
Скачать (4,84 Mb)
Биосинтез белка – чрезвычайно сложный и энергозатратный процесс. Он является основой жизнедеятельности клетки. Синтез белка осуществляется в рибосомах и проходит в несколько этапов. Двухцепочечная молекула ДНК на основе принципа комплементарности транскрибируется в одноцепочечную молекулу РНК. В результате получается матричная РНК, которая содержит информацию об аминокислотной последовательности белка. Далее мРНК поступает в рибосому и по ней, как по матрице, синтезируется белок, путем перевода генетической информации с языка нуклеотидной последовательности на язык аминокислотной последовательности…
1. Введение2. Информационная РНК3. Генетический код4. Транспортные РНК и аминоацил-тРНК-синтетазы5. Рибосомы6. Трансляция7. Сворачивание и транспорт белков8. Заключение9. Список литературы
Жизнь есть способ существования белковых тел. Это определение, данное Фридрихом Энгельсом, указывает на исключительную роль белков в функционировании организмов. Биосинтез белка – чрезвычайно сложный и энергозатратный процесс. Он является основой жизнедеятельности клетки.Синтез белка осуществляется в рибосомах и проходит в несколько этапов по схеме ДНК→РНК→белок. Двухцепочечная молекула ДНК на основе принципа комплементарности транскрибируется в одноцепочечную молекулу РНК. В результате получается матричная РНК, которая содержит информацию об аминокислотной последовательности белка. Далее мРНК поступает в рибосому и по ней, как по матрице, синтезируется белок, путем перевода генетической информации с языка нуклеотидной последовательности на язык аминокислотной последовательности. Шаг за шагом строится полипептидная цепь, которая в процессе синтеза и после него модифицируется в биологически активный протеин. Синтезированный белок транспортируется в разные участки клетки для выполнения своих функций.Кодирование аминокислотной последовательности белков осуществляется по определенным правилам, называемых генетическим кодом. Расшифровка генетического кода – очень значимое достижение науки. Код объясняет механизм синтеза белка, происхождение мутаций и другие биологические явления.Рентгеноструктурный анализ и другие современные методы исследования позволили далеко продвинутся в изучении биосинтеза белка и других аспектов молекулярной биологии. Но тем не менее все еще не установлены пространственные структуры некоторых жизненно важных макромолекул. Науке предстоит ответить на многие вопросы, касающиеся белкового синтеза.
Общая схема биосинтеза белков в клетке: ДНК→РНК→белок (Рисунок 1).
Рисунок 1. Общая схема биосинтеза белков в клетке
Транскрипция. Отдельные участки двухцепочечной ДНК (гены) служат матрицами для синтеза на них однотяжевых цепей РНК по принципу комплементарности. Транскрипция проходит в три стадии: инициация, элонгация, терминация.
Процессинг и транспорт. В процессе синтеза РНК подвергается изменениям, в результате которых превращается в зрелую молекулу, пригодную для синтеза белка. Получающаяся информационная (матричная) РНК (мРНК) затем поступает к рибосомам в качестве программы, определяющей аминокислотную последовательность в синтезируемом белке.
Активация и акцептирование аминокислот. Белки состоят из аминокислот, но свободные аминокислоты клетки не могут быть непосредственно использованы рибосомой. Каждая аминокислота сначала активируется с помощью АТФ, а затем присоединяется к специальной молекуле РНК – трансферной (транспортной) РНК (тРНК) вне рибосомы. Получающаяся аминоацил-тРНК поступает в рибосому в качестве субстрата для синтеза белка.
Трансляция. Поток информации в виде мРНК и поток материала в виде аминоацил-тРНК поступают в рибосомы, которые осуществляют перевод (трансляцию) генетической информации с языка нуклеотидной последовательности мРНК на язык аминокислотной. Каждая рибосома движется вдоль мРНК от одного конца к другому и соответственно выбирает из среды те аминоацил-тРНК, которые соответствуют (комплементарны) триплетным комбинациям нуклеотидов, находящимся в данный момент в рибосоме. Аминокислотный остаток выбранной аминоацил-тРНК каждый раз ковалентно присоединяется рибосомой к растущей полипептидной цепи, а деацилированная тРНК освобождается из рибосомы в раствор. Так последовательно строится полипептидная цепь.
Формирование функционального белка. По ходу синтеза полипептидная цепь высвобождается из рибосомы и сворачиваться в глобулу. Сворачивание и транспорт белка сопровождаются ферментативными модификациями (процессинг белка).
Несмотря на большую сложность аппарата биосинтеза белков, он протекает с чрезвычайно высокой скоростью. Синтез тысяч различных белков в каждой клетке строго упорядочен – при данных условиях метаболизма синтезируется лишь необходимое число молекул каждого белка.
Информационная (матричная) РНК (мРНК) – РНК, являющаяся комплементарной копией участков значащих цепей генов ДНК, содержащих информацию об аминокислотных последовательностях полипептидных цепей белков.
Первичная структура
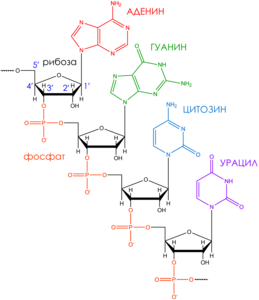
Рисунок 2. Химическое строение полинуклеотида РНК
Матричная РНК — одноцепочечный полинуклеотид (Рисунок 2). Он состоит из четырех нуклеотидов. Нуклеотид ы состоят из азотистого основания (аденин – А, гуанин – G, цитозин – C и урацил – U), сахара рибозы и фосфатной группы. 5'-гидроксил концевого нуклеозида (молекула, содержащая азотистое основание, связанное с сахаром) не образует связи между нуклеотидами. Он обозначается как 5'-конец РНК, а другой концевой нуклеозид со свободным З'-гидроксилом называют З'-концом РНК. мРНК читается рибосомой в направлении от 5'-конца к З'-концу .В природных мРНК 5'-концевой гидроксил всегда замещен. мРНК эукариотов в большинстве случаев несут на 5'-конце специальную группу – кэп (Рисунок 3). Кэп представляет собой остаток 7-метилгуанозина (Рисунок 4).

Рисунок 3. Строение 5'-конца кэпированной мРНК
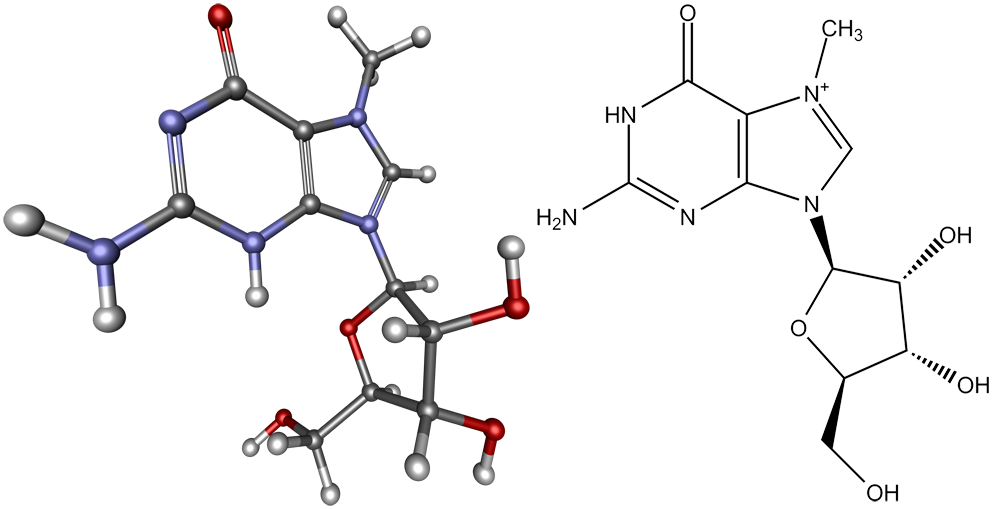
Рисунок 4. Модель молекулы 7-метилгуанозина
Функциональные участки мРНК
Чаще всего началом (инициаторным кодоном) кодирующей части мРНК яв¬ляется AUG. Не любой триплет может стать инициаторным. Это определяется собственной структурой кодона и положением в структуре мРНК.мРНК может содержать нуклеотидные последовательности для кодирования нескольких белков. Это характерно для прокариот. Такие мРНК называются полицистронными. У эукариот мРНК обычно кодируют одну полипептидную цепь (моноцистронные мРНК).
Пространственная структура
Рисунок 5. Вторичная структура РНК
Трехмерная структура мРНК еще не установлена. Измерения физических параметров мРНК свидетельствуют о том, что они являются сильно свернутыми структурами, с внутрицепными взаимодействиями между азотистыми основаниями. Вторичная структура мРНК образована благодаря комплементарному спариванию отдельных участков одной и той же цепи друг с другом, с образованием большого набора относительно коротких двуспиральных участков (Рисунок 5).Вторичная и третичная структуры мРНК играют определенную роль в трансляции. Однако роль вторичной и третичной структуры мРНК в скорости считывания цепи не установлена.Некодирующие последовательности мРНК участвуют в определении специальных пространственных структур, ответственных за регулирование инициации трансляции, элонгации и других процессов.
Так как существует только 4 нуклеотида в мРНК и 20 аминокислот в белке, то трансляция не может быть осуществляется на основе прямого соотношения между нуклеотидами РНК и аминокислотами в белке. Нуклеотидная последовательность гена через посредничество мРНК транслируется в аминокислотную последовательность по правилам, известным как генетический код.Генетический код – способ сохранения наследственной информации в виде последовательности нуклеотидов в молекулах нуклеиновых кислот. Этот код был расшифрован в 1960-ых. Генетический код, основан на использовании алфавита, состоящего из четырех букв: А, Г, Ц и Т. Эти буквы соответствуют нуклеотидам, найденным в ДНК: аденин, гуанин, цитозин, тимин.Последовательность нуклеотидов в молекуле мРНК читается непрерывными группами из трех нуклеотидов, называемых триплетами или кодонами. РНК представляет собой линейные полимер, состоящий из четырех разных нуклеотидов, поэтому возможны 4•4•4=64 комбинации трех нуклеотидов. Белки состоят из 20 аминокислот. Поэтому либо некоторые триплеты не используются, либо некоторые аминокислоты кодируются более, чем одним триплетом.Различают два типа кодонов— смысловые, или значащие кодоны, и бессмысленные кодоны, или нонсенс-кодоны. Большинство (61) кодонов — значащие и только 3 (UAA, UAG, UGA) – нонсенс-кодоны. Смысловые кодоны соответствуют аминокислотам, а кодон AUG, помимо кодирования митионина, является инициирующим, или стартовым кодоном. Нонсенс-кодоны являются терминирующими кодонами, или стоп-кодонами.
Генетический код является неперекрываемым, непрерывным, специфичным, универсальным и вырожденным.
Неперекрываемость кода означает, что каждый нуклеотид входит только в один кодон, и поэтому изменения любого нуклеотида изменяют смысл только одного кодона.
Генетический код непрерывен. Он имеет линейный непрерывающийся порядок считывания. Кодоны транслируются всегда целиком. Расположение остатков аминокислот в синтезируемом полипептиде определяется антикодоном тРНК (триплет нуклеотидов, комплементарный одному из кодонов) .
Специфичность кода означает, что код является однозначным, поскольку каждый кодонный триплет кодирует только одну аминокислоту, и с одной мРНК можно синтезировать только одинаковые пептиды,
Генетический код универсален для всех живых существ – у всех живых организмов, включая вирусы и бактерии, одинаковые кодоны (триплеты нуклеотидов) кодируют одинаковые аминокислоты. Исключение составляют 4 кодона митохондрий грибов и животных, имеющих информационный смысл, отличный от универсального кода.
Вырожденность кода означает его избыточность, синонимичность, то есть одну аминокислоту может кодировать более одного триплета. Однако вырожденность не абсолютна. Например, метионину соответствует только один кодон.
До расшифровки генетического кода было невозможно понять механизм синтеза белка и объяснить происхождение мутаций. Открытие генетического кода позволило ответить на вопрос о том, как связаны между собой дефекты определенных белков человека и наследственные заболевания.
Генетический код
| 1-ая позиция (5’ конец) | 2-ая позиция | 3-ая позиция (3’ конец) | |||
| ↓ | U | C | A | G | ↓ |
| U | PhePheLeuLeu | SerSerSerSer | TyrTyrSTOPSTOP | CysCysSTOPTrp | UCAG |
| C | LeuLeuLeuLeu | ProProProPro | HisHisGlnGln | ArgArgArgArg | UCAG |
| A | llellelleMet | ThrThrThrThr | AsnAsnLysLys | SerSerArgArg | UCAG |
| G | ValValValVal | AlaAlaAlaAla | AspAspGluGlu | GlyGlyGlyGly | UCAG |
| Аминокислоты и их символы | Кодоны | |||
| ACDEFGHIKLMNPARSTVWY | AlaCysAspGluPheGlyHisIleLysLeuMetAsnProGlnArgSerThrValTrpTyr | AlanineCysteineAspartic acidGlutamic acidPhenylalaniGlycineHistidineIsoleucineLysineLeucineMethionineAsparagineProlineGlutamineArginineSerineThreonineValineTryptophanTyrosine | АланинЦистеинАспарагиновая кислотаГлутаминовая кислотаФенилаланинГлицинГистидинИзолейцинЛизинЛейцинМетионинАспарагинПролинГлутаминАргининСеринТреонинВалинТриптофанТирозин | GCA GCC GCG GCUUGC UGUGAC GAUGAA GAGUUC UUUGGA GGC GGG GGUCAC CAUAUA AUC AUUAAA AAGUUA UUG CUA CUC CUG CUUAUGAAC AAUCCA CCC CCC CCUCAA CAGAGA AGG CGA CGC CGG CGUAGC AGU UCA UCC UCG UCUACA ACC ACG ACUGUA GUC GUG GUUUGGUAC UAU |
Транспортные РНК (тРНК) небольшие по размеру молекулы (73- 97 нуклеотидных остатков в цепи). Все тРНК имеют одинаковый 3'-конец, построенный из двух остатков цитозина и одного аденозина (CCA-конец). В середине цепи тРНК находится антикодон. В молекулах тРНК присутствет множество разнообразных модифицированных нуклеозидов (минорные нуклеозиды), образующиеся путем ферментативной модификации обычных нуклеозидных остатков.
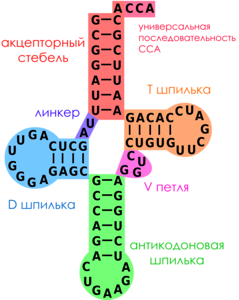
Рисунок 6. Вторичная структура тРНК
Вторичная структура тРНК складывается за счет взаимокомплементарности участков цепи. Они формируют структуру «клеверного листа», состоящую из четырех стеблей и трех петель. Стебель с петлей формируют ветвь. В дополнение к трем петлям клеверного листа в структуре тРНК выделяют также дополнительную, или вариабельную, петлю (V петлю). Двухцепочечные стебли с постоянным числом спаренных нуклеотидов представляют собой двойную спираль.

Рисунок 7. Третичная структура тРНК
Третичная структура формируется за счет взаимодействия элементов вторичной структуры. Пространственная структура тРНК называется L-формой (из-за сходства с латинской буквой L).Третичные взаимодействия (стекинг оснований и другие) скрепляют разные участки L-структуры в непрерывные двойные спирали.Молекулам тРНК присущи индивидуальные различия, проявляющиеся на уровне вторичной и третичной структур, например, разная величина угла между доменами L-структуры.
Две основные функции тРНК:
Аминоацилирование тРНК – процесс активации аминокислот. Он происходит на первом этапе биосинтеза белка – двадцать различных аминокислот присоединяются эфирной связью к соответствующим тРНК под действием двадцати различных активирующих ферментов, называемыми аминоацил-тРНК-синтетазами. Каждый фермент специфичен по отношению к определенной аминокислоте и к соответствующей тРНК.

Рисунок 8. Обобщенная структура аминоацил-тРНК
Аминоацилирование состоит из двух стадий, проходящих в каталитическом центре фермента. На первой стадии в результате взаимодействия АТР и аминокислоты образуется промежуточное соединение – аминоациладенилат. На второй стадии аминоацильный остаток переносится с аминоациладенилата, связанного с ферментом, на соответствующую специфическую тРНК (Рисунок 8).
Аминоацилирование может быть выражено схемой:
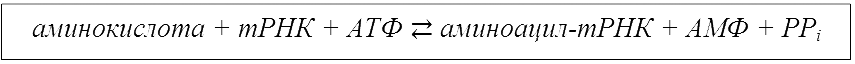
АТФ – аденозинтрифосфат, АМФ – аденозинмонофосфат, PPi – пирофосфаты.Исключительно низкая частота ошибок при аминоацилировании тРНК является непременным условием реализации генетического кода – если на предрибосомном этапе произошла ошибка и к тРНК присоединилась аминокислота, не соответствующая специфичности антикодона, то эта ошибка уже не может быть исправлена на последующих этапах белкового синтеза.В ходе эволюции выработались специфические механизмы отбора «правильных» субстратов для аминоацил-тРНК-синтетаз, обеспечивающие безошибочное аминоацилирование тРНК.
Узнавание тРНК аминоацил-тРНК-синтетазами
Каждая тРНК, сохраняя универсальную L-образную форму, имеет отличительные признаки, безошибочно распознаваемые «своим» ферментом как «притягательные», а остальными 19 ферментами – как «отталкивающие».Это следующие участки тРНК (Рисунок 9):
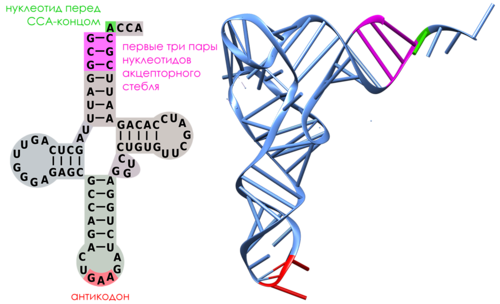
Рисунок 9. Участки, по которым происходит узнавание тРНК аминоацил-тРНК-синтетазами
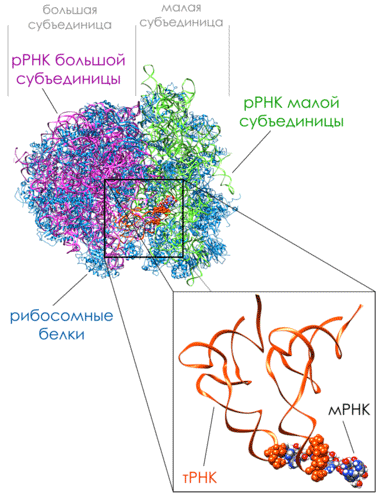
Рисунок 10. Комплекс 80S рибосома-мРНК-тРНК клетки дрожжей
Синтез белка происходит в рибосоме. Рибосома – сложный макромолекулярный аппарат, состоящий из более 50 белков, называемых рибосомными белками, и нескольких молекул РНК, называемых рибосомными РНК. Число рибосом в клетке различно. Оно зависит от интенсивности белкового синтеза в данном типе клеток. Обычная эукариотческая клетка содержит миллионы рибосом. Эукариотические и прокариотические рибосомы схожи в строении и функциях и различаются лишь числом и размером рРНК и рибосомных протеинов. Строение рибосомы приведено на рисунке 10 на примере рибосомы дрожжей (Рисунок 10).Рибосомы имеют размер 25-30 нм. Они состоят из двух неравных субъединиц. Субъединицы эукариотических рибосом формируются в ядре из рРНК, ассоциированных с рибосомными белками, которые транспортируются в ядро после синтеза в цитоплазме. Две субъединицы рибосомы затем выходят в цитоплазму, где соединяются воедино для участия в синтезе белка.Рибосомные рибонуклеиновые кислоты (рРНК) – основные компоненты рибосом, составляют большую часть их массы. Молекулы рРНК определяют структуру, физические и химические свойства, функции рибосом, а также расположение рибосомных белков в субчастицах рибосом.Малые субчастицы рибосом содержат одну молекулу рРНК, большие – две.Молекулы рРНК являются совокупностью коротких одноцепочечных и двухспиральных участков, образующихся за счет комплементарного спаривания участков одной и той же полинуклеотидной цепи.В субъединицах рибосом рРНК компактно упакованы благодаря ионам двухвалентных металлов и рибосомным белкам. Основная часть рРНК располагается внутри рибосомных субчастиц. Отдельные участки рРНК находятся на поверхности субчастиц. Они выполняют важную биололическую роль, формируя функциональные центры рибосом (центры связывания матричных и транспортных РНК и белковых факторов трансляции).Рибосомная РНК концентрируется в основном ближе к центру частиц, тогда как масса рибосомных белков занимает в среднем более периферическое положение. Можно сделать вывод, что свернутая молекула высокополимерной рибосомной РНК – это структурное ядро рибосомной субчастицы, определяющее и ее компактность, и ее форму, и организацию на ней рибосомных белков. То есть рибосома есть прежде всего ее РНК (Рисунок 11).
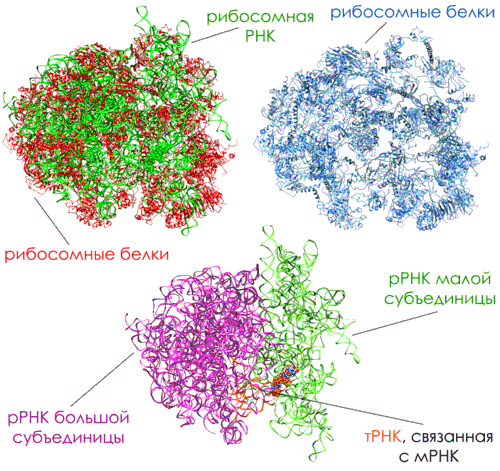
Рисунок 11. Рибосомные белки и рибосомная РНК
Многочисленные рибосомные белки могут участвовать в функциях связывания субстратов и каталитических функциях рибосомы, локализуясь в соответствующих функциональных центрах и обеспечивая их своими активными группами; рибосомные белки могут служить стабилизаторами или модификаторами определенных локальных структур рибосомной РНК и таким образом поддерживать их в функционально активном состоянии или способствовать их переключениям из одного состояния в другое.Когда рибосома не участвует в синтезе белков, две субъединицы разделены. Они соединяются с мРНК (обычно возле 5'-конца) для инициации синтеза белков. Затем мРНК продвигается через рибосому, и по мере вхождения кодонов в ядро рибосомы, нуклеотидная последовательность мРНК транслируется в аминокислотную поледовательность с помощью тРНК в качестве адаптора, чтобы приоединять каждую аминокислоту в правильном порядке к концу растущего белка. Когда считывается терминаторный кодон, из рибосомы выходит синтезированный белок, и две субъединицы снова разделяются. Эти субъединицы могут снова быть использованы для синтеза другого белка по другой молекуле мРНК.Обычно одна молекула мРНК читается сразу несколькими рибосомами, двигающимися вдоль мРНК друг за другом и, таким образом, независимо синтезирующими идентичные молекулы белка, но с соответствующим отставанием. Такой динамический комплекс одной мРНК с несколькими рибосомами называется полирибосомой.

Рисунок 12. Функциональные участки рибосомы
Рибосомы работают чрезвычайно продуктивно: в секунду одна рибосома эукариотической клетки присоединяет 2 аминокислоты к полипептидной цепи, рибосомы бактериальных клеток функционируют еще быстрее — около 20 аминокислот в секунду. Такая продуктивность объясняется наличием четырех функциональных участков (сайтов) для РНК молекул (Рисунок 12) – один для мРНК и три для тРНК:
Молекула тРНК крепится к А- и Р-сайтам только если её антикодон образует пары оснований с комплементарным кодоном молекулы мРНК. А- и Р-сайты расположены близко друг к другу чтобы две их тРНК молекулы формировали пары нуклеотидов с примыкающим кодоном молекулы мРНК. Эта особенность рибосомы обеспечивает правильное считывание мРНК.Рибосомы разных клеток различаются по размерам, которые определяются по скорости осаждения при центрифугировании. Скорость осаждения измеряется в единицах Сведберга (S). Сведберг – это отношение скорости седиментации к центробежному ускорению, 1S = 10-13 секунд. Коэффициенты седиментации разных рибосом варьируют от 5S до 80S. У прокариот рибосомы имеют коэффициент 70S, а рибосомы цитоплазмы эукариот — 80S.Диссоциация рибосом в случае прокариот и эукариот соответственно:70S→50S + 30S80S→60S + 40S
Трансляция – непосредственный процесс синтеза белка рибосомой. Трансляция проходит в три стадии: инициация, элонгация, терминация.
Трансляция у бактерий и в эукариотических клетках в общем схожи, но различаются механизмом инициации. Для инициация белкового синтеза в бактериях необходимы 30S и 50S рибосомные единицы, мРНК, молекула тРНК, аминоацилированная N-формилметионином – fMet-tRNAfMet, три белка, называемые факторами инициации, гуанозинтрифосфат (ГТФ), Mg2+. В процессе работы рибосома потребляет энергию гидролиза гуанозинтрифосфата (Рисунок 13).

Рисунок 13. Структура гуанозинтрифосфата
Комплекс 30S субъединицы с факторами инициации распознает участки связывания рибосомы (сайты), которые содержат инициаторный кодон AUG, и специальную последовательность Шайна- Дальгарно, служащую для отличия AUG от внутренних кодонов, кодирующих метионин. В результате инициации образуется 70S рибосома – инициирующий комплекс, содержащий мРНК и fMet-tRNAfMet, связанную с P-участком рибосомы. Комплекс готов к элонгации.В эукариотических клетках имеется как минимум девять факторов инициации. Инициирующий кодон AUG распознается не последовательностью Шайна-Дальгарно, а сканированием мРНК с 5'-конца до первого AUG и соответствующим расположением рамки считывания.
Стадия элонгации требует комплекс инициации, аминоацил-тРНК, факторы элонгации (растворимые цитозольные белки) и гуанозинтрифосфат (ГТФ).Синтез происходит на рибосоме путем последовательного добавления одного аминокислотного остатка за другим к строящейся полипептидной цепи; таким путем осуществляется элонгация (удлинение) пептида. Каждый новый аминокислотный остаток добавляется к карбоксильному концу (С-концу) пептида, т. е. С-конец пептида является растущим. Добавление одного аминокислотного остатка соответствует прочтению одного нуклеотидного триплета.Элонгация проходит в три этапа, которые повторяются пока есть остатки аминокислот для присоединения.На первом шаге аминоацил-тРНК молекула, нагруженная аминокислотой крепится к А-сайту рибосомы, а "отработавшая" тРНК высвобождается с Е-сайта (от exit — выход).На втором шаге формируется новая пептидная связь под действием фермента пептидилтрансферазы.На третьем шаге происходит транслокация: большая субъединица занимает позицию относительно малой субъединицы, оставляя две тРНК в гибридных сайтах: в Р-сайте на большой субъединице и А-сайте на малой для одной тРНК и в E-сайте на большой субъединице и Р-сайте на малой для другой. Затем малая субъединица перемещается вместе с мРНК на три нуклеотида, освобождая А-сайт для следующей тРНК и цикл повторяется снова. Молекула мРНК транслируется с 5'-конца к 3'-третьему, а синтез протеина начинается с N-конца. С началом каждого цикла аминокислота крепится к C-концу полипептидной цепи.
Элонгация продолжается до тех пор пока рибосома не присоединит последнюю аминокислоту. Терминация начинается при присутствии одного из трех терминаторных кодонов в мРНК (UAA, UAG, UGA), которые следуют сразу за последней аминокислотой. Когда рибосома достигнет терминирующего кодона мРНК, синтез полипептида прекращается. В бактериях, если терминаторный кодон имеет место быть в А-сайте рибосомы, в дело вступают три фактора терминации, которые участвуют в гидролизе пептидил-тРНК связи; освобождении полипептида и последней, уже ненагруженной тРНК из Р-сайта; диссоциации 70S рибосомы на 30S и 50S субъединицы, готовых начать новый цикл синтеза белка.Таким образом, каждая рибосома проходит полный цикл трансляции, включающий инициацию, элонгацию и терминацию; в результате такого эпицикла прочитывается вся кодирующая последовательность мРНК и синтезируется законченная полипептидная цепь белка. После этого рибосома может повторить цикл с другой цепью мРНК или другой кодирующей последовательностью той же цепи.
Процесс экспрессии генов не заканчивается на построении аминокислотной последовательности, составляющей протеин, с помощью генетического кода. Чтобы быть полезным клетке, новый пептид должен подвергнутся процессингу: свернутся в трехмерную нативную конформацию, присоединить какие-либо молекулы, необходимые для его активности, модифицироваться под действием протеинкиназ и других энзимов и правильно соединиться с другими частями белка, с которыми он функционирует.Информация, нужная для всех этих процессов содержится в последовательности связанных аминокислот, которые производит рибосома, когда транслирует мРНК в полипептидную цепь. Когда протеин сворачивается в компактную структуру, гидрофобные звенья обращаются внутрь глобулы. Формируется большая часть нековалентных взаимодействий между различными участками молекул. Итогом всех этих энергетически выгодных взаимодействий, определяющих свернутую структуру полипептидной цепи является конформация с самой низкой энергией.За миллионы лет эволюции аминокислотная последовательность каждого протеина была выбрана определенным образом не только для конформации, которую он принимает, но также для быстрого сворачивания. Для некоторых белков сворачивание начинается с N-конца сразу после выхода полипептида из рибосомы. В этом случае, как только протеин покидает рибосому, через несколько секунд он формирует компактное строение, содержащее окончательную вторичную структуру (спирали и β-листы).Большинство белков не сворачиваются во время синтеза. Вместо этого они "встречаются" у рибосомы с отдельным классом белков, называемых шаперонами. Связывание с шаперонами обеспечивает правильное сворачивание белка в нативную конформацию.Основные этапы процессинга:
Эукариотические клетки состоят из множества структур, органелл и отсеков со специфичными функциями, которые требуют определенный набор белков. Эти белки (за исключением тех, которые синтезируются в митохондрих и пластидах) производятся рибосомами в цитозоле.Белки предназначенные для секреции, интеграции в плазматическую мембрану, включения в лизосомы, обычно проходят первые несколько стадий внутриклеточной сортировки. Белки для митохондрий, пластид и ядра используют при отдельных механизм. Белки, предназначенные для цитозоля, остаются там, где были синтезированы.Важнейшим элементом в процессе сортировки белков является короткая последовательность аминокислот – сигнальная последовательность белка (лидер). Она направляет белок на предназначенное ему место и удаляется во время транспорта или по прибытию белка в конечный пункт.В процессе синтеза любого белка, в том числе предназначенного на «экспорт», сигнальные лидеры, будучи расположены на N-конце, образуются первыми. Такие лидеры узнаются особыми рецепторными участками на внешней поверхности эндоплазматического ретикулума, причем это происходит даже раньше, чем рибосома полностью завершит синтез белка. Гидрофобная жирорастворимая часть лидирующей последовательности проникает сквозь мембрану внутрь цистерн эндоплазматического ретикулума, протаскивая за собой растущую полипептидную цепь. Внутри цистерн под действием особой пептидазы сигнальный лидер отщепляется. После этого зрелый белок направляется в аппарат Гольджи, инкапсулируется и в виде секреторного пузырька покидает наконец клетку.
Белки являются структурными блоками клетки, а также осуществляют большую часть её функций. Биосинтез белка является одним из основополагающих процессов клетки.Расшифровка генетического кода, правил, по которым нуклеотидная последовательность РНК переводится в аминокислотную последовательность полипептида, позволила понять принцип биосинтеза белка и связанные с этих явления.Синтез протеина начинается с построения цепи матричной РНК из ДНК по принципу комплементарности.После определенных преобразований молекула матричной РНК становится матрицей для построения по ней белка.Биосинтез белка происходит в рибосомах, которые состоят из белков и рРНК. Рибосомы диссоциируют на две неравные субъединицы.
Транспортная РНК состоит из небольшого числа нуклеотидов (73-97), некоторые из которых могут быть модифицированы. тРНК имеют общее строение:
Транспортные РНК выполняют акцепторную и адапторную функции.Рост полипептида в рибосоме начинается с N-конца добавлением новых аминокислотных остатков и завершается на C-конце.Для синтез белка необходима активация аминокислот. Аминокислоты активируются в цитозоле аминоацил-тРНК синтетазой. Эти ферменты катализируют образование аминоацил-тРНК и расщепление АТФ на АМФ и пирофосфаты (PPi). Точность белкового синтеза зависит от правильного протекания этой реакции.Непосредственный синтез белка (трансляция) проходит в три стадии.
Полипептид сворачивается в биологически активную конформацию и транспортируется к "рабочему месту".Несмотря на множество открытий в биохимии и значимую работу, проделанную разными учеными в области этой науки, некоторые аспекты синтеза белков еще не установлены. Например, зачем нужны молекулы рРНК и как случилось, что они приобрели главенствующую роль в структуре и функции рибосом? На этот и другие вопросы науке предстоит ответить.
Структуры молекул взяты из RCSB Protein Data Bank и ChemSpider
Извините, ничего не найдено.
studentoriy.ru
В отличие от фотосинтеза биосинтез белка происходит иначе. В биосинтезе белка активное участие принимают нуклеиновые кислоты — ДНК и РНК, а в качестве необходимой энергии используется энергия химических связей высокомолекулярных органических соединений, имеющихся в клетке, главным образом — АТФ.
Последовательность нуклеотидов ДНК (т. е. генов), или генетический код, представляет собой систему записи информации о последовательности расположения аминокислот в белках и фактически является шифром, обеспечивающим биосинтез белка.
Генетическая информация в соответствии с генетическим кодом в какой-то момент переписывается с ДНК, как с матрицы, в нуклеотидную последовательность нити информационной РНК (иРНК). Она и определяет затем последовательность сборки аминокислот соответствующей белковой молекулы.
Важно отметить, что генетический код является универсальным для всех организмов, существующих на Земле. Это свойство универсальности кода позволяет сделать важный мировоззренческий вывод о единстве происхождения всех живых организмов — прокариот, эукариот и вирусов.
В настоящее время расшифрованы триплеты для всех 20 аминокислот, входящих 8 состав природных белков. Генетический код был расшифрован в 60-е гг. XX в. Это осуществили учёные-биохимики X. Корана, М. Ниренберг и Р. Холли. За расшифровку генетического кода и его роли в синтезе белка названным учёным в 1968 году была присуждена Нобелевская премия.
В биосинтезе активное участие принимают многие структурные компоненты клетки: различные молекулы РНК, рибосомы и молекулы разных аминокислот, из которых строится полимерная молекула белка. Хотя план строения белка закодирован в ДНК, сама она участия в синтезе белковых молекул не принимает, а служит лишь матрицей для синтеза информационной РНК (иРНК). Поэтому процесс синтеза белка складывается из двух этапов: создание иРНК и сборка молекулы белка по информации в этой молекуле иРНК.
Синтез белковых молекул происходит непрерывно. Он идёт с большой скоростью: в 1 минуту образуется от 50 до 60 тысяч пептидных связей. Синтез одной молекулы длится обычно 3-4 сек. Длительность жизни белков в среднем составляет около двух суток, хотя отдельные белки не разрушаются в течение нескольких месяцев. В результате половина белков тела человека (всего это около 17 кг белка) обновляется примерно за 80 дней. Материал с сайта http://doklad-referat.ru
Процесс биосинтеза на всех его этапах идёт с участием многих ферментов и с непременным потреблением большого количества энергии.
Чёткая последовательность происходящих процессов, их матричная организованность и распределение функций между всеми задействованными компонентами приводят к выводу, что биосинтез белка — это целостная молекулярная система выполнения сложных реакций, обеспечивающая создание веществ, необходимых для жизни.
Биосинтез белка — пластическая часть обмена веществ клетки. Характеризуется матричной основой сборки молекул белка. Синтез происходит в рибосомах при непосредственном участии иРНК, тРНК, рРНК и мономеров — аминокислот. В отличие от фотосинтеза биосинтез белка идёт под строгим контролем генетической информации, списанной иРНК с генетического кода ДНК. Процесс биосинтеза белковой молекулы обусловлен двумя этапами: транскрипцией (списание) и трансляцией (передача).
 На этой странице материал по темам:
На этой странице материал по темам:  Вопросы по этому материалу:
Вопросы по этому материалу: doklad-referat.ru
Министерство образования и науки Российской Федерации
Марийский государственный технический университет
Реферат на тему: «Биосинтез белка»
Выполнила: студентка гр. Ларх-11
Попова М. Ю.
Проверил: Карасев В.Н.
Йошкар-Ола
2012
Содержание
Любая живая клетка способна синтезировать белки, и эта способность представляет одно из наиболее важных и характерных ее свойств. С особенной энергией идет биосинтез белков в период роста и развития клеток. В это время активно синтезируются белки для построения клеточных органоидов, мембран. Синтезируются ферменты и белки. Биосинтез белков идет интенсивно и у многих взрослых, т. е. закончивших рост и развитие клеток, например у клеток пищеварительных желез, синтезирующих белки-ферменты (пепсин, трипсин) или у клеток желез с внутренней секрецией, синтезирующих белки-гормоны (инсулин, тироксин). Способность к синтезу белков присуща, впрочем, не только растущим или секреторным клеткам: любая клетка в течение всей жизни постоянно синтезирует белки, так как в ходе нормальной жизнедеятельности молекулы белков постепенно изнашиваются, структура и функции их нарушаются. Такие пришедшие в негодность молекулы белков удаляются из клетки. Взамен синтезируются новые полноценные молекулы, состав и деятельность клетки не нарушаются.
Любая клетка по внешнему виду и по свойствам похожа на материнскую. Так как свойства клетки зависят от ее белков, то ясно, что клетка способна синтезировать белки такие же, какие синтезировала материнская клетка. Следовательно, способность к синтезу белка передается по наследству от клетки к клетке и сохраняется ею в течение всей жизни. Вопросы о том, как происходит синтез столь большой и сложной молекулы белка, как отбираются нужные аминокислоты, расставляются и соединяются в определенном и строгом порядке, еще сравнительно недавно представляли неразрешимую загадку. Эти вопросы в настоящее время в основном выяснены, и решение их представляет величайшее достижение биологии и биохимии XX в.
Основная роль в определении структуры белка принадлежит ДНК. Мы уже знаем, что молекулы ДНК очень велики. Их длина в десятки и сотни раз превышает длину самых крупных молекул белков: по длине цепочки ДНК можно было бы уложить одну за другой десятки, а то и сотни молекул белков. В настоящее время установлено, что разные участки ДНК определяют синтез различных белков. Одна молекула ДНК участвует в синтезе нескольких десятков белков.
Каждый участок ДНК, определяющий синтез одной молекулы белка, называется геном. Каждый ген — участок двойной спирали ДНК, на котором содержится информация о структуре какого-то одного белка.
Чтобы разобраться в том, каким образом структура ДНК определяет структуру белка, приведем такой пример. Многие знают об азбуке Морзе, при помощи которой передают сигналы и телеграммы. По азбуке Морзе все буквы алфавита обозначены сочетаниями коротких и длинных сигналов — точками и тире. Буква А обозначается •—, Б - -и т. д. Собрание условных обозначений называется кодом или шифром. Азбука Морзе представляет собой пример кода. Получив телеграфную ленту с точками и тире, знающий код Морзе легко расшифрует (раскодирует) написанное.
Макромолекула ДНК, состоящая из нескольких тысяч последовательно расположенных четырех видов нуклеотидов, представляет собой код, определяющий структуру белка. Так же как в коде Морзе каждой букве соответствует определенное сочетание точек и тире, так в коде ДНК каждой аминокислоте соответствует определенное сочетание последовательно связанных нуклеотидов.
Код ДНК
Код ДНК удалось расшифровать почти полностью. Сущность кода ДНК состоит в следующем. Каждой аминокислоте соответствует участок, цепи ДНК из трех рядом стоящих нуклеотидов. Например, участок Т—Т—Т соответствует аминокислоте лизину, отрезок А—Ц—А—цистеину, Ц—А—А — валину и т. д.
Допустим, что в гене нуклеотиды следуют в таком порядке:
А—Ц—А—Т—Т—Т—А—А—Ц—Ц—А—А— Г— Г.
Разбив этот ряд на тройки (триплеты), мы сразу же расшифруем, какие аминокислоты и в каком порядке следуют в молекуле белка:
А—Ц—А Т—Т—Т А—А—Ц Ц—А—А Г—Г—Г
Цистеиц Лизин Лейцин Валин Пролин
В коде Морзе всего два знака. Для обозначения всех букв, всех цифр и знаков препинания приходится брать на некоторые буквы или цифры до 5 знаков. В коде ДНК проще. Разных нуклеотидов 4. Число возможных комбинаций из 4 элементов по 3 равно 64. Разных же аминокислот всего 20. Таким образом, различных триплетов нуклеотидов с избытком хватает для кодирования всех аминокислот.
Транскрипция.
Установлено, что сами ДНК непосредственного участия в синтезе белка не принимают, ДНК находятся в ядре клетки, а синтез белка происходит в рибосомах — мельчайших структурах, находящихся в цитоплазме. В ДНК только содержится и хранится информация о структуре белков. Для синтеза белка в рибосомы направляются точные копии этой информации. Это осуществляется с помощью РНК, которые синтезируются на ДНК и точно копируют ее структуру. Последовательность нуклеотидов РНК точно повторяет последовательность нуклеотидов в одной из цепей гена. Таким образом, информация, содержащаяся в структуре данного гена, как бы переписывается на РНК. Этот процесс называется транскрипцией («транскрипциа» — переписывание, лат.). С каждого гена можно снять любое число копий РНК. Эти РНК, несущие в рибосомы информацию о составе белков, называются информационными (и - РНК).
Для того, чтобы понять, каким образом состав и последовательность расположения нуклеотидов в гене могут быть «переписаны» на РНК, вспомним принцип комплементарности, на основании которого построена двуспиральная молекула ДНК. Нуклеотиды одной цепи обусловливают характер противолежащих нуклеотидов другой цепи. Если на одной цепи находится А, то на том же уровне другой цепи стоит Т, а против Г всегда находится Ц. Других комбинаций не бывает. Принцип комплементарности действует и при синтезе информационной РНК. Против каждого нуклеотида одной из цепей ДНК встает комплементарный к нему нуклеотид информационной РНК. Таким образом, против Гднк встает Црнк против Цднк — Грнк, против Аднк — Урнк, против Тднк — Арнк. В результате образующаяся цепочка РНК по составу и последовательности своих нуклеотидов представляет собой точную копию состава и последовательности нуклеотидов одной из цепей ДНК. Молекулы информационной РНК направляются к месту, где происходит синтез белка, т. е. к рибосомам. Туда же идет из цитоплазмы поток материала, из которого строится белок, т. е. аминокислоты. В цитоплазме клеток всегда имеются аминокислоты, образующиеся в результате расщепления белков пищи.
Транспортные РНК
Аминокислоты попадают в рибосому не самостоятельно, а в сопровождении особых молекул РНК, специально приспособленных для транспорта аминокислот к рибосомам. Они так и называются: транспортные РНК (т - РНК). Транспортные РНК — это сравнительно короткие цепочки, состоящие всего из нескольких десятков нуклеотидов. На одном конце их молекулы имеется структура, к которой может присоединиться аминокислота. На другом конце транспортной РНК находится триплет нуклеотидов, который соответствует по коду данной аминокислоте. Например, молекула транспортной РНК для аминокислоты лизина на одном конце имеет «площадку» для «посадки» лизина, а на другом конце — триплет нуклеотидов: У—У—У. Так как существует не менее 20 различных аминокислот, то, очевидно, существует не менее 20 различных транспортных РНК. На каждую аминокислоту имеется своя транспортная РНК.
Реакция матричного синтеза. Для изучавшего неорганическую и органическую химию привычны, реакции, протекающие в растворах, в которых молекулы веществ находятся в хаотическом движении. Реакции в таких системах осуществляются в результате случайного столкновения молекул. Чем концентрация веществ выше, тем больше вероятность столкновения, тем выше скорость реакции. Напротив, при понижении концентрации веществ вероятность встречи молекул невелика и скорость реакции может быть ничтожной.
В живых системах мы встречаемся с новым типом реакций, наподобие описанной выше реакции редупликации ДНК или реакции синтеза РНК. Такие реакции неизвестны в неживой природе. Они называются реакциями матричного синтеза.
Термином «матрица» в технике обозначают форму, употребляемую для отливки монет, медалей, типографского шрифта: затвердевший металл в точности воспроизводит все детали формы, служившей для отливки. Матричный синтез напоминает отливку на матрице: новые молекулы синтезируются в точном соответствии с планом, заложенным в структуре уже существующих молекул. Матричный принцип лежит в основе важнейших синтетических реакций клетки, таких, как синтез нуклеиновых кислот и белков. В этих реакциях обеспечивается точная, строго специфичная последовательность мономерных звеньев в синтезируемых полимерах. Здесь происходит направленное стягивание мономеров в определенное место клетки — на молекулы, служащие матрицей, где реакция и осуществляется. Если бы такие реакции происходили путем случайного столкновения молекул, они протекали бы бесконечно медленно. Синтез сложных молекул на основе матричного принципа осуществляется быстро и точно.
Роль матрицы в матричных реакциях играют макромолекулы нуклеиновых кислот — ДНК или РНК. Мономерные молекулы, из которых синтезируется полимер, — нуклеотиды или аминокислоты - соответствии с принципом комплементарности располагаются и фиксируются на матрице в строго определенном, наперед заданном порядке. Затем происходит «сшивание» мономерных звеньев в полимерную цепь, и готовый полимер сбрасывается с матрицы. После этого матрица готова к сборке новой полимерной молекулы. Понятно, что, как на данной форме может производиться отливка только какой-то одной монеты, или медали, или какой-то одной буквы, так и на данной матричной молекуле может идти «сборка» только какого-то одного полимера.
Матричный тип реакций — специфическая особенность химизма живых систем. Они являются основой фундаментального свойства всего живого — его способности к воспроизведению себе подобного.
Кроме живой клетки, матричный тип реакций нигде в природе обнаружен не был.
Трансляция
Информация о структуре белка, записанная в и - РНК в виде последовательности нуклеотидов, переносится далее в виде последовательности аминокислот в синтезируемой полипептидной цепи. Этот процесс называется трансляцией («трансляциа»— перенесение, перевод, лат.). Для того чтобы разобраться в том, как в рибосомах происходит трансляция, т. е. перевод информации с языка нуклеиновых кислот на язык белков. Рибосомы на рисунке изображены в виде яйцевидных тел, унизывающих и-РНК. Первая рибосома вступает на нитевидную молекулу и-РНК с левого конца и начинает синтез белка. По мере сборки белковой молекулы рибосома ползет по и-РНК (на рисунке слева направо). Когда рибосома продвинется вперед на 50—100 А, с того же конца на и-РНК входит вторая рибосома, которая, как и первая, начинает синтез и движется вслед за первой рибосомой. Затем на и-РНК вступает третья рибосома, четвертая и т. д. Все они выполняют одну и ту же работу: каждая синтезирует один и тот же белок, запрограммированный на данной и-РНК. Чем дальше вправо продвинулась рибосома по и-РНК, тем больший отрезок белковой молекулы «собран». Когда рибосома достигает правого конца и-РНК, синтез окончен и рибосома вместе со своим «изделием» сваливается в окружающую среду. Здесь они расходятся: рибосома — на любую и-РНК (так как она способна к синтезу любого белка; характер белка зависит от матрицы), белковая молекула — в эндоплазматическую сеть и по ней перемещается в тот участок клетки, где требуется данный вид белка. Через короткое время заканчивает работу вторая рибосома, затем третья и т. д. А с левого конца и-РНК на нее вступают все новые и новые рибосомы, и синтез белка идет непрерывно. Число рибосом, умещающихся одновременно на молекуле и-РНК, зависит от длины и-РНК. Так, например, на молекуле и-РНК, программирующей синтез белка-гемоглобина, длина которой около 1500 А0, помещается до 5 рибосом (диаметр рибосомы приблизительно 230 А). Группа рибосом, помещающихся одновременно на одной молекуле и-РНК, называется полирибосомой или, сокращенно, полисомой.
Теперь остановимся подробнее на механизме работы рибосомы. Рибосома во время своего движения по и-РНК в каждый данный момент находится в контакте с небольшим участком ее молекулы. Возможно, что размер этого участка составляет всего один триплет нуклеотидов. Рибосома передвигается по и-РНК не плавно, а прерывисто, «шажками» — триплет за триплетом. На некотором, расстоянии от места контакта рибосомы с и-РНК находится пункт «сборки» белка: здесь помещается и работает фермент белок-синтетаза, создающий полипептидную цепь, т. е. образующий пептидные связи между аминокислотами.
Сам механизм «сборки» белковой молекулы в рибосомах осуществляется следующим образом. В каждую рибосому, входящую в состав полисомы, т.е. движущуюся по и-РНК, из окружающей среды непрерывным потоком идут молекулы т-РНК с «навешанными» на них аминокислотами. Они проходят, задевая своим кодовым концом место контакта рибосомы с и-РНК, дотрагиваются до триплета нуклеотидов и-РНК, который в данный момент находится в рибосоме. Противоположный конец т-РНК (несущий аминокислоту) оказывается при этом вблизи пункта «сборки» белка. Однако только в том случае, если кодовый триплет т-РНК окажется комплементарным к триплету и-РНК (находящемуся в данный момент в рибосоме), аминокислота, доставленная т-РНК, попадет в состав молекулы белка и отделится от т-РНК. Тотчас же рибосома делает «шаг» вперед по и-РНК на один триплет, а свободная т-РНК выбрасывается из рибосомы в окружающую среду. Здесь она захватывает новую молекулу аминокислоты и несет ее в любую из работающих рибосом. Так постепенно, триплет за триплетом, движется по и-РНК рибосома и растет звено за звеном — полипептидная цепь. Так работает рибосома — этот удивительный органоид клетки, который с полным правом называют «молекулярным автоматом» синтеза белка.
Мы уже упоминали о синтезе белка, недавно осуществленном химиками в лабораторных условиях. Этот искусственный синтез потребовал огромных усилий, много времени и средств. А в живой клетке синтез одной молекулы белка завершается в 3—4 секунды. Вот пример, насколько совершеннее работает синтетический аппарат живой клетки.
Роль ферментов в биосинтезе белка
Не следует забывать, что ни один шаг в процессе синтеза белка не идет без участия ферментов. Все реакции белкового синтеза катализируются специальными ферментами. Синтез информационной РНК ведет фермент, который «ползет» вдоль молекулы ДНК от начала гена до его конца и оставляет позади себя готовую молекулу информационной РНК. Ген в этом процессе дает только программу для синтеза, а сам процесс осуществляет фермент. Без участия ферментов не происходит и соединения аминокислот с транспортной РНК.
myunivercity.ru
Цель: углубить знания о метаболизме клеток путем реализации наследственной информации в процессе биосинтеза белка; продолжить формирование знаний о хранении информации о белках в ДНК, познакомить с понятием генетический код и основными его свойствами; рассмотреть особенности транскрипции у эукариот как одного из этапов биосинтеза.
Свойства белков:
Для этого вы должны вспомнить про ДНК: что это такое, для чего служит в клетке? ДНК это двуспиральная молекула дезоксирибозы, являющаяся носителем наследственной информации.
2. Изучение нового материала.
Синтез белка состоит из двух этапов – транскрипции и трансляции.
Транскрипция – для того, чтобы синтезировать и- РНК, участок ДНК «разматывается», деспирализуется, а затем по принципу комплементарности на одной из цепочек ДНК с помощью ферментов синтезируются молекулы РНК. Это происходит следующим образом: против, например, гуанина молекулы ДНК становится цитозин молекулы РНК, против аденина - урацил, т.к. У РНК нет такой нуклеиновый кислоты как тимин., напротив тимина- аденин, цитозина- гуанин. Таким образом, формируется цепочка и- РНК, представляющая собой точную копию второй цепи ДНК. Таким образом, информация о последовательности нуклеотидов какого-либо гена ДНК «переписывается» последовательность нуклеотидов и-РНК. У прокариот синтезированные мо и- РНК сразу же могут взаимодействовать с рибосомами, и начинается синтез белка, у эукариот и- РНК взаимодействует в ядре со специальными белками и переносятся через поры в ядерной оболочке в цп.
Строение т- РНК. Для переноса каждого вида аминокислоты, а их как мы помним всего 20 участвуют в синтезе белка, столько же существует и видов т-РНК. Они очень схожи по строению. Они «носятся» по цитоплазме захватывая аминокислоты и перенося их к рибосомам - непосредственному месту синтеза белка. По форме т-РНК напоминает лист клевера. Виды т- РНК обязательно различаются по триплету нуклеотидов, расположенному «на верхушке». Это антикодон по генетическому коду соответствует той аминокислоте, которую ему предстоит переносить. К «черешку листа » специальный фермент присоединяет ту аминокислоту, которая кодируется триплетом, комплементарным антикодону.
Трансляция. В цп происходит трансляция. На тот конец и- РНК, с которого нужно начать синтез белка, нанизывается рибосома, она перемещается по цепочке и- РНК прерывисто, скачками, задерживаясь на каждом триплете. За это мгновение одна т –РНК из многих способна опознать своим антикодоном триплет, на котором находится рибосома. И если антикодон комплементарен триплету и –РНК, аминокислота отсоединяется от «черешка листа» и присоединяется пептидной связью к растущей белковой цепочке. Эта операция повторяется столько раз, сколько аминокислот должен содержать «строящийся» белок. Когда же в рибосоме оказывается один из триплетов , являющийся стоп-сигналом между генами, то ни одна т- РНК к такому триплету присоединиться не может, т.к. антикодонов к ним у т- РНК нет.
Генетический код - свойственный всем живым организмам способ кодирования аминокислотной последовательности белков при помощи последовательности нуклеотидов.
Свойства генетического кода
Триплетность — значащей единицей кода является сочетание трёх нуклеотидов (триплет, или кодон).
Непрерывность — между триплетами нет знаков препинания, то есть информация считывается непрерывно.
Неперекрываемость — один и тот же нуклеотид не может входить одновременно в состав двух или более триплетов (не соблюдается для некоторых перекрывающихся генов вирусов, митохондрий и бактерий, которые кодируют несколько белков, считывающихся со сдвигом рамки).
Однозначность (специфичность) — определённый кодон соответствует только одной аминокислоте (однако, кодон UGA у Euplotes crassus кодирует две аминокислоты — цистеин и селеноцистеин)[1]
Вырожденность (избыточность) — одной и той же аминокислоте может соответствовать несколько кодонов.
Универсальность — генетический код работает одинаково в организмах разного уровня сложности — от вирусов до человека (на этом основаны методы генной инженерии; есть ряд исключений, показанный в таблице раздела «Вариации стандартного генетического кода» ниже).
Помехоустойчивость — мутации замен нуклеотидов не приводящие к смене класса кодируемой аминокислоты, называют консервативными; мутации замен нуклеотидов, приводящие к смене класса кодируемой аминокислоты, называют радикальными.

bukvasha.ru
