
|
|
|
|
 Far Far |
| WinNavigator |
| Frigate |
| Norton
Commander |
| WinNC |
| Dos
Navigator |
| Servant
Salamander |
| Turbo
Browser |
|
|
| Winamp,
Skins, Plugins |
| Необходимые
Утилиты |
| Текстовые
редакторы |
| Юмор |
|
|

|
File managers and best utilites |
Читать лекция по биологии: "Цитоплазма эукариотической клетки. Мембранный принцип организации клеток". Клетка эукариотических организмов мембранный принцип ее организации реферат
Андреев в.П. Принципы структурно-функциональной организации и биология эукариотической клетки
Для всякого живого существа
клетка является последним
морфологическим элементом,
из которого исходит всякая
жизнедеятельность – нормальная,
и патологическая»
Рудольф Вирхов
Содержание лекции
Введение
Организация плазматической мембраны
Функции плазматической мембраны
Управление ионными каналами
Цитозоль
Ядро
Эндоплазматический ретикулум
Цитоскелет
Аппарат Гольджи
Везикулы транспортные
Лизосомы
Пероксисомы
Митохондрии
Протеасомы
Межклеточные взаимодействия
Межклеточные сигнальные вещества
Органоиды клетки
Цитоскелет
Передача сигналов внутри клетки
Передача внешнего сигнала в клетку
Концепции современной цитологии
Основные хронологические вехи в изучении клетки
Введение
Эукариотическая клетка – это элементарная самовоспроизводящая живая система, обладающая способностью к обмену веществом, энергией и информацией с окружающей средой. Клетки существуют в форме самостоятельных одноклеточных организмов (амеба, малярийный плазмодий и др.) или входят в состав тканей многоклеточных организмов. В многоклеточных организмах клетки представляют собой элементы, подчиненные целостному организму. Интеграцию клеток осуществляют гуморальная, нервная и иммунная система. Для адекватной реакции на регуляторные сигналы «центра» каждая клетка обладает системой управления. Согласно взглядам В.Я. Александрова и М.Д. Голубовского клетка владеет разными видами целесообразного поведения, что у человека называется эрудиция и ум. Клеточная «эрудиция» – хранение большого набора генетических программ; клеточный «ум» – способность включать в определенных условиях соответствующую программу поведения. Например, в ответ на стресс клетки генерируют изменение своих компонентов и приступают к поиску адаптивных генетических программ поведения.
Организм человека построен приблизительно из десяти триллионов клеток, которые формируют четыре основных типа тканей: эпителиальную, соединительную, мышечную и нервную. Каждая ткань представлена многочисленными клеточными типами. К настоящему времени изучена структура и функция более 200 фенотипов клеток, приспособленных для выполнения многочисленных функций. (Открытие клеточных типов продолжается; так несколько лет назад в сетчатке глаза ученые обнаружили третий тип клеток, участвующих в формировании ритма сон-бодрствование).
В эукариотической клетке, как элементарной целостной живой системе, можно выделить четыре общебиологические эволюционно обусловленные подсистемы. К ним относятся: пограничная, представленная внешней клеточной мембраной-плазмалеммой, скелетно-двигательная (цитоскелет и сократительный аппарат), гомеостатическая (ядро и комплекс органелл) и подсистема, обеспечивающая реактивные свойства за счет рецепторно-транспортных механизмов.
Организация эукариотической клетки
Эукариотическая клетка организована системой мембран. Снаружи она ограничена плазматической мембраной. Внутренний объем клетки заполнен цитоплазмой, которая разделена на хорошо различимые, окруженные внутриклеточными мембранами отделы, называемыми мембранными органеллами. В них сосредоточено около 50% объема клетки. Остальное внутриклеточное пространство занимает компартмент, называемый цитозолем ( син. матрикс цитоплазмы, гиалоплазма).
Самой крупной органеллой является ядро клетки, его можно легко видеть в световой микроскоп. В нем находятся хромосомы, и осуществляется ДНК-направляемый синтез РНК (транскрипция). Перинуклеарное пространство (лат. nucleus – ядро), ограниченное наружной и внутренней мембранами, связано с эндоплазматической сетью, представляющей собой ограниченную мембранами систему связанных друг с другом канальцами полостей в виде уплощенных мешочков и цистерн. Другая, ограниченная мембранами органелла, также представляющая собой систему мембран, – комплекс Гольджи напоминает сложенные в стопку расширенные по краям уплощенные цистерны, в которых происходит модификация и сортировка белков и липидов. Митохондрии отвечают за производство энергии в виде аденозинтрифосфата (АТФ). Лизосомы и пероксисомы – маленькие глобулярные органеллы, предназначенные для выполнения специфических функций. Экзосомы и эндосомы – пузыреобразные органеллы (везикулы), участвующие в процессе обмена веществ между клеткой и ее окружением. Кроме «мембранных», в клетке находятся и немембранные органоиды – рибосомы, клеточный центр, реснички, жгутики и др. Регуляторные, а также состарившиеся, содержащие ошибки транскрипции белки разрушаются в протеасомах. Способность клетки изменять свою форму, перемещаться, передвигать органеллы в цитоплазме и разделять хромосомы во время митоза обеспечивает система белковых нитевидных структур – микрофиламентов, актиновых филаментов, микротрубочек, – напоминающая строительные леса (так называемый цитоскелет).
Состав биомембран
Все биомембраны, несмотря на огромное разнообразие, построены однотипно; они состоят из двух слоев полярных липидных молекул, в которые встроены белки. В клеточной мембране присутствуют липиды трех главных типов – фосфолипиды, холестерол и гликолипиды. Все они являются амфипатическими, то есть обладающими двумя противоположными свойствами. Каждая молекула мембранного липида имеет заряженную гидрофильную «головку» и два незаряженных гидрофобных «хвоста». Каждый «хвост» представляет собой длинную углеводородную цепь жирной кислоты, причем у фосфолипидов одна из этих цепей – предельная, т.е. не содержит двойных связей, а вторая – непредельная имеет одну или более двойных связей. Неполярные, гидрофобные участки фосфолипидных молекул, образуемые цепями жирных кислот, обращены внутрь липидного бислоя, формируя сплошную пленку – своеобразный барьер для растворенных в воде ионов и полярных молекул. Полярные гидрофильные группы фосфолипидных молекул обращены наружу и как бы смачиваются окружающим водным раствором. Структуру такого типа называют жидкокристаллической (рис. 1).
У разных клеток состав мембранных липидов может существенно различаться. Различный липидный состав характерен и для разных мембран одной и той же клетки. Они могут включать такие липиды как фосфатидилхолин, фосфатидилэтаноламин, фосфатидилсерин, кардиолипин, триглицериды, холестерол и др., от которых в основном зависят физико-химические свойства мембран. Наиболее распространенными мембранными липидами являются производные глицерин-3-фосфата, называемые глицерофосфолипидами, в которых две гидроксильные группы глицерола замещены остатками различных или одинаковых жирных кислот. Некоторые мембраны содержат, кроме того, сахарные остатки, связанные с липидами и белками ковалентными связями. Такими углеводными остатками могут быть глюкоза, галактоза, N-ацетилгалактозамин и др.
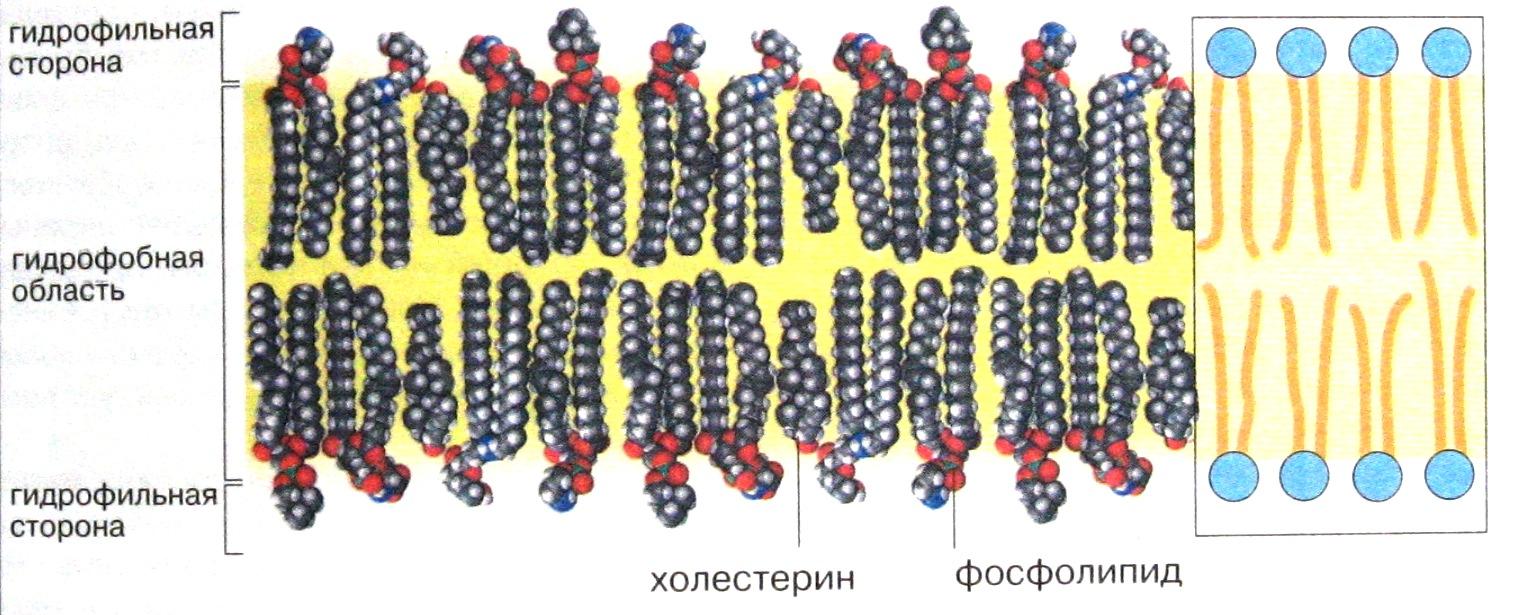
Рис.1. Мембранные липиды. (Из кн. Кольман Я., Рём К, – Г. «Наглядная биохимия»)
Жидкий кристалл – это как бы промежуточное состояние вещества между твердым кристаллом и аморфной жидкостью: в расположении образующих его молекул есть порядок, обусловленный правильной ориентацией молекул друг относительно друга. Для образования жидких кристаллов необходимо, чтобы молекулы были длинными и узкими. Такими молекулами в плазматической мембране являются жирные кислоты.
Соотношение этих компонентов является характерным для клетки или мембраны и существенно варьирует в зависимости от типа клеток или мембран. В мембранах эритроцитов имеется производное от многоатомного аминоспирта сфингозина – церамид (рис.2). К этому липиду ферменты присоединяют разветвленные цепочки различных сахаров. Такие гликолипиды называют ганглиозидами. В настоящее время идентифицирована структура более 60-ти молекул ганглиозидов. В нервных клетках ганглиев они составляют около 6 % общей массы липидов; в других типах клеток они встречаются в меньших количествах.
Все ганглиозиды выполняют важные функции в тех или иных клетках, являясь рецепторами для медиаторов, осуществляющих связь между клетками. Ганглиозиды в эритроцитах обладают антигенными свойствами и определяют группы крови людей 0, А, В-системы. Следует отметить, что молекулы ганглиозидов нередко являются «посадочными площадками» для вирусов или бактериальных токсинов, то есть выполняют несвойственные им в норме рецепторные функции. Так, холерный токсин проникает внутрь эпителиальных клеток кишечника, присоединяясь к ганглиозиду Gм1.
Плазматическая мембрана постоянно обновляется. Это происходит за счет процессов синтеза и разрушения отслуживших окисленных гликолипидов.
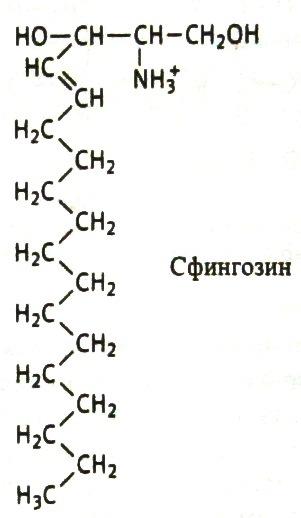
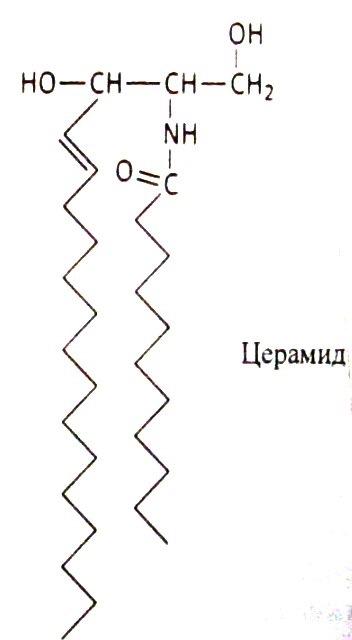
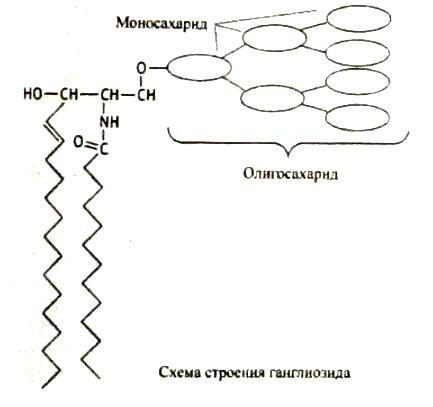
Рис. 2. Структура некоторых компонентов плазматической мембраны (из кн. В. Элиот, Д. Элиот «Биохимия и молекулярная биология».
Липиды мембран синтезируются на эндоплазматической сети и переносятся в плазматическую мембрану. Процесс переноса осуществляется транспортными пузырьками. При некоторых наследственных заболевания человека ферменты, необходимые для разрушения этих гликолипидов, оказываются дефектными, и клетка не может расщеплять гликолипиды. Это приводит к их накоплению в клетке, и, в конце концов, к гибели клетки. Заболевания, вызванные неспособностью клетки к расщеплению сложных гликолипидов, называются мукополисахаридозами.
Текучесть мембран зависит от липидного состава и температуры окружающей среды. Существенное значение для структуры клеточных мембран имеют полиненасыщенные (с двойными связями между углеродными атомами) жирные кислоты (ПНЖК). В промежутках между изгибами ПНЖК располагается спирт холестерол (холестерин). Он предотвращает кристаллизацию мембраны или переход в гель, придавая им текучесть. Однако полиненасыщенные жирные кислоты не синтезируются в организме человека и высших животных, поэтому их поступление с пищей необходимо для правильной деятельности клеточных мембран. Отсутствие ПНЖК приводит к тяжелым общим расстройствам, провалам памяти, выпадению волос, шелушению кожи. (Источниками ПНЖК являются растительные масла, не подвегнувшиеся термической обработке и рыба холодных морей).
Специфические функции биологических мембран осуществляются главным образом белками. Типы белков и их количества в мембране отражают ее функцию. Так в плазматической мембране белки составляют приблизительно 50% от ее массы, а в мембране митохондрии – 75%. По своей функциональной роли мембранные белки разделяются на ферментативные, транспортные и регуляторные. Но такое разделение носит только условный характер, поскольку, например, ионный канал может одновременно участвовать во всех этих функциях.
Большинство мембранных белков являются интегральными. Эти протеины или погружены в толщу липидного слоя, или пронизывают мембрану насквозь (трансмембранные белки). Они удерживаются в билипидном слое за счет нековалентных связей. Их гидрофильные аминокислоты взаимодействуют с фосфатными группами фосфолипидов, а гидрофобные – с цепями жирных кислот. Между белками и жирными кислотами могут возникать ковалентные связи. Для интегральных белков некоторых клеток характерна латеральная подвижность; они могут перераспределяться в мембранах в результате взаимодействия с периферическими белками, элементами цитоскелета, молекулами в мембране соседней клетки и компонентами внеклеточного матрикса. Кроме того, выделяют так называемые периферические белки, молекулы которых не встроены в мембрану, а за счет слабых взаимодействий удерживаются на ее поверхности. Периферические мембранные белки (фибриллярные, глобулярные) находятся как на наружной, так и внутренней поверхности мембраны и нековалентно – за счет водородных, гидрофобных, электростатических взаимодействий – связаны с интегральными мембранными белками. К периферическим белкам внутренней поверхности мембраны относятся белки цитоскелета (например, анкирин от лат. аnkir – якорь), которые обеспечивают механическую связь мембран с цитоскелетом (микрофиламентами и микротрубочками), семейство G-белков (гуанозинтрифосфатаз), посредством которых активируется фермент аденилатциклаза и мн. др.
Примером периферического белка наружной поверхности мембраны является фибронектин. Этот гликопротеин локализован на наружной поверхности мембраны почти всех клеток. Фибронектин обеспечивает прикрепление клеток к другим клеткам, а также способствует соединению клетки с внеклеточным матриксом. Отмечено резкое снижение содержания фибронектина в мембранах опухолевых клеток; это придает им большую подвижность и способствует метастазированию. Белки и некоторые липидные молекулы, расположенные на наружной стороне плазматической мембраны, несут ковалентно связанные углеводные компоненты – олигосахариды (рис3).
Эти гликопротеины и гликолипиды вместе с дополнительными несвязанными с мембраной гликопротеинами и полисахаридами образуют клеточную оболочку – гликокаликс. Гликокаликс, покрывающий микроворсинки каемчатых клеток эпителия кишечника, содержит ферменты, завершающие расщепление белков, углеводов (пристеночное пищеварение). Однако основные функции гликокаликса – межклеточное узнавание и межклеточные взаимодействия.
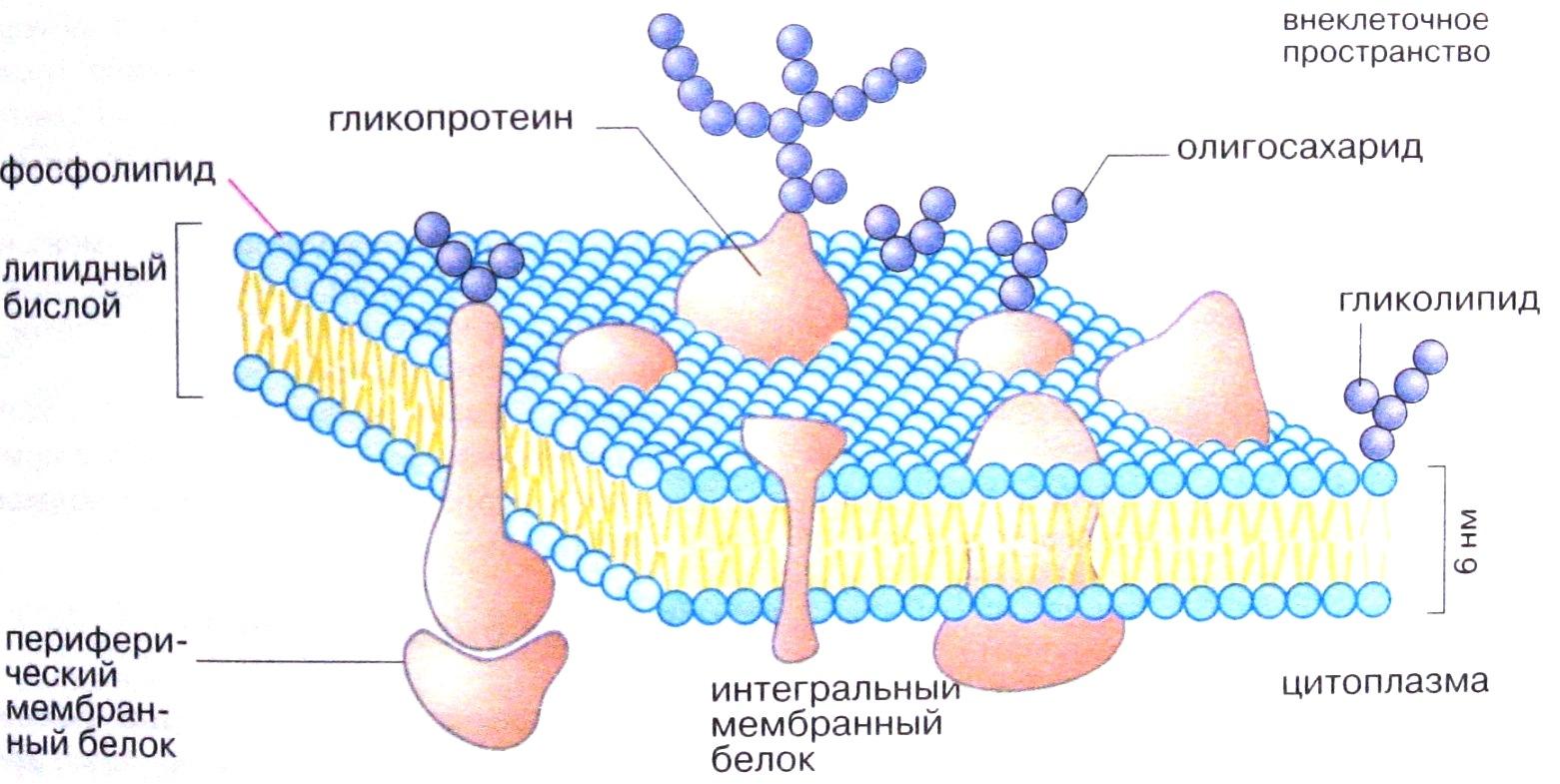
Рис.3. Структура плазматической мемраны. (Из кн. Кольман Я., Рём К, – Г. «Наглядная биохимия»)
Углеводные остатки гликопротеинов и гликолипидов наружной мембраны клетки обладают специфическими антигенными свойствами. Все клетки одного организма несут сходные поверхностные антигены, которые отличаются от поверхностных антигенов клеток любого другого организма. Это свойство используется иммунной системой для разделения всех клеток «на свои» и «чужие». Успех переливания крови или трансплантации другой ткани во многом определяют антигены групп крови 0, А, В-системы. Их антигенные свойства обусловлены структурой углеводных остатков гликолипидов на поверхности эритроцитов. (Схема строения ганглиозидов, определяющих IV, III, II и I-ю группы крови 0, А, В-системы представлена на рис. 4-7.)
Кроме этих антигенов индивидуальную поверхность клеток определяют так называемые трансплантационные антигены или антигены гистосовместимости. В этом случае антигенами служат полипептидные цепи группы трансмембранных белков. Эти белки-антигены кодируются в геноме млекопитающих многими генами, так называемого главного комплекса гистосовместимости. Бóльшая часть Т-лимфоцитов узнает чужеродные антигены только в том случае, если эти антигены ассоциированы на клеточных поверхностях с антигенами главного комплекса гистосовместимости.
Антигéны главного комплекса гистосовместимости (ГКГ) или антигены МНС (англ.majorhistocompatibilitycomplex, произносят как эм, эйч, си) – семейство антигенов, определяющих отторжение чужеродного трансплантата, т.е. тканевую несовместимость. У человека они впервые были выявлены в лейкоцитах и поэтому получили названиеHLA(англ.humanleucocyteantigen, произносят как эйч, эль, эй). Существует два основных класса антигенов (молекул) ГКГ: антигены ГКГ классаIи антигены ГКГ классаII. Все они являются мембранными гликопротеинами (рис. 8).
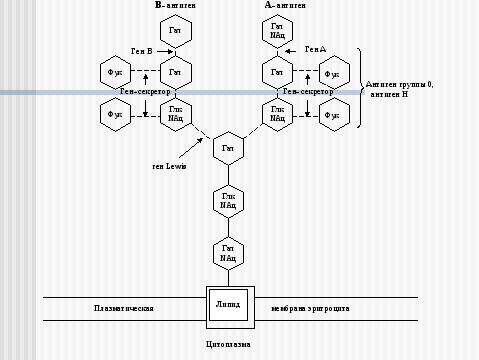
Рис. 4. Схема связи остатков сахаров с наружной мембраной эритроцита у людей с IV (АВ) группой крови 0, А, В-системы.
Глк – D-глюкоза, Гал – D-галактоза, Фук – L-фукоза, Глк NАц – N-ацетил- D-глюкозамин, Гал NАц – N-aцетил- D-галактозамин.
Прерывистые связи между остатками сахаров означают, что существует много типов соединений.
Ген Lewis, доминантный ген-секретор, ген-А, ген-В кодируют ферменты-трансферазы, переносящие и присоединяющие остатки сахаров. При наличии гена А или В, и рецессивного гена-секретора в гомозиготном состоянии антигены А и В не образуются (бомбейский феномен).
Примечание: Отличительной особенностью антигена Н является наличие его в биологических жидкостях секреторов групповых веществ и отсутствие – у несекреторов. Антиген 0, в отличие от антигена Н, А и В, с секретами (слюна, сперма) не выделяется.
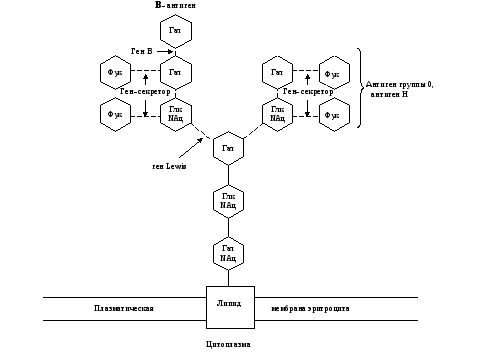
Рис. 5. Схема связи остатков сахаров с наружной мембраной эритроцита у людей с III (В) группой крови 0, А, В-системы.
Глк – D-глюкоза, Гал – D-галактоза, Фук – L-фукоза, Глк NАц – N-ацетил- D-глюкозамин, Гал NАц – N-aцетил- D-галактозамин.
Прерывистые связи между остатками сахаров означают, что существует много типов соединений.
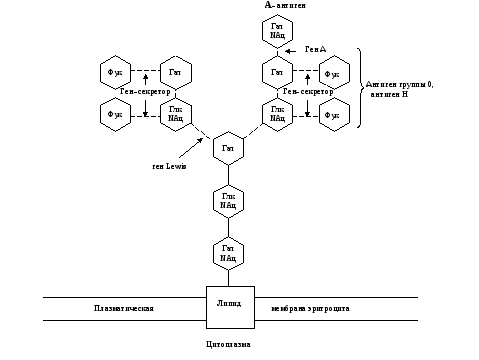
Рис. 6. Схема связи остатков сахаров с наружной мембраной эритроцита у людей с II (А) группой крови 0, А, В-системы.
Глк – D-глюкоза, Гал – D-галактоза, Фук – L-фукоза, Глк NАц – N-ацетил- D-глюкозамин, Гал NАц – N-aцетил- D-галактозамин.
Прерывистые связи между остатками сахаров означают, что существует много типов соединений.
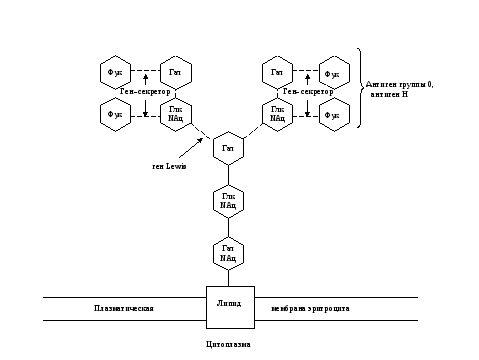
Рис. 7. Схема связи остатков сахаров с наружной мембраной эритроцита у людей с I (0) группой крови 0, А, В-системы.
Глк – D-глюкоза, Гал – D-галактоза, Фук – L-фукоза, Глк NАц – N-ацетил- D-глюкозамин, Гал NАц – N-aцетил- D-галактозамин.
Прерывистые связи между остатками сахаров означают, что существует много типов соединений.
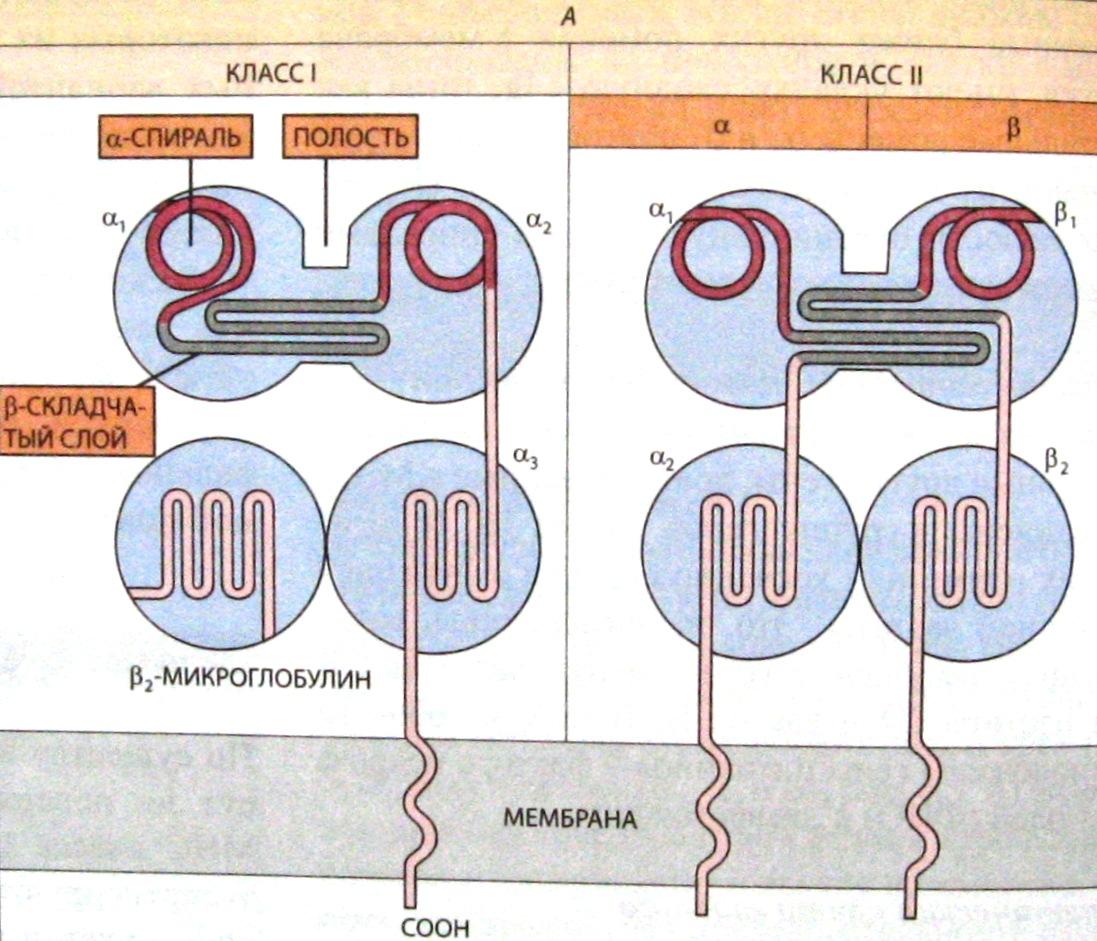
Рис. 8. Схема структуры трансмембранных гликопротеинов МНС класс 1 (слева) и МНС класса 2 (справа). (Из кн. Б. Албертс и др. «Молекулярная биология клетки», том 5.)
Класс IГКГ содержится на оболочках всех соматических клеток, обладающих ядром. Они презентируют вирусные антигены цитотоксическим Т-лимфоцитам. ГКГ классаIIсвязаны с плазматической мембраной В-лимфоцитов, макрофагов, дендритных и некоторых других антигенпредставляющих (презентирующих) клеток, которые представляют в иммунных ответах для распознавания Т-лимфоцитам переработанные ими до пептидов чужеродные антигены, вслед, за чем запускается развитие иммунных защитных реакций.
Функции мембран
Ограничение и обособление клеток и органелл. Обособление клеток от межклеточной среды обеспечивается плазматической мембраной, защищающей клетки от механического и химического воздействий.
Контролируемый транспорт метаболитов и ионов через поры и посредством переносчиков определяет внутреннюю среду, что существенно для гомеостаза, т.е. поддержания постоянной концентрации метаболитов и неорганических ионов, и других физиологических параметров.
Восприятие внеклеточных сигналов и их передача внутрь клетки, а также инициация сигналов.
Ферментативный катализ. В мембранах локализованы наиболее важные реакции энергетического обмена, такие, как окислительное фосфорилирование.
Контактное взаимодействие с межклеточным матриксом и взаимодействие с другими клетками при образовании тканей.
Заякоривание цитоскелета, обеспечивающее поддержание формы клеток и органелл и клеточной подвижности.
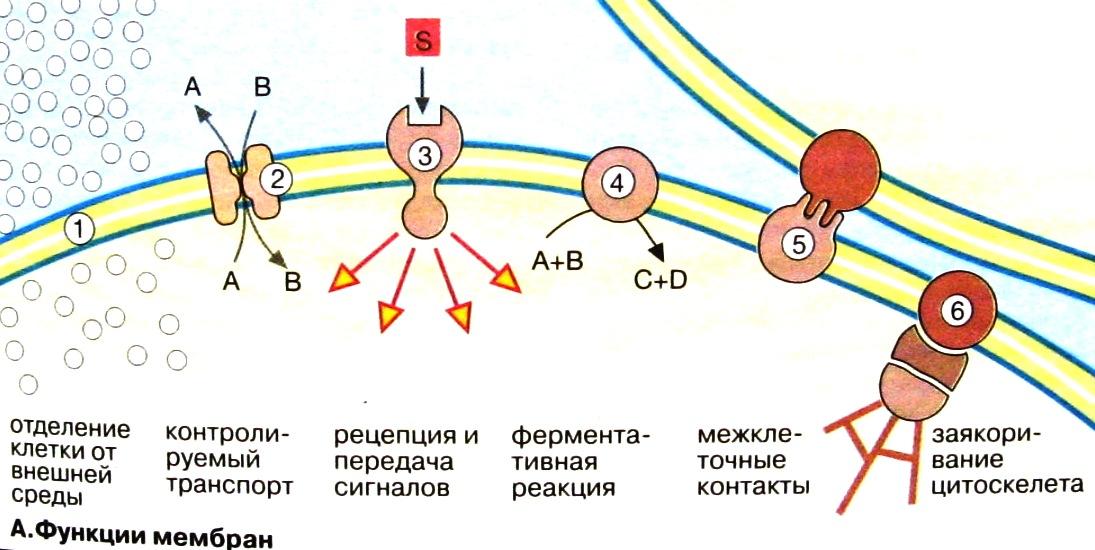
Основная функция наружной клеточной мембраны – сохранение внутренней среды клетки. При участии плазматической мембраны происходит узнавание и агрегация, как соседних клеток, так и клеток с компонентами внеклеточного матрикса. Формирование клеточной поверхности, которая способна к узнаванию других клеток, контакту с ними и восприятию разнообразных раздражителей, воздействующих на клетку, обеспечивается, прежде всего, углеводными группами, входящими в состав гликопротеидов клеточной мембраны.
studfiles.net
Читать лекция по биологии: "Цитоплазма эукариотической клетки. Мембранный принцип организации клеток"
 (Назад)
(Назад) (Cкачать работу)
(Cкачать работу)
Функция "чтения" служит для ознакомления с работой. Разметка, таблицы и картинки документа могут отображаться неверно или не в полном объёме!
Тема 15. Цитоплазма эукариотической клетки. Мембранный принцип организации клеток СОДЕРЖАНИЕ 1. Роль ученых в открытии клеточных мембран 2. Цитоплазма эукариотической клетки 3. Гиалоплазма - внутренняя среда клетки 4. Мембраны цитоплазмы 4.1. Химия и свойства клеточных мембран 4.2. Структура клеточных мембран 5. Плазматическая мембрана 5.1. Рост плазматической мембраны 6. Функции плазматической мембраны 6.1. Транспортные функции 6.2. Рецепторные функций плазматической мембраны 6.3. Межклеточные контакты 6.4. Простой контакт. Соединение типа «замка» 6.5. Плотный замыкающий контакт 6.6. Промежуточный контакт 6.7. Щелевидный контакт Список литературы 1. РОЛЬ УЧЕНЫХ В ОТКРЫТИИ КЛЕТОЧНЫХ МЕМБРАН До конца XVIII века предполагали, что клетки разделены общей стенкой и не могут быть изолированными друг от друга. В дальнейшем развитии данной науки большую роль сыграли исследования французского ботаника Бриссо де Мирбеля (1776-1854), в которых впервые высказывалась идея сравнительного изучения растительных тканей. Мирбель впервые предположил, что соприкасающиеся клетки разделены общей перегородкой - мембраной. Немецкий естествоиспытатель Рудольфи пришел к заключению об обособленности клеток, о наличии у них собственных мембран, окружающих их со всех сторон. Всякие сомнения о том, что клетки могут быть выделены и изолированы, окончательно исчезли после работы ботаника И. Молденгауера (1766-1827), из которой следовало, что клетка является самостоятельной структурой, покрытой оболочкой. Итак, сформировалось новое представление о клетке. Наиболее четко его сформулировал несколько позже Ф. Мейен. Его определение таково: «Клетка растительного организма представляет собой пространство, вполне замкнутое вегетативной мембраной». Ф. Мейен ясно изложил этот взгляд, в начале ХIХ в. сменивший первоначальное воззрение на клетку как на пору. Но конкретное содержание анатомии растений по-прежнему принципиально мало чем отличалось от старого. Исследователи продолжали системно описывать строение различных органов растений, а также детализировать и уточнять изученное. Этот период справедливо считается периодом собирания материала, периодом накопления многочисленных сведений о тончайшей структуре растений. Итак, сам факт наличия клеток в различных тканях сомнения не вызывал. Но исчерпывается ли все разнообразие структур растительных организмов только клетками? Вопрос оставался еще не до конца ясным. Более того, большинство ученых отвечало на этот вопрос отрицательно, указывая, наряду с клетками, на сосуды и волокна растений как на вполне своеобразные структуры. Обобщая все знания в этой области, Мейен прямо писал, что растительные ткани состоят из клеток, волокон и сосудов. Возникновение этих последних из метаморфизирующихся клеток было окончательно установлено лишь несколько позднее - в 30-х гг. прошлого века. 2. ЦИТОПЛАЗМА ЭУКАРИОТИЧЕСКОЙ КЛЕТКИ Цитоплазма представляет собой метаболический рабочий аппарат клетки. В ней сосредоточены общие и специальные органоиды, в цитоплазме протекают основные метаболические процессы. В последние десятилетия достигнуты особенно большие успехи в дискретном морфофункциональном анализе отдельных органоидов и мембранных систем метаболического аппарата цитоплазмы. Это делает возможным представить себе работу метаболического аппарата как целостной системы. В современных исследованиях метаболического аппарата клетки особенно ярко проявляется тенденция к изучению функционального значения морфологических структур - анализу функции через структурно-биохимическую организацию этих структур. Важнейшие успехи в этом плане достигнуты в изучении основных органоидов белкового синтеза (рибосом) и систем, обеспечивающих энергетический обмен (митохондрий, хлоропластов и сопрягающих мембран прокариотных клеток). Крупным обобщением в изучении метаболического аппарата цитоплазмы являются представления о нем как о пластичной дифференцированной трехфазной системе, состоящей из основной цитоплазмы, внутриклеточных мембран и содержимого мембранной системы. Таким образом метаболический аппарат цитоплазмы, с одной стороны, сложен специализированными структурами, выполняющими частные функции, а с другой стороны, является единой целостной системой, находящейся в тесной взаимосвязи и с поверхностным, и с ядерным аппаратами клетки. Начало такому подходу положил еще в 60-х гг. Де Дюв в учении об экзоплазматических мембранных системах. В дальнейшем этот подход развивался и в учении о лизосомах, и в современных представлениях об аппарате Гольджи и эндоплазматической сети, а также в представлениях о динамичном взаимодействии внутриклеточных мембранных систем. Весьма перспективным, на наш взгляд, является также оформившееся в последнее время представление об основной цитоплазме - гиалоплазме. В противовес бытовавшему раньше взгляду на гиалоплазму как на однородную живую коллоидную систему оно трактует основную цитоплазму как дифференцированную, гетерогенную фазу цитоплазмы, способную к формированию сложных структур. С этим свойством гиалоплазмы мы уже частично познакомились на примере субмембранной системы поверхностного аппарата с ее динамичными механо-химическими и опорными структурами. В настоящее время становится ясно, что такие примембранные системы гиалоплазмы формируются не только в области плазматических мембран, но и в области всех мембранных, а иногда даже и немембранных (клеточный центр) структур цитоплазмы. Эту часть цитоплазмы предлагается выделить в особую субсистему гиалоплазмы, обозначив ее термином «цитозоль». Естественно, что функции и морфобиохимическая организация такого цитозоля в области шероховатой и гладкой эндоплазматической сети (ЭПС), мембран аппарата Гольджи, митохондрий, клеточного центра, ядерной оболочки и т. д. будут различны и специфичны для тех структур, вокруг которых формируются соответствующие участки цитозоля. С учетом структурного и функционального разнообразия органоидов, обеспечивающих внутриклеточный метаболизм в цитоплазме клеток, функция соответствующих каждому конкретному органоиду и практически входящих в его состав участков цитозоля не будет ограничиваться только опорно-сократительной. Структуры и ферменты, входящие в состав цитозоля, специфичны для каждого органоида и принимают непосредственное участие в обеспечении протекающих в данном органоиде биохимических процессов. Рассмотрение цитоплазмы целесообразно начать с характеристики проблемы структурно-биохимической организации рибосом - молекулярных машин белкового синтеза. В этом случае особенно ярко проявляется значение структурной организации биохимических процессов в клеточных системах и достигнуты большие успехи в ее изучении. Второй областью биологии клетки, где структурная организация биохимических процессов также изучена весьма детально, является анализ так называемых сопрягающих мембран - аппарата энергетического обмена клеток. Их мы рассмотрим при характеристике митохондрий, пластид и других структур цитоплазмы, отвечающих за энергетический обмен в клетке. Далее мы обсудим результаты дискретного анализа основных мембран. Собственно тело клетки и ее содержимое отделено от внешней среды или от соседних элементов у многоклеточных организмов плазматической мембраной. Кнаружи от плазматической мембраны, экстрацеллюлярно, расположена клеточная оболочка, или стенка, особенно хорошо выраженная у растений и прокариотических организмов; у клеток животных она отсутствует или выражена очень слабо. Все внутреннее содержимое клетки, за исключением ядра, носит название цитоплазмы. Это общий термин, который подчеркивает разделение клетки на два главных компонента: цитоплазму и ядро. Цитоплазма эукариотических клеток неоднородна по своему строению и составу и включает в себя гиалоплазму, мембранные и немембранные компоненты. К мембранным компонентам относится вакуолярная система (эндоплазматическая сеть, аппарат Гольджи, лизосомы, вакуоли растений), мембранные органеллы (митохондрии и пластиды). К немембранным компонентам относятся центриоли, характерные для животных клеток, и немембранные макромолекулярные комплексы и структуры, такие как рибосомы, микротрубочки и микрофиламенты. Такое разделение
referat.co
Реферат - Типы клеточной организации. Строение про- и эукариотических клеток. Гипотезы происхождения эукариотических клеток (симбиотическая, инвагинационная).
Биологические системы, их фундаментальные свойства. Эволюционно обусловленные уровни организации жизни. Элементарные единицы, элементарные явления на различных уровнях организации жизни.
Биологические системы — биологические объекты различной сложности ( клетки, ткани, органы, системы органов и организмы, биоценозы и экосистемы, вплоть до биосферы в целом), имеющие несколько уровней структурно — функциональной организации, представляя собой совокупность взаимосвязанных и взаимодействующих элементов.
Фундаментальные свойства живых систем:
- Самовоспроизведение
- Специфичность организации( клеточные структуры, ткани, органы, системы органов)
- Упорядоченность структуры
- Обмен веществ ( препятствует увеличению энтропии)
- Рост и развитие
- Целостность и дискретность
- Раздражимость и возбудимость
- Движение
- Наследственность и изменчивость
- Саморегуляция
| Уровень организации живого | Элементарная единица | Элементарное явление |
| Молекулярно-генетический | ген | Способность гена к ковариантной редупликации (всегда появляется новое: при кроссинговере, половом размножении, мутациях) |
| Клеточный | клетка | Поток веществ ( белки, энергия АТФ, информация в виде ДНК) |
| Онтогенетический | особь | Клеточная дифференциация |
| Популяционно-видовой | популяция | Изменение генофондов |
| Экосистемный | биогеоценоз | Круговорот вещества и энергии |
| Биосферный | биосфера | Глобальный круговорот вещества и энергии |
2.Клеточная теория Т. Шванна и М. Шлейдена, её основные положения. Современное состояние клеточной теории.
- Клетка — единственная форма существования живого, является элементарной структурной и функциональной единицей живого.
- Новые клетки образуются только делением исходной клетки
- Клетка является структурно- функциональной единицей многоклеточного живого организма
Типы клеточной организации. Строение про- и эукариотических клеток. Гипотезы происхождения эукариотических клеток (симбиотическая, инвагинационная).
Среди всего многообразия ныне существующих на Земле организмов выделяют две группы: вирусы и фаги, не имеющие клеточного строения; все остальные организмы представлены разнообразными клеточными формами жизни. Различают два типа клеточной организации: прокариотический и эукариотический.
| Прокариоты | Эукариоты |
| Нет ядра | Есть ядро |
| Нет внутриклеточной мембраны | Сеть внутриклеточной мембраны |
| Кольцевая ДНК – генетический аппарат | ДНК в хромосомах |
| Нет хромосомных белков — гистонов | Есть гистоны и негистоны |
| Нет клеточного центра | Есть клеточный центр |
| Клеточная оболочка клетки – муреиновый мешок. | Клеточная стенка: у растений – из целлюлозы, у грибов- хитина, у животных ее нет. |
Эукариотические клетки произошли от предка, имевшего прокариотическое строение.
Наиболее популярна в настоящее время симбиотическая гипотеза происхождения эукариотических клеток, согласно которой основой в эволюции клетки эукариотическго типа послужил анаэробный прокариот, способный лишь к амебовидному движению. Митохондрии и жгутики произошли путем изменений проникших в клетку-хозяина симбионтов — аэробных прокариотов и бактерий. Хлоропласты клеток зеленых растений возникли из симбионтов — прокариотических клеток сине-зеленых водорослей.
Особое значение имело приобретение клеткой в процессе эволюции жгутиков с банальными тельцами, близкими к центриолям.
Трудным является вопрос о происхождении ядра. Предполагают, что оно также могло образоваться из симбионта- прокариота. Происхождение внутриплазматических мембран шероховатой и гладкой цитоплазматической сети, пластинчатого комплекса объясняют эволюционными преобразованиями наружной мембраны ядерной оболочки. Симбиотическая гипотеза не объясняет ряд фактов. Так, белок бациллин, из которого состоят реснички и жгутики современных прокариот, отличен от белка тубулина эукариот. У бактерий не обнаружено структур с типичным для жгутиков, ресничек, базальных телец или центриолей эукариотической клетки составом микротрубочек «9 + 2» или «9 + 0».
Согласно инвагинационной гипотезе, предковый формой был аэробный прокариот. Он содержал несколько геномов, первоначально прикреплявшихся к клеточной оболочке. Органеллы, имеющие ДНК, а также ядро, возникли путем впячивания и отшнуровки участков с последующей функциональной специализацией в ядро, митохондрии, хлоропласты. В процессе дальнейшей эволюции произошло усложнение ядерного генома, появилась система цитоплазматических мембран.
Эта гипотеза удовлетворительно объясняет наличие двух мембран в оболочке ядра, митохондрий и хлоропластов. Однако она не может ответить на вопрос, почему биосинтез белка в хлоропластах и митохондриях в деталях соответствуют таковому в современных прокариотических клетках, но отличается от биосинтеза белка в цитоплазме.
Симбиотическая и инвагинационная гипотезы не исчерпывают все точки зрения на происхождение эукариотического типа клеточной организации.
4.Клеточная оболочка, её структуры. Молекулярная организация и функции биологической мембраны. Виды транспорта веществ.
Клетки многоклеточных организмов, как животных, так и растительных, обособленны от своего окружения оболочкой.
Оболочки в растительных клетках состоят из клетчатки или пектина.
Клеточная оболочка, или плазмалемма, животных клеток образованна мембраной, покрытой снаружи слоем гликокаликса толщиной 10-20 нм.
Основными составляющими гликокаликса служат комплексы полисахаридов с белками (гликопротеины) и жирами (гликолипиды). Изнутри к мембране примыкает кортикальный слой цитоплазмы (0,1-0,5 мкм), в котором не встречаются рибосомы и пузырьки, но в значительном количестве находятся микротрубочки и микрофиламенты, имеющие в своем составе сократимые белки.
Плазмалемма выполняет отграничивающую, барьерную, транспортную и рецепторную функцию, регулирует химический состав внутренней среды клетки, в ней расположены молекулы рецепторов, которые избирательно распознают определенные биологически активные в-ва (гормоны).
Биологическая мембрана — тонкие пограничные структуры молекулярных размеров, расположенные на поверхности клеток и субклеточных частиц, а также канальцев и пузырьков. Особенности мембраны: она плотная, тонкая, пластичная, пронизана каналами и полярна (снаружи +, внутри -)
Мембрана состоит из бимолекулярного слоя липидов. Гидрофобные участки их молекул повернуты друг к другу, а гидрофильные – находятся на поверхности слоя.
Разнообразные белковые молекулы встроены в этот слой или размещены на его поверхности. Они выполняют ряд функций:
· Отграничивающую
· регуляции и обеспечения избирательной проницаемости веществ (транспорт ионов, сахаров, аминокислот, и других продуктов обмена веществ)
· образования поверхностей раздела между водной и неводной фазами с размещением на этих поверхностях ферментных коплексов.
Благодаря присутствию липидов (жировых в-в) мембраны образуют гидрофоюную внутриклеточную фазу как компартмент для химических реакций в неводной среде. Молекулярный состав мембран – набор соединений и ионов, размещающихся на поверхностях, различаются от структуры к структуре. Этим достигается функциональная специализация мембран клетки. Включение в мембрану клетки молекул рецепторов делает ее восприимчивой к биологически активным соединениям, например, гормонам, что способствует проявлению разности биоэлектрических потенциалов.
Виды транспорта веществ:
Пассивный транспорт — перенос веществ из области высокой концентрации в область низкой концентрации
- прямая диффузия (транспорт h3O, газов, неполярных молекул через липидный слой)
- облегченная диффузия через мембранные каналы транспорт с помощью каналообразующих белков
Активный транспорт – с затратой энергии против электрохимического градиента. Это происходит с помощью белков – переносчиков, а источник энергии молекулы АТФ
Виды активного транспорта:
- эндоцитоз (поглощение клеткой макромолекул), пинацитоз и фагоцитоз-только животные
- экзоцитоз ( из клетки выводятся различные макромолекулы)
- калий-натриевый насос.
www.ronl.ru

|
|
..:::Счетчики:::.. |
|
|
|
|
|
|
|
|