой ступени эволюции. У некоторых бактерий мембранные фотесинтезирующие структуры клеток тесно упакованных органелл расположены по периферии клетки и связаны с цитоплазматическими мембранами; кроме того, в клетках зеленых водорослей процесс фотосинтеза связан с системой двойных замкнутых мембран — тилакоидов, локализованных в периферической части клетки. У данной группы фотосинтетических организмов впервые появляется хлорофилл, а образование специализированных органелл – хлоропластов встречается у криптофитовых водорослей. В них находятся по два хлоропласта, содержащих от одного до нескольких тилакоидов. Сходное строение фотосинтетического аппарата имеет место и у других групп водорослей: красных, бурых, и др. В процессе эволюции мембранная структура фотосинтетического процесса усложняется.
Микроскопические исследования хлоропласта, техника криоскопии позволили сформулировать пространственную модель объемной организация хлоропластов. Наиболее известна гранулярно-решетчатая модель Дж. Хеслоп-Харрисона (1964).
Таким образом, фотосинтез – это сложный процесс преобразования световой энергии в энергию химических связей органических веществ, необходимых для жизнедеятельности как самих фотосинтезирующих организмов, так и других организмов, не способных к самостоятельному синтезу органических веществ.
Изучение проблем фотосинтеза, помимо общебиологических, имеет и прикладное значение. В частности, проблемы питания, создания систем жизнеобеспечения при космических исследованиях, использования фотосинтезирующих организмов для создания различных биотехнических устройств непосредственно связаны с фотосинтезом.
Список литературы
1. Д.Халл, К.Рао «Фотосинтез». М.,1983
2. Мокроносов А.Г. «Фотосинтетическая реакция и целостность растительного организма». М.,1983
3. Мокроносов А.Г., Гавриленко В.Ф. «Фотосинтез: физиолого – экологические и биохимические аспекты» М.,1992
4. «Физиология фотосинтеза» под ред. Ничипоровича А.А., М.,1982
5. Вечер А.С. «Пластиды растнеий»
6. Виноградов А.П. «Изотопы кислорода и фотосинтез»
7. Годнев Т.Н. «Хлорофилл и его строение».
8. Гуринович Г.П., Севченко А.Н., Соловьев К.Н. «Спектроскопия хлорофилла»
9. Красновский А.А. «Преобразование энергии света при фотосинтезе»
Для подготовки данной работы были использованы материалы с сайта www.ronl.ru/
www.ronl.ru
Реферат на тему: Фотосинтез
СОДЕРЖАНИЕ
История фотосинтеза
В течение тысячелетий люди считали, что питается растение исключительно благодаря корням, поглощая с их помощью все необходимые вещества из почвы. Проверить эту точку зрения взялся в начале девятнадцатого века голландский натуралист Ян Ван Гельмонт. Он взвесил землю в горшке и посадил туда побег ивы. В течение пяти лет он поливал деревце, а затем высушил землю и взвесил её и растение. Ива весила семьдесят пять килограмм, а вес земли изменился всего на несколько сот граммов. Вывод учёного был таков — растения получают питательные вещества, прежде всего, не из почвы, а из воды. На два столетия в науке утвердилась теория водного питания растений. Листья, по этой теории, лишь помогали растению испарять излишнюю влагу.
К самому неожиданному, но правильному предположению о воздушном питании растений ученые пришли лишь к началу девятнадцатого века. Важную роль в понимании этого процесса сыграло открытие, совершенное английским химиком Джозефом Пристли в 1771 году. Он поставил опыт, в результате которого он сделал вывод: растения очищают воздух и делают его пригодным для дыхания. Позднее выяснилось: для того, чтобы растение очищало воздух, необходим свет. Десять лет спустя, учёные поняли, что растение не просто превращает углекислый газ в кислород. Углекислый газ необходим растениям для жизни, он служит для них настоящей пищей (вместе с водой и минеральными солями).
Воздушное питание растений называется фотосинтезом. Кислород в процессе фотосинтеза выделяется в качестве необычного продукта. Миллиарды лет назад на земле не было свободного кислорода. Весь кислород, которым дышат почти все живые существа нашей планеты, выделен растениями в процессе фотосинтеза. Фотосинтез сумел изменить весь облик нашей планеты!
Начиная с семидесятых годов прошлого столетия, крупные успехи в области фотосинтеза были получены в России. Работами русских учёных Пуриевича, Ивановского, Риктера, Иванова, Костычева были изучены многие стороны этого процесса. Значение фотосинтеза не осознавалось до сравнительно недавнего времени. Аристотель и другие учёные Греции, наблюдая, что жизненные процессы животных зависят от потребления пищи,
полагали, что растения добывают свою «пищу» из почвы. Немногим более трехсот лет назад в одном из первых тщательно продуманных биологических экспериментов голландский врач Ян Ван Гельмонт представил доказательства того, что не одна почва кормит растение. Ван Гельмонт выращивал маленькое дерево ивы в глиняном горшке, добавляя в него только воду. Через пять лет масса игл увеличилась на 74,4 кг, в то время, как масса почвы уменьшилась только на 57 гр.
В конце XVIII века английский ученый Джозеф Пристли сообщил, что он «случайно обнаружил метод исправления воздуха, который был испорчен горением свечей». 17 августа 1771 г. Пристли «… поместил живую веточку мяты в закрытый сосуд, в котором горела восковая свеча», а 21 числа того же месяца обнаружил, что «… другая свеча снова могла гореть в этом же сосуде». «Исправляющим началом, которым для этих целей пользуется природа, — полагал Пристли, — было растение». Он расширил свои наблюдения и скоро показал, что воздух, «исправляемый» растением, не был «совсем не подходящим для мыши».
Опыты Пристли впервые позволили объяснить, почему воздух на Земле остается «чистым» и может поддерживать жизнь, несмотря на горение бесчисленных огней и дыхание множества живых организмов. Он говорил: «Благодаря этим открытиям мы уверены, что растения произрастают не напрасно, а очищают и облагораживают нашу атмосферу».
Позднее голландский врач Ян Ингенхауз (1730-1799) подтвердил работу Пристли и показал, что воздух «исправляется» только на солнечном свету и только зелеными частями растения. В 1796 году Ингенхауз предположил, что углекислота разлагается при фотосинтезе на С и О2, а О2 выделяется в виде газа. В последствие было обнаружено, что соотношение атомов углерода, водорода и кислорода в сахарах и крахмале таково, что один атом углерода приходится на одну молекулу воды, на что и указывает слово «углеводы». Считалось общепринятым, что углеводы образуются из С и Н2О, а О2 выделяется из углекислоты. Это вполне разумная гипотеза была широко признана, но, как позднее выяснилось, она была совершенно неверной.
Исследователем, который опроверг эту общепринятую теорию, был Корнелиус ван Ниль из Стамфордского университета, когда он, будучи еще студентом — дипломником, исследовал метаболизм различных фотосинтезирующих бактерий. Одна группа таких бактерий, а именно пурпурные серные бактерии, восстанавливает С до углеводов, но не выделяет О2. Пурпурным серным бактериям для фотосинтеза необходим сероводород. В результате фотосинтеза внутри бактериальных клеток накапливаются частицы серы. Ван Ниль обнаружил, что для этих бактерий уравнение фотосинтеза может быть записано как:
свет
С О2 + 2Н2S → (Ch3O) + Н2О + 2S
Этот факт не привлекал внимания исследователей до тех пор, пока ван Ниль не сделал смелого сообщения и не предложил следующего суммарного уравнения фотосинтеза:
свет
С О2 + 2Н2А → (Ch3O) + Н2О + 2А
В этом уравнении Н2А представляет собой либо воду, либо другое окисляемое вещество, например, сероводород или свободный Н2. У зеленых растений и водорослей Н2А = Н2О. То есть ван Ниль предположил, что Н2О, а не углекислота, разлагается при фотосинтезе. Эта блестящая идея, выдвинутая в тридцатые годы, экспериментально была доказана позднее, когда исследователи, использую тяжелый изотоп О2(18О2), проследили путь кислорода от воды до газообразного состояния:
свет
С О2 + 2Н218О2 → (Ch3O) + Н2О + 18О2
Таким образом, для водорослей или зеленых растений, у которых вода служит донором электронов, суммарное уравнение фотосинтеза записывается следующим образом:
свет
6СО2 + 12Н2О → C6h22O6 + 6О2 + 6Н2О
Процессы, происходящие в листе
Лист осуществляет три важных процесса – фотосинтез, испарение воды и газообмен. В процессе фотосинтеза в листьях из воды и двуокиси углерода под действием солнечных лучей синтезируются органические вещества. Днем, в результате фотосинтеза и дыхания, растение выделяет кислород и двуокись углерода, а ночью – только двуокись углерода, образующуюся при дыхании.
Большинство растений способно синтезировать хлорофилл при слабом освещении. При прямом солнечном освещении хлорофилл синтезируется быстрее.
Необходимая для фотосинтеза световая энергия в известных пределах поглощается тем больше, чем меньше затемнен лист. Потому у растений в процессе эволюции выработалась способность поворачивать пластину листа к свету так, чтобы на нее падало больше солнечных лучей. Листья на растении располагаются так, чтобы не притеснять друг друга.
Тимирязев доказал, что источником энергии для фотосинтеза служат преимущественно красные лучи спектра. На это указывает спектр поглощения хлорофилла, где наиболее интенсивная полоса поглощения наблюдается в красной, и менее интенсивное – в сине-фиолетовой части.
В хлоропластах вместе с хлорофиллом имеются пигменты каротин и ксантофилл. Оба этих пигмента поглощают синие и, отчасти, зеленые лучи и пропускают красные и желтые. Некоторые ученые приписываю каротину и ксантофиллу роль экранов, защищающих хлорофилл от разрушительного действия синих лучей.
Процесс фотосинтеза слагается из целого ряда последовательных реакций, часть которых протекает с поглощением световой энергии, а часть – в темноте. Устойчивыми окончательными продуктами фотосинтеза являются углеводы (сахара, а затем крахмал), органические кислоты, аминокислоты, белки.
Фотосинтез при различных условиях протекает с разной интенсивностью.
Интенсивность фотосинтеза также зависит от фазы развития растения. Максимальная интенсивность фотосинтеза наблюдается в фазе цветения.
Обычное содержание углекислоты в воздухе составляет 0,03% по объему. Уменьшение содержания углекислоты в воздухе снижает интенсивность фотосинтеза. Повышение содержания углекислоты до 0,5% увеличивает интенсивность фотосинтеза почти пропорционально. Однако при дальнейшем повышении содержания углекислоты, интенсивность фотосинтеза не возрастает, а при 1% — растение страдает.
Растения испаряют или трансперируют очень большое количество воды. Испарение воды является одной из причин восходящего тока. Вследствие испарения воды растением в нем накапливаются минеральные вещества, и происходит полезное для растения понижение температуры во время солнечного нагрева. Иногда трансперация снижает температуру растения на 6о.
Растение регулирует процесс испарения воды посредством работы устьиц. Отложение кутикулы или воскового налета на эпидерме, образование его волосков и другие приспособления направлены к сокращению нерегулируемой трансперации.
Процесс фотосинтеза и постоянное протекающее дыхание живых клеток листа требуют газообмена между внутренними тканями листа и атмосферой. В процессе фотосинтеза из атмосферы поглощается ассимилируемый углекислый газ и возвращается в атмосферу кислородом.
Применение изотопного метода анализа показало, что кислород, возвращаемый в атмосферу (16О) принадлежит воде, а не углекислому газу воздуха, в котором преобладает другой его изотоп — 15О. При дыхании живых клеток (окисление свободным кислородом органических веществ внутри клетки до углекислого газа и воды) необходимо поступление из атмосферы кислорода и возвращение углекислоты. Этот газообмен также в основном осуществляется через устьичный аппарат.
Современные представления о фотосинтезе
В настоящее время известно, что фотосинтез проходит две стадии, но только одна из них – на свету. Доказательства двухстадийности процесса впервые были получены в 1905 году английским физиологом растений Ф.Ф. Блэклином, который исследовал влияние освещенности и температуры на объем фотосинтеза.
На основании экспериментов, Блэклин сделал следующие выводы:
1. Имеется одна группа светозависимых реакций, которые не зависят от температуры. Объем этих реакций в диапазоне низких освещенностей мог возрастать с увеличением освещенности, но не с увеличением температуры.
2. Имеется вторая группа реакций, зависимых от температуры, а не от света. Оказалось, что обе группы реакций необходимы для осуществления фотосинтеза. Увеличение объема только одной группы реакций увеличивает объем всего процесса, но только до того момента, пока вторая группа реакций не начнет удерживать первую. После этого необходимо ускорить вторую группу реакций, чтобы первые могли проходить без ограничений.
Таким образом, было показано, что обе стадии светозависимы: «световая и темновая». Важно помнить, что темновые реакции нормально проходят на свету и нуждаются в продуктах световой стадии. Выражение «темновые реакции» просто означает, что свет как таковой в них не участвует.
Объем темновых реакций возрастает с увеличением температуры, но только до 30о, а затем начинает падать. На основании этого факта предположили, что темновые реакции катализируются ферментами, поскольку обмен ферментативных реакций, таким образом, зависит от температуры. В последствие оказалось, что данный вывод был сделан неправильно.
На первой стадии фотосинтеза (световые реакции) энергия света используется для образования АТР (молекула аденозин-трифосфата) и высокоэнергетических переносчиков электронов. На второй стадии фотосинтеза (темновые реакции) энергетические продукты, образовавшиеся в световых реакциях, используются для восстановления СО2 до простого сахара (глюкозы).
Процесс фотосинтеза все больше и больше привлекает к себе внимание ученых. Наука близка к разрешению важнейшего вопроса – искусственного создания при помощи световой энергии ценных органических веществ из широко распространенных неорганических веществ. Проблема фотосинтеза усиленно разрабатывается ботаниками, химиками, физиками и другими специалистами.
В последнее время уже удалось искусственно получить синтез формальдегида и сахаристых веществ из водных растворов карбонатной кислоты; при этом роль поглотителя световой энергии играли вместо хлорофилла карбонаты кобальта и никеля. Недавно синтезирована молекула хлорофилла.
Успехи науки в области синтеза органических веществ наносят сокрушительный удар по идеалистическому учению – витализму, который доказывал, что для образования органических веществ из неорганических необходима особая «жизненная сила» и что человек не сможет синтезировать сложные органические вещества.
Фотосинтез в растениях осуществляется в хлоропластах. Он включает преобразования энергии (световой процесс), превращение вещества (темновой процесс). Световой процесс происходит в гилакоидах, темновой – в строме хлоропластов. Обобщенное циркулирование фотосинтеза выглядит следующим образом:
свет
6СО2 + 12Н2О → C6h22O6 + 6Н2О + 6О2
Два процесса фотосинтеза выражаются отдельными уравнениями
свет
12Н2О → 12h3 + 6О2 + энергия АТР
(световой процесс)
свет
12h3 + 6О2 + энергия АТР → С6Н12О6 + Н2О
(темновой процесс)
Значение фотосинтеза в природе
Фотосинтез – единственный процесс в биосфере, ведущий к увеличению ее свободной энергии за счет внешнего источника. Запасенная в продуктах фотосинтеза энергия – основной источник энергии для человечества.
Ежегодно в результате фотосинтеза на Земле образуется 150 млрд. тонн органического вещества и выделяется около 200 млн. тонн свободного кислорода.
Круговорот кислорода, углерода и других элементов, вовлекаемых в фотосинтез, поддерживает современный состав атмосферы, необходимый для жизни на Земле. Фотосинтез препятствует увеличению концентрации СО2, предотвращая перегрев Земли вследствие так называемого «парникового эффекта».
Поскольку зеленые растения представляют собой непосредственную или опосредованную базу питания всех других гетеротрофных организмов, фотосинтез удовлетворяет потребность в пище всего живого на нашей планете. Он – важнейшая основа сельского и лесного хозяйства. Хотя возможности воздействия на него еще не велики, но все же и они, в какой то мере используются. При повышении концентрации углекислого газа в воздухе до 0,1% (против 0,3% в естественной атмосфере) удалось, например, повысить урожайность огурцов и томатов втрое.
Квадратный метр поверхности листьев в течение одного часа продуцирует около одного грамма сахара; это значит, что все растения, по приблизительной оценке, изымают из атмосферы от 100 до 200 млрд. тонн С в год. Около 60% этого количества поглощают леса, занимающие 30% непокрытой льдами поверхности суши, 32% — окультуренные земли, а оставшиеся 8% — растения степей и пустынных мест, а также городов и поселков.
Зеленое растение способно не только использовать углекислый газ и создавать сахар, но и превращать азотные соединения, и соединения серы в вещества, слагающие его тело. Через корневую систему растение получает растворенные в почвенной воде ионы нитратов и перерабатывает их в своих клетках в аминокислоты – основные компоненты всех белковых соединений. Компоненты жиров также возникают из соединений, образующихся в процессах обмена веществ и энергии. Из жирных кислот и глицерина возникают жиры и масла, которые служат для растения, главным образом, запасными веществами. В семенах приблизительно 80% всех растений, в качестве богатого энергией запасного вещества, содержатся жиры. Получение семян, жиров и масел играет важную роль в сельскохозяйственной и пищевой промышленности.
СПИСОК ЛИТЕРАТУРЫ
referati-besplatno.ru
works.tarefer.ru


