Доклад: Бактериофаги, их строение, классификация, применение. Реферат бактериофаги
Доклад - Бактериофаги - Биология и химия
Бактериофагами называют вирусы, живущие в бактериях.
Бактериофаги (от бактерии и греч. phagos — пожиратель; буквально — пожиратели бактерий), фаги, бактериальные вирусы, вызывающие разрушение (лизис) бактерий и других микроорганизмов. Бактериофаги размножаются в клетках, лизируют их и переходят в др., как правило, молодые, растущие клетки. Впервые перевиваемый лизис бактерий (сибиреязвенной палочки) наблюдал в 1898 русский микробиолог Н. Ф. Гамалея. В 1915 английский учёный Ф. Туорт описал это же явление у гнойного стафилококка, а в 1917 французский учёный Ф. Д'Эрелль назвал литический агент, проходящий через бактериальные фильтры.
Строение и химический состав. Частицы многих бактериофагов состоят из головки округлой, гексагональной или палочковидной формы диаметром 45—140 нм и отростка толщиной 10—40 и длиной 100—200 нм. Другие бактериофаги не имеют отростка; одни из них округлы, другие — нитевидны, размером 8х800 нм. Содержимое головки состоит преимущественно из дезоксирибонуклеиновой кислоты (ДНК) (длина её нити во много раз превышает размер головки и достигает 60—70 мкм, эта нить плотно скручена в головке) или рибонуклеиновой кислоты (РНК) и небольшого количества (около 3%) белка и некоторых других веществ. Отросток имеет вид полой трубки, окруженной чехлом, содержащим сократительные белки, подобные мышечным. У ряда бактериофагов чехол способен сокращаться, обнажая часть стержня. На конце отростка у многих бактериофагов имеется базальная пластинка с несколькими шиловидными или другие формы выступами. От пластинки отходят тонкие длинные нити, которые способствуют прикреплению фага к бактерии. Оболочки головки и отростка состоят из белков. Общее количество белка в частице фага 50—60%, нуклеиновых кислот — 40—50%. Каждый бактериофаг обладает специфическими антигенными свойствами, отличными от антигенов бактерии-хозяина и других фагов. Имеются антигены, общие для ряда фагов (особенно содержащих РНК).
Распространение. Бактериофаги найдены для большинства бактерий, в том числе патогенных и сапрофитных, а также.для актиномицетов (актинофаги) и сине-зелёных водорослей. Встречаются бактериофаги в кишечнике человека и животных, в растениях, почве, водоёмах, сточных водах, навозе и т. д. Бактериофаги почвенных микроорганизмов влияют на течение микробиологических процессов в почве.
Размножение. Бактериофаг прикрепляется своим отростком к бактериальной клетке и, выделяя фермент, растворяет клеточную стенку; затем содержимое его головки через канадец отростка переходит внутрь клетки, где под влиянием нуклеиновой кислоты фага останавливается синтез бактериальных белков, ДНК и РНК и начинается синтез нуклеиновой кислоты, а затем и белков фага. Часть этих белков — ферменты, другая часть образует оболочку зрелой частицы бактериофага Более мелкие, сферические фаги попадают в бактерии без участия отростка. Если клетка бактерии заражена одновременно частицами бактериофага, различающимися между собой по ряду свойств, то среди потомства, кроме частиц, подобных родителям, будут и такие, у которых эти свойства встречаются в новой комбинации, т. к. при размножении бактериофагов наблюдается
рекомбинация — обмен кусками нитей нуклеиновой кислоты, являющейся носителем наследственной информации. Частицы крупных фагов выходят из бактерии, разрушая её, а некоторых мелких и нитевидных — из живых бактерий. Одни бактериофаги весьма специфичны и способны лизировать клетки только одного какого-либо вида микроорганизмов (монофаги), другие — клетки разных видов (полифаги).
Бактериофагов делят на вирулентные, вызывающие лизис клетки с образованием новых частиц, и умеренные (симбиотические), которые адсорбируются клеткой и проникают в неё, но лизиса не вызывают, а остаются в клетке в латентной (скрытой) неинфекционной форме (профаг). Культуры, содержащие латентный фаг, называются лизогенными. Лизогения передаётся потомству бактерии. Лизогенная культура может содержать 2—3 и более фагов; она, как правило, устойчива против находящихся в ней фагов (лишь небольшая часть клеток лизируется и освобождает зрелые фаги). Воздействуя на лизогенную культуру ультрафиолетовыми или рентгеновскими лучами, перекисью водорода и некоторыми другими веществами, можно значительно увеличить количество клеток, освобождающих фаг (т. н. индукция бактериофагов). Лизогения широко распространена среди всех видов бактерий и актиномицетов. В ряде случаев многие свойства лизогенной культуры (токсичность, подвижность бактерий и др.) зависят от наличия в ней определённых профагов. Описано много мутаций бактериофагов, сопровождающихся изменением их литической активности, строения частиц и «колоний», устойчивости против неблагоприятных воздействий и другие свойств. Бактериофаги играют большую роль в изменчивости и эволюции микробов, причём механизмы воздействия их на клетку разные. Бактериофаги могут резко изменять азотфиксирующую способность азотобактера, токсичность и антигенные свойства патогенных бактерий и др.
Практическое значение бактериофагов Некоторые фаги (одни или в сочетании с антибиотиками) применяли для профилактики (фагопрофилактики) и лечения (фаготерапии) ряда бактериальных инфекционных болезней человека (дизентерия, брюшной тиф, холера, чума, стафилококковые и анаэробная инфекции и др.) и животных. Однако антибиотики и другие химиотерапевтические средства оказались эффективнее фагов, в связи с чем применение их с лечебной целью сузилось. Бактериофаги успешно применяются при определении вида бактерий, актиномицетов. Бактериофаги могут вредить производству антибиотиков, аминокислот, молочных продуктов, бактериальных удобрений и в других отраслях микробиологического синтеза. Велико значение бактериофагов для теоретических работ по генетике и молекулярной биологии.
www.ronl.ru
«Бактериофаги»
ФГБОУ ВПО «Марийский Государственный Университет»
Биолого-Химический факультет
кафедра Биохимии и Физиологии
Реферат по Микробиологии и основам Вирусологии
на тему:
Выполнила:
студентка III курса
группы БПГ-21
Чеснокова Елена
Проверила:
доцент, к.б.н.
Гажеева Т.П.
Йошкар-Ола, 2011
Оглавление
Оглавление 2
Вступление 3
Бактериофаги. Их роль в биосфере 4
Строение бактериофагов 6
Взаимодействие бактериофага с бактериальными клетками 7
Жизненный цикл 9
Систематика бактериофагов 10
Применение 11
В медицине 11
В биологии 11
В микробиологической промышленности 12
Основные стадии развития и простейшие методы исследования бактериофагов 13
Список источников информации 17
Вступление
Английский бактериолог Фредерик Туорт1 в статье 1915 года описал инфекционную болезнь стафилококков, инфицирующий агент проходил через фильтры, и его можно было переносить от одной колонии к другой.
Независимо от Фредерика Туорта французско-канадский микробиолог Феликс Д’Эрель2 3 сентября 1917 год сообщил об открытии бактериофагов. Наряду с этим известно, что российский микробиолог Николай Фёдорович Гамалея3 ещё в 1898 году, впервые наблюдал явление лизиса бактерий (сибиреязвенной палочки) под влиянием перевиваемого агента.
После открытия явлений бактериофагии Д’Эрелль развил учение о том, что бактериофаги патогенных бактерий, являясь их паразитами, играют большую роль в патогенезе инфекций, обеспечивая выздоровление больного организма, а затем создания специфического иммунитета. Это положение привлекло к явлению бактериофагии внимание многих исследователей, которые предполагали найти в фагах важное средство борьбы с наиболее опасными инфекционными болезнями человека и животных.
Также Феликс Д’Эрель выдвинул предположение, что бактериофаги имеют корпускулярную природу. Однако только после изобретения электронного микроскопа удалось увидеть и изучить ультраструктуру фагов. Долгое время представления о морфологии и основных особенностях фагов основывались на результатах изучения фагов Т-группы — Т1, Т2,…, Т7, которые размножаются на Е. coli (кишечная палочка) штамма B. Однако с каждым годом появлялись новые данные, касающиеся морфологии и структуры разнообразных фагов, что обусловило необходимость их морфологической классификации.
Бактериофаги. Их роль в биосфере
Б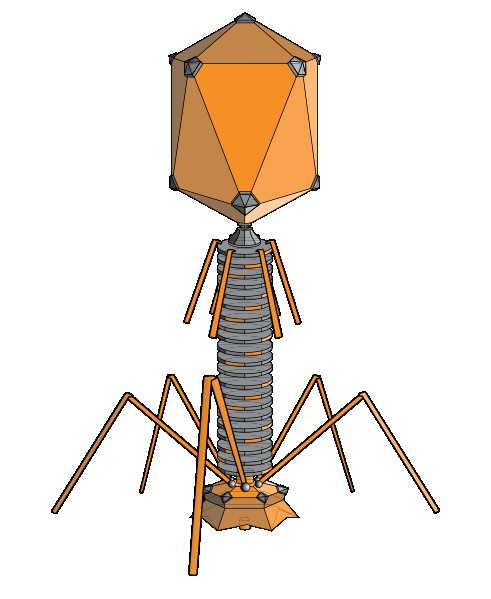 актериофаги (фаги) (от др.-греч. φᾰγω — «пожираю») — вирусы, избирательно поражающие бактериальные клетки. Чаще всего бактериофаги размножаются внутри бактерий и вызывают их лизис4. Как правило, бактериофаг состоит из белковой оболочки и генетического материала одноцепочечной или двуцепочечной нуклеиновой кислоты (ДНК или, реже, РНК). Размер частиц приблизительно от 20 до 200 нм.
актериофаги (фаги) (от др.-греч. φᾰγω — «пожираю») — вирусы, избирательно поражающие бактериальные клетки. Чаще всего бактериофаги размножаются внутри бактерий и вызывают их лизис4. Как правило, бактериофаг состоит из белковой оболочки и генетического материала одноцепочечной или двуцепочечной нуклеиновой кислоты (ДНК или, реже, РНК). Размер частиц приблизительно от 20 до 200 нм.
Структура типичного миовируса бактериофага (рис. 1).
Бактериофаги представляют собой наиболее многочисленную, широко распространенную в биосфере и, предположительно, наиболее эволюционно древнюю группу вирусов. Приблизительный размер популяции фагов составляет более 1030 фаговых частиц.
В природных условиях фаги встречаются в тех местах, где есть чувствительные к ним бактерии. Чем богаче тот или иной субстрат (почва, выделения человека и животных, вода и т. д.) микроорганизмами, тем в большем количестве в нём встречаются соответствующие фаги. Так, фаги, лизирующие клетки всех видов почвенных микроорганизмов, находятся в почвах. Особенно богаты фагами черноземы и почвы, в которые вносились органические удобрения.
Бактериофаги выполняют важную роль в контроле численности микробных популяций, в автолизе стареющих клеток, в переносе бактериальных генов, выступая в качестве векторных «систем».
Действительно, бактериофаги представляют собой один из основных подвижных генетических элементов. Посредством трансдукции они привносят в бактериальный геном новые гены. Было подсчитано, что за 1 секунду могут быть инфицированы 1024 бактерий. Это означает, что постоянный перенос генетического материала распределяется между бактериями, обитающими в сходных условиях.
Высокий уровень специализации, долгосрочное существование, способность быстро репродуцироваться в соответствующем хозяине способствует их сохранению в динамичном балансе среди широкого разнообразия видов бактерий в любой природной экосистеме. Когда подходящий хозяин отсутствует, многие фаги могут сохранять способность к инфицированию на протяжении десятилетий, если не будут уничтожены экстремальными веществами либо условиями внешней среды.
studfiles.net
Реферат Бактериофаг
скачатьРеферат на тему:
План:
Введение- 1 История
- 2 Роль бактериофагов в биосфере
- 3 Строение бактериофагов
- 4 Систематика бактериофагов
- 5 Взаимодействие бактериофага с бактериальными клетками
- 6 Жизненный цикл
- 7 Применение
- 7.1 В медицине
- 7.2 В биологии
Введение
1 — головка, 2 — хвост, 3 — нуклеиновая кислота, 4 — капсид, 5 — «воротничок», 6 — белковый чехол хвоста, 7 — фибрилла хвоста, 8 — шипы, 9 — базальная пластинка
Бактериофа́ги (фаги) (от др.-греч. φᾰγω — «пожираю») — вирусы, избирательно поражающие бактериальные клетки. Чаще всего бактериофаги размножаются внутри бактерий и вызывают их лизис. Как правило, бактериофаг состоит из белковой оболочки и генетического материала одноцепочечной или двуцепочечной нуклеиновой кислоты (ДНК или, реже, РНК). Размер частиц приблизительно от 20 до 200 нм.
1. История
Английский бактериолог Туорт, Фредерик в статье 1915 года описал инфекционную болезнь стафилококков, инфицирующий агент проходил через фильтры, и его можно было переносить от одной колонии к другой.
Независимо от Фредерика Туорта французско-канадский микробиолог Д’Эрель, Феликс 3 сентября 1917 год сообщил об открытии бактериофагов. Наряду с этим известно, что российский микробиолог Гамалея, Николай Фёдорович ещё в 1898 году, впервые наблюдал явление лизиса бактерий (сибиреязвенной палочки) под влиянием перевиваемого агента [1][2].
После открытия явлений бактериофагии Д’Эрелль развил учение о том, что бактериофаги патогенных бактерий, являясь их паразитами, играют большую роль в патогенезе инфекций, обеспечивая выздоровление больного организма, а затем создания специфического иммунитета. Это положение привлекло к явлению бактериофагии внимание многих исследователей, которые предполагали найти в фагах важное средство борьбы с наиболее опасными инфекционными болезнями человека и животных.
Также Феликс Д’Эрель выдвинул предположение, что бактериофаги имеют корпускулярную природу. Однако только после изобретения электронного микроскопа удалось увидеть и изучить ультраструктуру фагов. Долгое время представления о морфологии и основных особенностях фагов основывались на результатах изучения фагов Т-группы — Т1, Т2,…, Т7, которые размножаются на Е. coli штамма B. Однако с каждым годом появлялись новые данные, касающиеся морфологии и структуры разнообразных фагов, что обусловило необходимость их морфологической классификации.
2. Роль бактериофагов в биосфере
Бактериофаги представляют собой наиболее многочисленную, широко распространенную в биосфере и, предположительно, наиболее эволюционно древнюю группу вирусов [3][4]. Приблизительный размер популяции фагов составляет более 1030 фаговых частиц[5].
В природных условиях фаги встречаются в тех местах, где есть чувствительные к ним бактерии. Чем богаче тот или иной субстрат (почва, выделения человека и животных, вода и т. д.) микроорганизмами, тем в большем количестве в нём встречаются соответствующие фаги. Так, фаги, лизирующие клетки всех видов почвенных микроорганизмов, находятся в почвах. Особенно богаты фагами черноземы и почвы, в которые вносились органические удобрения.
Бактериофаги выполняют важную роль в контроле численности микробных популяций, в автолизе стареющих клеток, в переносе бактериальных генов, выступая в качестве векторных «систем»[6].
Действительно, бактериофаги представляют собой один из основных подвижных генетических элементов. Посредством трансдукции они привносят в бактериальный геном новые гены. Было подсчитано, что за 1 секунду могут быть инфицированы 1024 бактерий [7]. Это означает, что постоянный перенос генетического материала распределяется между бактериями, обитающими в сходных условиях.
Высокий уровень специализации, долгосрочное существование, способность быстро репродуцироваться в соответствующем хозяине способствует их сохранению в динамичном балансе среди широкого разнообразия видов бактерий в любой природной экосистеме. Когда подходящий хозяин отсутствует, многие фаги могут сохранять способность к инфицированию на протяжении десятилетий, если не будут уничтожены экстремальными веществами либо условиями внешней среды [8].
3. Строение бактериофагов
Бактериофаги различаются по химической структуре, типу нуклеиновой кислоты, морфологии и характеру взаимодействия с бактериями. По размеру бактериальные вирусы в сотни и тысячи раз меньше микробных клеток.
Типичная фаговая частица (вирион) состоит из головки и хвоста. Длина хвоста обычно в 2 — 4 раза больше диаметра головки. В головке содержится генетический материал — одноцепочечная или двуцепочечная РНК или ДНК с ферментом транскриптазой в неактивном состоянии, окруженная белковой или липопротеиновой оболочкой — капсидом, сохраняющим геном вне клетки [8][9].
Нуклеиновая кислота и капсид вместе составляют нуклеокапсид. Бактериофаги могут иметь икосаэдральный капсид, собранный из множества копий одного или двух специфичных белков. Обычно углы состоят из пентамеров белка, а опора каждой стороны из гексамеров того же или сходного белка. Более того, фаги по форме могут быть сферические, лимоновидные или плеоморфные [10]. Хвост представляет собой белковую трубку — продолжение белковой оболочки головки, в основании хвоста имеется АТФаза, которая регенерирует энергию для инъекции генетического материала. Существуют также бактериофаги с коротким отростком, не имеющие отростка и нитевидные [11].
Фаги, как и все вирусы, являются абсолютными внутриклеточными паразитами. Хотя они переносят всю информацию для запуска собственной репродукции в соответствующем хозяине, у них отсутствуют механизмы для выработки энергии и рибосомы для синтеза белка. У некоторых фагов в геноме содержится несколько тысяч оснований, тогда как фаг G, самый крупный из секвенированных фагов, содержит 480 000 пар оснований — вдвое больше среднего значения для бактерий, хотя всё же недостаточного количества генов для важнейшего бактериального органоида как рибосомы [8].
4. Систематика бактериофагов
Большое количество выделенных и изученных бактериофагов определяет необходимость их систематизации. Классификация вирусов бактерий претерпевала изменения: основывалась на характеристике хозяина вируса, учитывались серологические, морфологические свойства, а затем строение и физико-химический состав вириона [12].
В настоящее время согласно Международной классификации и номенклатуре вирусов бактериофаги, в зависимости от типа нуклеиновой кислоты разделяют на ДНК- и РНК- содержащие.
По морфологическим характеристикам ДНК-содержащие фаги выделены в следующие семейства: Myoviridae, Siphoviridae, Podoviridae, Lipothrixviridae, Plasmaviridae, Corticoviridae, Fuselloviridae, Tectiviridae, Microviridae, Inoviridae Plectovirus и Inoviridae Inovirus.
РНК-содержащие: Cystoviridae, Leviviridae [13].
5. Взаимодействие бактериофага с бактериальными клетками

Адсорбция бактериофагов на поверхности бактериальной клетки
По характеру взаимодействия бактериофага с бактериальной клеткой различают вирулентные и умеренные фаги [11]. Вирулентные фаги могут только увеличиваться в количестве посредством литического цикла [8]. Процесс взаимодействия вирулентного бактериофага с клеткой складывается из нескольких стадий: адсорбции бактериофага на клетке, проникновения в клетку, биосинтеза компонентов фага и их сборки, выхода бактериофагов из клетки [7][14].
Первоначально бактериофаги прикрепляются к фагоспецифическим рецепторам на поверхности бактериальной клетки. Хвост фага с помощью ферментов, находящихся на его конце (в основном лизоцима), локально растворяет оболочку клетки, сокращается и содержащаяся в головке ДНК инъецируется в клетку, при этом белковая оболочка бактериофага остается снаружи. Инъецированная ДНК вызывает полную перестройку метаболизма клетки: прекращается синтез бактериальной ДНК, РНК и белков. ДНК бактериофага начинает транскрибироваться с помощью собственного фермента транскриптазы, который после попадания в бактериальную клетку активируется. Синтезируются сначала ранние, а затем поздние иРНК, которые поступают на рибосомы клетки-хозяина, где синтезируются ранние (ДНК-полимеразы, нуклеазы) и поздние (белки капсида и хвостового отростка, ферменты лизоцим, АТФаза и транскриптаза) белки бактериофага. Репликация ДНК бактериофага происходит по полуконсервативному механизму и осуществляется с участием собственных ДНК-полимераз. После синтеза поздних белков и завершения репликации ДНК наступает заключительный процесс — созревание фаговых частиц или соединение фаговой ДНК с белком оболочки и образование зрелых инфекционных фаговых частиц [15].
Продолжительность этого процесса может составлять от нескольких минут до нескольких часов [8]. Затем происходит лизис клетки, и освобождаются новые зрелые бактериофаги [11]. Иногда фаг инициирует лизирующий цикл, что приводит к лизису клетки и освобождению новых фагов. В качестве альтернативы фаг может инициировать лизогенный цикл, при котором он вместо репликации обратимо взаимодействует с генетической системой клетки-хозяина, интегрируясь в хромосому или сохраняясь в виде плазмиды [8]. Таким образом, вирусный геном реплицируется синхронно с ДНК хозяина и делением клетки, а подобное состояние фага называется профагом. Бактерия, содержащая профаг, становится лизогенной до тех пор, пока при определенных условиях или спонтанно профаг не будет стимулирован на осуществление лизирующего цикла репликации. Переход от лизогении к лизису называется лизогенной индукцией или индукцией профага. На индукцию фага оказывает сильное воздействие состояние клетки хозяина предшествующее индукции, также как наличие питательных веществ и другие условия, имеющие место быть в момент индукции. Скудные условия для роста способствуют лизогенному пути, тогда как хорошие условия способствуют лизирующей реакции [8, 11, 15].
Очень важным свойством бактериофагов является их специфичность: бактериофаги лизируют культуры определенного вида, более того, существуют так называемые типовые бактериофаги, лизирующие варианты внутри вида, хотя встречаются поливалентные бактериофаги, которые паразитируют в бактериях разных видов [16][17].
6. Жизненный цикл
Умеренные и вирулентные бактериофаги на начальных этапах взаимодействия с бактериальной клеткой имеют одинаковый цикл.
- Адсорбция бактериофага на фагоспецифических рецепторах клетки.
- Инъекция фаговой нуклеиновой кислоты в клетку хозяина.
- Совместная репликация фаговой и бактериальной нуклеиновой кислоты.
- Деление клетки.
- Далее бактериофаг может развиваться по двум моделям: лизогенный либо литический путь. Умеренные бактериофаги после деления клетки находятся в состоянии профага (Лизогенный путь). Вирулентные бактериофаги развиваются по Литической модели:
- Нуклеиновая кислота фага направляет синтез ферментов фага, используя для этого белоксинтезирующий аппарат бактерии. Фаг тем или иным способом инактивирует ДНК и РНК хозяина, а ферменты фага совсем расщепляют её; РНК фага «подчиняет» себе клеточный аппарат синтеза белка.
- Нуклеиновая кислота фага реплицируется, и направляет синтез новых белков оболочки. Образуются новые частицы фага в результате спонтанной самосборки белковой оболочки (капсид) вокруг фаговой нуклеиновой кислоты; под контролем РНК фага синтезируется лизоцим.
- Лизис клетки: клетка лопается под воздействием лизоцима; высвобождается около 200—1000 новых фагов; фаги инфицируют другие бактерии.
7. Применение
7.1. В медицине
Одной из областей использования бактериофагов является антибактериальная терапия, альтернативная приёму антибиотиков. Например, применяются бактериофаги: стрептококковый, стафилококковый, клебсиеллёзный, дизентерийный поливалентный, пиобактериофаг, коли, протейный и колипротейный и другие.
Бактериофаги применяются также в генной инженерии в качестве векторов, переносящих участки ДНК, возможна также естественная передача генов между бактериями посредством некоторых фагов (трансдукция).
Фаговые векторы обычно создают на базе умеренного бактериофага λ, содержащего двухцепочечную линейную молеклул ДНК. Левое и правое плечи фага имеют все гены, необходимые для литического цикла (репликации, размножения). Средняя часть генома бактериофага λ (содержит гены, контролирующие лизогению, то есть его интеграцию в ДНК бактериальной клетки) не существенна для его размножения и составляет примерно 25 тысяч пар нуклеотидов. Данная часть может быть заменена на чужеродный фрагмент ДНК. Такие модифицированные фаги проходят литический цикл, но лизогения не происходит. Векторы на основе бактериофага λ используют для клонирования фрагментов ДНК эукариот (то есть более крупных генов) размером до 23 т.п.н. Причем, фаги без вставок — менее 38 т.п.н или, напротив, со слишком большими вставками — более 52 т.п.н не развиваются и не поражают бактерии[18].
7.2. В биологии
Бактериофаги M13, фаг Т4, T7 и фаг λ используют для изучения белок-белковых, белок-пептидных и ДНК-белковых взаимодействий методом фагового дисплея.
Поскольку размножение бактериофага возможно только в живых клетках бактериофаги могут быть использованы для определения жизнеспособности бактерий. Данное направление имеет большие перспективы, поскольку, одним из основных вопросов при разных биотехнологических процессах является определение жизнеспособности используемых культур. С помощью метода электрооптического анализа клеточных суспензий была показана возможность изучения этапов взаимодействия фаг-микробная клетка[19].
wreferat.baza-referat.ru
Реферат- Бактериофаги
Работа добавлена на сайт samzan.ru: 2016-06-20 Бактериофаги
Бактериофагами называют вирусы, живущие в бактериях.
Бактериофаги (от бактерии и греч. phagos пожиратель; буквально пожиратели бактерий), фаги, бактериальные вирусы, вызывающие разрушение (лизис) бактерий и других микроорганизмов. Бактериофаги размножаются в клетках, лизируют их и переходят в др., как правило, молодые, растущие клетки. Впервые перевиваемый лизис бактерий (сибиреязвенной палочки) наблюдал в 1898 русский микробиолог Н. Ф. Гамалея. В 1915 английский учёный Ф. Туорт описал это же явление у гнойного стафилококка, а в 1917 французский учёный Ф. Д'Эрелль назвал литический агент, проходящий через бактериальные фильтры.
Строение и химический состав. Частицы многих бактериофагов состоят из головки округлой, гексагональной или палочковидной формы диаметром 45140 нм и отростка толщиной 1040 и длиной 100200 нм. Другие бактериофаги не имеют отростка; одни из них округлы, другие нитевидны, размером 8х800 нм. Содержимое головки состоит преимущественно из дезоксирибонуклеиновой кислоты (ДНК) (длина её нити во много раз превышает размер головки и достигает 6070 мкм, эта нить плотно скручена в головке) или рибонуклеиновой кислоты (РНК) и небольшого количества (около 3%) белка и некоторых других веществ. Отросток имеет вид полой трубки, окруженной чехлом, содержащим сократительные белки, подобные мышечным. У ряда бактериофагов чехол способен сокращаться, обнажая часть стержня. На конце отростка у многих бактериофагов имеется базальная пластинка с несколькими шиловидными или другие формы выступами. От пластинки отходят тонкие длинные нити, которые способствуют прикреплению фага к бактерии. Оболочки головки и отростка состоят из белков. Общее количество белка в частице фага 5060% , нуклеиновых кислот 4050% . Каждый бактериофаг обладает специфическими антигенными свойствами, отличными от антигенов бактерии-хозяина и других фагов. Имеются антигены, общие для ряда фагов (особенно содержащих РНК).
Распространение. Бактериофаги найдены для большинства бактерий, в том числе патогенных и сапрофитных, а также .для актиномицетов (актинофаги) и сине-зелёных водорослей. Встречаются бактериофаги в кишечнике человека и животных, в растениях, почве, водоёмах, сточных водах, навозе и т. д. Бактериофаги почвенных микроорганизмов влияют на течение микробиологических процессов в почве.
Размножение. Бактериофаг прикрепляется своим отростком к бактериальной клетке и, выделяя фермент, растворяет клеточную стенку; затем содержимое его головки через канадец отростка переходит внутрь клетки, где под влиянием нуклеиновой кислоты фага останавливается синтез бактериальных белков, ДНК и РНК и начинается синтез нуклеиновой кислоты, а затем и белков фага. Часть этих белков ферменты, другая часть образует оболочку зрелой частицы бактериофага Более мелкие, сферические фаги попадают в бактерии без участия отростка. Если клетка бактерии заражена одновременно частицами бактериофага, различающимися между собой по ряду свойств, то среди потомства, кроме частиц, подобных родителям, будут и такие, у которых эти свойства встречаются в новой комбинации, т. к. при размножении бактериофагов наблюдается
рекомбинация обмен кусками нитей нуклеиновой кислоты, являющейся носителем наследственной информации. Частицы крупных фагов выходят из бактерии, разрушая её, а некоторых мелких и нитевидных из живых бактерий. Одни бактериофаги весьма специфичны и способны лизировать клетки только одного какого-либо вида микроорганизмов (монофаги), другие клетки разных видов (полифаги).
Бактериофагов делят на вирулентные, вызывающие лизис клетки с образованием новых частиц, и умеренные (симбиотические), которые адсорбируются клеткой и проникают в неё, но лизиса не вызывают, а остаются в клетке в латентной (скрытой) неинфекционной форме (профаг). Культуры, содержащие латентный фаг, называются лизогенными. Лизогения передаётся потомству бактерии. Лизогенная культура может содержать 23 и более фагов; она, как правило, устойчива против находящихся в ней фагов (лишь небольшая часть клеток лизируется и освобождает зрелые фаги). Воздействуя на лизогенную культуру ультрафиолетовыми или рентгеновскими лучами, перекисью водорода и некоторыми другими веществами, можно значительно увеличить количество клеток, освобождающих фаг (т. н. индукция бактериофагов). Лизогения широко распространена среди всех видов бактерий и актиномицетов. В ряде случаев многие свойства лизогенной культуры (токсичность, подвижность бактерий и др.) зависят от наличия в ней определённых профагов. Описано много мутаций бактериофагов, сопровождающихся изменением их литической активности, строения частиц и «колоний», устойчивости против неблагоприятных воздействий и другие свойств. Бактериофаги играют большую роль в изменчивости и эволюции микробов, причём механизмы воздействия их на клетку разные. Бактериофаги могут резко изменять азотфиксирующую способность азотобактера, токсичность и антигенные свойства патогенных бактерий и др.
Практическое значение бактериофагов Некоторые фаги (одни или в сочетании с антибиотиками) применяли для профилактики (фагопрофилактики) и лечения (фаготерапии) ряда бактериальных инфекционных болезней человека (дизентерия, брюшной тиф, холера, чума, стафилококковые и анаэробная инфекции и др.) и животных. Однако антибиотики и другие химиотерапевтические средства оказались эффективнее фагов, в связи с чем применение их с лечебной целью сузилось. Бактериофаги успешно применяются при определении вида бактерий, актиномицетов. Бактериофаги могут вредить производству антибиотиков, аминокислот, молочных продуктов, бактериальных удобрений и в других отраслях микробиологического синтеза. Велико значение бактериофагов для теоретических работ по генетике и молекулярной биологии.
Список литературы
Для подготовки данной работы были использованы материалы с сайта http://www.school188spb.narod.ru
samzan.ru
Доклад - Бактериофаги, их строение, классификация, применение.
Бактериофаги– вирусы бактерий. История открытия бактериофага связана с именами Н.Ф. Гамалеи, наблюдавшего спонтанный лизис сибиреязвенных бактерий в 1898г. Английский бактериолог Ф. Туорт описал способность фильтрата стафилококков растворять свежую культуру этих же бактерий (1915г.). Французский ученый Ф.Д, Эррель подробно изучил взаимодействие фага и бактерий и сделал заключение, что открытый им литический агент является вирусом бактерий и назвал его «бактериофагом» — пожирателем бактерий.
Бактериофаги широко распространены: они выявлены у большинства бактерий, а также у других микроорганизмов, например, у грибов и поэтому их часто называют фагами. Наиболее детально изучена структура крупных фагов, к которым относятся фаги E. Coli (Т2, Т4, Т6). Они состоят из головки икосаэдрического типа, в которой заключена или ДНК, или РНК. Большинство фагов содержат двунитевую ДНК, замкнутую в кольцо. Хвостовой отросток имеет внутри полый цилиндрический стержень, сообщающийся с головкой, а снаружи – чехол, способный к сокращению наподобие мышцы. Чехол присоединен к воротничку, окружающему стержень около головки. Хвостовой отросток заканчивается шестиугольной базальной пластинкой с шипами от которых отходят нитевидные структуры – фибриллы.
По морфологии фаги подразделяются на 6 групп: 1) фаги с длинным отростком, чехол которого сокращается; 2) фаги с длинным отростком, чехол которого не сокращается; 3) фаги с короткими отростками; 4) фаги с аналогом отростка; 5)фаги без отростка; 6) нитевидные фаги.
Бактериофаги содержат группоспецифические и типоспецифические антигены, обладают иммуногенными свойствами, т.е. синтезируют специфические антитела в организме.
По специфичности взаимодействия различают следующие бактериофаги: 1) поливалентные – взаимодействуют с родственными видами бактерий; 2) моновалентные – взаимодействуют с бактериями определенного вида; 3) типовые – взаимодействуют с отдельными типами бактерий данного вида.
Взаимодействие фагов с бактериями может протекать: 1) по продуктивному типу – образуется фаговое потомство и бактерии лизируются; 2) по абортивному типу – фаговое потомство не образуется и бактерии сохраняют свою жизнедеятельность; 3) по интегративному типу – геном фага встраивается в хромосому бактерии и сосуществует с ней.
В зависимости от типа взаимодействия различают вирулентные и умеренные бактериофаги.
Вирулентные бактериофаги взаимодействуют с бактерией по продуктивному типу. Адсорбция фагов на бактериальной клетке происходит при наличии комплементарных рецепторов в ее клеточной стенке. На бактериях, лишенных клеточной стенки бактериофаги не адсорбируются. Фаги, имеющие хвостовой отросток, прикрепляются к бактериальной клетке свободным концом отростка (фибриллами, базальной пластинкой). В результате активации АТФ чехол хвостового отростка сокращается и стержень с помощью лизоцима, растворяющего прилегающий фрагмент клеточной стенки как бы просверливает оболочку клетки. При этом ДНК фага, содержащаяся в его головке, проходит в форме нити через канал хвостового стержня и инъецируется в клетку, а капсид фага остается снаружи бактерии. Инъецированная внутрь бактерии нуклеиновая кислота подавляет биосинтез компонентов клетки, заставляя ее синтезировать нуклеиновую кислоту и белки фага, затем происходит самосборка частиц фага. В результате изменения внутриклеточного осмотического давления и действия фагового лизоцима происходит разрушение оболочки, лизис бактерии и выход фагов из нее.
Умеренные бактериофаги в отличие от вирулентных взаимодействуют с чувствительными бактериями либо по продуктивному, либо по интегративному типам. При интегративном типе взаимодействия ДНК умеренного фага встраивается в хромосому бактерии, реплицируется синхронно с геном размножающейся бактерии, не вызывая ее лизиса. ДНК бактериофага, встроенная в хромосому бактерии, называется профагом, а культура бактерий – лизогенной. Такое сосуществование бактерии и умеренного бактериофага называется лизогенией.
Бактериофаги используют: 1) в лабораторной диагностике инфекций при внутривидовой идентификации бактерий, т.е. определения фаговара. Для этого применяют метод фаготипирования. На чашку с плотной питательной средой засевают «газоном» чистой культурой возбудителя и наносят капли различных диагностических типоспецифических фагов. Фаговар бактерии определяется тем типом фага, который вызвал ее лизис. Методику фаготипирования используют для выявления источника и путей распространения инфекции (эпидемиологическое маркирование). Например, при возникновении массовых заболеваний стафилококковой этиологии в родильных домах, детских и больничных учреждениях большое значение приобретает выявление источников инфекции и установление эпидемиологических связей. Решение этой задачи возможно только методом фаготипирования стафилококков, подтверждающим идентичность микроорганизмов, выделяемых у больных, носителей и объектов внешней среды.
Таким образом, при проведении эпидемиологического обследования метод фаготипирования бактерий дает возможность: 1) устанавливать или исключать предполагаемые источники инфекции; 2) прослеживать эпидемические связи; 3) отличать местные случаи от «привозных» и спородические заболевания от эпидемических.
2) фаги применяют также для лечения и профилактики ряда бактериальных инфекций. Производят брюшнотифозный, сальмонеллезный, дизентерийный, синегнойный, стафилококковый, стрептококковый фаги; комбинированные: колипротейный, пиобактериофаги. Бактериофаги назначают по показаниям перорально, парентерально или местно в виде жидких, таблетированных форм, свечей, аэрозолей.
3) бактериофаги широко применяют в генной инженерии в качестве векторов для получения рекомбинантных ДНК.
Тесты по теме:
1. Требования, предъявляемые к питательным средам:
а) плотные
б) рН 5,0-5,2
в) рН 7,2-7,6
г) стерильные
д) концентрация NaCl 0,5%
2. Концентрация агара в полужидких питательных средах:
а) 0,1%
б) 0,5%
в) 1,0%
г) 1,5-2,0%
3. К простым питательным средам относятся:
а) кровяной агар
б) пептонная вода
в) сахарный бульон
г) мясопептонный агар
4. К сложным питательным средам относятся:
а) кровяной агар
б) пептонная вода
в) сахарный бульон
г) мясопептонный агар
5. Среды, на которых лучше растет какой-то определенный микроорганизм:
а) дифференциально-диагностические
б) элективные
в) полужидкие
г) среды обогащения
6. Среды, стимулирующие рост определенного микроорганизма, но при этом ингибируют рост других:
а) дифференциально-диагностические
б) элективные
в) полужидкие
г) среды обогащения
7. Среды, служащие для изучения ферментативной активности бактерий:
а) дифференциально-диагностические
б) элективные
в) полужидкие
г) среды обогащения
8. Питательная среда, используемая при культивировании бактерий кишечной группы:
а) среда Чистович
б) среда Эндо
в) среда Сабуро
г) среда Вильсон-Блер
9. Питательная среда, используемая при культивировании стафилококков:
а) среда Чистович
б) среда Эндо
в) среда Сабуро
г) среда Вильсон-Блер
10. Питательная среда, используемая при культивировании грибов:
а) среда Чистович
б) среда Эндо
в) среда Сабуро
г) среда Вильсон-Блер
11. При культивировании анаэробов используют:
а) среда Чистович
б) среда Эндо
в) среда Сабуро
г) среда Вильсон-Блер
12. При культивировании вирусов используют:
а) куриные эмбрионы
б) простые питательные среды
в) сложные питательные среды
г) культуры клеток
13. Перечислите фазы размножения бактерий:
а) первичная
б) стационарная
в) роста
г) логарифмическая
14. Скопление микроорганизмов одного вида:
а) колония
б) чистая культура
в) бактериофаг
г) пигмент
15. Рост бактерий на жидких питательных средах:
а) в виде пленки
б) колоний
в) сплошного налета
г) осадка
д) равномерного помутнения среды
е) по «уколу»
16. К каким свойствам относится характер роста выделенной культуры на питательных средах:
а) морфологические
б) культуральные
в) сахаролитические
г) гемолитические
д) протеолитические
17. Способность разрушать эритроциты на средах с кровью относится к свойствам:
а) морфологические
б) культуральные
в) сахаролитические
г) гемолитические
д) протеолитические
18. Способность расщеплять белки на средах, содержащих желатин, молоко, сыворотку относится к свойствам:
а) морфологические
б) культуральные
в) сахаролитические
г) гемолитические
д) протеолитические
19. Бактерии, использующие в питании неорганический углерод:
а) гетеротрофы
б) автотрофы
в) фототрофы
г) хемотрофы
20. Бактерии, использующие в питании органический углерод:
а) гетеротрофы
б) автотрофы
в) фототрофы
г) хемотрофы
21. Бактерии, получающие энергию за счет окислительно-восстановительных реакций:
а) гетеротрофы
б) автотрофы
в) фототрофы
г) хемотрофы
22. Бактерии, получающие энергию за счет света:
а) гетеротрофы
б) автотрофы
в) фототрофы
г) хемотрофы
23. Бактерии, которые питаются мертвым органическим материалом:
а) сапрофиты
б) паразиты
в) гетеротрофы
г) факультативные паразиты
24. Бактерии, которые питаются за счет макроорганизма:
а) сапрофиты
б) паразиты
в) гетеротрофы
г) факультативные паразиты
25. Микроорганизмы, которые растут и размножаются в присутствии кислорода:
а) облигатные аэробы
б) микроаэрофилы
в) строгие анаэробы
г) аэротолерантные м/о
д) факультативные анаэробы
26. Микроорганизмы, которые способны расти и размножаться как в присутствии кислорода, так и в его отсутствии:
а) облигатные аэробы
б) микроаэрофилы
в) строгие анаэробы
г) аэротолерантные м/о
д) факультативные анаэробы
27. Микроорганизмы, для которых кислород является токсичным:
а) облигатные аэробы
б) микроаэрофилы
в) строгие анаэробы
г) аэротолерантные м/о
д) факультативные анаэробы
28. Вирусы, проникающие в бактерии, паразитирующие в них вплоть до гибели:
а) ДНК-вирусы
б) РНК-вирусы
в) бактериофаги
г) аденовирусы
29. Бактериофаги состоят:
а) капсид
б) головка
в) хвостовой отросток
г) базальная пластина
30. По специфичности взаимодействия бактериофаги различают:
а) поливалентные
б) продуктивные
в) абортивные
г) моновалентные
31. Взаимодействие фагов с бактериями при котором образуется фаговое потомство и лизис бактерий протекает по типу:
а) абортивному
б) интегративному
в) продуктивному
32. Взаимодействие фагов с бактериями при котором геном фага встраивается в хромосому бактерии и сосуществует в ней протекает по типу:
а) абортивному
б) интегративному
в) продуктивному
33. Фаги, взаимодействие которых с бактерией происходит либо по продуктивному, либо по интегративному типу называют:
а) вирулентные
б) поливалентные
в) умереннные
34. Практическое применение фагов:
а) для культивирования вирусов
б) для лечения и профилактики инфекционных заболеваний
в) в генной инженерии
www.ronl.ru
Реферат Бактериофаги
скачатьРеферат на тему:
План:
Введение- 1 История
- 2 Роль бактериофагов в биосфере
- 3 Строение бактериофагов
- 4 Систематика бактериофагов
- 5 Взаимодействие бактериофага с бактериальными клетками
- 6 Жизненный цикл
- 7 Применение
- 7.1 В медицине
- 7.2 В биологии
Введение
1 — головка, 2 — хвост, 3 — нуклеиновая кислота, 4 — капсид, 5 — «воротничок», 6 — белковый чехол хвоста, 7 — фибрилла хвоста, 8 — шипы, 9 — базальная пластинка
Бактериофа́ги (фаги) (от др.-греч. φᾰγω — «пожираю») — вирусы, избирательно поражающие бактериальные клетки. Чаще всего бактериофаги размножаются внутри бактерий и вызывают их лизис. Как правило, бактериофаг состоит из белковой оболочки и генетического материала одноцепочечной или двуцепочечной нуклеиновой кислоты (ДНК или, реже, РНК). Размер частиц приблизительно от 20 до 200 нм.
1. История
Английский бактериолог Туорт, Фредерик в статье 1915 года описал инфекционную болезнь стафилококков, инфицирующий агент проходил через фильтры, и его можно было переносить от одной колонии к другой.
Независимо от Фредерика Туорта французско-канадский микробиолог Д’Эрель, Феликс 3 сентября 1917 год сообщил об открытии бактериофагов. Наряду с этим известно, что российский микробиолог Гамалея, Николай Фёдорович ещё в 1898 году, впервые наблюдал явление лизиса бактерий (сибиреязвенной палочки) под влиянием перевиваемого агента [1][2].
После открытия явлений бактериофагии Д’Эрелль развил учение о том, что бактериофаги патогенных бактерий, являясь их паразитами, играют большую роль в патогенезе инфекций, обеспечивая выздоровление больного организма, а затем создания специфического иммунитета. Это положение привлекло к явлению бактериофагии внимание многих исследователей, которые предполагали найти в фагах важное средство борьбы с наиболее опасными инфекционными болезнями человека и животных.
Также Феликс Д’Эрель выдвинул предположение, что бактериофаги имеют корпускулярную природу. Однако только после изобретения электронного микроскопа удалось увидеть и изучить ультраструктуру фагов. Долгое время представления о морфологии и основных особенностях фагов основывались на результатах изучения фагов Т-группы — Т1, Т2,…, Т7, которые размножаются на Е. coli штамма B. Однако с каждым годом появлялись новые данные, касающиеся морфологии и структуры разнообразных фагов, что обусловило необходимость их морфологической классификации.
2. Роль бактериофагов в биосфере
Бактериофаги представляют собой наиболее многочисленную, широко распространенную в биосфере и, предположительно, наиболее эволюционно древнюю группу вирусов [3][4]. Приблизительный размер популяции фагов составляет более 1030 фаговых частиц[5].
В природных условиях фаги встречаются в тех местах, где есть чувствительные к ним бактерии. Чем богаче тот или иной субстрат (почва, выделения человека и животных, вода и т. д.) микроорганизмами, тем в большем количестве в нём встречаются соответствующие фаги. Так, фаги, лизирующие клетки всех видов почвенных микроорганизмов, находятся в почвах. Особенно богаты фагами черноземы и почвы, в которые вносились органические удобрения.
Бактериофаги выполняют важную роль в контроле численности микробных популяций, в автолизе стареющих клеток, в переносе бактериальных генов, выступая в качестве векторных «систем»[6].
Действительно, бактериофаги представляют собой один из основных подвижных генетических элементов. Посредством трансдукции они привносят в бактериальный геном новые гены. Было подсчитано, что за 1 секунду могут быть инфицированы 1024 бактерий [7]. Это означает, что постоянный перенос генетического материала распределяется между бактериями, обитающими в сходных условиях.
Высокий уровень специализации, долгосрочное существование, способность быстро репродуцироваться в соответствующем хозяине способствует их сохранению в динамичном балансе среди широкого разнообразия видов бактерий в любой природной экосистеме. Когда подходящий хозяин отсутствует, многие фаги могут сохранять способность к инфицированию на протяжении десятилетий, если не будут уничтожены экстремальными веществами либо условиями внешней среды [8].
3. Строение бактериофагов
Бактериофаги различаются по химической структуре, типу нуклеиновой кислоты, морфологии и характеру взаимодействия с бактериями. По размеру бактериальные вирусы в сотни и тысячи раз меньше микробных клеток.
Типичная фаговая частица (вирион) состоит из головки и хвоста. Длина хвоста обычно в 2 — 4 раза больше диаметра головки. В головке содержится генетический материал — одноцепочечная или двуцепочечная РНК или ДНК с ферментом транскриптазой в неактивном состоянии, окруженная белковой или липопротеиновой оболочкой — капсидом, сохраняющим геном вне клетки [8][9].
Нуклеиновая кислота и капсид вместе составляют нуклеокапсид. Бактериофаги могут иметь икосаэдральный капсид, собранный из множества копий одного или двух специфичных белков. Обычно углы состоят из пентамеров белка, а опора каждой стороны из гексамеров того же или сходного белка. Более того, фаги по форме могут быть сферические, лимоновидные или плеоморфные [10]. Хвост представляет собой белковую трубку — продолжение белковой оболочки головки, в основании хвоста имеется АТФаза, которая регенерирует энергию для инъекции генетического материала. Существуют также бактериофаги с коротким отростком, не имеющие отростка и нитевидные [11].
Фаги, как и все вирусы, являются абсолютными внутриклеточными паразитами. Хотя они переносят всю информацию для запуска собственной репродукции в соответствующем хозяине, у них отсутствуют механизмы для выработки энергии и рибосомы для синтеза белка. У некоторых фагов в геноме содержится несколько тысяч оснований, тогда как фаг G, самый крупный из секвенированных фагов, содержит 480 000 пар оснований — вдвое больше среднего значения для бактерий, хотя всё же недостаточного количества генов для важнейшего бактериального органоида как рибосомы [8].
4. Систематика бактериофагов
Большое количество выделенных и изученных бактериофагов определяет необходимость их систематизации. Классификация вирусов бактерий претерпевала изменения: основывалась на характеристике хозяина вируса, учитывались серологические, морфологические свойства, а затем строение и физико-химический состав вириона [12].
В настоящее время согласно Международной классификации и номенклатуре вирусов бактериофаги, в зависимости от типа нуклеиновой кислоты разделяют на ДНК- и РНК- содержащие.
По морфологическим характеристикам ДНК-содержащие фаги выделены в следующие семейства: Myoviridae, Siphoviridae, Podoviridae, Lipothrixviridae, Plasmaviridae, Corticoviridae, Fuselloviridae, Tectiviridae, Microviridae, Inoviridae Plectovirus и Inoviridae Inovirus.
РНК-содержащие: Cystoviridae, Leviviridae [13].
5. Взаимодействие бактериофага с бактериальными клетками
Адсорбция бактериофагов на поверхности бактериальной клетки
По характеру взаимодействия бактериофага с бактериальной клеткой различают вирулентные и умеренные фаги [11]. Вирулентные фаги могут только увеличиваться в количестве посредством литического цикла [8]. Процесс взаимодействия вирулентного бактериофага с клеткой складывается из нескольких стадий: адсорбции бактериофага на клетке, проникновения в клетку, биосинтеза компонентов фага и их сборки, выхода бактериофагов из клетки [7][14].
Первоначально бактериофаги прикрепляются к фагоспецифическим рецепторам на поверхности бактериальной клетки. Хвост фага с помощью ферментов, находящихся на его конце (в основном лизоцима), локально растворяет оболочку клетки, сокращается и содержащаяся в головке ДНК инъецируется в клетку, при этом белковая оболочка бактериофага остается снаружи. Инъецированная ДНК вызывает полную перестройку метаболизма клетки: прекращается синтез бактериальной ДНК, РНК и белков. ДНК бактериофага начинает транскрибироваться с помощью собственного фермента транскриптазы, который после попадания в бактериальную клетку активируется. Синтезируются сначала ранние, а затем поздние иРНК, которые поступают на рибосомы клетки-хозяина, где синтезируются ранние (ДНК-полимеразы, нуклеазы) и поздние (белки капсида и хвостового отростка, ферменты лизоцим, АТФаза и транскриптаза) белки бактериофага. Репликация ДНК бактериофага происходит по полуконсервативному механизму и осуществляется с участием собственных ДНК-полимераз. После синтеза поздних белков и завершения репликации ДНК наступает заключительный процесс — созревание фаговых частиц или соединение фаговой ДНК с белком оболочки и образование зрелых инфекционных фаговых частиц [15].
Продолжительность этого процесса может составлять от нескольких минут до нескольких часов [8]. Затем происходит лизис клетки, и освобождаются новые зрелые бактериофаги [11]. Иногда фаг инициирует лизирующий цикл, что приводит к лизису клетки и освобождению новых фагов. В качестве альтернативы фаг может инициировать лизогенный цикл, при котором он вместо репликации обратимо взаимодействует с генетической системой клетки-хозяина, интегрируясь в хромосому или сохраняясь в виде плазмиды [8]. Таким образом, вирусный геном реплицируется синхронно с ДНК хозяина и делением клетки, а подобное состояние фага называется профагом. Бактерия, содержащая профаг, становится лизогенной до тех пор, пока при определенных условиях или спонтанно профаг не будет стимулирован на осуществление лизирующего цикла репликации. Переход от лизогении к лизису называется лизогенной индукцией или индукцией профага. На индукцию фага оказывает сильное воздействие состояние клетки хозяина предшествующее индукции, также как наличие питательных веществ и другие условия, имеющие место быть в момент индукции. Скудные условия для роста способствуют лизогенному пути, тогда как хорошие условия способствуют лизирующей реакции [8, 11, 15].
Очень важным свойством бактериофагов является их специфичность: бактериофаги лизируют культуры определенного вида, более того, существуют так называемые типовые бактериофаги, лизирующие варианты внутри вида, хотя встречаются поливалентные бактериофаги, которые паразитируют в бактериях разных видов [16][17].
6. Жизненный цикл
Умеренные и вирулентные бактериофаги на начальных этапах взаимодействия с бактериальной клеткой имеют одинаковый цикл.
- Адсорбция бактериофага на фагоспецифических рецепторах клетки.
- Инъекция фаговой нуклеиновой кислоты в клетку хозяина.
- Совместная репликация фаговой и бактериальной нуклеиновой кислоты.
- Деление клетки.
- Далее бактериофаг может развиваться по двум моделям: лизогенный либо литический путь. Умеренные бактериофаги после деления клетки находятся в состоянии профага (Лизогенный путь). Вирулентные бактериофаги развиваются по Литической модели:
- Нуклеиновая кислота фага направляет синтез ферментов фага, используя для этого белоксинтезирующий аппарат бактерии. Фаг тем или иным способом инактивирует ДНК и РНК хозяина, а ферменты фага совсем расщепляют её; РНК фага «подчиняет» себе клеточный аппарат синтеза белка.
- Нуклеиновая кислота фага реплицируется, и направляет синтез новых белков оболочки. Образуются новые частицы фага в результате спонтанной самосборки белковой оболочки (капсид) вокруг фаговой нуклеиновой кислоты; под контролем РНК фага синтезируется лизоцим.
- Лизис клетки: клетка лопается под воздействием лизоцима; высвобождается около 200—1000 новых фагов; фаги инфицируют другие бактерии.
7. Применение
7.1. В медицине
Одной из областей использования бактериофагов является антибактериальная терапия, альтернативная приёму антибиотиков. Например, применяются бактериофаги: стрептококковый, стафилококковый, клебсиеллёзный, дизентерийный поливалентный, пиобактериофаг, коли, протейный и колипротейный и другие.
Бактериофаги применяются также в генной инженерии в качестве векторов, переносящих участки ДНК, возможна также естественная передача генов между бактериями посредством некоторых фагов (трансдукция).
Фаговые векторы обычно создают на базе умеренного бактериофага λ, содержащего двухцепочечную линейную молеклул ДНК. Левое и правое плечи фага имеют все гены, необходимые для литического цикла (репликации, размножения). Средняя часть генома бактериофага λ (содержит гены, контролирующие лизогению, то есть его интеграцию в ДНК бактериальной клетки) не существенна для его размножения и составляет примерно 25 тысяч пар нуклеотидов. Данная часть может быть заменена на чужеродный фрагмент ДНК. Такие модифицированные фаги проходят литический цикл, но лизогения не происходит. Векторы на основе бактериофага λ используют для клонирования фрагментов ДНК эукариот (то есть более крупных генов) размером до 23 т.п.н. Причем, фаги без вставок — менее 38 т.п.н или, напротив, со слишком большими вставками — более 52 т.п.н не развиваются и не поражают бактерии[18].
7.2. В биологии
Бактериофаги M13, фаг Т4, T7 и фаг λ используют для изучения белок-белковых, белок-пептидных и ДНК-белковых взаимодействий методом фагового дисплея.
Поскольку размножение бактериофага возможно только в живых клетках бактериофаги могут быть использованы для определения жизнеспособности бактерий. Данное направление имеет большие перспективы, поскольку, одним из основных вопросов при разных биотехнологических процессах является определение жизнеспособности используемых культур. С помощью метода электрооптического анализа клеточных суспензий была показана возможность изучения этапов взаимодействия фаг-микробная клетка[19].
wreferat.baza-referat.ru
Реферат - Бактериофаги, их строение, классификация, применение.
Бактериофаги– вирусы бактерий. История открытия бактериофага связана с именами Н.Ф. Гамалеи, наблюдавшего спонтанный лизис сибиреязвенных бактерий в 1898г. Английский бактериолог Ф. Туорт описал способность фильтрата стафилококков растворять свежую культуру этих же бактерий (1915г.). Французский ученый Ф.Д, Эррель подробно изучил взаимодействие фага и бактерий и сделал заключение, что открытый им литический агент является вирусом бактерий и назвал его «бактериофагом» — пожирателем бактерий.
Бактериофаги широко распространены: они выявлены у большинства бактерий, а также у других микроорганизмов, например, у грибов и поэтому их часто называют фагами. Наиболее детально изучена структура крупных фагов, к которым относятся фаги E. Coli (Т2, Т4, Т6). Они состоят из головки икосаэдрического типа, в которой заключена или ДНК, или РНК. Большинство фагов содержат двунитевую ДНК, замкнутую в кольцо. Хвостовой отросток имеет внутри полый цилиндрический стержень, сообщающийся с головкой, а снаружи – чехол, способный к сокращению наподобие мышцы. Чехол присоединен к воротничку, окружающему стержень около головки. Хвостовой отросток заканчивается шестиугольной базальной пластинкой с шипами от которых отходят нитевидные структуры – фибриллы.
По морфологии фаги подразделяются на 6 групп: 1) фаги с длинным отростком, чехол которого сокращается; 2) фаги с длинным отростком, чехол которого не сокращается; 3) фаги с короткими отростками; 4) фаги с аналогом отростка; 5)фаги без отростка; 6) нитевидные фаги.
Бактериофаги содержат группоспецифические и типоспецифические антигены, обладают иммуногенными свойствами, т.е. синтезируют специфические антитела в организме.
По специфичности взаимодействия различают следующие бактериофаги: 1) поливалентные – взаимодействуют с родственными видами бактерий; 2) моновалентные – взаимодействуют с бактериями определенного вида; 3) типовые – взаимодействуют с отдельными типами бактерий данного вида.
Взаимодействие фагов с бактериями может протекать: 1) по продуктивному типу – образуется фаговое потомство и бактерии лизируются; 2) по абортивному типу – фаговое потомство не образуется и бактерии сохраняют свою жизнедеятельность; 3) по интегративному типу – геном фага встраивается в хромосому бактерии и сосуществует с ней.
В зависимости от типа взаимодействия различают вирулентные и умеренные бактериофаги.
Вирулентные бактериофаги взаимодействуют с бактерией по продуктивному типу. Адсорбция фагов на бактериальной клетке происходит при наличии комплементарных рецепторов в ее клеточной стенке. На бактериях, лишенных клеточной стенки бактериофаги не адсорбируются. Фаги, имеющие хвостовой отросток, прикрепляются к бактериальной клетке свободным концом отростка (фибриллами, базальной пластинкой). В результате активации АТФ чехол хвостового отростка сокращается и стержень с помощью лизоцима, растворяющего прилегающий фрагмент клеточной стенки как бы просверливает оболочку клетки. При этом ДНК фага, содержащаяся в его головке, проходит в форме нити через канал хвостового стержня и инъецируется в клетку, а капсид фага остается снаружи бактерии. Инъецированная внутрь бактерии нуклеиновая кислота подавляет биосинтез компонентов клетки, заставляя ее синтезировать нуклеиновую кислоту и белки фага, затем происходит самосборка частиц фага. В результате изменения внутриклеточного осмотического давления и действия фагового лизоцима происходит разрушение оболочки, лизис бактерии и выход фагов из нее.
Умеренные бактериофаги в отличие от вирулентных взаимодействуют с чувствительными бактериями либо по продуктивному, либо по интегративному типам. При интегративном типе взаимодействия ДНК умеренного фага встраивается в хромосому бактерии, реплицируется синхронно с геном размножающейся бактерии, не вызывая ее лизиса. ДНК бактериофага, встроенная в хромосому бактерии, называется профагом, а культура бактерий – лизогенной. Такое сосуществование бактерии и умеренного бактериофага называется лизогенией.
Бактериофаги используют: 1) в лабораторной диагностике инфекций при внутривидовой идентификации бактерий, т.е. определения фаговара. Для этого применяют метод фаготипирования. На чашку с плотной питательной средой засевают «газоном» чистой культурой возбудителя и наносят капли различных диагностических типоспецифических фагов. Фаговар бактерии определяется тем типом фага, который вызвал ее лизис. Методику фаготипирования используют для выявления источника и путей распространения инфекции (эпидемиологическое маркирование). Например, при возникновении массовых заболеваний стафилококковой этиологии в родильных домах, детских и больничных учреждениях большое значение приобретает выявление источников инфекции и установление эпидемиологических связей. Решение этой задачи возможно только методом фаготипирования стафилококков, подтверждающим идентичность микроорганизмов, выделяемых у больных, носителей и объектов внешней среды.
Таким образом, при проведении эпидемиологического обследования метод фаготипирования бактерий дает возможность: 1) устанавливать или исключать предполагаемые источники инфекции; 2) прослеживать эпидемические связи; 3) отличать местные случаи от «привозных» и спородические заболевания от эпидемических.
2) фаги применяют также для лечения и профилактики ряда бактериальных инфекций. Производят брюшнотифозный, сальмонеллезный, дизентерийный, синегнойный, стафилококковый, стрептококковый фаги; комбинированные: колипротейный, пиобактериофаги. Бактериофаги назначают по показаниям перорально, парентерально или местно в виде жидких, таблетированных форм, свечей, аэрозолей.
3) бактериофаги широко применяют в генной инженерии в качестве векторов для получения рекомбинантных ДНК.
Тесты по теме:
1. Требования, предъявляемые к питательным средам:
а) плотные
б) рН 5,0-5,2
в) рН 7,2-7,6
г) стерильные
д) концентрация NaCl 0,5%
2. Концентрация агара в полужидких питательных средах:
а) 0,1%
б) 0,5%
в) 1,0%
г) 1,5-2,0%
3. К простым питательным средам относятся:
а) кровяной агар
б) пептонная вода
в) сахарный бульон
г) мясопептонный агар
4. К сложным питательным средам относятся:
а) кровяной агар
б) пептонная вода
в) сахарный бульон
г) мясопептонный агар
5. Среды, на которых лучше растет какой-то определенный микроорганизм:
а) дифференциально-диагностические
б) элективные
в) полужидкие
г) среды обогащения
6. Среды, стимулирующие рост определенного микроорганизма, но при этом ингибируют рост других:
а) дифференциально-диагностические
б) элективные
в) полужидкие
г) среды обогащения
7. Среды, служащие для изучения ферментативной активности бактерий:
а) дифференциально-диагностические
б) элективные
в) полужидкие
г) среды обогащения
8. Питательная среда, используемая при культивировании бактерий кишечной группы:
а) среда Чистович
б) среда Эндо
в) среда Сабуро
г) среда Вильсон-Блер
9. Питательная среда, используемая при культивировании стафилококков:
а) среда Чистович
б) среда Эндо
в) среда Сабуро
г) среда Вильсон-Блер
10. Питательная среда, используемая при культивировании грибов:
а) среда Чистович
б) среда Эндо
в) среда Сабуро
г) среда Вильсон-Блер
11. При культивировании анаэробов используют:
а) среда Чистович
б) среда Эндо
в) среда Сабуро
г) среда Вильсон-Блер
12. При культивировании вирусов используют:
а) куриные эмбрионы
б) простые питательные среды
в) сложные питательные среды
г) культуры клеток
13. Перечислите фазы размножения бактерий:
а) первичная
б) стационарная
в) роста
г) логарифмическая
14. Скопление микроорганизмов одного вида:
а) колония
б) чистая культура
в) бактериофаг
г) пигмент
15. Рост бактерий на жидких питательных средах:
а) в виде пленки
б) колоний
в) сплошного налета
г) осадка
д) равномерного помутнения среды
е) по «уколу»
16. К каким свойствам относится характер роста выделенной культуры на питательных средах:
а) морфологические
б) культуральные
в) сахаролитические
г) гемолитические
д) протеолитические
17. Способность разрушать эритроциты на средах с кровью относится к свойствам:
а) морфологические
б) культуральные
в) сахаролитические
г) гемолитические
д) протеолитические
18. Способность расщеплять белки на средах, содержащих желатин, молоко, сыворотку относится к свойствам:
а) морфологические
б) культуральные
в) сахаролитические
г) гемолитические
д) протеолитические
19. Бактерии, использующие в питании неорганический углерод:
а) гетеротрофы
б) автотрофы
в) фототрофы
г) хемотрофы
20. Бактерии, использующие в питании органический углерод:
а) гетеротрофы
б) автотрофы
в) фототрофы
г) хемотрофы
21. Бактерии, получающие энергию за счет окислительно-восстановительных реакций:
а) гетеротрофы
б) автотрофы
в) фототрофы
г) хемотрофы
22. Бактерии, получающие энергию за счет света:
а) гетеротрофы
б) автотрофы
в) фототрофы
г) хемотрофы
23. Бактерии, которые питаются мертвым органическим материалом:
а) сапрофиты
б) паразиты
в) гетеротрофы
г) факультативные паразиты
24. Бактерии, которые питаются за счет макроорганизма:
а) сапрофиты
б) паразиты
в) гетеротрофы
г) факультативные паразиты
25. Микроорганизмы, которые растут и размножаются в присутствии кислорода:
а) облигатные аэробы
б) микроаэрофилы
в) строгие анаэробы
г) аэротолерантные м/о
д) факультативные анаэробы
26. Микроорганизмы, которые способны расти и размножаться как в присутствии кислорода, так и в его отсутствии:
а) облигатные аэробы
б) микроаэрофилы
в) строгие анаэробы
г) аэротолерантные м/о
д) факультативные анаэробы
27. Микроорганизмы, для которых кислород является токсичным:
а) облигатные аэробы
б) микроаэрофилы
в) строгие анаэробы
г) аэротолерантные м/о
д) факультативные анаэробы
28. Вирусы, проникающие в бактерии, паразитирующие в них вплоть до гибели:
а) ДНК-вирусы
б) РНК-вирусы
в) бактериофаги
г) аденовирусы
29. Бактериофаги состоят:
а) капсид
б) головка
в) хвостовой отросток
г) базальная пластина
30. По специфичности взаимодействия бактериофаги различают:
а) поливалентные
б) продуктивные
в) абортивные
г) моновалентные
31. Взаимодействие фагов с бактериями при котором образуется фаговое потомство и лизис бактерий протекает по типу:
а) абортивному
б) интегративному
в) продуктивному
32. Взаимодействие фагов с бактериями при котором геном фага встраивается в хромосому бактерии и сосуществует в ней протекает по типу:
а) абортивному
б) интегративному
в) продуктивному
33. Фаги, взаимодействие которых с бактерией происходит либо по продуктивному, либо по интегративному типу называют:
а) вирулентные
б) поливалентные
в) умереннные
34. Практическое применение фагов:
а) для культивирования вирусов
б) для лечения и профилактики инфекционных заболеваний
в) в генной инженерии
www.ronl.ru