
|
|
|
|
 Far Far |
| WinNavigator |
| Frigate |
| Norton
Commander |
| WinNC |
| Dos
Navigator |
| Servant
Salamander |
| Turbo
Browser |
|
|
| Winamp,
Skins, Plugins |
| Необходимые
Утилиты |
| Текстовые
редакторы |
| Юмор |
|
|

|
File managers and best utilites |
Читать реферат по химии: "Нуклеиновые кислоты". Нуклеиновые кислоты химия реферат
Читать реферат по химии: "Нуклеиновые кислоты"
 (Назад)
(Назад) (Cкачать работу)
(Cкачать работу)
Функция "чтения" служит для ознакомления с работой. Разметка, таблицы и картинки документа могут отображаться неверно или не в полном объёме!
Содержание
История открытия и названия нуклеиновых кислотНахождение нуклеиновых кислот в природеПолучение нуклеиновых кислот Химические свойства нуклеиновых кислотПрименение нуклеиновых кислотЗанимательные факты о нуклеиновых кислотах
1. История открытия нуклеинов и их названия Открытие нуклеиновых кислот связано с именем молодого врача из города Базеля (Швейцария) Фридриха Мишера. После окончания медицинского факультета Мишер был послан для усовершенствования и работы над диссертацией в Тюбинген (Германия) в физиолого-химическую лабораторию, возглавляемую Ф. Гоппе-Зейлером. Тюбингенская лаборатория в то время была известна ученому миру. Пройдя практику по органической химии, Мишер приступил к работе в биохимической лаборатории. Ему было поручено заняться изучением химического состава гноя. Молодой ученый не возражал против предложенной темы, так как считал лейкоциты, присутствующие в гное, одними из самых простых клеток.
Путём многочисленных опытов он получил из гнойных клеток вещество ядерного происхождения. Мишер был уверен именно в ядерном его источнике. Поэтому он начал более тщательное выделение ядер. В то время еще никто в биохимических лабораториях не пытался выделить ядра или какие-либо другие субклеточные компоненты, так что и здесь он был пионером.
Продолжив дальше очищать ядро от других клеточных фрагментов, он получил странное вещетво. Оно не разлагалось протеолитическими ферментами, значит, не являлось белком. Отсутствие растворимости в горячем спирте указывало на то, что это вещество не являлось и фосфолипидом. По-видимому, оно относилось к новому классу биохимических соединений.
Но Мишер с большой горячностью настаивал на точности своих результатов и добивался разрешения опубликовать их в печати. Тогда Гоппе-Зейлер решил проверить данные Мишера лично. Он и два его ассистента (одним из них был русский химик Любавин) в течение года шаг за шагом прошли все этапы аналитической работы Мишера и полностью подтвердили его данные, выделив нуклеин из клеток крови и из дрожжей. В 1871 г. работа Мишера вместе с подтверждающими ее контрольными работами Гоппе-Зейлера и его ассистентов увидела свет. Существование нуклеина как специфического ядерного вещества стало научным фактом. Вскоре методика Мишера была применена для выделения нуклеина из различных тканей.
Термин «нуклеиновые кислоты» был предложен в 1889: нуклеиновыми они были названы потому, что впервые были открыты в ядрах клеток, а кислотами — из-за наличия в их составе остатков фосфорной кислоты. Позже было показано, что нуклеиновые кислоты построены из большого числа нуклеотидов (от нескольких десятков до сотен миллионов). В состав каждого нуклеотида входит азотистое основание, углевод (пентоза) и фосфорная кислота. 2. Нахождение нуклеиновых кислот в природе Нуклеиновые кислоты в природе встречаются во всех живых клетках. Живые клетки, за исключением сперматозоидов, в норме содержат значительно больше рибонуклеиновой, чем дезоксирибонуклеиновой кислоты. На методы выделения дезоксирибонуклеиновых кислот оказало большое влияние то обстоятельство, что, тогда как рибонуклеопротеиды и рибонуклеиновые кислоты растворимы в разбавленном (0,15 М) растворе хлористого натрия, дезоксирибонуклеопротеидные комплексы фактически в нем нерастворимы.
Поэтому гомогенизированный орган или организм тщательно промывают разбавленным солевым раствором, из остатка с помощью крепкого солевого раствора экстрагируют дезоксирибонуклеиновую кислоту, которую осаждают затем добавлением этанола.
В клетках эукариот (например, животных или растений) ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органоидах (митохондриях и пластидах). В клетках прокариотических организмов (бактерий и архей) кольцевая или линейная молекула ДНК, так называемый нуклеотид, прикреплена изнутри к клеточной мембране. У них и у низших эукариот (например, дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами. Кроме того, одно- или двухцепочечные молекулы ДНК могут образовывать геном ДНК-содержащих вирусов. 3. Получение нуклеиновых кислот В клетках нуклеиновые кислоты связаны с белками, образуя нуклеопротеиды. Выделение нуклеиновых кислот сводится к очистке их от белков. Для этого препараты, содержащие нуклеиновые кислоты, обрабатывают ПАВ и экстрагируют белки фенолом. Послед, очистка и фракционирование нуклеиновых кислот проводятся с помощью ультрацентрифугирования, различных видов жидкостной хроматографии и гель - электрофореза. Для получения индивидуальных нуклеиновых кислот обычно используют различные варианты последнего метода.
Современные методы химического синтеза нуклеиновых кислот позволяют получать крупные фрагменты ДНК, в том числе целые гены. Методические основы химически - ферментативных методов синтеза ДНК разработаны X. Кораной.
Они включают:
химический синтез комплементарных, взаимоперекрывающихся олигонуклеотидов, из которых затем в результате комплементационных взаимодействий выстраиваются дуплексы - фрагменты молекулы синтезируемой ДНК с несовпадающими разрывами в обеих цепях;соединение (лигирование) таких олигонуклеотидов в составе дуплекса с помощью фермента Т4 ДНК-лигазы. Сборку протяженных ДНК из синтетически однотяжевых олигонуклеотидов проводят в несколько этапов. Сначала собирают небольшие дуплексы с "липкими" концами (однотяжевыми комплементарными участками), из которых затем последовательно формируют более протяженные структуры. Таким образом могут быть получены искусственные фрагменты ДНК большой длины и с любой нуклеотидной последовательностью. С помощью генетической инженерии возможно клонирование (получение в индивидуальном виде и размножение) искусственных ДНК.
Несмотря на малую эффективность этого метода, были синтезированы олигонуклеотиды, содержащие до 16 звеньев, из которых были собраны первые синтетические гены. Фосфодиэфирный метод образования межнуклеотидных связей, использованный Кораной, имеет историческое значение. Однако разработанные им приемы введения и избирательные удаления защитных групп широко используются в других методах синтеза нуклеиновых кислот.
Важным шагом в совершенствовании синтеза олигонуклеотидов явилась разработка так называемого фосфотриэфирного метода. Образующийся динуклеотид после частичного деблокирования фосфата конденсируют аналогичным образом с другими динуклеотидом и т.д. Применение этого способа, в котором используют защиту фосфатной группы, позволило значительно сократить время синтеза и повысить выходы олигонуклеотидов.
Параллельно этим методам, которые осуществляют в растворах, разрабатывались твердофазные способы синтеза нуклеиновых кислот. В последнем случае процесс проводят в двухфазной системе; нуклеозидный компонент связан ковалентно с нерастворимым полимером, а нуклеотидный компонент и необходимые реагенты находятся в растворе.
Обычно в этом случае на первой стадии нуклеозид присоединяют с помощью "якорной" группы к нерастворимому полимеру. Затем его 5'-гидроксильную группу деблокируют и конденсируют с нуклеотидным компонентом. У образующегося полностью защищенного динуклеозидмонофосфата деблокируют защитную группу в положении 5' и присоединяют следующему нуклеотид и т.д.
Наиболее распространенные методы твердофазного синтеза олигонуклеотидов основаны на использовании нуклеотидного компонента, содержащего Р(III). В так называемом амидофосфитном способе нуклеотидным компонентом является эфир 3'-амидофосфита дезоксинуклеозида. Достаточно устойчивые амидофосфиты при протонировании в присутствии тетразола превращаются в сильные фосфорилирующие агенты. После завершения синтеза удаляют защитные группы с межнуклеотидных фосфатов, отделяют олигонуклеотид от носителя, деблокируют группы Nh3 гетероциклов. Липофильную группу (МеО)2Тr удаляют после первого хроматографического разделения.
Стандартность операций в твердофазном синтезе олигонуклеотидов явилась основой для автоматизации процесса. Принцип
referat.co
Реферат : Нуклеиновые кислоты (работа 3)
Нуклеиновые кислоты
Виды нуклеиновых кислот. Нуклеиновые кислоты — фосфорсодержащие биополимеры живых организмов, обеспечивающие хранение и передачу наследственной информации. Они были открыты в 1869 г. швейцарским биохимиком Ф. Мишером в ядрах лейкоцитов, сперматозоидов лосося. Впоследствии нуклеиновые кислоты обнаружили во всех растительных и животных клетках, вирусах, бактериях и грибах.
В природе существует два вида нуклеиновых кислот — дезок-сирибонуклеиновые (ДНК) и рибонуклеиновые (РНК). Различие в названиях объясняется тем, что молекула ДНК содержит пяти-углеродный сахар дезоксирибозу, а молекула РНК— рибозу. В настоящее время известно большое число разновидностей ДНК и РНК, отличающихся друг от друга по строению и значению в метаболизме.
ДНК находится преимущественно в хромосомах клеточного ядра (99% всей ДНК клетки), а также в митохондриях и хлоропластах. РНК входит в состав рибосом; молекулы РНК содержатся также в цитоплазме, матриксе пластид и митохондрий.
Нуклеотиды — структурные компоненты нуклеиновых кислот. Нуклеиновые кислоты представляют собой биополимеры, мономерами которых являются нуклеотиды.
Нуклеотиды —сложные вещества. В состав каждого нуклео-тида входит азотистое основание, пятиуглеродный сахар (рибоза или дезоксирибоза) и остаток фосфорной кислоты.
Существует пять основных азотистых оснований: аденин, гуанин, урацил, тимин и цитозин. Первые два являются пуриновыми; их молекулы состоят из двух колец, первое содержит пять членов, второе — шесть. Следующие три являются пиримидинами и имеют одно пятичленное кольцо.
Вот как выглядит, например, формула тимидилового нуклеотида(тимидин):

Названия нуклеотидов происходят от названия соответствующих азотистых оснований; и те и другие обозначаются заглавными буквами: аденин — аденилат (А), гуанин — гуанилат (Г), цитозин — цитидилат (Ц), тимин — тимидилат (Т), урацил — уридилат (У).
Количество нуклеотидов в молекуле нуклеиновых кислот бывает разным — от 80 в молекулах транспортных РНК до нескольких сотен миллионов у ДНК.
ДНК. Молекула ДНК состоит из двух полинуклеотидных, спирально закрученных относительно друг друга цепочек.
В состав нуклеотидов молекулы ДНК входят четыре вида азотистых оснований: аденин, гуанин, тимин и цитоцин. В полинук-леотидной цепочке соседние нуклеотиды связаны между собой ковалентными связями, которые образуются между фосфатной группой одного нуклеотида и З'-гидроксильной группой пентозы другого. Такие связи называются фосфодиэфирными. Фосфатная группа образует мостик между З'-углеродом одного пентоз-ного цикла и 5-углеродом следующего. Остов цепей ДНК образован, таким образом, сахарофосфатными остатками (рис. 1.2).
Хотя в состав ДНК входит четыре типа нуклеотидов, благодаря различной последовательности их расположения в длинной цепочке достигается огромное разнообразие этих молекул.
Полинуклеотидная цепь ДНК закручена в виде спирали наподобие винтовой лестницы и соединена с другой, комплементарной ей цепью с помощью водородных связей, образующихся между адени-ном и тимином (две связи), а также гуанином и цитозином (три связи). Нуклеотиды А и Т, Г и Ц называются комплементарными.

Рис 1.2. Фрагмент молекулы ДНК (между А—Т— две водородные связи; между Г—Ц — три водородные связи).
В результате у всякого организма число адениловых нуклеотидов равно числу тимидиловых, а число гуаниловых — числу цитидиловых. Эта закономерность получила название «правило Чаргаффа». Благодаря этому свойству последовательность нуклеотидов в одной цепи определяет их последовательность в другой. Такая способность к избирательному соединению нуклеотидов называется комплемен-тарностью, и это свойство лежит в основе образования новых молекул ДНК на базе исходной молекулы (репликации, т. е. удвоения).
Цепи в молекуле ДНК противоположно направлены (антипа-раллелъностъ). Так, если для одной цепи мы выбираем направление от З'-конца к 5'-концу, то вторая цепь с таким направлением будет ориентирована противоположно первой — от 5-конца к З'-концу, иначе говоря, «голова» одной цепи соединяется с «хвостом» другой и наоборот.
Впервые модель молекулы ДНК была предложена в 1953 г. американским ученым Дж. Уотсоном и англичанином Ф. Криком на основе данных Э. Чаргаффа о соотношении пуриновых и пиримидиновых оснований молекул ДНК и результатов рентге-но-структурного анализа, полученных М. Уилкинсом и Р. Франклин. За разработку двухспиральной модели молекулы ДНК Уот-сон, Крик и Уилкинс были удостоены в 1962 г. Нобелевской премии.
ДНК — самые крупные биологические молекулы. Их длина составляет от 0,25 (у некоторых бактерий) до 40 мм (у человека). Это значительно больше самой крупной молекулы белка, которая в развернутом виде достигает длины не более 100—200 нм. Масса молекулы ДНК составляет 6x10-12 г.
Диаметр молекулы ДНК 2 нм, шаг спирали 3,4 нм; каждый виток спирали содержит 10 пар нуклеотидов. Спиральная структура поддерживается многочисленными водородными связями, возникающими между комплементарными азотистыми основаниями, и гидрофобными взаимодействиями. Молекулы ДНК эука-риотических организмов линейны. У прокариот ДНК, напротив, замкнута в кольцо и не имеет ни 3-, ни 5-концов.
При изменении условий ДНК, подобно белкам, может под-. вергаться денатурации, которая называется плавлением. При постепенном возврате к нормальным условиям ДНК ренатурирует.
Функцией ДНК является хранение, передача и воспроизведение в ряду поколений генетической информации. В ДНК любой клетки закодирована информация обо всех белках данного организма, о том, какие белки, в какой последовательности и в каком количестве будут синтезироваться. Последовательность аминокислот в белках записана в ДНК так называемым генетическим (триплетным) кодом.
Основным свойством ДНК является ее способность к репликации.
Репликация — это процесс самоудвоения молекул ДНК, происходящий под контролем ферментов. Репликация осуществляется перед каждым делением ядра. Начинается она с того, что спираль ДНК временно раскручивается под действием фермента ДНК-полимеразы. На каждой из цепей, образовавшихся после разрыва водородных связей, по принципу комплементарности синтезируется дочерняя цепь ДНК. Материалом для синтеза служат свободные нуклеотиды, которые есть в ядре (рис. 1.3).

Рис. 1.3.. Схема репликации ДНК
Таким образом, каждая полинуклеотидная цепь выполняет роль матрицы для новой комплементарной цепи (поэтому процесс удвоения молекул ДНК относится к реакциям матричного синтеза). В результате получается две молекулы ДНК, у каждой из которых ' одна цепь остается от родительской молекулы (половина), а другая — вновь синтезированная. Причем одна новая цепь синтезируются сплошной, а вторая — сначала в виде коротких фрагментов, которые затем сшиваются в длинную цепь специальным ферментом—ДНК-лигазой. В результате репликации две новые молекулы ДНК представляют собой точную копию исходной молекулы.
Биологический смысл репликации заключается в точной передаче наследственной информации от материнской клетки к дочерним, что и происходит при делении соматических клеток.
РНК. Строение молекул РНК во многом сходно со строением молекул ДНК. Однако имеется и ряд существенных отличий. В молекуле РНК вместо дезоксирибозы в состав нуклеотидов входит рибоза, вместо тимидилового нуклеотида (Т) — уридило-вый (У). Главное отличие от ДНК состоит в том, что молекула РНК представляет собой одну цепь. Однако ее нуклеотиды способны образовывать водородные связи между собой (например, в молекулах тРНК, рРНК), но в этом случае речь идет о внутри-цепочечном соединении комплементарных нуклеотидов. Цепочки РНК значительно короче ДНК.
В клетке существует несколько видов РНК, которые различаются по величине молекул, структуре, расположению в клетке и функциям:
Информационная (матричная) РНК(иРНК). Этот вид наиболее разнороден по размерам и структуре. иРНК представляет собой незамкнутую полинуклеотидную цепь. Она синтезируется в ядре при участии фермента РНК-полимеразы, комплементарна участку ДНК, на котором происходит ее синтез. Несмотря на относительно низкое содержание (3—5% РНК клетки), она выполняет важнейшую функцию в клетке: служит в качестве матрицы для синтеза белков, передавая информацию об их структуре с молекул ДНК. Каждь|й белок клетки кодируется специфической иРНК, поэтому число их типов в клетке соответствует числу видов белков.
Рибосомная РНК (рРНК). Это одноцепочечные нуклеиновые кислоты, образующие в комплексе с белками рибосомы — орга-неллы, на которых происходит синтез белка. Рибосомные РНК синтезируются в ядре. Информация об их структуре закодирована в участках ДНК, которые расположены в области вторичной перетяжки хромосом. Рибосомные РНК составляют 80% всей РНК клетки, поскольку в клетке имеется огромное количество рибосом. Рибосомные РНК обладают сложной вторичной и третичной структурой, образуя петли на комплементарных участках, что приводит к самоорганизации этих молекул в сложное по форме тело. В состав рибосом входит три типа рРНК у прокариот и четыре типа рРНК у эукариот. 3. Транспортная (трансферная) РНК(тРНК). Молекула тРНК состоит в среднем из 80 нуклеотидов. Содержание тРНК в клетке — около 15% всей РНК. Функция тРНК — перенос аминокислот к месту синтеза белка. Число различных типов тРНК в клетке невелико (20—60). Все они имеют сходную пространственную организацию. Благодаря внутрицепочечкым водо-родным связям молекула тРНК приобретает характерную вторичную структуру, называемую клеверным листам. Трехмерная же модель тРНК выглядит несколько иначе. В тРНК выделяют четыре петли: акцепторную (служит местом присоединения аминокислоты), антикодоновую (узнает кодон в иРНК в процессе трансляции) и две боковые.
Источник : Н.А. Лемеза Л.В.Камлюк Н.Д. Лисов "Пособие по биологии для поступающих в ВУЗы"
Список литературы
Для подготовки данной работы были использованы материалы с сайта http://biology.asvu.ru
topref.ru
Реферат - Нуклезоиды, нуклеотиды и нуклеиновые кислоты
НУКЛЕОЗИДЫ, НУКЛЕОТИДЫ И НУКЛЕИНОВЫЕ
КИСЛОТЫ, ИХ СТРУКТУРА И СВОЙСТВА
Нуклеозиды и нуклеотиды являются продуктами гидролиза нуклеиновых кислот, но они присутствуют в живых организмах также в несвязанном состоянии, выполняя исключительно важную роль в обмене веществ.
Нуклеозиды
— это природные гликозиды гетероциклических азотистых оснований (пиримидиновых и пуриновых ), которые связаны с пентозами через атом азота. В зависимости от природы углеводного остатка (пентозы) различают рибонуклеозиды и
дезоксирибонуклеозиды.

Пиримидиновые основания:

Пуриновые основания:


Названия нуклеозидов
производятся от тривиального названия соответствующего гетероциклического азотистого основания с суффиксами - идин у пиримидиновых и
- озин у пуриновых нуклеозидов. Исключение их этого правила сделанодля нук-
леозидов тимина.

Нуклеозиды сокращенно чаще обозначают однобуквенным индексом, но существует также система трехбуквенного индекса.
Нуклеозиды, являясь N-гликозидами, устойчивы к гидролизу в слабощелочной среде, но расщепляются в кислой среде. Пуринонуклеозиды гидролизуются легко, пирими-диновые — труднее.
Нуклеотиды
являются сложными эфирами нуклеозидов и фосфорной кислоты, которая обычно этерифицирует гидроксогруппы при С-5' пентозы. В связи с наличием в молекуле остатка фосфорной кислоты нуклеотиды проявляют свойства двухосновной кислоты с pKa1 = 0,9 — 1,5 и рKaП = 6 — 6,5.

Нуклеотиды называют или как соответствующие кислоты (монозамещенные производные фосфорной кислоты), или как соли (монофосфаты) с указанием в обоих случаях положения фосфатного остатка:

Большое значение в живых системах играют нуклеотиды, содержащие в своем составе ди- и трифосфатные группировки. Важнейшими среди этих производных являются аденозиндифосфат (АДФ) и аденозинтрифосфат (АТФ), которые способны к взаимо-превращениям путем наращивания или отщепления фосфатных групп:

В этих соединениях фосфатные группы в физиологических условиях почти полностью ионизованы, поэтому их часто записывают в виде анионов АТФ4-, АДФ3-. Главная особенность этих нуклеотидов состоит в том, что их полифосфатные группы содержат одну или две ангидридные группы
—Р—О—Р—
II II
О О
. При гидролизе такой группы разрывается связь, называемая макроэргической, и выделяется около 33 кДж/моль. Именно с этим связана роль АТФ в клетке как поставщика химической энергии для биохимических и физиологических процессов.
При участии АТФ в организме также осуществляется реакция фосфорилирования гидроксилсодержащих соединений с образованием сложных эфиров фосфорной кислоты:

При фосфорилировании карбоксилсодержащих соединений образуются ацилфосфаты, которые содержат ангидридную группировку:

 – С—О—Р
– С—О—Р
׀׀ ׀׀
О О

Фосфорилированные производные выступают активными метаболитами во многих биохимических процессах.
. Важнейшая биологическая роль нуклеотидов заключается в том, что из них построены нуклеиновыекислоты (полинуклеотиды).
Нуклеиновые кислоты -
в живых организмах играют главную роль в передаче наследственных признаков (генетической информации) и управлении процессом биосинтеза белка. Нуклеиновые кислоты — высокомолекулярные соединения с молекулярной массой от 20 тысяч до десятка миллиардов. Их полимерные цепи построены из мономерных звеньев -нуклеотидов.
Особенность нуклеотидного звена заключается в том, что оно представляет собой трехкомпонентное образование, состоящее из гетероциклического азотсодержащего основания, углеводного компонента – пентозы (β, D-дезоксирибозы или β, D-рибозы) — и фосфатной группы. Каркас полимерной цепи состоит из чередующихся пентозных и фосфатных остатков, связанных сложноэфирными связями (у С-3' и С-5'), а гетероциклические основания являются «боковыми» группами, присоединенными к пентозным остаткам за счет N-гликозидной cвязи:

Первичная структура нуклеиновых кислот определяется природой и последовательностью нуклеотидных звеньев, связанных сложноэфирными связями между пентозами и фосфатными группами (рис.1).
Вторичная структура нуклеиновых кислот . Согласно вторичной структуре полинуклеотидная цепь представляет собой двойную спираль, в которой пуриновые и пиримидиновые основания направлены внутрь. Между пуриновым основанием одной цепи и пиримидиновым основанием другой цепи имеются водородные связи, стабилизирующие такую структуру.
Основания, образующие пары, связанные водородными связями, называются комплементарными. В ДНК комплементарными будут: аденин — тимин, образующие между собой две водородные связи, и гуанин — цитозин, связанные тремя водородными связями (рис.2). Это означает, что пуриновым основаниям — аденину и гуанину в одной цепи будут соответствовать пиримидиновые основания — тимин и цитозин в другой цепи. Полинуклеотидные цепи, образующие двойную спираль, не идентичны, но
комплементарны между собой.

Комплементарность цепей и последовательность звеньев составляют химическую основу важнейших функций нуклеиновых кислот: ДНК — хранение и передача наследственной информации, а РНК — непосредственное участие в биосинтезе белка.
Молекулярная масса ДНК варьирует от нескольких миллионов до десятка миллиардов, у РНК — от десятка тысяч до нескольких миллионов.
Молекула ДНК, в отличие от молекулы РНК, в большинстве случаев состоит из двух комплементарных взаимозакрученных цепей. В зависимости от длины витка и угла спирали, а также ряда других ее геометрических параметров, различают более десяти разнообразных упорядоченных спиральных структур ДНК.
В стабилизации этих структур наряду с водородными связями, действующими поперек спирали, большую роль играют межмолекулярные взаимодействия, направленные вдоль спирали между соседними пространственно сближенными азотистыми основаниями. Поскольку эти взаимодействия направлены вдоль стопки азотистых оснований молекулы ДНК, их называют стэкинг-взаимодействиями. Таким образом, взаимодействия азотистых оснований между собой скрепляют двойную спираль молекулы ДНК и вдоль, и поперек ее оси.
Сильное стэкинг-взаимодействие всегда усиливает водородные связи между основаниями, способствуя уплотнению спирали. Вследствие этого молекулы воды из окружающего раствора связываются в основном с пентозофосфатным остовом ДНК, по-
лярные группы которого находятся на поверхности спирали.
При ослаблении стэкинг-взаимодействия молекулы воды, проникая внутрь спирали, конкурентно взаимодействуют с полярными группами оснований, инициируют дестабилизацию и способствуют дальнейшему распаду двойной спирали. Все это свидетельствует о динамичности вторичной структуры ДНК под воздействием компонентов окружающего раствора.
Биспиральные структуры в молекулах РНК возникают в пределах одной и той же цепи в тех зонах, где расположены комплементарные азотистые основанияаденин — урацил и гуанин-цитозин (рис.3). В результате вторичная структура молекулы РНК содержит биспиральные участки и петли, число и размеры которых определяются первичной структурой молекулы и составом окружающего раствора.
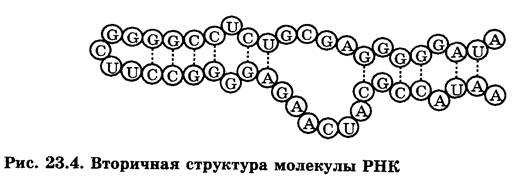
Третичная структура нуклеиновых кислот.
Двойная спираль молекул ДНК существует в виде линейной, кольцевой, суперкольцевой и компактных клубковых форм. Между этими формами совершаются взаимные переходы при действии особой группы ферментов — топоизомераз , изменяющих пространственную структуру (рис. 4).
Третичная структура многих молекул РНК пока еще требует окончательного выяснения, но уже установлено, что она зависит не только от первичной и вторичной структуры, но и от состава окружающего раствора.
Биологические функции и ДНК, и РНК полностью определяются только совокупностью первичной, вторичной и третичной структур. При этом следует отметить, что стабилизация вторичной и третичной структур нуклеиновых кислот, так же как у
белков, происходит за счет ассоциации по принципу самоорганизации под влиянием и при участии компонентов окружающего раствора, и прежде всего молекул воды.


Некоторые свойства нуклеиновых кислот
Поверхностные свойства . Макромолекулы нуклеиновых кислот состоят из полярных групп, и поэтому их поверхность достаточно гидрофильна. Вследствие этого в водных растворах нуклеиновые кислоты при их малой концентрации, низкой молекулярной массе и при достаточно большой концентрации свободных молекул воды самопроизвольно образуют истинные растворы, а в случае большой молекулярной массы — лио-
фильные коллоидные растворы.
Наличие на поверхности макромолекул нуклеиновых кислот отрицательного заряда, возникающего за счет диссоциации фосфатных групп, способствует образованию ассоциативных комплексов — нуклеопротеинов, состоящих из нуклеиновых кислот
и основных белков (рI > 8,0).
Структурно-информационные свойства . Нуклеиновые кислоты — информационные биополимеры, осуществляющие хранение и передачу генетической информации во всех живых организмах, а также участвующие в биосинтезе белков. ДНК является носителем генетической информации, которая записана через определенную последовательность расположения в цепи четырех гетероциклических оснований. Первый этап реализации генетической информации заключается в том, что на конкретных участках одной из нитей молекулы ДНК происходит синтез молекул РНК. Биосинтез РНК, называемый транскрипцией , обычно происходит в результате комплементарного копирования ДНК-матрицы с помощью фермента РНК-полимеразы. Синтезированная РНК содержит точную копию конкретного участка ДНК.
В результатетранскрипции образуются четыре различных вида РНК:
рибосомальная рРНК, матричная мРНК (информационная), транспортная тРНК и малые ядерные РНК, роль которых разнообразна, но до конца еще не выяснена. Каждая из синтезированных РНК играет строго определенную роль на втором этапе реализации генетической информации — трансляции . Реализация генетической информации с помощью нуклеиновых кислот происходит по схеме:

Рибосомальная РНК входит совместно с белками в состав рибосом. Матричная РНК, объединяясь с рибосомами, образует полирибосому, в которой с помощью ферментов и транспортных РНК, поставляющих определенные аминокислоты, происходит трансляция — синтез белков в соответствии с информацией, записанной на мРНК. Информация о последовательности аминокислот в молекуле белка считывается с последовательности ге-
тероциклических оснований в мРНК. Конкретная группа из трех гетероциклических оснований в молекуле нуклеиновой кислоты, которая соответствует отдельной аминокислоте, называется кодоном. ( Например, кодон GUA соответствует аспарагиновой кислоте, UGU- валину, UUU- фенилаланину, ACA –аспаргину). Совокупность кодонов составляет генетический код. Генетический код един для всего живого: у любого вида организмов каждая из аминокислот кодируется одним и тем же кодоном или одними и теми же кодонами. (Несколько кодонов могут кодировать одну и ту же аминокислоту, но один и тот же код не способен кодировать разные аминокислоты.)
Денатурация . Подобно денатурации белков происходит денатурация нуклеиновых кислот, сопровождаемая разрушением их третичной и вторичной структур и сохранением первичной структуры. Это происходит под влиянием тех же факторов, что и в случае белков, но интенсивность фактора в случае нуклеиновых ислот, естественно, может быть другой, чем при денатурации белка. Под воздействием того или иного фактора снижается прочность водородных связей и уменьшается эффективность стэкинг-взаимодействия между азотистыми основаниями в макромолекуле. Это способствует раскручиванию двухцепочечных спиралей с образованием неупорядоченных одноцепочечных клубков. Поскольку при денатурации сохраняется первичная структура нуклеиновых кислот, то данный процесс может иметь обратимый характер.
Кислотно-основные свойства . Сильнополярные фосфатные группы нуклеиновых кислот характеризуются значением рКа1 < 2.
Таким образом, нуклеиновые кислоты — это довольно сильные поликислоты, полностью ионизованные при рН выше 4, и поэтому их поверхность несет отрицательный заряд. Именно это обстоятельство объясняет большую склонность нуклеиновых кислот к взаимодействию с полиаминами, у которых между атомами азота содержатся две или три метиленовые (-СН2 -) группы. Однако особый интерес вызывает кислотно-основное взаимодействие нуклеиновых кислот с белками, которые являются полиамфолитами, образуя комплексные ассоциаты (соли), называемые нуклеопротеинами. Так, ДНК образует прочный комплекс с белками- гистонами, входящими в состав хромосом. Гистоны содержат 25-30 % остатков лизина и аргинина, основные функциональные группы которых при рН = 7 заряжены положительно. Они, электростатически взаимодействуя с отрицательно заряженными фосфатными группами, расположенными на периферии двойной спирали ДНК, образуют достаточно прочный комплексный ассоциат, в котором структура ДНК дополнительно стабилизирована.
Рибонуклеиновые кислоты также образуют с белками нуклеопротеины. Так, рибосомы состоят из 50-65 % рибосомной РНК и 35-50 % белков, содержащих до 25 % основных аминокислот.
Вирусы представляют собой устойчивые комплексные ассоциаты, содержащие до 30 % нуклеиновой кислоты и большое число белковых молекул, уложенных в определенном порядке и образующих специфическую трехмерную структуру. В состав вируса может входить как ДНК, так и РНК.
Окислительно-восстановительные свойства . Нуклеиновые кислоты не содержат групп, склонных к окислительно-восстановительным превращениям при мягком воздействии. Поэтому они относительно устойчивы к воздействию мягких окислителей и восстановителей. При жестком окислении в водной среде нуклеиновые кислоты превращаются, как все органические соединения в организме, в СО2 и Н2 О, а из-за присутствия в их составе атомов азота образуют мочевую кислоту, мочевину или соли аммония; кроме того, из-за наличия фосфатных групп образуются неорганические фосфаты.
www.ronl.ru
Реферат: Нуклеиновые кислоты
Нуклеиновые кислоты, биополимеры, состоящие из остатков фосфорной кислоты, сахаров и азотистых оснований (пуринов и пиримидинов). Имеют фундаментальное биологическое значение, поскольку содержат в закодированном виде всю генетическую информацию любого живого организма, от человека до бактерий и вирусов, передаваемую от одного поколения другому.
Нуклеиновые кислоты были впервые выделены из клеток гноя человека и спермы лосося швейцарским врачом и биохимиком Ф.Мишером между 1869 и 1871. Впоследствии было установлено, что существует два типа нуклеиновых кислот: рибонуклеиновая (РНК) и дезоксирибонуклеиновая (ДНК), однако их функции долго оставались неизвестными.
В 1928 английский бактериолог Ф.Гриффит обнаружил, что убитые патогенные пневмококки могут изменять генетические свойства живых непатогенных пневмококков, превращая последние в патогенные. В 1945 микробиолог О.Эвери из Рокфеллеровского института в Нью-Йорке сделал важное открытие: он показал, что способность к генетической трансформации обусловлена переносом ДНК из одной клетки в другую, а следовательно, генетический материал представляет собой ДНК. В 1940–1950 Дж.Бидл и Э.Тейтум из Станфордского университета (шт. Калифорния) обнаружили, что синтез белков, в частности ферментов, контролируется специфическими генами. В 1942 Т.Касперсон в Швеции и Ж.Браше в Бельгии открыли, что нуклеиновых кислот особенно много в клетках, активно синтезирующих белки. Все эти данные наводили на мысль, что генетический материал – это нуклеиновая кислота и что она как-то участвует в синтезе белков. Однако в то время многие полагали, что молекулы нуклеиновых кислот, несмотря на их большую длину, имеют слишком простую периодически повторяющуюся структуру, чтобы нести достаточно информации и служить генетическим материалом. Но в конце 1940-х годов Э.Чаргафф в США и Дж.Уайатт в Канаде, используя метод распределительной хроматографии на бумаге, показали, что структура ДНК не столь проста и эта молекула может служить носителем генетической информации.

Структура ДНК была установлена в 1953 М.Уилкинсом, Дж.Уотсоном и Ф.Криком в Англии. Это фундаментальное открытие позволило понять, как происходит удвоение (репликация) нуклеиновых кислот. Вскоре после этого американские исследователи А.Даунс и Дж.Гамов предположили, что структура белков каким-то образом закодирована в нуклеиновых кислотах, а к 1965 эта гипотеза была подтверждена многими исследователями: Ф.Криком в Англии, М.Ниренбергом и С.Очоа в США, Х.Кораной в Индии. Все эти открытия, результат столетнего изучения нуклеиновых кислот, произвели подлинную революцию в биологии. Они позволили объяснить феномен жизни в рамках взаимодействия между атомами и молекулами.
Типы и распространение. Как мы уже говорили, есть два типа нуклеиновых кислот: ДНК и РНК. ДНК присутствует в ядрах всех растительных и животных клеток, где она находится в комплексе с белками и является составной частью хромосом. У особей каждого конкретного вида содержание ядерной ДНК обычно одинаково во всех клетках, кроме гамет (яйцеклеток и сперматозоидов), где ДНК вдвое меньше. Таким образом, количество клеточной ДНК видоспецифично. ДНК найдена и вне ядра: в митохондриях («энергетических станциях» клеток) и в хлоропластах (частицах, где в растительных клетках идет фотосинтез). Эти субклеточные частицы обладают некоторой генетической автономией.
Бактерии и цианобактерии (сине-зеленые водоросли) содержат вместо хромосом одну или две крупные молекулы ДНК, связанные с небольшим количеством белка, и часто – молекулы ДНК меньшего размера, называемые плазмидами. Плазмиды несут полезную генетическую информацию, например содержат гены устойчивости к антибиотикам, но для жизни самой клетки они несущественны.
Некоторое количество РНК присутствует в клеточном ядре, основная же ее масса находится в цитоплазме – жидком содержимом клетки. Б льшую ее часть составляет рибосомная РНК (рРНК). Рибосомы – это мельчайшие тельца, на которых идет синтез белка. Небольшое количество РНК представлено транспортной РНК (тРНК), которая также участвует в белковом синтезе. Однако оба этих класса РНК не несут информации о структуре белков – такая информация заключена в матричной, или информационной, РНК (мРНК), на долю которой приходится лишь небольшая часть суммарной клеточной РНК.
льшую ее часть составляет рибосомная РНК (рРНК). Рибосомы – это мельчайшие тельца, на которых идет синтез белка. Небольшое количество РНК представлено транспортной РНК (тРНК), которая также участвует в белковом синтезе. Однако оба этих класса РНК не несут информации о структуре белков – такая информация заключена в матричной, или информационной, РНК (мРНК), на долю которой приходится лишь небольшая часть суммарной клеточной РНК.
Генетический материал вирусов представлен либо ДНК, либо РНК, но никогда обеими одновременно.
Общие свойства
Молекулы нуклеиновых кислот содержат множество отрицательно заряженных фосфатных групп и образуют комплексы с ионами металлов; их калиевая и натриевая соли хорошо растворимы в воде. Концентрированные растворы нуклеиновых кислот очень вязкие и слегка опалесцируют, а в твердом виде эти вещества белые. Нуклеиновые кислоты сильно поглощают ультрафиолетовый свет, и это свойство лежит в основе определения их концентрации. С этим же свойством связан и мутагенный эффект ультрафиолетового света.
Длинные молекулы ДНК хрупки и легко ломаются, например при продавливании раствора через шприц. Поэтому работа с высокомолекулярными ДНК требует особой осторожности.
Химическая структура. Нуклеиновые кислоты это длинные цепочки, состоящие из четырех многократно повторяющихся единиц (нуклеотидов). Их структуру можно представить следующим образом:

Символ Ф обозначает фосфатную группу. Чередующиеся остатки сахара и фосфорной кислоты образуют сахарофосфатный остов молекулы, одинаковый у всех ДНК, а огромное их разнообразие обусловливается тем, что четыре азотистых основания могут располагаться вдоль цепи в самой разной последовательности.
Сахаром в нуклеиновых кислотах является пентоза; четыре из пяти ее углеродных атомов вместе с одним атомом кислорода образуют кольцо. Атомы углерода пентозы обозначают номерами от 1 до 5. В РНК сахар представлен рибозой, а в ДНК дезоксирибозой, содержащей на один атом кислорода меньше. Фрагменты полинуклеотидных цепей ДНК и РНК показаны на рисунке.
Поскольку фосфатные группы присоединены к сахару асимметрично, в положениях 3 и 5, молекула нуклеиновой кислоты имеет определенное направление. Сложноэфирные связи между мономерными единицами нуклеиновых кислот чувствительны к гидролитическому расщеплению (ферментативному или химическому), которое приводит к высвобождению отдельных компонентов в виде небольших молекул.
Азотистые основания – это плоские гетероциклические соединения. Они присоединены к пентозному кольцу по положению 1. Более крупные основания имеют два кольца и называются пуринами: это аденин (А) и гуанин (Г). Основания, меньшие по размерам, имеют одно кольцо и называются пиримидинами: это цитозин (Ц), тимин (Т) и урацил (У). В ДНК входят основания А, Г, Т и Ц, в РНК вместо Т присутствует У. Последний отличается от тимина тем, что у него отсутствует метильная группа (Ch4). Урацил встречается в ДНК некоторых вирусов, где он выполняет ту же функцию, что и тимин.
Трехмерная структура. Важной особенностью нуклеиновых кислот является регулярность пространственного расположения составляющих их атомов, установленная рентгеноструктурным методом. Молекула ДНК состоит из двух противоположно направленных цепей (иногда содержащих миллионы нуклеотидов), удерживаемых вместе водородными связями между основаниями:

Водородные связи, соединяющие основания противоположных цепей, относятся к категории слабых, но благодаря своей многочисленности в молекуле ДНК они прочно стабилизируют ее структуру. Однако если раствор ДНК нагреть примерно до 60 С, эти связи рвутся и цепи расходятся – происходит денатурация ДНК (плавление).
Обе цепи ДНК закручены по спирали относительно воображаемой оси, как будто они навиты на цилиндр. Эта структура называется двойной спиралью. На каждый виток спирали приходится десять пар оснований.
Правило комплементарности. Уотсон и Крик показали, что образование водородных связей и регулярной двойной спирали возможно только тогда, когда более крупное пуриновое основание аденин (А) в одной цепи имеет своим партнером в другой цепи меньшее по размерам пиримидиновое основание тимин (Т), а гуанин (Г) связан с цитозином (Ц). Эту закономерность можно представить следующим образом:

Соответствие АТ и ГЦ называют правилом комплементарности, а сами цепи комплементарными. Согласно этому правилу, содержание аденина в ДНК всегда равно содержанию тимина, а количество гуанина – количеству цитозина. Следует отметить, что две цепи ДНК, различаясь химически, несут одинаковую информацию, поскольку вследствие комплементарности одна цепь однозначно задает другую.
Структура РНК менее упорядочена. Обычно это одноцепочечная молекула, хотя РНК некоторых вирусов состоит из двух цепей. Но даже такая РНК более гибка, чем ДНК. Некоторые участки в молекуле РНК взаимно комплементарны и при изгибании цепи спариваются, образуя двухцепочечные структуры (шпильки). В первую очередь это относится к транспортным РНК (тРНК). Некоторые основания в тРНК подвергаются модификации уже после синтеза молекулы. Например, иногда происходит присоединение к ним метильных групп.
Функция нуклеиновых кислот
Одна из основных функций нуклеиновых кислот состоит в детерминации синтеза белков. Информация о структуре белков, закодированная в нуклеотидной последовательности ДНК, должна передаваться от одного поколения к другому, и поэтому необходимо ее безошибочное копирование, т.е. синтез точно такой же же молекулы ДНК (репликация).
Репликация и транскрипция. С химической точки зрения синтез нуклеиновой кислоты – это полимеризация, т.е. последовательное присоединение строительных блоков. Такими блоками служат нуклеозидтрифосфаты; реакцию можно представить следующим образом:

Энергия, необходимая для синтеза, высвобождается при отщеплении пирофосфата, а катализируют реакцию особые ферменты – ДНК-полимеразы.
В результате такого синтетического процесса мы получили бы полимер со случайной последовательностью оснований. Однако большинство полимераз работает только в присутствии уже существующей нуклеиновой кислоты –матрицы, диктующей, какой именно нуклеотид присоединится к концу цепи. Этот нуклеотид должен быть комплементарен соответствующему нуклеотиду матрицы, так что новая цепь оказывается комплементарной исходной. Используя затем комплементарную цепь в качестве матрицы, мы получим точную копию оригинала.
ДНК состоит из двух взаимно комплементарных цепей. В ходе репликации они расходятся, и каждая из них служит матрицей для синтеза новой цепи:

Так образуются две новые двойные спирали с той же последовательностью оснований, что и у исходной ДНК. Иногда в процессе репликации происходит «сбой», и возникают мутации .
В результате транскрипции ДНК образуются клеточные РНК (мРНК, рРНК и тРНК):

Они комплементарны одной из цепей ДНК и являются копией другой цепи, за исключением того, что место тимина у них занимает урацил. Таким способом можно получить множество РНК-копий одной из цепей ДНК.
В нормальной клетке передача информации осуществляется только в направлении ДНК ДНК и ДНК РНК. Однако в клетках, инфицированных вирусом, возможны и другие процессы: РНК РНК и РНК ДНК. Генетический материал многих вирусов представлен молекулой РНК, обычно одноцепочечной. Проникнув в клетку-хозяина, эта РНК реплицируется с образованием комплементарной молекулы, на которой, в свою очередь, синтезируется множество копий исходной вирусной РНК:

Вирусная РНК может транскрибироваться ферментом обратной транскриптазой в ДНК, которая иногда включается в хромосомную ДНК клетки-хозяина. Теперь эта ДНК несет вирусные гены, и после транскрипции в клетке может появиться вирусная РНК. Таким образом, спустя длительное время, в течение которого никакого вируса в клетке не обнаруживается, он снова в ней появится без повторного заражения. Вирусы, генетический материал которых включается в хромосому клетки-хозяина, часто являются причиной рака.
Трансляция нуклеиновых кислот в белки. Генетическая информация, закодированная в нуклеотидной последовательности ДНК, переводится не только на язык нуклеотидной последовательности РНК, но и на язык аминокислот – мономерных единиц белков.
Белковая молекула – это цепочка из аминокислот. Каждая аминокислота содержит кислую карбоксильную группу –COOH и оснвную аминогруппу
–Nh3. Карбоксильная группа одной аминокислоты связывается с аминогруппой другой, образуя амидную связь, и этот процесс продолжается, пока не образуется цепь, содержащая до 1000 аминокислот .
В белках присутствует 20 разных аминокислот, от последовательности которых зависят их природа и функции. Эта последовательность определяется нуклеотидной последовательностью соответствующего гена – участка ДНК, кодирующего данный белок. Однако сама ДНК не является матрицей при синтезе белка. Сначала она транскрибируется в ядре с образованием матричной РНК (мРНК), которая диффундирует в цитоплазму, и на ней как на матрице синтезируется белок. Процесс ускоряется благодаря тому, что на каждой молекуле мРНК может одновременно синтезироваться множество белковых молекул.
Репликация нуклеиновых кислот осуществляется благодаря образованию водородных связей между комплементарными основаниями исходной и дочерней цепей. Аминокислоты не образуют водородных связей с основаниями, так что прямое копирование матрицы невозможно. Они взаимодействуют с матрицей опосредованно, через «адапторные» нуклеиновые кислоты – небольшие молекулы транспортных РНК (тРНК), состоящие примерно из 80 оснований и способные связываться с мРНК.
Каждая тРНК содержит специфическую последовательность из трех оснований, антикодон, который комплементарен группе из трех оснований, кодону, в мРНК. Антикодоны взаимодействуют с кодонами по правилу комплементарности, примерно так же, как взаимодействуют две цепи ДНК. Таким образом, последовательность оснований в мРНК определяет порядок присоединения тРНК, несущих аминокислоты. Схематически перенос информации от ДНК к белку можно представить следующим образом:

Последовательность оснований в ДНК задает порядок следования аминокислот в белке, поскольку каждая аминокислота присоединяется специфическим ферментом только к определенным тРНК, а те, в свою очередь, – только к определенным кодонам в мРНК. Комплексы тРНК-аминокислота связываются с матрицей по одному в каждый данный момент времени. Ниже перечислены основные этапы белкового синтеза (см. также рисунок).
1. Ферменты, называемые аминоацил-тРНК-синтетазами, присоединяют аминокислоты к соответствующим тРНК. Таких ферментов 20, по одному для каждой аминокислоты.
2. Молекула мРНК присоединяется своим первым кодоном к небольшой частице, называемой рибосомой. Рибосомы состоят из примерно равных количеств рРНК и белка. Структура и функция рибосом весьма сложны, но главная их задача – облегчение взаимодействия мРНК и тРНК и ускорение полимеризации аминокислот, связанных с разными тРНК.
3. тРНК, нагруженная аминокислотой, связывается с соответствующим кодоном мРНК, которая, в свою очередь, контактирует с рибосомой. Образуется комплекс рибосома-мРНК-тРНК-аминокислота.
4. мРНК, подобно ленте на конвейере, продвигается по рибосоме на один кодон вперед.
5. Следующая тРНК, нагруженная аминокислотой, присоединяется ко второму кодону.
6. Первая и вторая аминокислоты связываются между собой.
7. Первая тРНК отсоединяется от комплекса, и теперь вторая тРНК несет две аминокислоты, связанные между собой.
8. мРНК снова продвигается на один кодон вперед, и все события повторяются, а растущая аминокислотная цепь удлиняется на одну аминокислоту. Процесс продолжается, пока не будет достигнут последний, «стоп»-кодон и последняя тРНК не отделится от готовой белковой цепи. В бактериальных клетках цепь из 100–200 аминокислот собирается за несколько секунд. В животных клетках этот процесс занимает около минуты.
Генетический код. Итак, каждая аминокислота в белке опосредованно детерминируется определенным кодоном (группой из 3 оснований) в мРНК и в конечном счете в ДНК. Поскольку в нуклеиновых кислотах имеется четыре вида оснований, число возможных кодонов составляет 444 = 64. Соответствие между кодонами и аминокислотами, которые они кодируют, называется генетическим или биологическим кодом. Это соответствие было установлено опытным путем: к разрушенным клеткам добавляли синтетические полинуклеотиды известного состава и смотрели, какие аминокислоты включаются в белки. Позднее появилась возможность прямо сравнить последовательности аминокислот в вирусных белках и оснований в вирусных нуклеиновых кислотах. Чрезвычайно интересно, что генетический код, за редкими исключениями, одинаков для всех организмов – от вирусов до человека. Одно из таких исключений составляют изменения в генетическом коде, используемом митохондриями. Митохондрии это небольшие автономные субклеточные частицы (органеллы), присутствующие во всех клетках, кроме бактерий и зрелых эритроцитов. Предполагают, что когда-то митохондрии были самостоятельными организмами; проникнув в клетки, они со временем стали их неотъемлемой частью, но сохранили некоторое количество собственной ДНК и синтезируют несколько митохондриальных белков.
| Аланин | Аргинин | Аспарагин | Аспарагиновая кислота | |
| ГЦУ | ЦГУ | ГАУ | ААУ | |
| ГЦЦ | ЦГЦ | ГАЦ | ААЦ | |
| ГЦА | ЦГА |
|
| |
| ГЦГ | ЦГГ |
|
| |
|
| АГА |
|
| |
|
| АГГ |
|
| |
| Валин | Гистидин | Глицин | Глутаминовая кислота | |
| ГУУ | ЦАУ | ГГУ | ЦАА | |
| ГУЦ | ЦАЦ | ГГЦ | ЦАГ | |
| ГУА |
| ГГА |
| |
| ГУГ |
| ГГГ |
| |
| Глутамин | Изолейцин | Лейцин | Лизин | |
| ГАА | АУУ | УУА | ААА | |
| ГАГ | АУЦ | УУГ | ААГ | |
|
| АУА | ЦУУ |
| |
|
|
| ЦУЦ |
| |
|
|
| ЦУА |
| |
|
|
| ЦУГ |
| |
| Метионин | Пролин | Серин | Тирозин | |
| АУГ | ЦЦУ | АГУ | УАУ | |
|
| ЦЦЦ | АГЦ | УАЦ | |
|
| ЦЦА | УЦА |
| |
|
| ЦЦГ | УЦГ |
| |
|
|
| УЦУ |
| |
|
|
| УЦЦ |
| |
| Треонин | Триптофан | Фенилаланин | Цистеин | Нет |
| АЦУ | УГГ | УУУ | УГУ | УАА |
| АЦЦ |
| УУЦ | УГЦ | УАГ |
| АЦА |
|
|
| УГА |
| АЦГ |
|
|
|
|
| ГЕНЕТИЧЕСКИЙ СЛОВАРЬ: указаны аминокислоты, встречающиеся в белках, и соответствующие им кодоны в мРНК. «Буквы» в кодонах записаны в направлении 5' 3' . В этом же направлении идут транскрипции нуклеиновых кислот и синтез белка на матрице. «Нет» означает, что кодон не кодирует никаких аминокислот; такие кодоны называются «бессмысленными». Генетический словарь одинаков для всех живых организмов – от вирусов до человека. |
Вообще говоря, каждой аминокислоте соответствует более одного кодона. Большинство кодонов, кодирующих одну и ту же аминокислоту, имеют два одинаковых первых основания, но в трех случаях (для лейцина, серина и аргинина) имеются два альтернативных набора первых дублетов в кодонах, соответствующих одной и той же аминокислоте. Природа основания в третьем положении не столь важна; одна и та же аминокислота глицин может кодироваться по-разному: ГГУ, ГГЦ, ГГА и ГГГ. Однако кодоны для двух разных аминокислот могут иметь два одинаковых первых основания, и тогда различие между ними будет определяться природой третьего основания – пурином или пиримидином. Так, гистидин кодируется триплетами ЦАУ и ЦАЦ, а глутамин ЦАА и ЦАГ. Три кодона, УАА, УАГ и УГА, не кодируют никаких аминокислот и называются «бессмысленными».
Одна молекула ДНК кодирует много белковых цепей. Каждый отрезок, кодирующий одну цепь, называют цистроном. Начало и конец цистрона, а также граница раздела между ними помечаются с помощью своего рода знаков химической пунктуации. По крайней мере у бактерий в начале цистрона находится метиониновый кодон АУГ. Логично предположить, что первой аминокислотой в белке всегда должен быть метионин, но часто несколько первых аминокислот отщепляются ферментативно после окончания синтеза белка. Конец белковой цепи помечается одним или несколькими «бессмысленными» кодонами.
У бактерий (прокариот) практически вся ДНК кодирует какие-либо белки или тРНК. Однако у высших форм (эукариот) значительная часть ДНК состоит из простых повторяющихся последовательностей и «молчащих» генов, которые не транскрибируются в РНК и поэтому не транслируются в белки. Кроме того, исходно синтезированная мРНК содержит участки, не детерминирующие никаких белковых последовательностей. Такие участки (интроны), расположенные между кодирующими участками (экзонами), перед началом синтеза белка удаляются специальными ферментами. Почему в ДНК существуют эти казалось бы бесполезные сегменты – неясно; возможно, они выполняют регуляторные функции.
У простейшей Tetrahymena РНК сама удаляет свои интроны и соединяет свободные концы цепей, действуя как фермент по отношению к себе самой. Это единственное известное исключение из правила, согласно которому нуклеиновые кислоты не обладают ферментативной активностью.
Транспортные РНК и супрессия. Смысл информации, содержащейся в ДНК, если переводить ее на язык аминокислот, определяется как самой ДНК, так и считывающим механизмом, т.е. зависит не только от того, какие кодоны есть в ДНК и в какой последовательности они расположены, но также и от того, какие именно аминокислоты (и к каким тРНК) присоединяют аминоацил-тРНК-синтетазы. Конечно, природа синтетаз и тРНК тоже определяется ДНК, и в этом смысле ДНК является первичным детерминантом белковой последовательности. Тем не менее суммарная детерминация представляет собой функцию всей системы, поскольку результат зависит от исходных компонентов. Если бы соответствие между тРНК и аминокислотами было другим, смысл кодонов тоже изменился бы.
Известно, что мутации в ДНК изменяют считывающий механизм и в результате меняют – пусть и незначительно – смысл кодонов. Так, в бактерии Escherichia coli глициновая тРНК обычно узнает в мРНК кодон ГГА; мутация в ДНК, с которой транскрибируется эта тРНК, изменяет антикодон глициновой тРНК таким образом, что теперь он узнает кодон АГА, соответствующий аргинину, и в белковой молекуле вместо аргинина появляется глицин. Это не обязательно имеет фатальные последствия, поскольку не все аргинины кодируются триплетом АГА и есть аргининовые тРНК, по-прежнему узнающие «свои» АГА. В результате измененными оказываются не все белковые молекулы. Иногда такие мутации, изменяющие антикодон, подавляют (супрессируют) мутации в кодоне. Например, если в результате мутации глициновый кодон ГГА превращается в АГА, он все же может прочитываться как глицин, если антикодон глициновой тРНК, в свою очередь, изменился так, что эта тРНК стала узнавать АГА. В этом случае вторая «ошибка» устраняет первую.
Мутации, приводящие к изменению антикодонов, могут иметь разные последствия, поскольку один и тот же кодон может узнаваться несколькими тРНК. Вообще говоря, узнавание осуществляется благодаря комплементарности оснований кодона и антикодона, однако одно из оснований кодона может модифицироваться таким образом, что антикодон будет узнавать даже неполностью комплементарный кодон. В результате одна и та же тРНК может взаимодействовать с несколькими разными кодонами, кодирующими одну и ту же аминокислоту. Этот феномен неполного соответствия кодона и антикодона был назван Ф.Криком «шатанием».
Регуляция активности генов. Для организма было бы катастрофой, если бы во всех его клетках одновременно работали все гены и синтезировались все закодированные ими белки. Бактерии, например, должны все время приспосабливаться к условиям среды, синтезируя нужные ферменты. Все клетки высших организмов имеют один и тот же набор генов, но, к счастью, клетки мозга не продуцируют пищеварительные ферменты, а в хрусталике глаза не синтезируются мышечные белки.
Активность гена характеризуется тем, транскрибируется ли он с образованием соответствующей мРНК. ДНК длинная молекула, и в определенных ее участках имеются последовательности, называемые промоторами, которые распознаются специфическим транскрибирующим ферментом полимеразой. В этих участках и только в них начинается транскрипция, продолжаясь до тех пор, пока не достигнет последовательности оснований, означающей конец считывания.
Существуют особые репрессорные белки, которые связываются с ДНК поблизости от промотора в участке, называемом оператором. Образовавшийся комплекс блокирует транскрипцию, и мРНК не синтезируется. Таким образом, репрессорные белки являются ингибиторами транскрипции. С другой стороны, существуют небольшие молекулы, которые образуют комплекс с репрессорами и снимают их блокирующее действие на транскрипцию. Иными словами, они ингибируют ингибиторы. Так, у бактерий в норме отсутствуют ферменты, катализирующие расщепление некоторых сахаров; однако если один из этих сахаров появляется в среде, он образует комплекс с репрессором, ингибирование снимается и запускается синтез соответствующего фермента. Ферменты, синтез которых индуцируется собственными субстратами, называются индуцибельными. В ряде случаев, наоборот, репрессорный белок не блокирует транскрипцию мРНК, если он не связан с определенной молекулой. У бактерий некоторые ферменты, участвующие в синтезе определенных аминокислот, образуются только в отсутствие этих аминокислот, т.е. бактерии производят данные ферменты лишь по мере надобности. Если добавить в среду соответствующую аминокислоту, она образует комплекс с репрессором и активирует его, а тем самым ингибирует транскрипцию соответствующих генов. Уже образовавшаяся мРНК вскоре расщепляется, и синтез ферментов останавливается. Такие ферменты являются отрицально индуцибельными.
Поскольку репрессорные белки сами кодируются генами, работа которых, в свою очередь, может регулироваться другими генами, а синтез малых молекул-индукторов и гормонов также в конечном счете регулируется генами, механизмы регуляции генной активности могут быть очень сложными.
Список литературы
Ичас М. Биологический код. М., 1971
Шабарова З.А., Богданов А.А. Химия нуклеиновых кислот и их компонентов, М., 1978
Зенгер В. Принципы структурной организации нуклеиновых кислот. М., 1987
www.neuch.ru

|
|
..:::Счетчики:::.. |
|
|
|
|
|
|
|
|