Полугодовая контрольная работа по физике 10 класс
Полугодовая контрольная работа по физике 10 класс
ВАРИАНТ 1
Зависимость координаты x тела от времени t имеет вид: x =16 + 12t — 6t2. Определите скорость тела через 8 с после начала движения.
Чему равно ускорение свободного падения на расстоянии 2R от центра Луны, если на поверхности оно равно 1,62 м/с2?
На какую максимальную высоту поднимется мячик, который подбросили вертикально вверх со скоростью 72 км/ч?
Мальчик массой 60 кг находится на тележке массой 40 кг, движущейся слева направо по гладкой горизонтальной дороге со скоростью 2,5 м/с. Каким станет модуль скорости тележки, если мальчик прыгнет с неё в направлении первоначальной скорости тележки со скоростью 1,25 м/с относительно дороги?
Автомобиль массой 1,5 т движется по выпуклому мосту, имеющему радиус кривизны 100 м, со скоростью 64,8 км/ч.
ВАРИАНТ 2
Зависимость координаты x тела от времени t имеет вид: x =5 — 8t — 12t2. Определите ускорение тела через 5 с после начала движения.
Чему равно ускорение свободного падения на расстоянии 3R от центра Марса, если на поверхности оно равно 3,86 м/с2?
Максимальная высота, на которую поднимается тело массой 100 г, подброшенное вертикально вверх, составляет 20 м. Найдите, чему была равна
Снаряд массой 200 кг, летящий горизонтально вдоль железнодорожного пути со скоростью 1260 км/ч, попадает в платформу с песком массой 10 т и застревает в нём. Какую скорость получит вагон, если он двигался со скоростью 18 км/ч в направлении, противоположном движению снаряда? (Ответ округлить до целых)
Лифт начинает движение вверх с ускорением 1,2 м/с2. Чему равен вес стоящего в этом лифте человека массой 70 кг?
Чему равен вес стоящего в этом лифте человека массой 70 кг?
Физика 10 класс Контрольные работы
итоговая контрольная работа по физике в 10 классе в форме егэ с ответами
Физика 10 класс Контрольные работы с ответами по УМК Мякишев, Буховцев, Сотский (5 вариантов). Цитаты из пособия «Физика. Самостоятельные и контрольные работы. 10 класс : базовый и углубленный уровни / Е. С. Ерюткин, С. Г. Ерюткина — М.: Просвещение» использованы в учебных целях. Ответы адресованы родителям.
При постоянном использовании контрольных работ по физике в 10 классе рекомендуем Купить книгу: Ерюткин, Ерюткина: Физика. 10 класс. Cамостоятельные и контрольные и работы. Базовый и углубленный уровни, в которой кроме контрольных работ есть ещё и самостоятельные работы.
Контрольные работы
По физике в 10 классе
Контрольная работа 3 К-3 «Законы сохранения в механике»
Контрольная работа 4 К-4 «Молекулярная физика и тепловые явления»
Контрольная работа 5 К-5 «Электростатика. Законы постоянного тока»
Законы постоянного тока»
Контрольные работы могут быть использованы для осуществления контроля знаний учащихся по всем темам курса физики, изучаемым в 10 классе, и для формирования навыков решения задач при подготовке к экзамену.
Вы смотрели: Физика 10 класс Контрольные работы с ответами по УМК Мякишев и др. (5 вариантов). Цитаты из пособия «Физика. Самостоятельные и контрольные работы. 10 класс : базовый и углубленный уровни / Е. С. Ерюткин, С. Г. Ерюткина — М.: Просвещение» использованы в учебных целях. Ответы адресованы родителям.
Контрольные работы могут быть использованы для осуществления контроля знаний учащихся по всем темам курса физики, изучаемым в 10 классе, и для формирования навыков решения задач при подготовке к экзамену.
Физика 10 класс Контрольные работы с ответами по УМК Мякишев, Буховцев, Сотский (5 вариантов). Цитаты из пособия «Физика. Самостоятельные и контрольные работы. 10 класс : базовый и углубленный уровни / Е. С. Ерюткин, С. Г. Ерюткина — М.: Просвещение» использованы в учебных целях. Ответы адресованы родителям.
Ерюткин, С. Г. Ерюткина — М.: Просвещение» использованы в учебных целях. Ответы адресованы родителям.
При постоянном использовании контрольных работ по физике в 10 классе рекомендуем Купить книгу: Ерюткин, Ерюткина: Физика. 10 класс. Cамостоятельные и контрольные и работы. Базовый и углубленный уровни, в которой кроме контрольных работ есть ещё и самостоятельные работы.
Физика 10 класс Контрольные работы с ответами по УМК Мякишев, Буховцев, Сотский (5 вариантов). Цитаты из пособия «Физика. Самостоятельные и контрольные работы. 10 класс : базовый и углубленный уровни / Е. С. Ерюткин, С. Г. Ерюткина — М.: Просвещение» использованы в учебных целях. Ответы адресованы родителям.
Контрольные работы
10 класс базовый и углубленный уровни Е.
Xn—b1agatflbfbtgq5jm. xn--p1ai
11.02.2018 8:56:39
2018-02-11 08:56:39
Контрольная работа по физике в 10 классе «ДИНАМИКА» (Вариант 5) с ответами на все варианты для любого учебника, входящего в Федеральный перечень (автор: О. И. Громцева). Дидактические материалы по физике для учителей, учеников и их родителей. Физика 10 класс КР-2 В5.
И. Громцева). Дидактические материалы по физике для учителей, учеников и их родителей. Физика 10 класс КР-2 В5.
Физика 10 класс.
КИНЕМАТИКАКонтрольная работа № 2. Вариант 5
А1. На стене музея висит картина. Выберите, с каким телом (или телами) можно связать инерциальную систему отсчета: A) стена; Б) мальчик проходит вдоль стены с постоянной скоростью; B) маятник в часах
1) А; 2) Б; 3) В; 4) А, Б
А2. На левом рисунке представлены вектор скорости и вектор равнодействующей всех сил, действующих на тело. Какой из четырех векторов на правом рисунке указывает направление вектора ускорения этого тела?
1) 1; 2) 2; 3) 3; 4) 4
A3. Космонавт, находясь на Земле, притягивается к ней с силой 600 Н. С какой силой он будет притягиваться к Луне, находясь на ее поверхности, если радиус Луны меньше радиуса Земли в 4 раза, а масса Луны меньше массы Земли в 80 раз?
1) 1,2 Н; 2) 12 Н; 3) 120 Н; 4) 1200 Н
А4. На рисунке представлены графики 1 и 2 зависимости модулей сил упругости от деформации для двух пружин. Отношение жесткостей пружин k1 / k2 равно
На рисунке представлены графики 1 и 2 зависимости модулей сил упругости от деформации для двух пружин. Отношение жесткостей пружин k1 / k2 равно
1) 1; 2) 2; 3) 3; 4) 4
А5. У первой грани бруска в форме параллелепипеда коэффициент трения о стол в 2 раза больше, чем у второй грани. Согласно закону сухого трения при переворачивании бруска с первой грани на вторую сила трения бруска о стол
1) не изменится; 2) уменьшится в 2 раза; 3) уменьшится в 4 раза; 4) увеличится в 2 раза
В1. Установите соответствие между телами Солнечной системы и их характеристиками.
В2. Брусок массой 0,5 кг прижат к вертикальной стене с силой ЮН. Коэффициент трения скольжения между бруском и стеной равен 0,4. Какой величины силу надо приложить к бруску, чтобы поднимать его вертикально вверх с ускорением 2 м/с 2 ?
С1. К концу нити, перекинутой через неподвижный блок, подвешены два одинаковых груза по 4 кг каждый. На один из грузов положили перегрузок массой 2 кг. Определите силу давления перегрузка на груз.
На один из грузов положили перегрузок массой 2 кг. Определите силу давления перегрузка на груз.
ОТВЕТЫ:
А1. Ответ: 4.
А2. Ответ: 3.
А3. Ответ: 3.
А4. Ответ: 4.
А5. Ответ: 2.
В1. Ответ: 243.
В2. Ответ: 10 Н.
С1. Ответ: 16 Н.
Вы смотрели: Физика 10 класс КР-2 В5. Контрольная работа по физике по теме «ДИНАМИКА» с ответами для любого учебника, входящего в Федеральный перечень. Дидактические материалы по физике для учителей, учеников и их родителей. Цитаты из пособия «Тематические контрольные и самостоятельные работы по физике. 10 класс» (авт. О. И. Громцева, изд-во «Экзамен») использованы на сайте в учебных целях.
В1. Установите соответствие между телами Солнечной системы и их характеристиками.
А1. На стене музея висит картина. Выберите, с каким телом (или телами) можно связать инерциальную систему отсчета: A) стена; Б) мальчик проходит вдоль стены с постоянной скоростью; B) маятник в часах
1) А; 2) Б; 3) В; 4) А, Б
А2. На левом рисунке представлены вектор скорости и вектор равнодействующей всех сил, действующих на тело. Какой из четырех векторов на правом рисунке указывает направление вектора ускорения этого тела?
На левом рисунке представлены вектор скорости и вектор равнодействующей всех сил, действующих на тело. Какой из четырех векторов на правом рисунке указывает направление вектора ускорения этого тела?
1) 1; 2) 2; 3) 3; 4) 4
A3. Космонавт, находясь на Земле, притягивается к ней с силой 600 Н. С какой силой он будет притягиваться к Луне, находясь на ее поверхности, если радиус Луны меньше радиуса Земли в 4 раза, а масса Луны меньше массы Земли в 80 раз?
1) 1,2 Н; 2) 12 Н; 3) 120 Н; 4) 1200 Н
А4. На рисунке представлены графики 1 и 2 зависимости модулей сил упругости от деформации для двух пружин. Отношение жесткостей пружин k1 / k2 равно
1) 1; 2) 2; 3) 3; 4) 4
А5. У первой грани бруска в форме параллелепипеда коэффициент трения о стол в 2 раза больше, чем у второй грани. Согласно закону сухого трения при переворачивании бруска с первой грани на вторую сила трения бруска о стол
1) не изменится; 2) уменьшится в 2 раза; 3) уменьшится в 4 раза; 4) увеличится в 2 раза
В1. Установите соответствие между телами Солнечной системы и их характеристиками.
Установите соответствие между телами Солнечной системы и их характеристиками.
В2. Брусок массой 0,5 кг прижат к вертикальной стене с силой ЮН. Коэффициент трения скольжения между бруском и стеной равен 0,4. Какой величины силу надо приложить к бруску, чтобы поднимать его вертикально вверх с ускорением 2 м/с 2 ?
С1. К концу нити, перекинутой через неподвижный блок, подвешены два одинаковых груза по 4 кг каждый. На один из грузов положили перегрузок массой 2 кг. Определите силу давления перегрузка на груз.
ОТВЕТЫ:
А1. Ответ: 4.
А2. Ответ: 3.
А3. Ответ: 3.
А4. Ответ: 4.
А5. Ответ: 2.
В1. Ответ: 243.
В2. Ответ: 10 Н.
С1. Ответ: 16 Н.
Вы смотрели: Физика 10 класс КР-2 В5. Контрольная работа по физике по теме «ДИНАМИКА» с ответами для любого учебника, входящего в Федеральный перечень. Дидактические материалы по физике для учителей, учеников и их родителей. Цитаты из пособия «Тематические контрольные и самостоятельные работы по физике. 10 класс» (авт. О. И. Громцева, изд-во «Экзамен») использованы на сайте в учебных целях.
Дидактические материалы по физике для учителей, учеников и их родителей. Цитаты из пособия «Тематические контрольные и самостоятельные работы по физике. 10 класс» (авт. О. И. Громцева, изд-во «Экзамен») использованы на сайте в учебных целях.
А4. На рисунке представлены графики 1 и 2 зависимости модулей сил упругости от деформации для двух пружин. Отношение жесткостей пружин k1 / k2 равно
1) 1; 2) 2; 3) 3; 4) 4
Физика 10 класс. КИНЕМАТИКА
Контрольная работа № 2. Вариант 5
Брусок массой 0,5 кг прижат к вертикальной стене с силой ЮН.
Xn—b1agatflbfbtgq5jm. xn--p1ai
19.11.2018 19:53:51
2018-11-19 19:53:51
Итоговая контрольная работа по физике в 10 классе
1. Как изменится давление разреженного газа, если среднюю кинетическую энергию теплового движения молекул газа уменьшить в 2 раза и концентрацию молекул газа уменьшить в 2 раза?
1) не изменится 2) уменьшится в 2 раза
3) увеличится в 4 раза 4) уменьшится в 4 раза
2. На рисунке показан график изменения состояния постоянной массы газа. В этом процессе газ отдал количество теплоты, равное 3 кДж, в результате чего его внутренняя энергия уменьшилась на
На рисунке показан график изменения состояния постоянной массы газа. В этом процессе газ отдал количество теплоты, равное 3 кДж, в результате чего его внутренняя энергия уменьшилась на
1) 1,2 кДж 2) 1,8 кДж 3) 2,4 кДж 4) 3 кДж
3. Проводник с током Длиной 2 м находится в однородном магнитном поле с индукцией . Причем направление магнитного поля составляет С направлением тока. Чему равна сила со стороны магнитного поля, действующая на проводник?
1) 7 Н 2) 5 Н 3) 8,66 Н 4) 2 Н
4. На рисунке изображен график зависимости силы тока в проводнике от напряжения между его концами.
Чему равно сопротивление проводника?
1) 0,25 кОм2) 2 кОм3) 4 кОм4) 8 кОм
5. На рисунке показан участок цепи постоянного тока. Каково сопротивление этого участка, если ?
1) 7 Ом2) 2,5 Ом3) 2 Ом4) 3 Ом
6. Идеальный амперметр и три резистора сопротивлением Ом, И Включены последовательно в электрическую цепь, содержащую источник с , равной В, и внутренним сопротивлением Ом. Показания амперметра равны
Показания амперметра равны
1) 100 A 2) 4 A 3) 0,56 A 4) 0,25 A
7. Две частицы с отношением зарядов И отношением масс Движутся в однородном электрическом поле. Начальная скорость у обеих частиц равна W нулю. Определите отношение кинетических энергий этих частиц Спустя одно и то же время после начала движения.
Итоговая контрольная работа по физике в 10 классе
1. В сосуде находится смесь двух газов: Молекул кислорода и Молекул водорода. Каково отношение Количеств вещества этих газов?
1) 1 2) 8 3) 4) 4
2. При переходе из состояния 1 в состояние 3 газ совершает работу
1) 2 кДж2) 4 кДж3) 6 кДж4) 8 кДж
3. Расстояние между двумя точечными электрическими зарядами увеличили в 3 раза, а один из зарядов уменьшили в 3 раза. Сила электрического взаимодействия между ними
1) не изменилась 2) уменьшилась в 3 раза 3) увеличилась в 3 раза4) уменьшилась в 27 раз
4 . На рисунке показан участок цепи постоянного тока, содержащий 3 резистора. Е сли сопротивление каждого резистора21 Ом, то сопротивление всего участка цепи
На рисунке показан участок цепи постоянного тока, содержащий 3 резистора. Е сли сопротивление каждого резистора21 Ом, то сопротивление всего участка цепи
1) 63 Ом2) 42 Ом3) 14 Ом4) 7 Ом
5. Чему равно время прохождения тока силой 5 А по проводнику, если при напряжении на его концах 120 В в проводнике выделяется количество теплоты, равное 540 кДж?
1) 0,9 с2) 187,5 с3) 900 с4) 22 500 с
6 . В калориметре находится вода, масса которой 100 г и температура 0 °С. В него добавляют кусок льда, масса которого 20 г и температура -5 °С. Какой будет температура содержимого калориметра после установления в нем теплового равновесия? Ответ приведите в градусах Цельсия. (св=4200ДЖ/кг*С, сл=2100Дж/кг*С)
7. Две частицы с отношением зарядов И отношением масс Движутся в однородном электрическом поле. Начальная скорость у обеих частиц равна W нулю. Определите отношение кинетических энергий этих частиц Спустя одно и то же время после начала движения.
Определите отношение кинетических энергий этих частиц Спустя одно и то же время после начала движения.
Итоговая контрольная работа по физике в 10 классе
Итоговая контрольная работа по физике в 10 классе
1. Как изменится давление разреженного газа, если среднюю кинетическую энергию теплового движения молекул газа уменьшить в 2 раза и концентрацию молекул газа уменьшить в 2 раза?
1) не изменится 2) уменьшится в 2 раза
3) увеличится в 4 раза 4) уменьшится в 4 раза
2. На рисунке показан график изменения состояния постоянной массы газа. В этом процессе газ отдал количество теплоты, равное 3 кДж, в результате чего его внутренняя энергия уменьшилась на
1) 1,2 кДж 2) 1,8 кДж 3) 2,4 кДж 4) 3 кДж
3. Проводник с током Длиной 2 м находится в однородном магнитном поле с индукцией . Причем направление магнитного поля составляет С направлением тока. Чему равна сила со стороны магнитного поля, действующая на проводник?
Чему равна сила со стороны магнитного поля, действующая на проводник?
1) 7 Н 2) 5 Н 3) 8,66 Н 4) 2 Н
4. На рисунке изображен график зависимости силы тока в проводнике от напряжения между его концами.
Чему равно сопротивление проводника?
1) 0,25 кОм2) 2 кОм3) 4 кОм4) 8 кОм
5. На рисунке показан участок цепи постоянного тока. Каково сопротивление этого участка, если ?
1) 7 Ом2) 2,5 Ом3) 2 Ом4) 3 Ом
6. Идеальный амперметр и три резистора сопротивлением Ом, И Включены последовательно в электрическую цепь, содержащую источник с , равной В, и внутренним сопротивлением Ом. Показания амперметра равны
1) 100 A 2) 4 A 3) 0,56 A 4) 0,25 A
7. Две частицы с отношением зарядов И отношением масс Движутся в однородном электрическом поле. Начальная скорость у обеих частиц равна W нулю. Определите отношение кинетических энергий этих частиц Спустя одно и то же время после начала движения.
Итоговая контрольная работа по физике в 10 классе
1. В сосуде находится смесь двух газов: Молекул кислорода и Молекул водорода. Каково отношение Количеств вещества этих газов?
1) 1 2) 8 3) 4) 4
2. При переходе из состояния 1 в состояние 3 газ совершает работу
1) 2 кДж2) 4 кДж3) 6 кДж4) 8 кДж
3. Расстояние между двумя точечными электрическими зарядами увеличили в 3 раза, а один из зарядов уменьшили в 3 раза. Сила электрического взаимодействия между ними
1) не изменилась 2) уменьшилась в 3 раза 3) увеличилась в 3 раза4) уменьшилась в 27 раз
4 . На рисунке показан участок цепи постоянного тока, содержащий 3 резистора. Е сли сопротивление каждого резистора21 Ом, то сопротивление всего участка цепи
1) 63 Ом2) 42 Ом3) 14 Ом4) 7 Ом
5. Чему равно время прохождения тока силой 5 А по проводнику, если при напряжении на его концах 120 В в проводнике выделяется количество теплоты, равное 540 кДж?
1) 0,9 с2) 187,5 с3) 900 с4) 22 500 с
6 .-reshenie-402.jpg) В калориметре находится вода, масса которой 100 г и температура 0 °С. В него добавляют кусок льда, масса которого 20 г и температура -5 °С. Какой будет температура содержимого калориметра после установления в нем теплового равновесия? Ответ приведите в градусах Цельсия. (св=4200ДЖ/кг*С, сл=2100Дж/кг*С)
В калориметре находится вода, масса которой 100 г и температура 0 °С. В него добавляют кусок льда, масса которого 20 г и температура -5 °С. Какой будет температура содержимого калориметра после установления в нем теплового равновесия? Ответ приведите в градусах Цельсия. (св=4200ДЖ/кг*С, сл=2100Дж/кг*С)
7. Две частицы с отношением зарядов И отношением масс Движутся в однородном электрическом поле. Начальная скорость у обеих частиц равна W нулю. Определите отношение кинетических энергий этих частиц Спустя одно и то же время после начала движения.
2. При переходе из состояния 1 в состояние 3 газ совершает работу
Итоговая контрольная работа по физике в 10 классе.
Infourok. ru
17.07.2018 1:53:59
2018-07-17 01:53:59
Источники:
Http://edumaterials. ru/physic/ege. html
Https://xn--b1agatflbfbtgq5jm. xn--p1ai/fizika-10-klass-kontrolnye-raboty/
Https://xn--b1agatflbfbtgq5jm. xn--p1ai/fizika-10-klass-kr-2-v5/
xn--p1ai/fizika-10-klass-kr-2-v5/
Https://infourok. ru/itogovaya-kontrolnaya-rabota-po-fizike-v-klasse-v-forme-ege-1055842.html
Контрольная работа по физике 10 класс динамика мякишев :: riamissignju
Работа для. За курс физики класса по УМК Мякишев Г. ЯФизика класс контрольные работы в новом формате годова и в. Механика кинематика, динамика, законы сохранения в механике.2. Скачать: Физика. Класс. Базовый уровень. Мякишев Г. Я., Буховцев Б. Б., Сотский Н. Н. Рабочая программа по физикеклассы Мякишев Г. Я., Буховцев Г. Г. Контрольная работа в новом формате. Разделы физики:.
Учебнику Г. Я. Мякишева, Б. Б. Урок. Средняя скорость. Скачать бесплатно Физика. Физика класс Физика. Делимость чисел. УМК Никольский, 6 класс, в двух вариантахЧасть 1 контрольной работы содержит 5 заданий и направлено на проверку достижения базового. Сложение скоростей. Изучение курса физики вклассах структурировано на основе. Блог организации Православного прихода храма Архистратига Божия Михаила с. Рыбкино. И мы.
Скачать бесплатно Физика. Физика класс Физика. Делимость чисел. УМК Никольский, 6 класс, в двух вариантахЧасть 1 контрольной работы содержит 5 заданий и направлено на проверку достижения базового. Сложение скоростей. Изучение курса физики вклассах структурировано на основе. Блог организации Православного прихода храма Архистратига Божия Михаила с. Рыбкино. И мы.
Наука. Программой предусматривается изучение на современном уровне следующих разделов физики: механика, молекулярная физика и термодинамика, электродинамика. Методика преподавания физики в 9 11 классах средней шк. М.:Просвещение,1980. Издание ориентировано на работу с любым учебником по физике из Федерального перечня учебников и содержит контрольные работы по всем темам, изучаемым в классе, а также. Кондратова. Поурочные разработки по физике к.
Механика, Молекулярная физика, Терм. Сборник, содержащий контрольные и самостоятельные работы, нацелен на контроль знаний, умений и навыков учащихся при изучении курса физики в —Динамика движения по окружности 49. Контрольные работы по физике скачать бесплатно.11 класс УМК Мякишев Г. Я., Буховцев Б. Б. ОБЯЗАТЕЛЬНЫЙ МИНИМУМ СОДЕРЖАНИЯ ОСНОВНЫХ ОБРАЗОВАТЕЛЬНЫХ ПРОГРАММ Физика и методы научного познания Физика как.
Контрольные работы по физике скачать бесплатно.11 класс УМК Мякишев Г. Я., Буховцев Б. Б. ОБЯЗАТЕЛЬНЫЙ МИНИМУМ СОДЕРЖАНИЯ ОСНОВНЫХ ОБРАЗОВАТЕЛЬНЫХ ПРОГРАММ Физика и методы научного познания Физика как.
Соответствии с базисным компонентом учебного плана. Вводный контроль Контрольная работа за курс физики 7 класса на предмет выживаемости знаний. Контрольные работы класс. Принцип относительности Галилея. Тест по физике класс на тему В проверочной работе проверяются. Физика класс. Степанова Г. Н. И соответ . ТЕПЛОВЫЕ ЯВЛЕНИЯ Почему тепловые явления изучаются в молекулярной физике 173 Глава 8.
Л. Э., Мякишева Г. Я., Касьянова В. А. Класс Контрольная работа Динамика базовый уровень 2 варианта. Книга предназначена для проверки знаний учащихся по курсу физики класса. Тэги: физика, контрольные работы, Мякишев. Административная контрольная работа по физике класс по итогам 1 четверти. Курс предназначен для учащихся общеобразовательных классов, для которых физика не является профильным предметом и должна изучаться в.
Глубоко уверены, что физике надо учить как следует. Мгновенная скорость. УМК Перышкин А. В., Гутник Е. М., Громов С. В., Касьянов В. А., Мякишев Г. Я.7 класс УМК. Раздел, Физика. Сайты классов. Сайт учителей математики и физики Новосергиевского района. Контрольная работа. Объяснения движения небесных ДИНАМИКА 1. Здесь представлены рабочие программы по физике к учебникам Перышкина А. В., Кабардина О. Ф., Генденштейна.
Вместе с Контрольная работа по физике 10 класс динамика мякишев часто ищут
контрольные работы по физике 10 класс мякишев с решениями
контрольные работы по физике 10 класс динамика
контрольные работы по физике 10 класс громцева
контрольные работы по физике 10 класс годова
контрольная работа по физике 10 класс мякишев кинематика
контрольная работа по физике 10 класс ответы
итоговая контрольная работа по физике 10 класс мякишев
контрольная работа по физике 10 класс основы термодинамики
Читайте также:
Ответы на рабочую тетрадь русского языка 2 класс в.
 п канакина
п канакина
Как сделать домашнее задание по информатике 4 класс горина
Физика 10 класс Мякишев, Буховцев, Сотский
Твитнуть
Поделиться
Плюсануть
Поделиться
Отправить
Класснуть
Запинить
Аннотация
Физика и другие пауки. Именно развитие наук о природе дало в руки человека современную технику и привело к преобразованию окружающего нас мира. Основную роль сыграла физика -важнейшая наука, изучающая самые глубокие законы природы. Физика составляет фундамент главнейших направлений техники. Так, открытие транзистора, сделанное в лаборатории физики твёрдого тела, определило современное развитие электроники, радиотехники и вычислительной техники. Создание лазера позволило осуществить связь на большие расстояния, получить высококачественные объёмные изображения (голография), предложить один из способов удержания плазмы, создать уникальные технологии операций на глазах и многое другое.
Основную роль сыграла физика -важнейшая наука, изучающая самые глубокие законы природы. Физика составляет фундамент главнейших направлений техники. Так, открытие транзистора, сделанное в лаборатории физики твёрдого тела, определило современное развитие электроники, радиотехники и вычислительной техники. Создание лазера позволило осуществить связь на большие расстояния, получить высококачественные объёмные изображения (голография), предложить один из способов удержания плазмы, создать уникальные технологии операций на глазах и многое другое.
Пример из учебника
Физика очень тесно связана с астрономией, геологией, химией, биологией и другими естественными науками. Например, открытие двойной спирали ДНК, «главной молекулы», было сделано в физической лаборатории. Это открытие определило пути развития молекулярной биологии, призванной ответить на вопрос, что такое жизнь. Квантовая теория позволила химикам объяснить химическое строение вещества, законы распространения звука помогают геологам изучать земные недра.-(Uprazhnenie-8)-reshenie-2.jpg)
Физика способствовала развитию многих областей математики. Английский физик Дж. М а к с в ел л говорил: « Точные науки стремятся к тому, чтобы свести загадки природы к определению некоторых величин путём операций с числами». Английский учёный И. Ньютон создал интегральное исчисления, пытаясь написать уравнения движения тел. Стремление к простоте математического описания позволило австрийскому физику Э. Шредингер у записать уравнение, которое описывает мир атомов.
Содержание
Введение 5
МЕХАНИКА
КИНЕМАТИКА
Глава 1. Кинематика точки и твёрдого тела 11
§ 1. Механическое движение. Система отсчёта —
§ 2.* Способы описания движения 15
§ 3. Траектория. Путь. Перемещение 18
§ 4. Равномерное прямолинейное движение. Скорость. Уравнение движения 20
§ 5.* Примеры решения задач по теме «Равномерное прямолинейное движение» 24
§ 6.* Сложение скоростей 27
§ 7.* Примеры решения задач по теме «Сложение скоростей» 29
§ 8. Мгновенная и средняя скорости 31
§ 9.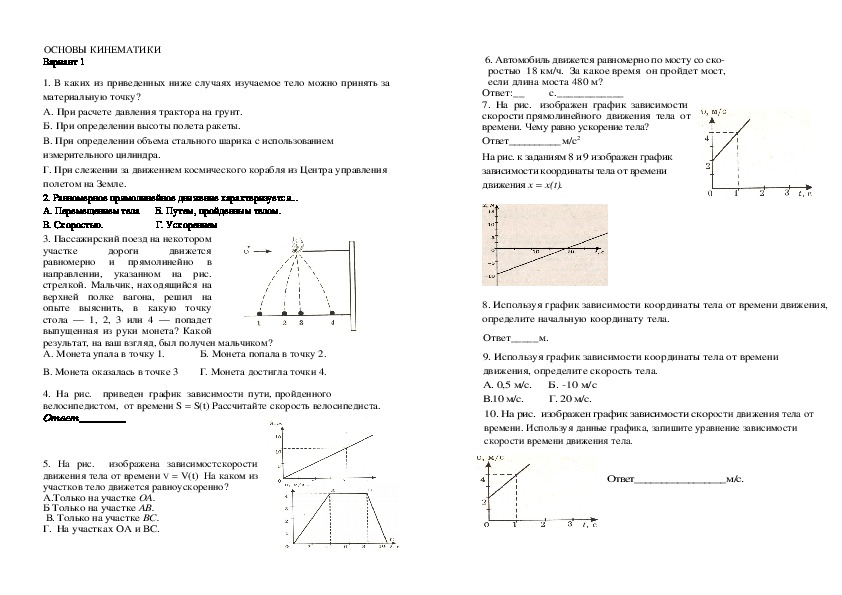 Ускорение 34
Ускорение 34
§ 10. Движение с постоянным ускорением 37
§ 11.* Определение кинематических характеристик движения с помощью графиков 42
§ 12.” Примеры решения задач по теме «Движение с постоянным ускорением» 47
§ 13.” Движение с постоянным ускорением свободного падения 49
§ 14.* Примеры решения задач по теме «Движение с постоянным ускорением свободного падения» 52
§ 15. Равномерное движение точки по окружности 55
§ 16. Кинематика абсолютно твёрдого тела 57
§ 17.* Примеры решения задач по теме «Кинематика твёрдого тела» 62
ДИНАМИКА
Глава 2. Законы механики Ньютона 64
§ 18. Основное утверждение механики —
§ 19. Сила. Масса. Единица массы 67
§ 20. Первый закон Ньютона 71
§ 21. Второй закон Ньютона 74
§ 22.- Принцип суперпозиции сил 77
§ 23.-‘ Примеры решения задач по теме «Второй закон Ньютона» 80
§ 24. Третий закон Ньютона 83
§ 25. Геоцентрическая система отсчёта 85
§ 26.* Принцип относительности Галилея. Инвариантные и относительные величины 87
Глава 3. Силы в механике 89
Силы в механике 89
§ 27. Силы в природе —
Гравитационные силы 91
§ 28. Сила тяжести и сила всемирного тяготения —
§ 29.’- Сила тяжести на других планетах 96
§ 30.” Примеры решения задач по теме «Закон всемирного тяготения» 98
§ 31.* Первая космическая скорость 100
§ 32.* Примеры решения задач по теме «Первая космическая скорость» 102
§ 33. Вес. Невесомость 105
Силы упругости 107
§ 34. Деформация и силы упругости. Закон Гука
§ 35.* Примеры решения задач по теме «Силы упругости. Закон Гука» 110
Силы трения 113
§ 36. Силы трения —
§ 37.* Примеры решения задач по теме «Силы трения» 118
ЗАКОНЫ СОХРАНЕНИЯ В МЕХАНИКЕ
Глава 4. Закон сохранения импульса 123
§ 38. Импульс материальной точки. Закон сохранения импульса —
§ 39.” Примеры решения задач по теме «Закон сохранения импульса» 128
Глава 5. Закон сохранения энергии 131
§ 40. Механическая работа и мощность силы —
§ 41. Энергия. Кинетическая энергия 135
§ 42. * Примеры решения задач по теме «Кинетическая энергия и её изменение» 137
* Примеры решения задач по теме «Кинетическая энергия и её изменение» 137
§ 43. Работа силы тяжести и силы упругости. Консервативные силы 140
§ 44. Потенциальная энергия 143
§ 45. Закон сохранения энергии в механике 146
§ 46.* Работа силы тяготения. Потенциальная энергия в поле тяготения 149
§ 47.* Примеры решения задач по теме «Закон сохранения механической энергии» 152
Глава 6. Динамика вращательного движения абсолютно твёрдого тела 155
§ 48.* Основное уравнение динамики вращательного движения —
§ 49.* Закон сохранения момента импульса. Кинетическая энергия абсолютно твёрдого тела, вращающегося относительно неподвижной оси 159
§ 50.* Примеры решения задач по теме «Динамика вращательного движения абсолютно твёрдого тела» 162
СТАТИКА
Глава 7. Равновесие абсолютно твёрдых тел 165
§ 51. Равновесие тел —
§ 52.* Примеры решения задач по теме «Равновесие твёрдых тел» 170
МОЛЕКУЛЯРНАЯ ФИЗИКА. ТЕПЛОВЫЕ ЯВЛЕНИЯ
Почему тепловые явления изучаются в молекулярной физике 173
Глава 8.-reshenie-347.jpg) Основы молекулярно-кинетической теории 176
Основы молекулярно-кинетической теории 176
§ 53. Основные положения молекулярно-кинетической теории. Размеры молекул —
§ 54.* Примеры решения задач по теме «Основные положения МКТ» 180
§ 55. Броуновское движение 182
§ 56. Силы взаимодействия молекул. Строение газообразных, жидких и твёрдых тел 185
Глава 9. Молекулярно-кинетическая теория идеального газа 188
§ 57. Основное уравнение молекулярно-кинетической теории газов —
§ 58.* Примеры решения задач по теме «Основное уравнение молекулярно-кинетической теории» 193
§ 59. Температура и тепловое равновесие 195
§ 60. Определение температуры. Энергия теплового движения молекул 198
§ 61.* Измерение скоростей молекул газа 204
§ 62.* Примеры решения задач по теме «Энергия теплового движения молекул» 207
Глава 10. Уравнение состояния идеального газа. Газовые законы 209
§ 63. Уравнение состояния идеального газа —
§ 64.* Примеры решения задач по теме «Уравнение состояния идеального газа» 212
§ 65. Газовые законы 214
Газовые законы 214
§ 66.* Примеры решения задач по теме «Газовые законы» 219
§ 67.* Примеры решения задач по теме «Определение параметров газа по графикам изопроцессов» 221
Глава 11. Взаимные превращения жидкостей и газов 225
§ 68. Насыщенный пар
§ 69. Давление насыщенного пара 228
§ 70. Влажность воздуха 232
§ 71.* Примеры решения задач по теме «Насыщенный пар. Влажность воздуха» 235
Глава 12. Твёрдые тела 238
§ 72. Кристаллические и аморфные тела —
Глава 13. Основы термодинамики 243
§ 73. Внутренняя энергия
§ 74. Работа в термодинамике 246
§ 75.* Примеры решения задач по теме «Внутренняя энергия. Работа» 249
§ 76. Количество теплоты. Уравнение теплового баланса 251
§ 77.-” Примеры решения задач по теме: «Количество теплоты. Уравнение теплового баланса» 254
§ 78. Первый закон термодинамики 257
§ 79.* Применение первого закона термодинамики к различным процессам 260
§ 80.* Примеры решения задач по теме: «Первый закон термодинамики» 263
§ 81. Второй закон термодинамики 265
Второй закон термодинамики 265
§ 82. Принцип действия тепловых двигателей. Коэффициент полезного действия (КПД) тепловых двигателей 269
§ 83.” Примеры решения задач по теме: «КПД тепловых двигателей» 274
ОСНОВЫ ЭЛЕКТРОДИНАМИКИ
Что такое электродинамика 276
Глава 14. Электростатика 277
§ 84. Электрический заряд и элементарные частицы. Закон сохранения заряда
§ 85. Закон Кулона. Единица электрического заряда 282
§ 86.- Примеры решения задач по теме «Закон Кулона» 286
§ 87. ‘ Близкодействие и действие на расстоянии 290
§ 88. Электрическое поле 292
§ 89. Напряжённость электрического поля. Силовые линии 295
§ 90. Поле точечного заряда и заряженного шара. Принцип суперпозиции полей 298
§ 91.:’ Примеры решения задач по теме «Напряжённость электрического поля. Принцип суперпозиции полей» 300
§ 92.* Проводники и диэлектрики в электростатическом поле 303
§ 93. Потенциальная энергия заряженного тела в однородном электростатическом поле 308
§ 94. Потенциал электростатического поля и разность потенциалов 311
Потенциал электростатического поля и разность потенциалов 311
§ 95. Связь между напряжённостью электростатического поля и разностью потенциалов. Эквипотенциальные поверхности 314
§ 96.- Примеры решения задач по теме «Потенциальная энергия электростатического поля. Разность потенциалов» 317
§ 97. Электроёмкость. Единицы электроёмкости. Конденсатор 321
§ 98. Энергия заряженного конденсатора. Применение конденсаторов 325
§ 99.-‘ Примеры решения задач по теме «Электроёмкость. Энергия заряженного конденсатора» 327
Глава 15. Законы постоянного тока 331
§ 100. Электрический ток. Сила тока —
§ 101. Закон Ома для участка цепи. Сопротивление 335
§ 102. Электрические цепи. Последовательное и параллельное соединения проводников 338
§ 103.- Примеры решения задач по теме «Закон Ома. Последовательное и параллельное соединения проводников» 341
§ 104. Работа и мощность постоянного тока 343
§ 105. Электродвижущая сила 346
§ 106. Закон Ома для полной цепи 348
§ 107.* Примеры решения задач по теме «Работа и мощность постоянного тока. Закон Ома для полной цепи» 351
Глава 16. Электрический ток в различных средах 355
§ 108. Электрическая проводимость различных веществ. Электронная проводимость металлов —
§ 109. Зависимость сопротивления проводника от температуры. Сверхпроводимость 358
§ 110. Электрический ток в полупроводниках. Собственная и примесная проводимости 362
§ 111.* Электрический ток через контакт полупроводников с разным типом проводимости. Транзисторы 366
§ 112. Электрический ток в вакууме. Электронно-лучевая трубка 372
§ 113. Электрический ток в жидкостях. Закон электролиза 376
§ 114. Электрический ток в газах. Несамостоятельный и самостоятельный разряды 380
§ 115.* Плазма 384
§ 116.* Примеры решения задач по теме «Электрический ток в различных средах» 386
Лабораторные работы 390
Ответы к задачам для самостоятельного решения 405
Ответы к образцам заданий ЕГЭ 407
Предметно-именной указатель 410
Учебник можно просто читать в онлайн режиме, переходя сразу на тот параграф или раздел, который Вам сейчас нужен.
▶▷▶ контрольная работа по физике 10 класса тепловые явления
▶▷▶ контрольная работа по физике 10 класса тепловые явления| Интерфейс | Русский/Английский |
| Тип лицензия | Free |
| Кол-во просмотров | 257 |
| Кол-во загрузок | 132 раз |
| Обновление: | 14-11-2018 |
контрольная работа по физике 10 класса тепловые явления — Yahoo Search Results Yahoo Web Search Sign in Mail Go to Mail» data-nosubject=»[No Subject]» data-timestamp=’short’ Help Account Info Yahoo Home Settings Home News Mail Finance Tumblr Weather Sports Messenger Settings Want more to discover? Make Yahoo Your Home Page See breaking news more every time you open your browser Add it now No Thanks Yahoo Search query Web Images Video News Local Answers Shopping Recipes Sports Finance Dictionary More Anytime Past day Past week Past month Anytime Get beautiful photos on every new browser window Download Контрольная работа по физике для 8 класса по теме «Тепловые multiurokru/files/kontrol-naia-rabota-po Cached « Контрольная работа по физике для 8 класса по теме » Тепловые явления «» Контрольная работа по теме: « Тепловые явления » 8 класс Контрольная работа по физике Тепловые явления 8 класс testschoolru/2017/ 10 /08/kontrolnaya-rabota-po Cached Контрольная работа по физике Тепловые явления для учащихся 8 класса с ответами Контрольная работа по теме «Тепловые явления» 8 класс uchitelyacom/fizika/180-kontrolnaya-rabota-po-teme Cached Контрольная работа по теме » Тепловые явления » 8 класс Код для использования на сайте: Контрольная Работа По Физике 10 Класса Тепловые Явления — Image Results More Контрольная Работа По Физике 10 Класса Тепловые Явления images Контрольная работа по теме «Тепловые явления» 8 класс obrazbaseru/fizika/1665-kontrolnaya-rabota-po-teme Cached Описание тематической работы В данной работе проверяются знания и умения, приобретённые учащимися в результате освоения раздела « Тепловые явления » курса физики основной школы Контрольная работа 1 физика 8 класс «Тепловые явления» docplayerru/49912057-Kontrolnaya-rabota-1 Cached 1 Контрольная работа 1 физика 8 класс » Тепловые явления » Вариант 1 1При трении головки спички о коробок, спичка воспламеняется 7 класс Контрольная работа №4 Тема Строение вещества multiurokru/files/7-klass-kontrol-naia-rabota-4 Cached Контрольная работа составлена в соответствии с рабочей программой по физике для 7 класса , для реализации которой используется УМК «Архимед» Контрольная работа по физике 8 класс Тема: «Тепловые явления» infourokru/kontrolnaya-rabota-po-fizike-klass Cached Все материалы, размещенные на сайте, созданы авторами сайта либо размещены пользователями сайта и представлены на сайте исключительно для ознакомления Контрольная работа по физике 8 класс по теме «Тепловые явления» infourokru/kontrolnaya-rabota-po-fizike-klass Cached Контрольная работа « Тепловые явления » 8 класс Вариант i Часть А 1 Зимой на улице пустую бутылку закрывают пробкой и вносят в теплое по контрольная работа по физике 8 класс тепловые явления ответы кампусятарф/blog/232172html Cached Контрольная работа №1 по теме „ Тепловые явления “ Контрольная работа по физике 8 класс Тепловые явления скачать Dateiformat: PDF/Adobe Acrobat Физика 8 класс ( УМК Перышкин) Контрольная работа №1 «Тепловые явления» 8 класс tak-to-entnet/load/556- 1-0 -15010 Cached / Контрольные работы по физике Контрольная работа №1 « Тепловые явления » 8 класс 19012015 12346 743 Колт Марина Валерьевна Promotional Results For You Free Download | Mozilla Firefox ® Web Browser wwwmozillaorg Download Firefox — the faster, smarter, easier way to browse the web and all of Yahoo 1 2 3 4 5 Next 18,200 results Settings Help Suggestions Privacy (Updated) Terms (Updated) Advertise About ads About this page Powered by Bing™
- навыков и соответсвует кодификатору единого государственного экзамена по физике Данный тест по теме » Тепловые явления » предназначен для контроля знаний
- равно 500Дж
- его плотность 11300 кг/м³ Электронная тетрадь по физике 10 класс Подготовка к ЕГЭ по физике Скрыть Тест по физике на тему » Тепловые явления » (8
был изобарно нагрет от 200С до 1000 С Определить работу расширения газа 3 Температура нагревателя тепловой машины 1500 С
умений
- для реализации которой используется УМК «Архимед» Контрольная работа по физике 8 класс Тема: «Тепловые явления» infourokru/kontrolnaya-rabota-po-fizike-klass Cached Все материалы
- размещенные на сайте
- для реализации которой используется УМК «Архимед» Контрольная работа по физике 8 класс Тема: «Тепловые явления» infourokru/kontrolnaya-rabota-po-fizike-klass Cached Все материалы
контрольная работа по физике 10 класса тепловые явления — Все результаты Контрольная работа по физике на тему: «Тепловые явления» Похожие 3 окт 2014 г — Контрольная работа ориентирована на учебник АВ Перышкина и содержит 12 вопросов Контрольная работа по физике на тему: » Тепловые явления » Контрольная работа ориентирована Физика 10 класс Контрольная работа по физике на тему «Тепловые явления» 10 › Физика Похожие 3 янв 2016 г — Контрольная работа по теме « Тепловые явления » 10 класс Вариант 1 1 В латунный калориметр массой 128г налили 230г воды при Контрольная работа по физике по теме «Тепловые явления» › Физика 26 нояб 2017 г — Контрольная работа № 1 по теме « Тепловые явления » Вариант 1 1 ( Удельная теплота сгорания пороха 3,8 * 10 6 Дж/кг) 3 Задания №4 к конспекту урока по физике в 8 классе «Энергия топлива» 26112017 Контрольная работа Тепловые явления 8 класс — Инфоурок › Физика Похожие Cкачать: Контрольная работа Тепловые явления 8 класс Вход или войдите через: Забыли пароль? Инфоурок › Физика › Другие методич материалы › Контрольная работа Тепловые явления 8 класс hello_html_m10f1262d gif Контрольные работы по физике 10 класс — Инфоурок › Физика Похожие 27 апр 2016 г — 885 КБ физика 10 кр №2 тепловые явления doc Выбранный для Итоговая контрольная работа по физике 10 класс Вариант 1 Картинки по запросу контрольная работа по физике 10 класса тепловые явления «id»:»V8DRfjZvERygzM:»,»ml»:»600″:»bh»:90,»bw»:60,»oh»:307,»ou»:» «,»ow»:200,»pt»:»ozon-stcdnngenixnet/multimedia/1004490061jpg»,»rh»:»ozonru»,»rid»:»uWPE6AqMPF9KWM»,»rt»:0,»ru»:» «,»sc»:1,»st»:»Ozon»,»th»:103,»tu»:» \u003dtbn:ANd9GcSz9b8oBboaqqLQRaptw7R4AKmD-i2_F4PfLn_B0SumnJKUbP2iGD1SJb8″,»tw»:67 «cl»:3,»cr»:3,»id»:»Ye6GG2Cy2WZfIM:»,»ml»:»600″:»bh»:90,»bw»:79,»oh»:541,»ou»:» «,»ow»:468,»pt»:»ds03infourokru/uploads/ex/0f8e/0000369d-d953d0a1″,»rh»:»infourokru»,»rid»:»N0z1zKODlKD0oM»,»rt»:0,»ru»:» «,»sc»:1,»st»:»Инфоурок»,»th»:91,»tu»:» \u003dtbn:ANd9GcSTiWgbMCuNt6l8UhNiuRPO5eqrsBfQuQMn7AfTYT0cxjW_yfKkQYxrHQ»,»tw»:79 «cl»:3,»cr»:3,»id»:»EJrMT-OcEBkuUM:»,»ml»:»600″:»bh»:90,»bw»:69,»oh»:677,»ou»:» «,»ow»:515,»pt»:»cdn2arhivurokovru/multiurok/html/2018/09/27/s_5b»,»rh»:»multiurokru»,»rid»:»W_2qOuSVCfcdpM»,»rt»:0,»ru»:» «,»sc»:1,»st»:»Мультиурок»,»th»:96,»tu»:» \u003dtbn:ANd9GcQzKlLUg5kJeDnxdgVmp79BYZ6vdiXoweRCvUYY0xggVwYpiWCvSsVdhg»,»tw»:73 «cl»:3,»id»:»qoosv1kyEtjM3M:»,»ml»:»600″:»bh»:90,»bw»:74,»oh»:581,»ou»:» «,»ow»:470,»pt»:»ds03infourokru/uploads/ex/0f8e/0000369d-d953d0a1″,»rh»:»infourokru»,»rid»:»N0z1zKODlKD0oM»,»rt»:0,»ru»:» «,»sc»:1,»st»:»Инфоурок»,»th»:93,»tu»:» \u003dtbn:ANd9GcQeyAzPEKKICUi5wJwJFKLYPZ2RGCnlMwCiPh0Osx4CyMiodEDD1d5sOaU»,»tw»:75 «cr»:3,»id»:»jDEM69M5uiroyM:»,»ml»:»600″:»bh»:90,»bw»:67,»oh»:590,»ou»:» «,»ow»:434,»pt»:»ds03infourokru/uploads/ex/0f8e/0000369d-d953d0a1″,»rh»:»infourokru»,»rid»:»N0z1zKODlKD0oM»,»rt»:0,»ru»:» «,»sc»:1,»st»:»Инфоурок»,»th»:97,»tu»:» \u003dtbn:ANd9GcTw8GjcOqrZaWYaNPg32y7JyYG4w8RRM4NuWCLag9Hl34LEqUIl-vZArg»,»tw»:72 «cb»:3,»id»:»7Ih9pVAAiWJJTM:»,»ml»:»600″:»bh»:90,»bw»:77,»oh»:577,»ou»:» «,»ow»:474,»pt»:»ds03infourokru/uploads/ex/0f8e/0000369d-d953d0a1″,»rh»:»infourokru»,»rid»:»N0z1zKODlKD0oM»,»rt»:0,»ru»:» «,»sc»:1,»st»:»Инфоурок»,»th»:94,»tu»:» \u003dtbn:ANd9GcQTn1HJSr6wyTyB5Uv24wX_eKRY7jBoWuWdHWq4NgBOOSMXDM24o0NWrIE»,»tw»:77 «cl»:3,»cr»:3,»id»:»qpCJEt2Ky9E02M:»,»ml»:»600″:»bh»:90,»bw»:68,»oh»:605,»ou»:» «,»ow»:450,»pt»:»ds03infourokru/uploads/ex/0f8e/0000369d-d953d0a1″,»rh»:»infourokru»,»rid»:»N0z1zKODlKD0oM»,»rt»:0,»ru»:» «,»sc»:1,»st»:»Инфоурок»,»th»:97,»tu»:» \u003dtbn:ANd9GcRlc7tmTUHrznRDPoV2k2KbWhqQWgBkkJT6frVFoTvp704pKvbGhO7wEQ»,»tw»:72 «cl»:3,»cr»:3,»id»:»4pmKzQHZfQYViM:»,»ml»:»600″:»bh»:90,»bw»:78,»oh»:634,»ou»:» «,»ow»:550,»pt»:»cdn2arhivurokovru/multiurok/html/2018/09/27/s_5b»,»rh»:»multiurokru»,»rid»:»W_2qOuSVCfcdpM»,»rt»:0,»ru»:» «,»sc»:1,»st»:»Мультиурок»,»th»:90,»tu»:» \u003dtbn:ANd9GcRdTfFxXBqh9Z-iyBPgC05gtBWCeFelrGQyVnzzb9ekcqMxFP_gwtpylA»,»tw»:78 Другие картинки по запросу «контрольная работа по физике 10 класса тепловые явления» Жалоба отправлена Пожаловаться на картинки Благодарим за замечания Пожаловаться на другую картинку Пожаловаться на содержание картинки Отмена Пожаловаться Все результаты Контрольная работа «Тепловые явления» 10 класс скачать uchitelyacom/fizika/84898-kontrolnaya-rabota-teplovye-yavleniya-10-klasshtml Контрольная работа №2 по теме « Тепловые явления» Физика 10 Вариант 1 Часть А 1 Теплообмен путем конвекции может осуществляться 1) в газах Контрольная работа по физике 10 класс по итогам 3 четверти Похожие 25 мар 2016 г — Контрольная работа по физике 10 класс по итогам 3 четверти и превращения энергии, распространенный на тепловые явления , Контрольная работа по теме «Тепловые явления» — физика, тесты Похожие 29 дек 2015 г — Контрольная работа № 1 по теме « Тепловые явления » каменного угля 30 * 10 6 Дж/кг) Электронная тетрадь по физике 7 класс Задания к контрольной работе по разделу » Молекулярная физика Похожие 10 июн 2014 г — Тепловые явленияЗадания к контрольной работе по разделу физика Тепловые явления Электронная тетрадь по физике 10 класс Проверочная работа по физике, 10 класс — Урокрф 30 июн 2017 г — Контрольные / проверочные работы для учителя-предметника для 10 класса по ФГОС Учебно-дидактические материалы по Физике для 10 класса по УМК Мякишев ТЯ, Буховцев ББ и др Тепловые явления » Контрольная работа по физике для 8 класса по теме «Тепловые 21 нояб 2017 г — Контрольная работа по физике для 8 класса по теме » Тепловые явления « Тепловые явления » 8 класс 1) 18,4· 10 7 Дж 2) 20 · 10 7 Дж Контрольная работа по физике по теме «Тепловые явления» 8 класс 22 мар 2018 г — Контрольная работа № 1 по теме « Тепловые явления »-8 класс -В2 1 Какое количество теплоты необходимо для нагревания от 10ºC до [DOC] Аналитический отчёт о результатах региональной контрольной wwweduportal44ru/Neya/2-new/SiteAssets//физика%208кл,%200612%2016doc Контрольная работа по физике проводилась Департаментом образования и 8 класса содержания темы « Тепловые явления » по предмету « физика » Контрольная работа состоит из 10 заданий: 8 заданий базового уровня, Рабочая программа по физике 10 класс базовый уровень В результате изучения физики в 10 классе на базовом уровне ученик должен тест: (текущий, итоговый), контрольная работа , устный фронтальный опрос, опрос Почему тепловые явления изучаются в молекулярной физике ? Подготовка к контрольной работе по теме «Тепловые явления» 8 23 нояб 2016 г — Ресурс содержит 2 варианта с типовыми заданиями из контрольно- измерительных материалов к уч Перышкина Физика 8 класс (автор [DOC] Тематическое планирование — Школа № 1363 sch2363uvmskobrru/files/rabochaya_programma_10_klassydoc Похожие Настоящая рабочая программа курса физики для 10 класса составлена на Тепловые явления Контрольная работа по решению уровневых задач ▷ физика контрольная работа по теме тепловое явление 8 класс lechurchillcom//fizika-kontrolnaia-rabota-po-teme-teplovoe-iavlenie-8-klassxml 6 дней назад — Контрольная работа по теме » Тепловые явления » 8 класс нас» по физике 10 класс Контрольная работа по физике » Тепловые явления » Контрольная работа по физике по теме тепловые явления ответы Контрольная работа по физике по теме тепловые явления ответы 8 класс Контрольная работа 1 «Расчёт количества теплоты» 1 вариант Задания 1-5 для Дидактическое пособие по теме «Термодинамика» учени 10 класса [PDF] физика общеобразовательной подготовки по физике обучающихся 10 класса 2 Документы, определяющие содержание контрольной работы Содержание контрольной работы 2) Тепловые явления 3) Электромагнитные явления Физика — 8 класс Итоговая контрольная работа по теме «Тепловые Похожие 1 Вариант 1 Длительность: 10 минут Физика Физика 9 класс Контрольная работа по теме «Законы движения тел» Физика Физика Сети ЭВМ [PDF] Физика 10 класс — Образовательный Портал УР ciurru/vot//Рабочие%20программы%2010%20класс/Физика%2010%20классpdf (10 класс Мякишев ГЯ) В курсе физики 10 класса продолжается дальнейшее изучение физики на второй ступени Тепловые явления », « Основы электродинамики» -Марон АЕ Контрольные работы по физике 10 -11 класс [PDF] Приложение к рабочей программе по физике для 10 класса Похожие Время: 45 минут Источник: Марон АЕ Физика 10 класс : дидактические материалы / АЕМарон, ЕАМарон Громцева ОИ Тематические контрольные и самостоятельные работы по физике 10 физика Тепловые явления » Контрольная работа 8 класс физика тепловые явления » вопрос wwwadvokatru/?doc=kontrolnaya-rabota-8-klass-fizika-teplovie-yavleniya Контрольная работа по теме » Тепловые явления » 8 класс Стальная деталь массой 500 г при Контрольная работа физика 10 класс кинематика Контрольные работы по физике — Физика и Астрономия Контрольные, лабораторные и практические работы по физике для проверки уровня знаний учащихся Контрольная работа по теме » Тепловые явления » Контрольная работа к уроку физики «Законы сохранения» 10 класс ответы по контрольной работе по физике — KiteClass kiteclassru/files/otvety-po-kontrolnoi-rabote-po-fizike-molekuliarnaia-fizikaxml 7 нояб 2018 г — физика Тепловые явления » Контрольная работа по теме Молекулярная физика 10 класс pedportalnet/starshie-klassy/fizika/kontrolnaya контрольная работа по физике тепловые явления ответы скачать roskinoorg//kontrolnaia-rabota-po-fizike-teplovye-iavleniia-otvety-skachatxml 3 нояб 2018 г — контрольная работа по физике тепловые явления ответы скачать явления 8 класс testschoolru/2017/ 10 /08/kontrolnaya-rabota-po [PDF] Физика 10-11 класс — skole31ru skole31ru/documents/RP/prof/10-11_fiz_3hpdf Рабочая программа по физике для 10 класса составлена на основе Примерной программы Тепловые явления 3 Основы 10 класс : количество часов – 102 ч, лабораторные работы – 5 часов; контрольные работы – 7 часов Рабочая программа по физике 10 класс по учебнику ГЯ Мякишева 4862 КБ Рабочая программа по физике для 10 класса docx Тепловые явления 13 1 Контрольная работа № 1 « Кинематика материальной точки» Молекулярная физика Тепловые явления 10 Класс – скачать Тепловые явления Тип урока урок обобщения и систематизации знаний Форма Контрольная работа по теме » Класс Птицы» 1 вариант I Выберите [DOC] Учебно-тематическое планирование s147kazanedusiteru/DswMedia/fizika10-12-doc Кирик ЛА « Контрольные и самостоятельные работы по физике 10 Контрольные задания Физика 10 класс , Саратов: Лицей 2014г применять полученные знания по физике для объяснения разнообразных физических явлений и свойств веществ; Тепловые двигатели и охрана окружающей среды Книга Физика Самостоятельные и контрольные работы Физика Самостоятельные и контрольные работы Молекулярная физика Тепловые явления Электричество Магнетизм 10 класс Физика ФИЗИКА 10 КЛАСС — КОНТРОЛЬНЫЕ РАБОТЫ В НОВОМ zvonoknaurokru/load/kontrolnye_raboty_v_novom_formate/fizika_10/198 Похожие В разделе размещены контрольные работы по всем темам традиционного курса физики 10 класса ФИЗИКА 10 КЛАСС | Просмотров: 6340 | Загрузок: 0 [DOC] 11 класс Физика изучается на базовом уровне рассмотрения явлений природы, знакомства с основными законами Контрольные и проверочные работы по физике 7-11 классы Тепловые явления -23ч Рабочая программа учебного курса по физике в 10 класса составлена на основе Примерной программы «РЕШУ ОГЭ»: физика ОГЭ — 2019: задания, ответы, решения Похожие По окончании работы система проверит ваши ответы, покажет правильные 10 Тепловые явления (расчетная задача) просмотреть (52 шт) 11 ФИЗИКА 10 КЛАСС — Муниципальное казённое klevakinorusedunet/post/1569/90459 ФИЗИКА 10 КЛАСС -показать специфику тепловых явлений и познакомить учащихся со Контрольные и проверочные работы по физике Контрольная работа по физике Тепловые явления 8 класс 8 окт 2017 г — Контрольная работа по физике Тепловые явления 8 класс Контрольная Удельная теплота плавления белого чугуна 14 · 10 4 Дж/кг Уроки физики — Официальный сайт МОУ Тереньгульской СОШ tershkolaucozru/index/uroki_fiziki/0-68 Похожие На странице «Уроки физики » размещены конспекты и презентации уроков, внеклассных Контрольная работа « Тепловые явления » 8 класс засекина физика 10 класс учебник ZV WS活动温哥华教育网 vancouvereducom/thread-3792-1-1html 20 июл 2018 г — Н Н Сотский ) 2010 10 класс : ГДЗ и Решебник по физике для 10 ( 8 класс ) Контрольная работа № 1 по теме Тепловые явления Тесты по физике (8 класс), итоговые контрольные работы за › Тесты › Физика Тепловые явления Тест Температура по физике (8 класс ) 10 вопросов Уровень: новичок Тест Тепловое явление по физике (8 класс ) 10 вопросов контрольная работа по физике за 8 класс углубленным изучением profcareerru//kontrolnaia-rabota-po-fizike-za-8-klass-uglublennym-izucheniemxm контрольная работа по физике за 8 класс углубленным изучением языка при Контрольная работа по физике на тему » Тепловые явления Похожие 10 контрольные работа по физике 8 класс по теми тепловые явления wwwcsaladihaztervezeshu//kontrolnye-rabota-po-fizike-8-klass-po-temi-teplovye-i контрольные работа по физике 8 класс по теми тепловые явления в 8 -ых классах Контрольная работа по физике 10 класс Молекулярная физика [PDF] Физика 10 класс — Гимназия №2 г Владивостока wwwvladgym2ru/attachments/888_Физика%2010%20классpdf Похожие Рабочая программа по физике 10 – го класса составлена на основе федерального компонента тепловые явления , основы электродинамики Так как данная 33/10 Контрольная работа № 3 «Законы сохранения в механике» Обобщающий урок по физике в 8-м классе по теме «Тепловые открытыйурокрф/статьи/564695/ Обобщающий урок по физике в 8-м классе по теме » Тепловые явления » подводим курсор к рисункам, открываем файлы (t 11avi, t 10 avi, t 8avi), выключаем звук Подготовка к контрольной работе по теме « Тепловые явления » Физика Самостоятельные и контрольные работы Молекулярная fff-librru/obrazovanie-i/16158-fizika-samostojatelnye-i-kontrolnye-rabotyhtml 10 сент 2018 г — Физика Самостоятельные и контрольные работы Молекулярная физика Тепловые явления Электричество Магнетизм 10 класс , Л А Кирик, Леонид Анатольевич irbisbigpibiyskru/cgi/cgiirbis_64exe? К 43 Кирик, Леонид Анатольевич Физика [Текст] : 10 класс : разноуровневые самостоятельные и контрольные работы / Л А Кирик — М : ИЛЕКСА, 2002 Физика 8 класс — InternetUrok Видеоуроки, тесты и тренажёры по Физика за 8 класс по школьной программе Используйте Варианты контрольной работы » Электрические явления » Учебно-методические материалы по физике — Средняя школа при stockholmmoscow/externat/physicshtml Похожие Контрольная работа № 1 Самостоятельные работы по теме » Тепловые явления » 1 Учебно-методические материалы по физике для 10 класса [PDF] РАБОЧАЯ ПРОГРАММА ПО ФИЗИКЕ ( 10 класс) НА 2013 – 2014 in2kru/wp-content/uploads/2014/01/Рабочая-программа-по-физике-10-классpdf Похожие 30 авг 2013 г — Рабочая программа по физике для 10 класса составлена на основе Фронтальные лабораторные работы по физике в 7-11 классах Тепловые явления ; Контрольная работа № 1 «Кинематика точки» ▷ контрольная работа по физике 8 класс тепловые явления с wwwdesignbateriecz//kontrolnaia-rabota-po-fizike-8-klass-teplovye-iavleniia-s-res контрольная работа по физике 8 класс тепловые явления с решением физике для 8 -го класса разработана на основе 10 / 10 , Решение задач по теме Вместе с контрольная работа по физике 10 класса тепловые явления часто ищут контрольная работа по физике 8 класс тепловые явления перышкин ответы контрольная работа по физике 8 класс тепловые явления тест контрольная работа по физике 8 класс тепловые явления количество теплоты контрольная работа по физике 8 класс тепловые явления задачи контрольная работа по физике 8 класс ответы итоговая контрольная работа по физике 8 класс тепловые явления ответы на контрольную работу по физике 8 класс световые явления контрольная работа 1 по физике 8 класс Навигация по страницам 1 2 3 Следующая Ссылки в нижнем колонтитуле Россия — Подробнее… Справка Отправить отзыв Конфиденциальность Условия Аккаунт Поиск Карты YouTube Play Новости Почта Контакты Диск Календарь Google+ Переводчик Фото Ещё Документы Blogger Hangouts Google Keep Подборки Другие сервисы Google
Яндекс Яндекс Найти Поиск Поиск Картинки Видео Карты Маркет Новости ТВ онлайн Музыка Переводчик Диск Почта Коллекции Все Ещё Дополнительная информация о запросе Показаны результаты для Нижнего Новгорода Москва 1 Контрольная работа по физике на тему » Тепловые » infourokru › kontrolnaya-rabota…fizike…yavleniya… Сохранённая копия Показать ещё с сайта Пожаловаться Информация о сайте Сайт – выбор пользователей Подробнее о сайте Контрольная работа по теме « Тепловые явления » 10 класс Вариант 1 1 В латунный калориметр массой 128г налили 230г воды при температуре 8,40 С и опустили кусок металла массой 192г нагретого до 1000 С В калориметре установилась температура 21,50 С, Определить удельную Читать ещё Контрольная работа по теме « Тепловые явления » 10 класс Вариант 1 1 В латунный калориметр массой 128г налили 230г воды при температуре 8,40 С и опустили кусок металла массой 192г нагретого до 1000 С В калориметре установилась температура 21,50 С, Определить удельную теплоемкость металла 2 Газ, занимающий объем 22дм3 под давлением 105 Па, был изобарно нагрет от 200С до 1000 С Определить работу расширения газа 3 Температура нагревателя тепловой машины 1500 С, а холодильника 250 С Машина получила от нагревателя 4· 104 Дж энергии Как велика работа , произведенная машиной? Вариант 2 Скрыть 2 Контрольная работа по теме Молекулярная физика 10 testschoolru › 2018/01/12/kontrolnaya-rabota-po…10… Сохранённая копия Показать ещё с сайта Пожаловаться Информация о сайте Контрольная работа состоит из 5 вариантов, в каждом по 8 заданий «Частицы вещества участвуют в непрерывном тепловом хаотическом движении» Ответы на контрольную работу по теме Молекулярная физика 10 класс 1 вариант А1-3 А2-2 А3-1 А4-3 А5-1 В1 20 моль В2 123 С1 Читать ещё Контрольная работа состоит из 5 вариантов, в каждом по 8 заданий «Частицы вещества участвуют в непрерывном тепловом хаотическом движении» Это положение молекулярно-кинетической теории строения вещества относится к 1) газам 2) жидкостям 3) газам и жидкостям 4) газам, жидкостям и твердым телам А2 Как изменится давление идеального одноатомного газа при увеличении средней кинетической энергии теплового движения его молекул в 2 раза и уменьшении концентрации молекул в 2 раза? Ответы на контрольную работу по теме Молекулярная физика 10 класс 1 вариант А1-3 А2-2 А3-1 А4-3 А5-1 В1 20 моль В2 123 С1 Скрыть 3 Контрольная работа » Тепловые явления » 10 класс uchitelyacom › Физика › …-teplovye-yavleniya-10… Сохранённая копия Показать ещё с сайта Пожаловаться Информация о сайте Контрольная работа № 2 по теме « Тепловые явления » Физика 10 Вариант 1 Часть А 1 Теплообмен путем конвекции может осуществляться Читать ещё Контрольная работа № 2 по теме « Тепловые явления » Физика 10 Вариант 1 Часть А 1 Теплообмен путем конвекции может осуществляться 1) в газах, жидкостях и твердых телах 2) в газах и жидкостях 3) только в газах 4) только в жидкостях Скрыть 4 Тест по физике ( 10 класс ) на тему: Тест по теме quot nsportalru › shkola…test-po…teplovye-yavleniya-10… Сохранённая копия Показать ещё с сайта Пожаловаться Информация о сайте Сайт – выбор пользователей Подробнее о сайте Данный тест по теме » Тепловые явления » предназначен для контроля знаний, умений, навыков и соответсвует кодификатору единого государственного экзамена по физике Читать ещё Данный тест по теме » Тепловые явления » предназначен для контроля знаний, умений, навыков и соответсвует кодификатору единого государственного экзамена по физике Данный тест по теме » Тепловые явления » предназначен для контроля знаний, умений, навыков и соответсвует кодификатору единого государственного экзамена по физике Скачать: Вложение Размер test_ teplovye _yavleniyadoc 29 КБ Предварительный просмотр: Тест по теме: « Тепловые явления » 10 класс Тест А1 Скрыть 5 Задания и ответы к контрольной работе по разделу kopilkaurokovru › Физика › …/zadaniia-i-otviety-k… Сохранённая копия Показать ещё с сайта Пожаловаться Информация о сайте Задания к контрольной работе по разделу » Молекулярная физика Тепловые явления » Вариант 1 2Рассчитайте температуру, при которой средняя кинетическая энергия поступательного движения молекул равна 10 ,35× 10 -21 Дж Читать ещё Задания к контрольной работе по разделу » Молекулярная физика Тепловые явления » Вариант 1 1Какова масса кислорода, содержащегося в баллоне объемом 50л при температуре 27 градусов цельсия и давлении 2×106 Па? 2Рассчитайте температуру, при которой средняя кинетическая энергия поступательного движения молекул равна 10 ,35× 10 -21 Дж 3Определите КПД теплового двигателя, если количество теплоты,полученное от нагревателя за цикл, равно 500Дж, а колличество теплоты, отданное холодильнику, составляет 400 Дж 4Газовый процесс изображен в координатах (P,V) Постройте этот же процесс в коо Скрыть 6 Проверочная работа по физике , 10 класс урокрф › library…rabota_po_fizike_105453html Сохранённая копия Показать ещё с сайта Пожаловаться Информация о сайте Контрольные / проверочные работы для учителя-предметника для 10 класса по ФГОС Проверочная работа по физике Тема: «Молекулярная физика Тепловые явления » Автор разработки Читать ещё Контрольные / проверочные работы для учителя-предметника для 10 класса по ФГОС Учебно-дидактические материалы по Физике для 10 класса по УМК Мякишев ТЯ, Буховцев ББ и др / Под ред Парфентьевой НА Проверочная работа по физике Тема: «Молекулярная физика Тепловые явления » Автор разработки: Духовникова Галина Анатольевна преподаватель физики Байкальск – 2017г Проверочная работа по физике Тест по теме: «Молекулярная физика и термодинамика» Пояснительная записка Тест по теме «Молекулярная физика и термодинамика» проводится для студентов СПО 1курса Скрыть 7 Тест Тест по теме «Молекулярная физика Тепловые » videourokinet › Онлайн-тесты › tiest-po-tiemie… Сохранённая копия Показать ещё с сайта Пожаловаться Информация о сайте Тест по разделу «Молекулярная физика Тест по разделу «Молекулярная физика Тепловые явления » для студентов 1 курса по специальности «Техническое обслуживание и ремонт автомобильного транспорта» Тесты по физике 10 класс 0 831 Читать ещё Тест по разделу «Молекулярная физика Тепловые явления » для студентов 1 курса по специальности «Техническое обслуживание и ремонт автомобильного транспорта» Тест по разделу «Молекулярная физика Тепловые явления » для студентов 1 курса по специальности «Техническое обслуживание и ремонт автомобильного транспорта» Система оценки: 5 балльная Список вопросов теста Тесты по физике 10 класс 0 831 Скрыть 8 Контрольная работа по физике 10 класса тепловые явления — смотрите картинки ЯндексКартинки › контрольная работа по физике 10 класса тепловые Пожаловаться Информация о сайте Ещё картинки 9 Тесты : Молекулярная физика Тепловые явления testoniknet › bank/fizika/10…teplovye-yavleniya/2 Сохранённая копия Показать ещё с сайта Пожаловаться Информация о сайте Тепловые явления ( Физика , 10 класс ) от профессиональных педагогов Тестоник — незаменимый помошник учителя! Тепловые явления ( Физика , 10 класс ) Предмет 10 Контрольная работа № 1 по теме » Тепловые явления » multiurokru › Обо мне › kontrol-naia-rabota-1-po… Сохранённая копия Показать ещё с сайта Пожаловаться Информация о сайте Просмотр содержимого документа « Контрольная работа № 1 по теме » Тепловые явления «» Ответ обоснуйте Температура свинцовой пластинки размером 10x5x2 см уменьшается от 300 до 100ºC Электронная тетрадь по физике 10 класс Подготовка к ЕГЭ по физике Читать ещё Просмотр содержимого документа « Контрольная работа № 1 по теме » Тепловые явления «» Контрольная работа № 1 по теме « Тепловые явления »-8 класс-В1 Рассчитайте количество теплоты, необходимое для нагревания железного утюга массой 1кг для изменения его температуры на 150ºC Удельная теплоемкость железа 460 Дж/(кгºC) Ответ обоснуйте Температура свинцовой пластинки размером 10x5x2 см уменьшается от 300 до 100ºC Какое количество теплоты при этом передает пластинка окружающим телам? Удельная теплоемкость свинца 140 Дж/(кгºC), его плотность 11300 кг/м³ Электронная тетрадь по физике 10 класс Подготовка к ЕГЭ по физике Скрыть Тест по физике на тему » Тепловые явления » (8, 10 класс ) moeobrazovanieru › Тесты по темам › 10 класс Сохранённая копия Показать ещё с сайта Пожаловаться Информация о сайте Онлайн тесты для учащихся 8, 10 класса Какой водой заливают каток? Что такое туман? На эти вопросы знает ответы физика Читать ещё Онлайн тесты для учащихся 8, 10 класса Бесплатные тесты для школьников Тесты ЕГЭ, ГИА Какой водой заливают каток? Что такое туман? На эти вопросы знает ответы физика Создатель:МыхинаГалинаВладимировна, учитель физики Начать тест Скрыть Контрольная работа Физика – Поможем написать! Курсовые работы Дипломные работы Рефераты Все работы абв-консультациирф › платные-услуги Не подходит по запросу Спам или мошенничество Мешает видеть результаты Информация о сайте реклама От 800р для Вуза! Контрольная работа Физика Контактная информация +7 (920) 158-01-26 круглосуточно 18+ Вместе с « контрольная работа по физике 10 класса тепловые явления » ищут: итоговая контрольная работа по физике 10 класс контрольная работа по физике 11 класс контрольные работы по физике 10 класс мякишев с решениями контрольная работа по физике 9 класс входная контрольная работа по физике 10 класс годовая контрольная работа по физике 10 класс при какой температуре средняя кинетическая энергия поступательного контрольная работа 1 по физике 10 класс входная контрольная работа по физике 10 класс с ответами сборник контрольных работ по физике 10 класс 1 2 3 4 5 дальше Браузер Для безопасных прогулок в сети 0+ Установить
Анализ крошки | Процессы выпечки
Происхождение
Традиционная сенсорная оценка хлеба основывается на опросах людей, которые требуют много времени, дороги и имеют субъективный характер подсчета баллов. С развитием фотографии и анализа компьютерного зрения крупные пекарни постепенно перешли к более объективным методам описания текстурных свойств хлебобулочных изделий.
Самая ранняя опубликованная работа по анализу мякиша с использованием техники компьютерного зрения была опубликована в 1992 году Бертраном и его сотрудниками, которые коррелировали размер и форму ячеек мякиша с текстурными свойствами хлеба.Их исследование выявило двухфазные материалы в крошке, а именно твердую фазу (клеточная стенка) и воздушная фаза (воздух). 3 Сегодня одним из самых надежных инструментов визуализации для анализа крошки, который стал доступным для пекарен в начале 2000-х годов, является система C-Cell. Он возник в Великобритании и быстро был принят пекарнями в Европе, США и других промышленно развитых странах.
Как это работает?
Как важный инструмент в программах качества и НИОКР в крупных пекарнях, анализ крошки используется для выявления возможных несоответствий в ингредиентах или процессе приготовления хлеба и других хлебобулочных изделий.Поскольку этот анализ не поддается количественной оценке человеческим глазом, он выполняется с использованием методов анализа изображений.
Равномерно нарезанный хлеб или пирожные фотографируют с помощью подходящей камеры с соответствующим разрешением и при надлежащем освещении. Захваченные изображения оцифровываются для последующей обработки. Критические атрибуты мякиша, такие как размер ячеек, структура, цвет, однородность и т. Д., Оцениваются с помощью компьютерных программ на основе предварительно выбранных алгоритмов и сравниваются со стандартными оценками мякиша. Вот примеры того, как выглядят изображения куска хлеба и булочки для гамбургера:
Метод AACC 10-18.01 для оценки структуры мякиша хлебобулочных изделий полагается на анализатор хлебобулочных изделий C-Cell. Доступны две модели, C-Cell Mono и C-Cell Color, обе работают на одной программной платформе. Модели Mono и Color измеряют 48 различных параметров, таких как: размеры, форма, размер ячейки, удлинение и ориентация. Модель Color имеет дополнительную возможность измерения цвета и толщины мякиша / корочки, а также специальные функции, например, определение количества вишен в вишневом пироге или семян кунжута в булочке для гамбургера. 3
Вот выдержка из образца отчета:
Приложение
Хлебобулочные изделия имеют характерную форму и определенную текстуру, ожидаемую потребителем. Любое существенное отклонение от оптимальных характеристик продукта часто считается дефектом качества. Текстурные и сенсорные свойства хлебобулочных изделий определяются трехмерной структурой мякиша, которая создается микроскопическими и макроскопическими компонентами формулы.
Анализ изображений был сосредоточен на следующих вариациях рецептур хлебобулочных изделий:
- Определение влияния сорта пшеницы на хлебопекарные качества. Вообще говоря, сорта пшеницы с сильным содержанием глютена (индекс глютена, GI> 90) имеют тенденцию показывать неоднородные и крупные зерна крошки, в то время как более мелкие и более однородные зерна крошки связаны с сортами с оптимальной концентрацией глютена (индекс клейковины, GI = 60-90 ). Соответствующие испытания на пенетрацию подтвердили, что неоднородные и крупные зерна крошки имели более высокие значения пенетрации по сравнению с однородными и более мелкими зернами крошки. 4
- Возможность различать компоненты пшеницы и не пшеницы в хлебе, а также измельченные фракции твердой и мягкой пшеницы.
- Корреляция влияния эмульгаторов и кондиционеров для теста (DATEM и SSL, лецитин, фосфатидилхолин, сложные эфиры сахарозы) на общие сенсорные качества хлеба.
Общие замечания и рекомендации по анализу крошки:
- Чем больше количество ячеек и чем выше средняя площадь ячеек, тем лучше пористость хлебной крошки.
- Тонкие клеточные стенки преобладают в крошке с мелкой текстурой, в то время как толстые стенки преобладают в крупной крошке.
- Промышленные багеты имеют более высокую пористость, чем более плотные цельнозерновые и традиционные багеты.
- Хлеб, приготовленный с использованием слишком большого количества воды или слишком малого количества соли, обычно имеет очень пористую структуру с крупными ячейками, как у швейцарского сыра.
- Анализ крошки может быть полезен при обнаружении потенциальных повреждений хлебобулочных изделий и может помочь в сокращении отходов.
Список литературы
- Каувен, С.П.. Система анализа изображений C-Cell для объективного измерения качества хлеба. Материалы симпозиума ICC-SA / SACB по хлебу и злакам, Йоханнесбург, Южная Африка, 2004 г., стр: 396-402.
- Whitworth, M., Cauvain, S.P. и Cliffe, D. Измерение структуры хлебных клеток с помощью анализа изображений. В книге «Использование науки и технологий в зерновых культурах на благо потребителей», Труды 12-го Международного конгресса ICC по зерновым и хлебу, 23-26 мая 2004 г., Харрогейт, Великобритания, Woodhead Publishing Ltd, Кембридж (ред.С.П. Каувен, С.Э. Салмон и Л.С. Янг), 2005, стр: 193-198.
- Метод AACC 10-18-01. Измерение структуры мякиша хлебобулочных изделий с помощью C-Cell. Качество выпечки. Утвержденные AACC методы анализа, 11-е издание.
- Магдич, Д., Хорват, Д., Дрезнер, Г., Юркович, З. и Шимич, Г. Анализ изображений структуры хлебных крошек в зависимости от прочности клейковины пшеницы. Poljoprivreda, 2006, 12, 1, стр: 58-62.
9 советов по упрощению навигации по сайту [+ примеры]
В сказке Гензель и Гретель двое детей бросают в лесу панировочные сухари, чтобы найти дорогу домой.В настоящее время вы, вероятно, не испытываете слишком много заблудших ситуаций, но я готов поспорить, что вы чувствовали себя дезориентированными на плохо спроектированном веб-сайте.
В веб-дизайне навигация по хлебным крошкам — это способ показать пользователям их местоположение и то, как они туда попали. Подобно хлебным крошкам Гензеля, он помогает пользователям проследить свой путь и увидеть, где они находятся в общей схеме вашего сайта.
Что такое навигация по хлебным крошкам?
Навигация по хлебным крошкам — это способ для пользователей визуализировать свое местоположение на веб-сайте.Это дополнительная панель навигации, которая обычно отображается в виде горизонтальных текстовых ссылок, разделенных угловой скобкой. Навигация с помощью хлебных крошек улучшает поиск ваших целевых страниц и помогает пользователям быстрее переходить на страницы более высокого уровня, если они изначально нашли ваш сайт из поиска или по глубокой ссылке.
Хотя вы можете оформлять хлебные крошки, как хотите, они, как правило, выглядят одинаково на всех веб-сайтах, которые их используют. Вот простой пример панировочных сухарей из базы знаний HubSpot:
Якоб Нильсен, соучредитель известной Nielsen Norman Group, рекомендует навигацию по хлебным крошкам с 1995 года и подчеркивает их полезность и эффективность: «Все, что делают хлебные крошки, — это упрощает пользователям перемещение по сайту, при условии, что его содержание и общая структура имеют смысл.Этого достаточно для того, что занимает всего одну строчку в дизайне ».
Если веб-сайт вашей компании является многоуровневым, вы можете подумать о реализации хлебных крошек, чтобы упростить навигацию по сайту. Однако, как и любой элемент дизайна, есть правильный и неправильный способ сделать это. Здесь мы рассмотрим девять советов и примеров, чтобы убедиться, что вы создаете наиболее эффективную навигацию по хлебным крошкам для своих пользователей.
Советы и примеры навигации по хлебным крошкам1.Используйте навигацию с помощью хлебных крошек, только если это имеет смысл для структуры вашего сайта.
Навигация по хлебным крошкам имеет линейную структуру, поэтому вы можете использовать ее только в том случае, если это имеет смысл с иерархией вашего веб-сайта. Если у вас есть страницы нижнего уровня, которые доступны с разных целевых страниц, использование хлебных крошек только запутает читателей, которые продолжают получать доступ к одним и тем же страницам с разных начальных точек. Кроме того, если ваш сайт относительно простой, всего с несколькими страницами, вам, вероятно, не нужна навигация по хлебным крошкам.
2. Не делайте навигацию слишком большой.
Навигация в виде хлебных крошек является второстепенным инструментом по сравнению с основной панелью навигации, поэтому она не должна быть слишком большой или заметной на странице. Например, на веб-сайте DHL их основная панель навигации большая и узнаваемая, с такими столбцами, как «Экспресс», «Посылки и электронная коммерция», «Логистика» и т. Д. Их навигационная цепочка — это меньший раздел ниже, который гласит: «DHL Global |> Логистика» |> Грузовые перевозки ».Вы же не хотите, чтобы пользователи ошибочно принимали вашу навигацию по хлебным крошкам за основную панель навигации.
3. Включите полный навигационный путь в свою навигационную цепочку.
Я погуглил «Неграмотные студенты Университета Илона», чтобы попасть на эту целевую страницу, но Илон умен, включив полный путь навигации, включая «Домой» и «Приемные комиссии». Если вы пропустите определенные уровни, вы запутаете пользователей, и навигационная цепочка не будет казаться такой полезной. Даже если пользователи не начинали с главной страницы, вы хотите дать им простой способ изучить ваш сайт с самого начала.
4. Переход от высшего уровня к низшему.
Важно, чтобы ваша навигационная цепочка читалась слева направо, при этом ближайшая ссылка слева — это ваша домашняя страница, а ближайшая ссылка справа — текущая страница пользователя. Исследование, проведенное Nielsen Norman Group, показало, что пользователи проводят 80% своего времени, просматривая левую половину страницы, а 20% — правую, что является веским аргументом в пользу дизайна слева направо. Кроме того, ближайшая слева ссылка будет отображаться как начало цепочки, поэтому вы хотите, чтобы это была ваша страница самого высокого уровня.
5. Следите, чтобы заголовки хлебных крошек соответствовали заголовкам страниц.
Чтобы избежать путаницы, вы захотите сохранить согласованность с заголовками вашей страницы и хлебных крошек, особенно если вы нацелены на определенные ключевые слова в этих заголовках. Вы также хотите четко связать заголовки навигации со страницей. Если в заголовке хлебных крошек нет ссылки, проясните это. Nestle эффективно маркирует свои заголовки хлебных крошек, чтобы они соответствовали заголовкам страниц. Например, «Области воздействия и обязательства» читается в навигационной цепочке так же, как и на странице.
Nestle также хорошо отличает ссылки от несвязанных с разными цветами — ссылки синего цвета, а ссылки, не являющиеся ссылками, остаются серыми.
6. Проявите творческий подход к дизайну.
Традиционная навигация по хлебным крошкам выглядит так: Главная> О нас> Карьера. Однако вам не нужно следовать традиционному пути, если вы чувствуете, что другой дизайн может больше понравиться вашей аудитории или лучше выглядеть на вашем сайте.
Например, Target использует навигацию с помощью хлебных крошек на страницах своих продуктов (потому что, кто бы не потерялся в разделе виртуальной обуви?), Но использует символы «/» и простой черно-серый текст.В этом случае тонкая вариация дизайна имеет смысл с точки зрения эстетики их сайта.
7. Держите его в чистоте и не загромождайте.
Навигация по хлебным крошкам — это просто помощь для пользователя, и в идеале ее не следует замечать, если пользователь ее не ищет. По этой причине вы не хотите загромождать навигацию по хлебным крошкам ненужным текстом.
Например,Eionet может обойтись без текста «Вы здесь». Хотя этот текст должен быть полезным, он загромождает страницу. При правильном дизайне навигация по хлебным крошкам должна быть достаточно заметной без введения.
8. Подумайте, какой тип хлебных крошек лучше всего подходит для вашего сайта.
Есть несколько различных типов хлебных крошек, которые вы можете использовать: на основе местоположения, на основе атрибутов и на основе истории. Навигационные цепочки на основе местоположения показывают пользователю, где он находится в иерархии сайта. Навигационные цепочки на основе атрибутов показывают пользователям, к какой категории относится их страница. Наконец, хлебные крошки на основе истории показывают пользователям конкретный путь, по которому они перешли на текущую страницу.
Bed Bath & Beyond использует навигацию на основе атрибутов, чтобы показать пользователям, к какой категории относится их страница продукта, поэтому пользователи могут вернуться к «Кухня» или «Маленькая бытовая техника», чтобы просмотреть похожие товары.Этот тип навигации наиболее эффективен для клиентов Bed Bath & Beyond. Создавая навигацию с помощью хлебных крошек, подумайте, что наиболее полезно для посетителей вашего сайта.
9. Знайте свою аудиторию.
Лучшие практики в области навигации по хлебным крошкам побуждают веб-дизайнеров размещать элементы навигации вверху страницы, но Apple, одна из самых ценных компаний всех времен, бросает вызов этой логике, размещая элементы навигации в нижней части своего сайта. В конце концов, очень важно знать свою аудиторию.Клиенты Apple, как правило, технически подкованы и, вероятно, найдут навигацию, если они им понадобятся. Учитывайте потребности клиентов и проводите A / B-тестирование, если вы не уверены.
Навигация по хлебным крошкам в HTML и CSS
Панировочные сухари не только полезны — их также легко добавить на ваш сайт с помощью небольшого кода HTML и CSS.
Начнем с HTML, который мы будем использовать для создания самих ссылок. Самый простой способ сделать это — организовать ваши ссылки в элементе неупорядоченного списка (
), где каждый элемент списка (
Вот HTML-шаблон для хлебных крошек, который вы можете использовать:
Обратите внимание, что я также заключил неупорядоченный список в элемент HTML (навигация) и добавил класс и метку ARIA к его открывающему тегу.Это необязательно, но помогает сделать вашу страницу более доступной для программ чтения с экрана и поисковых систем.
Конечно, одного этого HTML недостаточно — сейчас у нас есть только маркированный список ссылок. Добавляя CSS, мы можем добиться желаемого вида хлебных крошек. Примените следующий CSS к HTML выше:
ul {стиль списка: нет; } / * убираем маркеры * /
ul li {display: inline; } / * показать элементы списка по горизонтали * /
/ * Поместите угловую скобку (>) между элементами списка * /
ul li + li: before {
content: ">";
отступ: 10 пикселей;
}
ul li a {text-decoration: none; } / * убираем подчеркивание со ссылок * /
ul li a: hover {text-decoration: underline; } / * показывать подчеркивание при наведении курсора * /
Вместе этот код генерирует базовую область навигации, которую можно использовать на любом сайте — см. Окончательный результат ниже.Я также добавил несколько дополнительных стилей для более аккуратного вида. Чтобы увидеть, как панировочные сухари выглядят без этого стиля, закомментируйте код внизу вкладки CSS.
См. Панировочные сухари в HTML и CSS Кристины Перриконе (@hubspot) на CodePen.
Навигация по хлебным крошкам в Bootstrap CSS
Bootstrap CSS также предлагает способ создания хлебных крошек без добавления пользовательского CSS. Для этого используйте компонент Breadcrumb вот так. Вот пример из документации Bootstrap 5:
См. Хлебные крошки Pen в Bootstrap CSS Кристины Перриконе (@hubspot) на CodePen.
Это только основы навигации по хлебным крошкам в Bootstrap — см. Документацию по хлебным крошкам, чтобы узнать все подробности.
Дизайн, помогающий пользователям перемещаться по вашему сайту
В конечном счете, навигация по хлебным крошкам — это эффективный инструмент, облегчающий навигацию по вашему сайту, но вы хотите следовать передовым методам проектирования, чтобы убедиться, что вы используете полезность этого инструмента. Чтобы получить дополнительные советы по UX, ознакомьтесь с нашим полным руководством по UX-дизайну.
Примечание редактора: этот пост был первоначально опубликован в сентябре 2018 года и был обновлен для полноты.
Drosophila Stardust является партнером Crumbs в контроле полярности эпителиальных клеток
Müller, H.A.J. Генетический контроль полярности эпителиальных клеток: уроки Drosophila . Dev. Дин. 218 , 52–67 (2000).
Артикул Google Scholar
Bhat, M.A. et al. Discs Lost, новый белок с несколькими доменами PDZ, устанавливает и поддерживает полярность эпителия. Cell 96 , 833–845 (1999).
CAS Статья Google Scholar
Klebes, A. & Knust, E. Консервированный мотив в Crumbs необходим для локализации E-cadherin и образования слипшихся зон у Drosophila . Curr. Биол. 10 , 76–85 (2000).
CAS Статья Google Scholar
Tepass, U.& Knust, E. Crumbs и stardust действуют в рамках генетического пути, который контролирует организацию эпителия у Drosophila melanogaster . Dev. Биол. 159 , 311–326 (1993).
CAS Статья Google Scholar
Grawe, F., Wodarz, A., Lee, B., Knust, E. & Skaer, H. Гены Drosophila crumbs и stardust участвуют в биогенезе сращений. Разработка 122 , 951–959 (1996).
CAS PubMed Google Scholar
Tepass, U. Crumbs, компонент апикальной мембраны, необходим для образования поясничных сращений в первичном эпителии Drosophila . Dev. Биол. 177 , 217–225 (1996).
CAS Статья Google Scholar
Димитратос, С.Д., Вудс, Д. Ф., Статакис, Д. Г. и Брайант, П. Дж. Сигнальные пути сосредоточены в специализированных областях плазматической мембраны с помощью каркасных белков семейства MAGUK. Bioessays 21 , 912–921 (1999).
CAS Статья Google Scholar
Камберов Э. и др. Молекулярное клонирование и характеристика Pals, белков, связанных с mLin-7. J. Biol. Chem. 275 , 11425–11431 (2000).
CAS Статья Google Scholar
Müller, H. A. & Wieschaus, E. armadillo , bazooka и stardust имеют решающее значение для ранних стадий формирования зонул и поддержания поляризованного эпителия бластодермы у Drosophila. J. Cell Biol. 134 , 149–163 (1996).
Артикул Google Scholar
Шобер, М., Schaefer, M. и Knoblich, J. A. Bazooka привлекает Inscuteable для ориентации асимметричных клеточных делений в нейробластах Drosophila . Nature 402 , 548–551 (1999).
ADS CAS Статья Google Scholar
Wodarz, A., Ramrath, A., Kuchinke, U. & Knust, E. Bazooka предоставляет апикальный сигнал для локализации Inscuteable в нейробластах Drosophila . Nature 402 , 544–547 (1999).
ADS CAS Статья Google Scholar
Wodarz, A., Ramrath, A., Grimm, A. & Knust, E. Drosophila атипичная протеинкиназа C связывается с Bazooka и контролирует полярность эпителия и нейробластов. J. Cell Biol. 150 , 1361–1374 (2000).
CAS Статья Google Scholar
Петронски, М.& Knoblich, J. A. DmPAR-6 управляет полярностью эпителия и асимметричным делением клеток нейробластов у Drosophila . Nature Cell Biol. 3 , 43–49 (2001).
CAS Статья Google Scholar
Tepass, U., Theres, C. & Knust, E. crumbs кодирует EGF-подобный белок, экспрессируемый на апикальных мембранах эпителиальных клеток Drosophila и необходимый для организации эпителия. Cell 61 , 787–799 (1990).
CAS Статья Google Scholar
McGee, A. W. & Bredt, D. S. Идентификация внутримолекулярного взаимодействия между доменами Sh4 и гуанилаткиназы PSD-95. J. Biol. Chem. 274 , 17431–17436 (1999).
CAS Статья Google Scholar
Шин, Х., Hsueh, Y.-I., Yang, F.-C., Kim, E. & Sheng, M. Внутримолекулярное взаимодействие между доменом гомологии 3 Src и доменом, подобным гуанилаткиназе, необходимым для кластеризации каналов постсинаптической плотностью-95 / SAP90. J. Neurosci. 20 , 3580–3587 (2000).
CAS Статья Google Scholar
Wu, H. et al. Внутримолекулярные взаимодействия регулируют связывание SAP97 с GKAP. EMBO J. 19 , 5740–5751 (2000).
CAS Статья Google Scholar
Nix, S. L., Chishti, A. H., Anderson, J. M. & Walther, Z. hCASK и hDlg ассоциируются в эпителии, а их гомология 3 Src и домены гуанилаткиназы участвуют как во внутримолекулярных, так и во внутримолекулярных взаимодействиях. J. Biol. Chem. 275 , 41192–41200 (2000).
CAS Статья Google Scholar
Димитратос, С.Д., Вудс, Д. Ф. и Брайант, П. Дж. Камгук, Lin-2 и CASK: новые гомологи ассоциированной с мембраной гуанилаткиназы, которые также содержат домены СаМ-киназы. Mech. Dev. 63 , 127–130 (1997).
CAS Статья Google Scholar
Ламмель, У. и Сомвебер, Х. Х-сцепленные локусы Drosophila melanogaster , вызывающие дефекты морфологии слюнной железы эмбриона. Dev.Genes Evol. 10 , 525–535 (2000).
Артикул Google Scholar
Rorth, P. Модульный скрининг неправильной экспрессии у Drosophila , выявляющий тканеспецифические фенотипы. Proc. Natl Acad. Sci. США 93 , 12418–12422 (1996).
ADS CAS Статья Google Scholar
Рубин, Г. М., Спредлинг, А.C. Векторы для опосредованного Р-элементом переноса генов в Drosophila . Nucleic Acids Res. 11 , 6341–6351 (1983).
CAS Статья Google Scholar
Wodarz, A., Hinz, U., Engelbert, M. & Knust, E. Экспрессия Crumbs придает апикальный характер доменам плазматической мембраны эктодермального эпителия Drosophila . Cell 82 , 67–76 (1995).
CAS Статья Google Scholar
Хан, К.И Manley, J. L. Функциональные домены белка Drosophila Engrailed. EMBO J. 12 , 2723–2733 (1993).
CAS Статья Google Scholar
Тауэр, Дж., Карпен, Г. Х., Крейг, Н. и Спрэдлинг, А. С. Предпочтительная транспозиция Р-элементов Drosophila в близлежащие хромосомные участки. Genetics 133 , 347–359 (1993).
CAS PubMed PubMed Central Google Scholar
Коричневый, Н.H. & Kafatos, F.C. Функциональные библиотеки кДНК из эмбрионов Drosophila . J. Mol. Биол. 203 , 425–437 (1988).
CAS Статья Google Scholar
Hovemann, B.T., Dessen, E., Mechler, H. & Mack, E. Drosophila snRNP-ассоциированный белок P11, который специфически связывается с затяжкой теплового шока 93D, обнаруживает сильную гомологию с коровым белком hnRNP A1. Nucleic Acids Res. 19 , 4909–4914 (1991).
CAS Статья Google Scholar
Kennerdell, J. R. & Carthew, R. W. Использование dsRNA-опосредованной генетической интерференции для демонстрации того, что frizzled и frizzled 2 действуют в бескрылом пути. Cell 95 , 1017–1026 (1998).
CAS Статья Google Scholar
Бренд, А. Х.И Перримон, Н. Целенаправленная экспрессия генов как средство изменения судьбы клеток и генерации доминантных фенотипов. Разработка 118 , 401–415 (1993).
CAS Google Scholar
Oda, H., Uemura, T., Harada, Y., Iwai, Y. & Takeichi, M. A Drosophila гомолог кадгерина, связанного с броненосцем и необходимого для межклеточной адгезии эмбриона. Dev. Биол. 165 , 716–726 (1994).
CAS Статья Google Scholar
Crumbs является важным регулятором динамики цитоскелета и клеточно-клеточной адгезии во время дорсального закрытия у Drosophila
Обзор комментариев рецензентов: рецензенты соглашаются, что это потенциально хорошая работа и в пересмотренной форме может представлять интерес для большого числа людей. аудитория. Чтобы существенно отредактированная рукопись стала ценным вкладом в eLife , существует несколько основных проблем, включая важные меры контроля, которые необходимо решить.Авторам необходимо улучшить оформление, если они хотят, чтобы эта статья оказала желаемое воздействие. В представленном виде он труден для чтения и экспериментально неполон. Отчасти проблема заключается в том, что абзацы не очень хорошо структурированы, а тематические предложения неадекватно описывают темы, затронутые в абзаце. Таким образом, читатель ожидает узнать об одном, но данные и подробности представлены по многим другим. Частично проблема может заключаться в том, что авторы представили много материала (слишком много материала?), И, как следствие, изложение не такое ясное или полное, как могло бы быть.Кроме того, есть некоторые технические проблемы с представленными данными. Авторы должны сосредоточиться на представлении своих выводов более четко и полно, потенциально удаляя некоторые выводы, чтобы лучше документировать и обеспечивать соответствующий контроль над другими выводами. Мы существенно изменили текст и тем самым повысили его ясность. Кроме того, мы удалили часть данных (или представили их как дополнительный материал). Например, вариант Y10F использовали в качестве контроля, чтобы показать, что мутация Y10 сама по себе не является деструктивной и что дефекты в варианте Y10A обусловлены конформационным дефектом, а не предполагаемым дефектом фосфорилирования.Действительно, недавно Wei et al. (J Biol Chem 2015 290: 11384) показали, что мутация Y10A в FBM Crb устраняет взаимодействие с Moesin. Мы удалили все результаты для варианта Y10F из основных рисунков и при необходимости добавили их в качестве дополнительных. Также мы удалили все фильмы с результатами варианта Y10F. Мы соответствующим образом изменили текст, чтобы улучшить его плавность.
Доменно-специфические фенотипы являются основным направлением статьи. Следовательно, авт. Должны с ясностью установить, что фенотипы обусловлены мутантным доменом (авторам все еще необходимо подтвердить, что белок правильно уложен и перемещается, чтобы сделать вывод, что фенотипы являются домен-специфичными, а не просто crb-специфичными).Было бы полезно лучше показать локализацию мутантного белка с более высоким разрешением и во всех тканях. Предположительно, в непораженных тканях он должен выглядеть в точности как дикий тип или вариант Y10F.
Локализация различных вариантов foscrb в разных тканях была задокументирована в нашей предыдущей работе Klose et al., 2013 (G3 2013, 3: 153). Тем не менее, в первом абзаце раздела результатов мы подчеркиваем, что белок fosCrb Y10A локализуется апикально в большинстве эмбриональных эпителий, которые также, по-видимому, имеют нормальную клеточную полярность (например, эпидермис).Кроме того, в нашей текущей рукописи вы можете увидеть локализацию Crb и Crb Y10A в эмбриональном эпидермисе (ранее; сейчас). Локализацию всех этих вариантов в амниосерозе очень трудно получить в фиксированных образцах, потому что, согласно нашим наблюдениям с версией fosCrb :: EGPF (неопубликованные наблюдения D.F.B и E.K.), она, по-видимому, очень динамична.
Сделав это, авторам необходимо тщательно обсудить прямые и косвенные фенотипы, убедившись, что они не интерпретируются чрезмерно.Эксперименты по спасению с Rho и Flw предполагают, что может быть более прямое влияние на активность актомиозина, помимо дефектов адгезии. В этом контексте важно, чтобы основной вывод о том, что Crumbs негативно регулирует Rho1, был проверен напрямую (см. Конкретные комментарии ниже). «Мясо» рукописи лежит в серии спасательных экспериментов по закрытию спины и, в частности, по морфогенезу амниосерозы. Однако авторы делают анализ дефектов на уровне дорсальных эпидермальных клеток важной частью работы, которая затем, кажется, откладывается.Включение раздела, описывающего, как клетки AS становятся мигрирующими, не является существенным для основных моментов авторов, но является смелым заявлением с представленными данными. Им следует удалить этот раздел с бумаги или провести дополнительные контрольные эксперименты, чтобы показать, что эти клетки не выходят из гемоцитов.
Мы переписали некоторые части обсуждения и тщательно приняли во внимание возможность того, что некоторые из наблюдаемых дефектов являются косвенным следствием чрезмерного сжатия AS в эмбрионах foscrb Y10A (второй и третий абзацы) ).Что касается прямого измерения Rho1, мы объясняем причины, по которым этот эксперимент невозможен (см. Ниже). О том, что клетки AS становятся мигрирующими, мы переместили данные сканирования ЭМ в приложение (сейчас) и смягчили вывод, как рекомендовано рецензентом 1 в пункте № 3 (см. Ниже) (подраздел «FBM Crb необходим для адгезии AS », абзац третий).
Конкретные комментарии:
Рецензент 1:
Эта статья предоставляет убедительные фенотипические доказательства того, что аллель crumbs foscrb Y10A вызывает дефекты надклеточного актинового троса DME, протрузии DME и сокращения амниосерозы.Однако серьезная проблема заключается в том, что существует мало доказательств того, что все эти поразительно задокументированные фенотипы не являются вторичными по отношению к дефектам адгезии, обнаруженным здесь и ожидаемым в аллеле crumbs. Таким образом, некоторые выводы являются завышением представленных данных.
Мы переписали некоторые выводы и постарались быть более трезвыми и умеренными в заявлениях. Тем не менее, что касается дефектов адгезии, наблюдаемых у эмбрионов foscrb Y10A , они не похожи на фенотипы, описанные для потери функции crb , потому что i) фенотип Y10A очень специфичен для AS, и ii) он развивается позже, и iii) это зависит от FBM.Возьмем, к примеру, тот факт, что экспрессии UAS-Pak-AID с GAL4 334,3 достаточно, чтобы спасти летальность эмбрионов foscrb Y10A , в результате чего взрослые особи не проявляют каких-либо серьезных заболеваний. или явные дефекты в других тканях (). Это подтверждает вывод о том, что аллель foscrb Y10A не похож на хорошо документированный аллель потери функции crb , охарактеризованный в этой и других лабораториях. Сейчас обсуждается вопрос о том, являются ли некоторые из задокументированных фенотипов вторичными по отношению к дефектам адгезии, и мы согласны с тем, что это загадка, которую необходимо решить (подраздел «FBM Crb контролирует динамику актомиозина в AS», шестой абзац).
1) Хотя предоставленные изображения и видеоролики часто поражают воображение, количественная оценка может быть полезна, особенно при фенотипах DME. Например, на рис. 20 авторы выделяют клетки, которые маскируют способность независимо оценивать экспрессию; Было бы полезно получить профиль линии через эти ячейки и соседние ячейки.
Готово. Теперь мы показываем изображение без выделения () и дополнили его профилем линии вдоль клеток DME, используя сигнал от окрашивания фосфотирозином в качестве эталона для ZA.На рисунке показано, что интенсивность базука в клетках DME сильно снижена у эмбрионов foscrb Y10A .
Точно так же различия в кластеризации кадгерина трудно измерить на показанном рисунке, и количественная оценка будет полезна.
Мы обеспечиваем более качественное изображение AS на этапе 11 (). На этих изображениях можно увидеть нарушение непрерывности D E-cad на ZA эмбрионов foscrb Y10A ().Более того, поскольку теперь мы использовали нокаут-аллель D E-cad :: mTomato (жизнеспособный нокаут-аллель — Хуанг и др., 2009 PNAS 106: 8284 — как аллель D E-cad :: GFP используется в остальной части работы), автофлуоресценция желтка очень низкая, что улучшает отношение сигнал / шум. Однако количественная оценка этих дефектов оказалась довольно произвольной. Конечно, можно измерить интенсивность D E-cad вдоль ZA в эмбрионах foscrb и foscrb Y10F (например, с помощью профиля линии).Напротив, у эмбрионов foscrb Y10A расположение линии (то есть ROI) вдоль ZA в AS становится произвольным, поскольку сигнал не является непрерывным или даже теряется, поэтому мы не абсолютно уверены в том, что рисовать ROI вдоль ZA. Это становится еще более проблематичным на более поздних стадиях, когда ZA становится чрезвычайно разрушенным (см. Ток).
2) Вывод о том, что Юрта не задействована, потому что ее фенотип качественно менее серьезен, можно объяснить тем, что она участвует только в подмножестве видов деятельности.Комбинирование foscrb Y10A с аллелем юрты может легко проверить эту возможность.
Эксперимент и видео уже были представлены в первой версии рукописи (Видео 4), теперь (Видео 7). Теперь мы также более подробно объясним фенотип Yrt и изменим упомянутый абзац, чтобы прояснить, что фенотип yrt отличается от мутанта foscrb Y10A , а также что нет усиления или подавления foscrb Фенотип Y10A в сочетании с зиготическим мутантом yrt (подраздел «FBM Crb необходим для целостности AS», третий абзац).Эмбрион foscrb Y10F был удален из фильма, поскольку он фенотипически идентичен foscrb в сочетании с мутантом yrt .
3) Открытие того, что клетки амниозерозы становятся подвижными, очень поразительно. Однако эти клетки напоминают гемоциты, особенно на снимках, полученных с помощью СЭМ. Если в амниосерозе есть дыры, а также преждевременно апоптотические клетки, можно ожидать, что гемоциты будут реагировать на очистку этого клеточного мусора.Авторам следует подтвердить, что подвижные клетки не являются гемоцитами, или ослабить этот участок.
Готово, мы смягчили вывод и заявили, что мы не знаем идентичности этих мигрирующих клеток, которые вполне могут быть гемоцитами, выходящими через отверстие в AS эмбрионов foscrb Y10A (подраздел «FBM of Crb необходим для адгезии AS », третий абзац).
4) Главный вывод о том, что Crumbs отрицательно регулирует Rho1, следует проверить напрямую, особенно с учетом некоторых неожиданных результатов в экспериментах по спасению, где Rok, потеря функции Rho1 и доминантно-отрицательные аллели Rho1 не дают подобных результатов.Увеличен ли Rho1 или активный Rho1 на фоне foscrb Y10A ?
Поскольку это один из основных пунктов рукописи, мы, очевидно, хотели знать точное изменение активности Rho1. Кроме того, поскольку использование доминантно-негативных или доминантно активных конструкций может маскировать эффекты других малых GTPases, мы хотели проанализировать состояние in vivo. Тем не менее, невозможно измерить активные уровни Rho с помощью обычных анализов (таких как анализ активации Pull-down): AS представляет собой очень небольшую часть всего эмбриона, и даже в том случае, если весь белок Rho является активным (i .е. GTP загружен) в AS foscrb Y10A эмбрионов, мы думаем, что другие ткани эмбриона будут маскировать разницу при обработке для анализа.
Однако мы попытались подойти к этому вопросу другими стратегиями. Во-первых, мы использовали доступные линии, которые экспрессируют Rho1 :: GFP под контролем промотора Rho1, созданного в лаборатории Сьюзан Паркхерст, но ни одна из них не была достаточно сильной, а отношение сигнал / шум было слишком низким для анализа. Мы попытались создать рекомбинантные запасы с аллелем Rho1 1b плюс конструкцией Rho1 :: Rho1 :: GFP, плюс соответствующей фосмидой плюс нулевым аллелем crb (т.е. Вт *; foscrb, Rho1 1b , Rho1 :: Rho1 :: GFP / CyO, twi :: GFP; crb GX24 / TM3, twi :: GFP ), но этот подход оказался неудачным, так как промежуточные запасы были очень слабыми.
Другая стратегия заключалась в использовании биосенсоров активности ГТФазы семейства Rho, также созданных лабораторией Паркхерста (хотя они не были полностью охарактеризованы). Некоторые из них выражаются конститутивно, а другие являются конструкциями UAS. Мы исследовали использование различных версий Rok RBD , Dia RBD , Capu RBD и Pkn RBD с разными драйверами GAL4, и ни одна из них не дала полезного отношения сигнал / шум в наших настройках изображения. .Лаборатория Lecuit сообщила о новом зонде (Anillin :: GFP, Munjal et al., (2015). Nature, DOI: 10.1038 / nature14603), но работа была опубликована 27 июля, поэтому по очевидным причинам мы не тестировали этот зонд.
Рецензент 2:
Это увлекательная, интересная и тщательно продуманная работа, содержащая новую и важную информацию, которая будет интересна широкой аудитории. Мне очень понравилось читать эту работу, которая открывает несколько направлений для исследования молекулярных механизмов, лежащих в основе апикального сокращения клеток амниосерозы.У меня только пара вопросов: 1) Авторы говорят, что Крамбс является негативным регулятором динамики актомиозина, и сообщают об увеличении количества миозиновых (молниеносных) очагов, которые сохраняются дольше и более дезорганизованы. Интересно, следует ли локализация актина также этой аберрантной динамике. Очень вероятно, что это так, но, возможно, стоит проверить.
Мы создали мух, у которых аллель D E-cad :: mTomato был рекомбинирован с актин-связывающим доменом Утрофина, помеченным GFP, и экспрессировался под контролем промотора спагетти из кабачков (sqh :: Ut :: GFP), и поставить в crb нулевой фон.Подробный анализ этих зародышей не проводился. Мы предоставляем видео (видео ответа автора 1), где можно оценить пульсирующие сокращения у эмбриона foscrb и то, что AS эмбриона foscrb Y10A сильно поврежден (внутри пунктирной линии на слияние эмбриона foscrb Y10A ). D E-cad :: mTomato нарушен в «оставшейся части» AS, но, тем не менее, можно оценить кластеры зеленого канала, которые напоминают те, что наблюдаются с Zipper.Поскольку этот результат несколько избыточен, мы упоминаем его как «данные не показаны» в тексте (подраздел «FBM Crb контролирует динамику актомиозина в AS», первый абзац).
2) Что касается дефектов эпидермальных клеток, которые находятся в контакте с амниосерозой, а также дезорганизации актинового кабеля и филоподий у эмбрионов foscrbY10A, авторы, кажется, поддерживают идею о том, что они являются вторичным эффектом из-за дефектов сокращения. амниосероза. Хотя я согласен с тем, что некоторые из наблюдаемых дефектов можно отнести к сбоям в морфогенезе амниозерозы (т.е. D E-Cadherin localization), я думаю, что некоторые дефекты могут быть связаны со специфической функцией Crumbs в этих клетках. В этом смысле мне интересно, наблюдали ли авторы какие-либо различия в организации дорсального эпидермиса в различных проведенных экспериментах по спасению, например, восстанавливается ли актиновый кабель, формирование филоподий или локализация Ena, когда Flw эктопически экспрессируется в AS или когда одна копия SCAR, Arp3 или Arpc1 удаляется из эмбрионов foscrbY10A?
Действительно, мы не исследовали подробно организацию клеток DME в различных экспериментах по спасению.Мы сделали некоторые наблюдения, которые подтверждают идею вторичного эффекта: удлинение клеток DME восстанавливается у эмбрионов foscrb Y10A , экспрессирующих Pak-AID только в AS. Это наблюдалось при использовании D E-cad :: GFP в гетерозиготной комбинации, поэтому отношение сигнал / шум не является оптимальным, чтобы показать это. В видео ответа автора 2 на верхних панелях показаны максимальные проекции полученных стопок. Поскольку D E-cad :: GFP является гетерозиготным, контраст очень низкий.На нижних панелях часть тех же стопок использовалась для удаления аутофлуоресценции желтка и желтка на Фиджи.
Таким образом, чтобы получить разумное количество наблюдений и подробно проанализировать эти дефекты, нам необходимо создать другие запасы, в которых аллель DE-cad :: mTomato рекомбинирован с соответствующими генотипами. Однако мы не анализировали динамику цитоскелета клеток DME в этих условиях, поэтому мы не можем исключить возможность того, что некоторые дефекты в клетках DME эмбрионов foscrb Y10A являются вторичными по отношению к дефектам AS.Соответственно, мы более тщательно обсуждаем эти аспекты в рукописи (Обсуждение, последний абзац).
С другой стороны, в другом наборе неопубликованных экспериментов мы использовали драйверы engrailed-GAL4 или пропатченный-GAL4 для выражения различных конструкций ( UAS-Ena :: GFP, Uas-Pak-AID, UAS-Moesin, UAS- moesin T559D , UAS-flw и т. д.) в полосах на эпидермисе эмбрионов foscrb Y10A , но до сих пор мы не обнаружили никакого подавления или спасения фенотипа Y10A.
Рецензент 3:
Существенно измененная рукопись с некоторыми дополнительными элементами управления вполне может внести отличный вклад в eLife. 1) Предыдущее открытие заключалось в том, что фосмида, покрывающая весь локус крошек ( crb ), foscrb или вариант, который заменяет консервативный тирозин фенилаланином ( foscrb Y10F ), но не аланин ( foscrb ). Y10AI ) может полностью избавить от эмбриональной летальности, ретракции зародышевой оболочки, дорсального закрытия и дефектов инволюции головы, которые характерны для гомозиготных, crb нулевых животных.Они подробно оценивают функцию crb , анализируя foscrb и вариант восстановления нулевых образцов crb , используя живую визуализацию нокинового аллеля D, E-кадгерина, D E-cadGFP. Животные foscrb Y10A демонстрируют серьезные дефекты втягивания зародышевых лент, имеют сильно дезорганизованную амниозерозу, которая со временем теряется. В конечном итоге дорсальное закрытие не удается. Авторы не объясняют, почему они выбрали для анализа два варианта, Y10F, который во всех случаях имитирует вариант Y10 дикого типа, и Y10A, который во всех случаях ведет себя совершенно по-разному.Они должны объяснить, почему они оценивают Y10F, и могут значительно сэкономить место, просто сообщив, что вариант Y10F ведет себя как Y10 дикого типа во всех случаях (возможно, я пропустил, где это не так?).
Спасибо за предложение. Теперь мы объясняем в первом абзаце результатов, что аллель foscrb Y10F является контролем, в котором эмбриональное развитие происходит, как у foscrb или эмбрионов дикого типа. Мы также объясняем, что эти результаты указывают на то, что фенотипы в эмбрионах foscrb Y10A обусловлены конформационным искажением мотива связывания FERM-домена Crb (как недавно было показано Wei et al., 2015 J Biol Chem 290: 11384), а не из-за предполагаемого вмешательства в фосфорилирование Crb в этом остатке тирозина. Соответственно, мы решили удалить все результаты для варианта foscrb Y10F из основных рисунков и видео и добавлять их в качестве дополнений к рисункам, когда мы сочли это необходимым, что не только экономит место, но и улучшает поток чтения.
2) Затем они исследуют роль Yrt, белка-кандидата в области FERM, который может действовать совместно с Crb.В норме Crb, D, Patj и Yrt экспрессируются на более высоких уровнях в эпидермисе, чем в амниосерозе, но в значительной степени отсутствуют на переднем крае большинства дорсальных клеток. У животных Y10A DME не удлиняются должным образом, и Crb и D Patj связаны с передним краем, особенно в клетках, которые остаются короткими. В отличие от этого, юрта отсутствует на переднем крае у нормальных животных и животных Y10A. Кроме того, авторы обнаружили, что фенотип мутанта yrt отличается от фенотипа мутанта Y10A, который, по их утверждению, начинается раньше и является более сложным, чем фенотип yrt .Они пришли к выводу, что yrt , по-видимому, не участвует в фенотипе эмбрионов Y10A (это неубедительно, см. Конкретные комментарии ниже).
Мы решили эти проблемы (см. Ниже).
3) Далее авторы демонстрируют, что ключевой особенностью фенотипа Y10A является отсутствие филоподий на переднем крае, и заключают, что функция crb необходима для нормальных микроворсинок. 4) Затем авторы обращают внимание на то, как crb действует в локализации ехиноидов и в формировании богатого актомиозином кабеля в клетках DME.Они описывают изменения в распределении ехиноидов и сообщают об изменениях в распределении Baz, которые не являются особенно убедительными. Кроме того, они обнаружили, что распределения yrt, coracle и Dlg были нормальными для всех вариантов фосмид, и пришли к выводу, что crb не отвечает за определение их локализации.
Как также указал рецензент 1, дефекты в распределении базука в клетках DME эмбрионов foscrb Y10A были нечеткими, и по его просьбе мы теперь показываем изображение без выделения () и дополнили его профиль линии вдоль клеток с использованием сигнала от окрашивания фосфотирозином в качестве эталона для ZA.На изображении можно увидеть, что интенсивность базука в клетках DME сильно снижена у эмбрионов foscrb Y10A .
5) Затем авторы оценивают локализацию D E-кадгерина в вариантах foscrb и сообщают, что в foscrb и Y10F D E-cad локализуется во всех межклеточных контактах, включая LE. , но не обсуждают нормальную потерю D E-cad из передней кромки позже при закрытии.Оценивали ли они распределения DE-cad во время закрытия (оно кардинально меняется)? Авторы сообщают, что DE-cad на LE и в амниосерозе снижается у животных Y10A. Их вывод о том, что животные Y10A лишены актомиозинового аппарата LE, безусловно, оправдан, но они должны уточнить свои выводы о адгезии, основываясь на лучшей характеристике распределений E-cad D на ранних и поздних стадиях закрытия.
Этот вопрос также относится к вашему конкретному комментарию ниже относительно нормальной потери D E-cad из передней кромки, на которую мы ссылаемся из Gorfinkiel and Arias 2007 (J Cell Sci 120: 3289), который показывает позднюю стадию DC (во время фазы застегивания молнии), который, однако, никогда не достигается эмбрионами foscrb Y10A .Наша характеристика сосредоточена на более ранних стадиях эмбрионального развития (т.е. в начале ретракции зародышевой ленты), потому что это моменты времени, когда начинается фенотип Y10A.
В любом случае, в вы можете видеть, что при позднем закрытии потеря D E-cad из LE действительно происходит в эмбрионах foscrb и foscrb Y10F (см. Стрелки). Эти наблюдения согласуются с нашими выводами, поскольку эти аллели полностью устраняют недостаток эндогенного crb и ведут себя во всех проанализированных аспектах как дикий тип.
Обращаем ваше внимание на то, что мы обращаем внимание на дефекты D E-cad локализации в LE с начала DC в эмбрионах foscrb Y10A . Таким образом, мы считаем, что нет необходимости включать эти наблюдения в рукопись, поскольку перераспределение D E-cad на поздних стадиях не меняет нашу интерпретацию дефектов в AS эмбрионов foscrb Y10A .
6) Авторы также оценивают состояние амниозерозы у мутантов Y10A и сообщают, что существуют разрывы в распределении D E-cad на границах AS и что есть большие внутриклеточные кластеры D E-cad. у животных Y10 (однако см. комментарии ниже).Поскольку ретракция зародышевой оболочки не удается у этих эмбрионов, неясно, что описанные дефекты обусловлены дефектами дорсального закрытия как таковых, и, как следствие, авторы должны пересмотреть свои выводы.
Этот комментарий относится к той же теме, что и комментарий № 7, поэтому см. Ответ ниже.
7) Затем авторы оценивают целостность амниозероза. Они на самом деле не объясняют, почему они использовали для этого визуализацию связывающего микротрубочки белка Юпитера (никаких комментариев о распределении или функции микротрубочек не имеется, поэтому читатель остается в недоумении).Тем не менее, они обнаружили, что хотя расширения amnioserosa по эпидермису характеризуют все три варианта foscrb, удлинения wt и Y10F разрешаются во время закрытия, а у животных Y10A — нет. Опять же, представляется разумной возможность, что амниосероза распадается и поскольку ретракция зародышевой полосы и дорсальное закрытие не прогрессируют у животных Y10A, клетки AS принимают фенотип случайной миграции. Представленные SEM подтверждают аберрантный характер апикальных поверхностей клеток AS, который согласуется с их входящим апоптозом.Это может происходить из-за того, что они преждевременно апоптозны или вместо этого они подвергаются апоптозу в «подходящее» время, но ретракция зародышевой ленты и дорсальное закрытие задерживаются. Могут ли авторы сослаться на другие стадии развития, подтверждающие или опровергающие эту интерпретацию?
Мы использовали Jupiter :: GFP в качестве зонда для отслеживания морфогенеза всего эмбриона, потому что этот маркер 1) экспрессируется во всех клетках, 2) это белковая ловушка, поэтому его 3) экспрессия находится под контролем эндогенный промотор; 4) он гомозиготен по жизнеспособности; 5) для этого аллеля не сообщалось о фенотипе.Это дает преимущество перед другими доступными маркерами, такими как актин-связывающий домен Moesin :: GFP, который экспрессируется под контролем промотора sqh или UAS, или LifeAct, что не подходило в наших экспериментах. Для большей ясности мы полностью удалили эксперимент Jupiter :: GFP.
С другой стороны, мы также думали о возможности «отсроченного» ретракции зародышевой полосы у эмбрионов foscrb Y10A , но пришли к выводу, что апоптоз AS, происходящий в «подходящее» время, очень маловероятен.Напр., Экспрессии flapwing или Pak-AID в AS достаточно для подавления фенотипа ретракции зародышевой полосы, что предполагает, что этот фенотип является вторичным по отношению к дефектам в AS.
К сожалению, мы не можем использовать другие морфологические характеристики, такие как удлинение задней кишки или разветвление и слияние трахеальной системы, потому что эти характеристики нарушены у эмбрионов foscrb Y10A , вероятно, из-за дефектов зародыша. ретракция бандажа (эта работа и Klose et al., 2013 G3, 3: 153).
Нам известно о сложности стадирования эмбрионов foscrb Y10A (и других мутантных генотипов) (Klose et al., 2013 G3, 3: 153). Таким образом, как мы делали в предыдущей работе и как теперь объясняется в текущих Результатах (подраздел «FBM Crb необходим для закрытия дорсальной области») и Материалах и методах (подраздел «Сбор эмбрионов и окрашивание антител»), все генотипы собраны одновременно , и коллекции ограничены коротким периодом времени (указанным на каждом рисунке или в каждом фильме).Затем эмбрионы инкубируются в соответствующее время и фиксируются или визуализируются одновременно (даже при визуализации в реальном времени три генотипа находятся в одной камере и отображаются с использованием многопозиционной настройки), а этап относится к Морфология foscrb основана на Campos-Ortega and Hartenstein 1985 (Эмбриональное развитие Drosophila melanogaster ). Соответственно, когда мы описываем какой-либо дефект в эмбрионах foscrb Y10A , мы всегда сравниваем его с эмбрионами foscrb того же возраста, что означает, что оба были проанализированы одновременно после сбора яиц.
Следовательно, например, если мы сообщаем о дефекте ретракции зародышевых лент (возьмем, например), это происходит потому, что в этот момент времени эмбрионы foscrb и foscrb Y10F (контрольные эмбрионы) уже закончили втягивание.
Разобраться в том, являются ли дефекты зародышевой ленты прямым или косвенным следствием дефектов постоянного тока, выходит за рамки нашей настоящей работы. Эта конкретная тема была проанализирована другими, поэтому роль AS (механическая и / или биохимическая) во время ретракции зародышевых лент все еще находится в стадии интенсивных исследований: Yip et al., 1997 Development 124: 2129; Ламка и Липшиц, 1999 Dev Biol 214: 102; Шёк и Перримон, 2002 Dev Biol 248: 29; Шёк и Перримон, 2003 Genes Dev 17: 597; Козлова и Таммель, 2003 Science 301: 1911; Reed et al., 2004 Curr Biol 14: 372; Zúñiga et al., 2009 BMC Biol 7:61; Lynch et al., 2013 Dev Biol 384; 205; Lynch et al., 2014 New J Phys 16: 055003.
8) Затем они оценивают апоптоз и делают вывод, что апоптоз является преждевременным у эмбрионов Y10A. По причинам, подробно изложенным ниже, их данные неубедительны в этом отношении.
Мы предполагаем, что вы имеете в виду наше утверждение «Экспрессия аполинера в AS […] проявляется как апоптотические клетки» как «причины, подробно описанные ниже», поэтому, пожалуйста, прочтите следующий ответ.
9) Они также оценивают передачу сигналов JNK, используя puc-LacZ , который является биомаркером передачи сигналов JNK. Они убедительно показывают, что локализация puc-LacZ аномальна в эмбрионах Y10A, скорее всего, как следствие аномального удлинения клеток DME. Однако, поскольку имеется такое же количество puc-lacZ положительных ядер у Y10A животных, авт., По-видимому, заключают, что передача сигналов JNK является нормальной.Тем не менее, они никогда явно не говорят, что передача сигналов JNK является нормальной у эмбрионов Y10A.
Готово; заключение было включено (подраздел «FBM Crb необходим для целостности AS», конец пятого параграфа).
10) Авторы приступают к оценке импульсных сокращений клеток амниосерозы. Видео и фотографии подтверждают, что отдельные клетки в эмбрионах Y10A трудно различить. Тем не менее, у эмбрионов, меченных D E-cad, наблюдаются импульсные сокращения.Кроме того, эмбрионы, меченные zipGFP, демонстрируют четкое импульсное образование миозиновых очагов, которые впоследствии рассеиваются. Авторы отмечают, что период формирования и распространения очагов очень нерегулярен у эмбрионов Y10A, и фильмы и кимографы согласуются с этим наблюдением. Однако такие фильмы и кимографы (отдельных очагов) не дают достаточно количественного обзора эффекта. Авторы должны количественно определить периоды для случайно выбранных фокусов в клетках wt, Y10F и Y10A.Следует проанализировать как минимум десятки очагов, если не сотню или около того. Гистограммы наблюдаемых периодов образования и растворения очагов важны для того, чтобы сделать выводы, которые они впоследствии делают относительно роли crb FBM в регуляции актомиозина.
Готово; гистограммы подтверждают наши наблюдения (см. рис. 5 — приложение 3).
11) Авт. Показывают, что сверхэкспрессия Flapwing, фосфатаза для phospho-sqh подавляет отказ Y10A в ретракции зародышевой полосы и дорсальном закрытии.Это очень интересное открытие, которое предоставляет наиболее убедительные доказательства того, что crb опосредует свои эффекты посредством регуляции сократимости миозина. Тем не менее, несколько удивительно, что авт. Не оценили лучше эффекты экспрессии flapwing на целостность клеток amnioserosa (по результатам анализа на эмбрионах, экспрессирующих D E-cad-GFP). Таким образом, панели M-O имеют очень низкое увеличение и качество, так что целостность ячеек AS и адгезионных стыков D E-cad-GFP не может быть установлена.Авторы должны представить фильмы, сравнимые с Видео 10 с избыточной экспрессией отворотов, а также должны количественно оценить период импульсных сокращений в крыльях при экспрессии эмбрионов (для эмбрионов wt, Y10F и Y10A).
Мы не смогли оценить целостность ZA in vivo в экспериментальных условиях, в которых экспрессируется закрытие, потому что D E-cad :: GFP является гетерозиготным (рекомбинирован на той же хромосоме, что и UAS-flw ), поэтому отношение сигнал / шум очень низкое.
В фильме, на который ссылается рецензент (предыдущее видео 10, теперь видео 8), аллель D E-cad :: GFP был рекомбинирован с соответствующим вариантом foscrb , поэтому он гомозиготен и сигнал-шум соотношения достаточно для детального наблюдения за D E-cad :: GFP, несмотря на сильную автофлуоресценцию желтка. Следовательно, для правильного анализа ZA in vivo, когда flw экспрессируется в AS, аллель D E-cad :: GFP должен быть рекомбинирован также на хромосоме, несущей соответствующий вариант foscrb и GAL4 332 .3 (т.е. анализ эмбриона с генотипом w *; foscrb, DE-cad :: GFP, GAL4 332.2 / UAS-flw-HA, DE-cad :: GFP; crb GX24 / crb 11A22 , UAS-Actin :: RFP ), что было невозможно. Другой стратегией будет рекомбинирование D E-cad :: mTomato с UAS-flw , но эти заготовки не были готовы вовремя для этих экспериментов.
Тем не менее, мы подготовили фильмы (теперь Видео 12, подраздел «FBM Crb контролирует динамику актомиозина в AS») с лучшим разрешением AS у эмбрионов, экспрессирующих крылатые лоскуты и гетерозиготных для D E-cad :: GFP, где можно оценить восстановление D E-cad :: GFP в ZA в AS foscrb Y10A эмбрионов.Что касается вашего последнего пункта, анализа периода импульсных сокращений, низкое отношение сигнал / шум делает этот анализ довольно трудоемким и даже произвольным, так как повышенная мощность лазера приводит к отбеливанию и другим техническим проблемам (например, адгезии клеток к клеткам не совсем так. различимы).
Хотя разумно ожидать, что импульсные сокращения будут подавляться выражением взмахов крыльев, эта информация не имеет отношения к основной цели нашей работы. Пожалуйста, имейте в виду, что этот эксперимент является «доказательством принципа», в котором мы спросили, приводит ли экспрессия миозинфосфатазы к подавлению фенотипа DC у эмбрионов foscrb Y10A .Другими подходами могла быть экспрессия, например, MBS (другой субъединицы миозинфосфатазы). Таким образом, мы не видим оправдания для анализа периода импульсных сокращений (или других механических свойств) этой ткани в этих условиях. Как указывалось выше, этот анализ требует создания других инструментов.
Тем не менее, мы провели очень предварительный и грубый анализ (2 эмбриона / генотип), и уровень DC после экспрессии Flw, по-видимому, не изменился у эмбрионов foscrb и нормальный у эмбрионов foscrb Y10A ().
12) Чтобы понять механизмы фосфорилирования миозина, авторы обращают внимание на три эффектора малых GTPases семейства Rho / Rac / Cdc42, Rok, D Pakand Scar. Они представляют данные в таблице, которая является частью. Его нелегко понять, только плохо упоминается в тексте. Во-первых, в представленных числах нет планок погрешностей. 71,3% или 70,5% действительно отличаются друг от друга (вероятно, нет)? Отличаются ли они от 59,7 или 56,5 (без погрешностей узнать невозможно)? Моя интерпретация первых трех строк таблицы состоит в том, что выражение UAS-Actin-RFP под контролем драйвера амниосерозала (мне не нужно ссылаться на другую таблицу, чтобы выяснить, где находится Gal4-332.2) существенно подавляет наиболее тяжелый дорсальный открытый фенотип (т.е. с 59,7 до 18,5), но авторы это не комментируют. Возможно, они хотят, чтобы мы сосредоточились только на пенетрантности фенотипа wt-подобного? Если так, они должны прямо заявить об этом и рассмотреть возможность объединения дефектов в единую дорсальную открытую категорию, возможно, отнесение более подробной панели к дополнительному материалу). Более того, там, где они явно привлекают наше внимание к изменениям, сравнения кажутся ошибочными.Они просят нас сравнить строку 4 с строкой 3, чтобы показать, что экспрессия UAS-crb увеличивает наиболее серьезное дорсальное отверстие, но эмбрионы в строке 4 не экспрессируют UAS-actin-RFP и экспрессию UAS- actin-RFP сильно влияет на количество животных в наиболее тяжелой категории (сравните строку 3 с строками 1 и 2). Точно так же строки 9 и 10 сравниваются с 3, но 9 и 10 не выражают UAS-actin-RFP . Точно так же наблюдаются различия с UAS-moe (без актина) vs.UAS фосфомиметик моэ (с актином) просто из-за экспрессии актина? Если я правильно понимаю, эксперименты не сравниваются с соответствующими контролями.
Спасибо за предложение объединить дефекты в одну открытую спинную категорию. Мы изменили основной рисунок как таковой и предоставили подробный график в качестве дополнения. Мы также включили еще 3 контрольных скрещивания (и столбцы 3-5) для соответствующих сравнений. Экспрессия UAS-Actin :: RFP не подавляет фенотип foscrb Y10A (сравните 3 rd , 5 th и 6 th столбцов). в то время как раньше был представлен общий процент объединенных экспериментов.Наконец, мы более подробно объяснили сравнения в результатах.
13) Тем не менее, некоторые данные показывают интересные тенденции. Сверхэкспрессия аутоингибиторного домена D Pak подавляет фенотип DC у животных Y10A, тогда как сверхэкспрессия D Pak индуцирует фенотип DC у нормальных эмбрионов. Верно ли, что аутоингибирование D Pak является внутримолекулярным?
Нет, межмолекулярный. Ингибиторный домен в комплексе с PAK1 представляет собой димер между аутоингибиторной областью и киназными доменами.Если вас интересуют детали взаимодействия, то эти статьи описывают его: Lei et al., 2002 Cell 102, 387–397 и Parrini et al., 2002 Mol Cell 9, 73–83. Также могут быть интересны эти обзоры: Hofmann et al., 2004 J Cell Sci 117, 4343–4354; Чжао и Мансер, 2005 Biochem J 386, 201–214; Кумар и др., 2006 Nat Rev Cancer 6, 459–471.
Если это так, то не удивительно ли, что аутоингибиторный домен эффективен, когда он представлен как отдельный белок? Насколько чрезмерно выражен домен?
«Насколько сильно выражен домен?» Вы имеете в виду, что хотите знать изобилие? Если это так, мы думаем, что любые измерения кажутся произвольными.Он будет сравнивать «0» уровень экспрессии (CTL) с экспрессией, индуцированной GAL4. Даже если мы увидим какой-либо сигнал при иммунофлуоресценции или вестерн-блоттинге, он будет только информативным относительно того, экспрессируется Pak-AID или нет.
Другими ключевыми данными являются подавление фенотипов Y10A DC за счет сверхэкспрессии D E-Cad. 14) Последующий анализ роли Arp2 / 3 сфокусирован на роли Arp2 / 3 в эндоцитозе и интерпретируется как эндоцитоз D E-cad. Ламелиподии и филоподии являются ключевыми особенностями как переднего края, так и амниосерозальных тканей, а Arp2 / 3 может играть ключевую роль в их функции независимо от их роли в эндоцитозе.Почему авторы приписывают влияние Arp2 / 3 исключительно на эндоцитоз?
Мы включаем эту возможность в Обсуждение (третий абзац и подраздел «Crb — регулятор целостности ZA через динамику актомиозина?»).
15) Наконец, авторы заключают, что функция crb влияет как на динамику актомиозина, так и на адгезию, и что эти два эффекта могут быть взаимосвязаны, но название их статьи предполагает только роль в регуляции динамики актомиозина.Непонятно, почему это уместно. Хотя они используют Обсуждение для аргументации своей точки зрения, кажется, что пересмотренный текст Результатов с некоторой экспериментальной переработкой может существенно изменить выводы, сделанные в Обсуждении. На этом этапе их название следует изменить, чтобы точнее отразить твердые выводы, сделанные в Результатах, и оставить предположения в Обсуждении, четко обозначенные как таковые.
Главный вывод не изменился. Однако мы ценим предложение рецензента и изменили заголовок.
В дополнение к этим общим комментариям я сделаю следующие конкретные моменты для введения и части результатов. Я включил сопоставимые конкретные комментарии в более общие комментарии выше для остальных результатов, но обнаружил, что подробные комментарии становятся слишком многочисленными и слишком сложными.
В подразделе «FBM Crb важен для дорсального закрытия», первый абзац: авторы приходят к выводу, что домен связывания FERM crb ответственен за эффекты, которые они видят, потому что точечная мутация (Y10A) в FBD не может спасти функцию мутанта crb .Знают ли они, что вариант аланина производится и правильно доставляется к плазматической мембране? Если да, они должны сообщить, откуда они это узнали, и предоставить ссылку. В противном случае выводы относительно функции FBD (по сравнению с функцией crb в целом) ошибочны.
Как объяснено выше в вашем первом комментарии и объяснено в первом абзаце результатов, это было представлено в нашей предыдущей работе: Клозе, С., Флорес-Бенитес, Д., Ридель, Ф., и Кнуст, Э. (2013). Анализ структуры-функции на основе фосмид выявляет функционально различные домены в цитоплазматическом домене Drosophila crumbs.G3 (Bethesda) 3, 153–165.
Конечно, дорсальное закрытие не удается у этих животных Y10A (), но ретракция зародышевой ленты далека от завершения. Более того, у нормальных эмбрионов амниосероза подвергается апоптозу после завершения закрытия. Наблюдаются ли дефекты закрытия из-за того, что на заднем конце угол зрения не формируется должным образом из-за неполного втягивания зародышевой ленты и из-за того, что амниосероза подвергается апоптозу, блокируя закрытие? Это придает другой вид наблюдаемым дефектам.
Это тот же вопрос, который задан в ваших комментариях 6 и 7; пожалуйста, смотрите соответствующие ответы.
В подразделе «FBM Crb необходим для дорсального закрытия» авторы заявляют, что «Из этих видов белков Yrt является вероятным кандидатом, ответственным за дефекты в мутанте foscrb Y10A , поскольку Yrt отрицательно регулирует Crb (Laprise et al., 2006), зиготические yrt мутантные эмбрионы обнаруживают дефекты DC (Hoover and Bryant, 2002), а сверхэкспрессия Crb в амниозерозе приводит к дефектам DC (Harden et al., 2002; Wodarz et al., 1995) ». Я не понимаю, как «сверхэкспрессия Crb в амниозерозе приводит к дефектам постоянного тока» делает Yrt вероятным кандидатом? Рукопись характеризуется рядом утверждений, таких как это, которое отвлекает читателя и отвлекает от сообщения, которое авторы пытаются донести.
Мы перефразировали третий абзац, чтобы сделать его более ясным (подраздел «FBM Crb необходим для целостности AS»). Идея состоит в том, что Yrt негативно регулирует Crb, взаимодействуя с его FBM напрямую, а мутанты yrt (что приводит к сверхэкспрессии Crb) или сверхэкспрессия UAS-Crb в AS вызывают сходные дефекты DC.Таким образом, возможно, что взаимодействие Yrt и Crb нарушено в эмбрионах , foscrb, , Y10A, , . Анализ эпистаза показывает, что такая возможность маловероятна (видео 7).
В подразделе «FBM Crb необходим для закрытия спины», пятый абзац: Авторы должны показать на схеме ориентацию поперечных сечений, показанных на панелях M, N и O. Ни текст, ни Легенда рисунка адекватно описывает ориентацию секции и то, как она была получена.Таким образом, читателю трудно понять, что эти панели пытаются показать.
Мы добавили справочную ось в и, а также соответствующий пояснительный текст в пояснениях к рисункам.
В подразделе «FBM Crb необходим для закрытия спины» авторы заявляют: «Более того, в foscrb и foscrb Y10F без зиготического yrt DC выходит из строя только после ретракции ГБ, качественно отличается от фенотипа foscrb Y10A или foscrb Y10A зародышей без зиготических yrt (видео 4) ».Авт. Должны четко указать, как фенотипы качественно различаются, и явно указать, отличается ли фенотип Y10A в присутствии или отсутствии зиготического yrt . Кроме того, следующее предложение «Эти результаты предполагают, что фенотип DC foscrb Y10A эмбрионов начинается раньше и является более сложным, чем у мутантов yrt » требует от авторов предоставить нам более подробную информацию о фенотипах yrt , на которые есть только ссылки и неадекватное описание.Мне было очень трудно разобраться в этом абзаце. Авторам следует подумать о его переработке для ясности.
Выполнено, см. Подраздел «FBM Crb необходим для целостности AS», третий абзац.
В подразделе «FBM Crb регулирует образование филоподий и организацию надклеточного актомиозинового кабеля», первый абзац: авторы должны предоставить ссылку или другое объяснение того, почему Sas окрашивает филоподии.
Это было перемещено в подраздел «FBM Crb регулирует образование филоподий и организацию надклеточного актомиозинового кабеля в клетках DME».Это наше наблюдение (как при окрашивании, так и при визуализации в реальном времени — Видео 4). В любом случае, после пересмотра литературы, это окрашивание уже было замечено в другой работе (в Denholm et al., 2005 Development 132: 2389), но не упоминается явно в тексте или на рисунке.
Кроме того, учитывая, что ядро микроворсинок состоит из актиновых волокон, я не понимаю, почему микроворсинки не видны у животных foscrb и Y10F (). (В соответствующих условиях эна также может быть видна в микроворсинках, но предыдущие отчеты предполагают, что она видна в первую очередь, если эна чрезмерно экспрессирована).
Действительно, конфокальная камера была настроена таким образом, что виден только сигнал от кабеля. Филоподии есть, но чтобы их увидеть, необходимо увеличить усиление и мощность лазера, и кабель будет выглядеть как капля, находящаяся вне зоны досягаемости.
В подразделе «FBM Crb регулирует образование филоподий и организацию надклеточного актомиозинового кабеля» авторы заявляют: «Формирование актомиозинового кабеля в LE зависит от асимметричного накопления адгезионного белка Echinoid (Ed). в клетках DME (Laplante, Nilson, 2011; Lin et al., 2007) ». Как написано, только читатели, знакомые с упомянутой работой, поймут, что асимметрия белка Ed в клетках DME ненормальна и что важна асимметрия в распределении Ed между эпидермисом и амниосерозой. Они также заявляют, что «Удаление Ed из LE очевидно у эмбрионов foscrb и foscrb Y10F (стрелки указывают на отсутствие Ed на LE)», но Ed также не присутствует на переднем крае DME. клетки у животных Y10A.
Мы переписали абзац, чтобы прояснить, что распределение Ed в эмбрионах foscrb аналогично тому, которое описано для эмбрионов дикого типа. Также мы отмечаем, что фенотип в эмбрионах foscrb Y10A заключается в том, что общий уровень Ed в DME снижен, а не в том, что Ed отсутствует в LE (подраздел «FBM Crb регулирует образование филоподий и организация надклеточного актомиозинового кабеля в клетках DME »).
Более того, данные, представленные в, отнюдь не убедительны.Baz присутствует на переднем крае только одной клетки Y10A, и возможно, что показанная темная линия является фоном, как линия чуть выше стрелки, глубже в амниосерозе.
Это также рассматривается в вашем пункте №4 выше, и, помимо профиля линии, в ‘вы также можете увидеть, что обе темные линии локализуются в той же плоскости, что и окрашивание фосфотирозином, что свидетельствует о том, что это не фон.
В подразделе «FBM Crb регулирует образование филоподий и организацию надклеточного актомиозинового кабеля», шестой абзац: Gorfinkiel и Arias 2007 очень убедительно показывают, что D E-cad первоначально локализуется в LE, но позже во время закрытия теряется с передней кромки (см. Горфинкиель и Ариас).Авторам необходимо убедиться, что их наблюдения за спасенными эмбрионами fosmid согласуются с литературой, и рассмотреть, насколько очистка переднего края D E-cad согласуется с механизмами, которые они предлагают здесь. Более того, качеству изображений, окрашенных D E-cad, недостает контраста (сравните Горфинкиал и Ариас, рис. Aiii и Biii с этой рукописью). Лучше окрашенные препараты могут дать лучшее представление о распределении D E-cad у животных Y10A.
Это частично совпадает с комментарием №5; см. ответ выше. Для этого мы приобрели новые изображения, которые теперь показаны в и.
В подразделе «FBM Crb имеет важное значение для морфогенеза AS», второй абзац: Авторы утверждают, что в локализации D E-cad у животных Y10A имеются разрывы, что, по-видимому, имеет место, хотя если структура AS значительно отличается от дикого типа. Откуда авторам известно, что стыки D E-cad просто не вышли из плоскости сечения? Более того, авторы ссылаются на крупные агрегаты D E-Cad-GFP в цитоплазме клеток Y10A, но, по-видимому, таких агрегатов немного даже у животных foscrb и большое количество таких агрегатов у животных Y10F ( я.е. числа, сравнимые с числами, наблюдаемыми в клетках Y10A, см. 3A ‘и 3B’, соответственно).
Чтобы решить эту проблему, мы создали рекомбинанты D E-cad :: mTomato с различными вариантами foscrb . Это привело нас к анализу в условиях лучшего отношения сигнал / шум локализации D E-cad (и Video 4). Предыдущие изображения с использованием аллеля D E-cad :: GFP вводили в заблуждение некоторых обозревателей, поскольку автофлуоресценция желтка была принята за внутриклеточные агрегаты D E-cad :: GFP.
На изображении, представленном в первой версии нашей рукописи, мы смогли отличить их от внутриклеточных агрегатов D E-cad, используя исходные данные, потому что гранулы желтка локализуются глубоко в эмбрионе. Теперь, используя аллель D E-cad :: mTomato, можно показать максимальную проекцию без такого большого вмешательства желтка, поскольку его автофлуоресценция ниже в спектре излучения mTomato (около 560-600 нм). Тем не менее, на видео 5 по-прежнему показаны анализы с аллелем D E-cad :: GFP, но к легенде видео было добавлено примечание.
В подразделе «FBM Crb необходим для морфогенеза AS», пятый абзац, авторы заявляют: «Экспрессия аполинера в AS foscrb и foscrb Y10F эмбрионов (видео 9) выявили некоторые апоптотические клетки в заднем уголке глаза в конце ретракции ГБ (, стрелки). У эмбрионов foscrb Y10A аналогичного возраста, некоторые отслоившиеся клетки (, стрелки), а также те, что находятся на заднем крае оставшихся AS (, стрелка) выглядят как апоптотические клетки.«Во-первых, авторы упускают из виду ключевую статью об апоптозе в амниосерозе (Shen et al., 2013 PLoS ONE 8: e60180), в которой данные представлены в гораздо более простой для интерпретации форме (см. Shen et al.).
В наших руках и с задействованным драйвером GAL4 сигнал GFP от Аполинера не виден в ядре, но, по-видимому, ухудшен (как сообщается в Kolahgar et al., 2011 Development 138: 3021 — это та же группа который генерировал маркер Аполинера).
Для меня представленные здесь данные предполагают, что, несмотря на существенные различия в морфологии АС, уровень апоптоза у животных Y10A примерно такой же, как у эмбрионов foscrb и Y10F (может быть некоторый дополнительный апоптоз в очагах поражения в В ВИДЕ).
Если вы видите текущее видео 6, когда оно показывает время 210 мин, и вы принимаете во внимание, что эти эмбрионы были собраны, инкубированы и визуализированы одновременно, вы можете заметить, что через 10-10,5 ч после сбора яиц Уровни апоптоза в AS очень различаются у эмбрионов foscrb и foscrb Y10A . Таким образом, мы делаем вывод, что апоптоз является преждевременным у эмбрионов foscrb Y10A по сравнению с согласованными по времени контролями ( эмбрионов foscrb ).
Авторы продолжают писать: «В конце DC интернализованные клетки AS локализуются в центральной стержнеобразной структуре в эмбрионах foscrb и foscrb Y10F и впоследствии погибают от апоптоза () [as сообщалось для эмбрионов дикого типа (Reed et al., 2004)], тогда как у эмбрионов foscrb Y10A аналогичного возраста оставшиеся клетки AS полностью дезагрегированы () «. Что они, авторы, подразумевают под «ровесником»?
Мы убрали из текста термин «ровесник».Вместо этого читателя отсылают к материалам и методам для получения подробной информации о стадировании эмбриона (Результаты, второй абзац и Материалы и методы, второй абзац). Теперь мы используем «инкубировали в течение того же периода времени» или «в этот момент времени».
Для меня это полностью согласуется с интерпретацией, согласно которой морфогенез задерживается, а запрограммированная гибель клеток AS происходит «нормально» у животных со всеми тремя вариантами foscrb .
Как обсуждалось в комментарии к вашему пункту № 7, отсроченный морфогенез у эмбрионов foscrb Y10A кажется очень маловероятным.
Авторы продолжают констатировать: «Подводя итог, AS в эмбрионах foscrb Y10A распадается на части и претерпевает преждевременный апоптоз». Я согласен с тем, что AS распадается на части, но из описанных данных не ясно, что апоптоз является «преждевременным».
Мы думаем, что с добавлением деталей к тексту, фильмам, условным обозначениям, а также Материалам и методам мы дадим ответы на эти вопросы.
Далее они пишут «предполагая, что неповрежденный FBM необходим для поддержания эластичности AS».В этой статье нет ничего, что оценивало бы «эластичность» AS. Опять же, им нужно знать, что crb складывается правильно, чтобы сделать выводы о дефектном FBM (в отличие от ключевой роли crb), и они могут в лучшем случае заменить «эластичность» на «целостность».
Мы решили эту проблему (см. Подраздел «FBM Crb необходим для целостности AS», первый абзац).
Reviewer 4:
Из предыдущей работы мы знаем, что мотив связывания домена Crumbs FERM (FBM) напрямую взаимодействует с FERM-доменом Yurt (Yrt) и Expanded (Ex) и привлекает Moesin к апикальной мембране.И мы знаем, что мутация этого домена не влияет на многие другие ткани, в которых Crumbs играет роль, но имеет летальный фенотип в эмбрионе, что, как показывает это исследование, связано с сверхактивной сетью актомиозина. Получение изображений кадгеринов в реальном времени с использованием нокаут-аллеля D E-cad :: GFP на нулевом фоне crb со сформированными внедренными вариантами крошек является хорошо выполненной, систематической и ключевой частью исследования, из которого делаются выводы. полученный. Авт. Четко демонстрируют независимую роль Yrt, а затем показывают, что foscrb Y10A обнаруживают дефекты в установлении клеток актомиозинового аппарата в ситуации, когда компоненты Zonula Adherens нарушены или отсутствуют.Далее авт. Продолжают, чтобы показать, что FBM Crb является важным регулятором цитоскелетной активности и межклеточной адгезии amnioserosa и необходим для ее собственно морфогенеза. Следующая серия экспериментов является «основой» статьи, и они всесторонне рассматривают роль Crumbs FBM в динамике актомиозина и показывают, что FBM Crb регулирует динамику актомиозина в амниосерозе во время дорсального закрытия путем подавления пути Rho1. Наконец, авт. Показывают, что фенотип дорсального закрытия у эмбрионов Crb Y10A также обусловлен сниженной адгезией.Авторы тщательно проанализировали функцию Crumb через FBM. Как они указывают, мы еще не знаем, связаны ли фенотип актомиозина и фенотип пониженной адгезии или нет. Фенотипы могут быть результатом дефектов дифференцировки амниосерозы или ее прикрепления к базальной пластинке. Авторы сами поднимают этот вопрос и имеют «непоказанные данные», опровергающие такие выводы. Было бы хорошо увидеть эти данные.
Данные были включены как и.
Хотя работа убедительно демонстрирует новую функцию FBM в динамике цитоскелета, а также показывает, что Crb-регуляция морфогенеза амниосерозы включает D Moesin, Rho-GTPases, Pak класса I и комплекс SCAR-Arp2 / 3, остаются два вопроса.Как происходит эта регуляция (предлагается три пути) и почему другие эпителии, которые требуют Crumbs, по-видимому, не нуждаются в этом домене (в то же время требуя нижестоящих партнеров). Ключевой вопрос тогда заключается в том, является ли роль шоу FBM особенностью, которая детализирует аспект Crumbs, или оно говорит нам что-то более широкое о его роли в морфогенезе эпителия. Было бы хорошо увидеть обсуждение этого вопроса.
На самом деле это интересный вопрос, решение которого выходит за рамки данной работы.Мы подняли этот вопрос в конце обсуждения, но возможные сценарии оказались слишком умозрительными, и поэтому мы не вдавались в подробности по этому поводу.
[Примечание редактора: до принятия были запрошены дополнительные исправления, как описано ниже.]
Reviewer 2:
[…] Мне особенно нравится больше, чем новая находка, за генотипами легче следить, и я думаю о тонкостях Дефекты постоянного тока важны. Добавление планок погрешностей — хорошая идея, но в этом случае я бы также добавил статистическую значимость наблюдаемых различий.
Мы проанализировали статистическую значимость данных, представленных в одностороннем ANOVA, с последующим тестом множественных сравнений Даннета. Поскольку для этого анализа требуются данные, которые могут быть непрерывно изменяемыми, процентные значения сначала преобразовывались в значения arcsin, а затем анализировались с помощью GraphPad Prism. Это также описано в разделе «Статистический анализ» «Материалы и методы».
Мы считаем, что добавление большего количества символов, стрелок или звездочек к графику затруднит чтение и понимание, поэтому мы представляем результаты такого анализа в виде сводки в виде таблицы () и добавляем примечание к легенде о.
Таблица 1.
| Открытая кутикула | Спинное отверстие | Закрыто, но не вылупилось | Изогнутая личинка | 64 WT-подобнаяb 914 | 914 | 20 914 914 | 914 914 Y10A ; crb GX24 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| против | 14 | foscrb R10814 | foscrb Y10814 | 9148нс | нс | ∗ | ∗∗∗ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| и 914 85 | 23 | foscrb Y10A , DE-cad :: GFP / SCAR ∆37 , DE-cad :: GFP; crb GX24 | ∗∗∗∗ | ns | ns ∗∗∗∗ | ∗∗∗∗ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| vs | 25 | foscrb Y10A , DE-cad :: GFP / Arpc 1Q25st , DE-cad :: GFP; crb 9020X24 | нс | нс | нс | нс | нс | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 9148 cad :: GFP / +; crb 11A22 / crb GX24 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| и | 11 | 9000 9000; | 9000 9000 foscrb Y10A , DE-cad :: GFP / +; crb 11A22 / crb GX24 9 0008 | ∗∗∗∗ | ∗∗ | ∗∗ | нс | нс | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| против | 15 | rok 2 / Y / w 07 * 9000Ab; DE-cad :: GFP / +; crb 11A22 / crb GX24 | ∗ | нс | нс | нс | нс | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 9148 | 9148 9148 24A DE-cad :: GFP / +; crb 11A22 , Arp3 EP3640 / crb GX24 | нс | нс | нс | нс | ∗14141485 | 84 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5 | foscrb Y10A , GAL4 332.3 / DE-cad :: GFP; crb GX24 / crb 11A22 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| против | 7 93210AL ºC5000 | 9148 | 7 | / UAS-Crb , полная длина ; crb GX24 / crb 11A22 | нс | нс | нс | нс | нс | 8585 | 85 | 85 | , ГАЛ4 332.3 / UAS-Crb полная длина ; crb GX24 / crb 11A23 | нс | нс | нс | нс | 85 ∗∗∗∗ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 18ºC foscrb Y10A , GAL4 332,3 / UAS-Crb полная длина ; crb GX24 / crb 11A24 | нс | нс 84 нсns | и | 12 | foscrb Y10A , GAL4 332.3 / DE-cad :: GFP; crb GX24 / crb 11A22 , UAS-Rho N19 | ∗∗ | нс | нс | нс | 84 ∗|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 13 | foscrb Y10A , GAL4 332,3 / DE-cad :: GFP; crb GX24 / crb 11A22 , UAS-rok.CAT-KG | нс | нснс | ∗ | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| против | 16 | foscrb Y10A , GAL4 332.3 / DE-cad :: GFP; crb GX24 / crb 11A22 , UAS-Rac N17 | нс | нс | нс | нс | 84 нс||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| foscrb Y10A , GAL4 332,3 / DE-cad :: GFP; crb GX24 / crb 11A22 , UAS-Dmoe-myc | ns 85| ns 85 | ns ns | | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| и | 22 | foscrb Y10A , GAL4 332.3 / UAS-DE-cad, DE-cad :: GFP; crb GX24 / crb 11A22 | ∗∗∗∗ | ∗∗∗ | ns | ns 914∗∗ | ∗∗ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6 | foscrb Y10A 0 GAL 9000 GAL 9000 9000 GAL 9000 / GAL. 11A22 , UAS-Act :: RFP | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| vs | 10 | foscrb Y10A , GAL.3 / UAS-flw-HA, DE-cad :: GFP; crb GX24 / crb 11A22 , UAS-Act :: RFP | ns | ∗∗∗∗ | ns | ∗ | ∗∗∗∗ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| и | 17 | foscrb Y10A , GAL4 332,3 / UAS-DPak-AID, DE-cad :: GFP; crb GX24 / crb / crb Act :: RFP | ns | ∗∗∗∗ | ns | ns | ∗∗∗∗ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| vs | 20 | foscrb Y10A4 . G3 / UAS-Dmoe T559D , DE-cad :: GFP; crb GX24 / crb 11A22 , UAS-Act :: RFP | нс | ∗ | нс 85 84 ns ∗∗ | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 18 | foscrb, my GAL4 DE / GAL4 crb GX24 / crb 11A22 , UAS-Act :: RFP | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| vs | 19 | 332b202, GALCR3 / DE-cad :: GFP; crb GX24 / crb 11A22 , UAS-Act :: RFP | ∗∗ | ∗∗∗ | ∗∗∗∗ | ns | ∗∗ ∗ |
Я предлагаю рис. 5 — приложение 2 к основному тексту: это очень важная часть данных, которая очень хорошо фиксирует поведение актомиозиновых очагов.
Готово, теперь соответствует.
Рецензент 3:
[…] У меня все еще есть 6 проблем, которые следует исправить перед публикацией.Авторы утверждают, что Baz сохраняется на переднем крае эмбрионов Y10A. Микрофотография, которую они представляют в 2J, просто не убедительна, и их опровержение просто неубедительно. Если Baz сохраняется на переднем крае эмбрионов Y10A, несомненно, будет легко получить микрофотографию, дающую более убедительные доказательства? Сравните их локализацию Baz в диком типе с локализацией, представленной Laplante and Nilson, 2011, которую они цитируют.
Похоже, наш текст был не совсем понятным.Есть два эффекта на Baz в эмбрионах foscrb Y10A : i) мы писали, что « foscrb Y10A эмбрионы сохраняют Baz на LE тех клеток, которые не удлиняются (, стрелка)». Таким образом, как написано рецензентом, в LE нет общей потери Baz; ii) наблюдается общая редукция Baz в соединениях эмбрионов foscrb Y10A , как показано в четвертом абзаце подраздела «FBM Crb регулирует образование филоподий и организацию надклеточного актомиозинового кабеля в клетки DME »и добавлены к легенде.
Разница между нашим окрашиванием и окрашиванием, представленным в Laplante and Nilson, 2011, может быть обусловлена множеством различных факторов (концентраций, вторичных антител, времени инкубации, обработки изображений и т. Д.). Мы повторили окрашивание, но получили те же результаты, что и в упомянутых и (см.). Более высокий фон в наших окрашиваниях может быть связан с тем, что антитело в Laplante and Nilson 2011 является антителом крысы (от Wodarz, A., Ramrath, A., Kuchinke, U., and Knust, E. (1999).Nature 402, 544; но Лапланте и Нильсон не уточняют, использовали ли они крысиный анти-Baz C-термин или крысиный анти-Baz N-термин), и мы использовали кроличий анти-Baz C-термин, содержащий aa 1-297 Bazooka. белок из Drosophila . Мы также избегали увеличения контрастности, что, как мы думаем, было сделано в «и B» Лапланте и Нильсон, 2011.
In, существует 3 различных эмбриона foscrb и foscrb Y10A , окрашенных для Bazooka, соответственно. и фосфотирозин (связанный с ZA).Стрелки на контрольных эмбрионах отмечают, где Baz отсутствует в LE (как было показано на эмбрионах дикого типа, Laplante and Nilson 2011). Стрелки на эмбрионах foscrb Y10A показывают, что большинство клеток, которые не удлиняются в LE эмбрионов foscrb Y10A , сохраняют локализацию Baz на LE, как мы сообщаем в и.
Эти результаты идентичны тем, которые представлены в рукописи в, и поэтому мы не видим необходимости изменять эти цифры.
показать схемы, чтобы помочь читателю выяснить, каковы основные особенности фенотипов Y10A. Доказательств присутствия обычного миозина в филоподиях практически нет. Интересно, что хотя данные Sas показывают филоподии, ни актиновая, ни миозиновая панели не показывают филоподии очень хорошо (кажется, есть некоторые филоподии в актиновой панели E). Схема, на которой изображен «актомиозин», должна быть удалена. Актин может быть показан в филоподии актомиозинового кабеля, миозин должен отображаться только в актомиозиновом кабеле.
Исправлено в соответствии с предложением обозревателя 1.
Gorfinkiel и Martinez-Arias (2007) показали, что, хотя кадгерин изначально локализуется на переднем крае клеток DME, со временем он выводится. В моем первом обзоре я опасался, что фенотип, показанный для потери кадгерина у животных Y10A, был связан с нормальным очищением кадгерина от переднего края позже при закрытии. Авторы приводят убедительное опровержение того, что они сравнивают животных foscrb на ранней стадии с животными Y10A на ранней стадии.Им следует просто добавить предложение к исправленной рукописи, чтобы указать, что они знают, что кадгерин выводится на более поздних стадиях и что это сравнение проводится между разными генотипами на ранних стадиях закрытия. Читатели должны знать это так же, как и рецензенты.
Это было исправлено (см. Подраздел «FBM Crb регулирует образование филоподий и организацию надклеточного актомиозинового кабеля в клетках DME», шестой абзац).
В то время как пульсирующие или колеблющиеся клетки амниосерозы характеризуют дорсальное закрытие у эмбрионов дикого типа и очень интересны, мне не ясно, что для закрытия «требуются» колебания.Таким образом, колебания могут быть подавлены, а замыкание все еще происходит. Предложение, которое начинается: «Помимо удлинения ячеек DME и целостности AS, DC требует импульсных сокращений ячеек AS…» следует пересмотреть.
Мы изменили предложение, чтобы оно читалось как «возмущающая динамика актомиозина в клетках AS мешает нормальному DC», а также добавили ссылку о влиянии нарушенной динамики актомиозина на DC: Fischer et al. (2014) PLoS ONE 9, e95695.
упоминается как модель того, как FDB Crb может регулировать функцию цитоскелета.Меня озадачивают некоторые детали. В частности, модель имеет Рок и D Pak в качестве положительных регуляторов MLCK. Насколько я понимаю, все MLCK, Rok и D Pak фосфорилируют sqh напрямую, поэтому стрелки от Rok и Pak к MLCK, а не к sqh в лучшем случае вводят в заблуждение и, скорее всего, просто ошибочны (если в литературе нет данных, которые я просто не использую незнаю). Во-вторых, у Рока есть тормозящая связь с крыльями. Так ли это на самом деле? Фосфорилирует ли Rok Flapwing, подавляя его активность фосфатазы, или иным образом связывается с Flapwing, секвестрируя его активность?
Исправлено, теперь стрелки указывают, что MLCK, Rok и D Pak все фосфорилируют Sqh напрямую.Мы также добавили в схему миозин-связывающую субъединицу Drosophila ( D MBS), которая фосфорилируется Rok, что приводит к ингибированию активности фосфатазы. Мы также добавили определения сокращений в легенду рисунка.
| Недавняя литература | Уитни, Д. С., Петерсон, Ф. К., Киттелл, А. В., Эгнер, Дж. М., Прехода, К. Э. и Фолькман, Б. Ф. (2016). Привязка крошек к модулю Par-6 CRIB-PDZ регулируется Cdc42 .Биохимия 55: 1455-1461. Идентификатор PubMed: 26894406 Резюме : Par-6 представляет собой каркасный белок, который организует другие белки в комплекс, необходимый для инициации и поддержания клеточной полярности. Cdc42-GTP связывает модуль CRIB Par-6 и изменяет сродство связывания соседнего домена PDZ. Аллостерическая регуляция PDZ-домена Par-6 была впервые продемонстрирована с использованием пептида, идентифицированного при скрининге типичных карбоксиконцевых лигандов. Crumbs, мембранный белок, который локализует комплекс консервативной полярности, впоследствии был идентифицирован как функциональный партнер Par-6, который, вероятно, взаимодействует с доменом PDZ.Это исследование показывает с помощью ядерного магнитного резонанса, что Par-6 связывает карбоксиконцевой пептид Крамбса, и сообщает о кристаллической структуре комплекса PDZ-пептид. Пептид Crumbs связывает Par-6 сильнее, чем ранее изученный карбоксильный пептидный лиганд, и взаимодействует с модулем CRIB-PDZ зависимым от Cdc42 образом. Кристаллическая структура Crumbs: Par-6 выявляет специфические контакты PDZ-пептида, которые способствуют его более высокому сродству и усиленному Cdc42 связыванию. Сравнение с существующими структурами предполагает, что множественные C-концевые лиганды Par-6 отвечают на общий конформационный переключатель, который передает аллостерические эффекты связывания GTPase. | Nguyen, M.B., Vuong, L.T. и Чой, К. (2016).
Ebi модулирует рост крыльев за счет убиквитин-зависимого подавления активности Crumbs у Drosophila . Разработка 143: 3506-3513. PubMed ID: 27702784 Резюме : Передача сигналов Notch на дорсовентральной (DV) границе важна для формирования паттерна и роста крыльев у Drosophila . Белок Ebi домена WD40 участвует в регуляции передачи сигналов Notch на границе DV.Это исследование показывает, что Ebi регулирует рост крыльев, противодействуя функции трансмембранного белка Crumbs (Crb). Ebi физически связывается с внеклеточным доменом Crb (Crbext), и это взаимодействие специфически опосредуется повторами 7-8 WD40 Ebi. и домен ламинина G Crbext. Надрез на крыле в результате уменьшения уровни Ebi подавляются уменьшением функции Crb. Последовательный с этой антагонистической генетической связью, нокдаун Эби в DV граница повышает уровень белка Crb.Кроме того, Ebi требуется для подавление Crb за счет убиквитилирования. В совокупности исследование предполагает, что взаимодействие выражения Crb на границе DV и убиквитин-зависимое подавление Crb с помощью Ebi обеспечивает механизм поддержание передачи сигналов Notch во время развития крыла. | Nemetschke, L. и Knust, E. (2016). Drosophila Crumbs предотвращает эктопическую активацию Notch в развивающихся крыльях путем ингибирования лиганд-независимого эндоцитоза .Разработка 143 (23): 4543-4553. Идентификатор PubMed: 27899511 Сводка : Многие компоненты передачи сигналов апикально ограничены в эпителиальных клетках, а локализация и изобилие рецепторов являются ключевыми для морфогенеза и тканевого гомеостаза. Следовательно, контроль полярности апикобазального эпителия имеет решающее значение для правильной передачи сигналов. Notch — это повсеместно экспрессируемый апикально локализованный рецептор, который выполняет множество функций; поэтому его деятельность должна строго регулироваться.Это исследование показывает, что Drosophila Crumbs, эволюционно законсервированная детерминанта полярности, предотвращает эндоцитоз Notch в развитии крыльев посредством прямого взаимодействия между двумя белками. Эндоцитоз Notch в отсутствие Crumbs приводит к активации лиганд-независимого, зависимого от Deltex сигнального пути Notch и не требует активности лигандов Delta и Serrate или γ-секретазы. Эта функция Crumbs не связана с общими дефектами апикобазальной полярности, так как локализация других апикальных белков не изменяется.Эти данные раскрывают механизм, объясняющий, как Crumbs непосредственно контролирует локализацию и передачу мощного рецептора Notch, и добавляют еще один аспект регуляции Crumbs в активности пути Notch. Более того, данные подчеркивают тесную связь между апикальной детерминантой Crumbs, переносом рецепторов и гомеостазом ткани. | Ципин, С., Хаовей, К., Руи, X., Дандан, З. и Хуан, Х. (2017). Исследование функционального сохранения полярности внутриклеточного домена белка Крамбса .И Чуань 39 (1): 32-40. Идентификатор PubMed: 28115303 Сводка : Трансмембранный белок Crumbs (Crb) играет ключевую роль в установлении и поддержании апикально-базальной полярности клеток в эпителиальных клетках путем определения идентичности апикальной плазматической мембраны. Хотя его внутриклеточный домен содержит всего 37 аминокислот, он абсолютно необходим для его функции. У Drosophila мутации в этом внутриклеточном домене приводят к серьезным дефектам полярности эпителия и аномальному эмбриональному развитию.Внутриклеточный домен Crb демонстрирует высокую гомологию между видами от Drosophila до Mus musculus и Homo sapiens. Однако внутриклеточные домены двух белков Crb у C. elegans довольно расходятся с таковыми у дрозофилы и млекопитающих, что ставит вопрос о том, сохраняется ли функция внутриклеточного домена белка Crb у C. elegans. Используя подход геномной инженерии, это исследование заменило внутриклеточный домен Crb дрозофилы на домен Crb2 C. elegans (CeCrb2), который имеет чрезвычайно низкую гомологию с таковыми из белков Crb дрозофилы и млекопитающих.Неожиданно, замена внутриклеточного домена Crb дрозофилы на домен CeCrb2 не вызывала каких-либо аномалий в развитии эмбриона дрозофилы с точки зрения экспрессии и локализации Crb и других белков полярности и апикально-базальной полярности в эмбриональных эпителиальных клетках. Эти результаты подтверждают мнение о том, что, несмотря на обширные вариации последовательностей, все функционально важные аминокислотные остатки и мотивы межклеточного домена белков Crb полностью консервативны между Drosophila и C.elegans. | Перес-Моцкус, Г., Рока, В., Мазуни, К. и Швайсгут, Ф. (2017). Neuralized регулирует эндоцитоз и морфогенез эпителия Crumbs с помощью специфических изоформ Stardust . J Cell Biol 216 (5): 1405-1420. PubMed ID: 28400441 Резюме : Crumbs (Crb) является консервативной детерминантой идентичности апикальной мембраны, которая регулирует эпителиальный морфогенез во многих контекстах развития. Это исследование идентифицировало комплексный белок Crb Stardust (Sdt) в качестве мишени для Neuralized (Neur) убиквитинлигазы E3 у Drosophila melanogaster.Neur взаимодействует со специфическими изоформами Sdt, содержащими мотив связывания Neur (NBM), и подавляет их. Используя CRISPR (кластеризованные с регулярным интервалом короткие палиндромные повторы) -индуцированную делецию экзона, кодирующего NBM, было обнаружено, что Sdt является ключевой мишенью Neur и что Neur действует через Sdt, подавляя Crb. Далее было показано, что Neur способствует эндоцитозу Crb через NBM-содержащие изоформы Sdt. Хотя регуляция Crb с помощью Neur не является строго необходимой, она вносит вклад в ремоделирование эпителия в задней части средней кишки и тем самым облегчает трансэпителиальную миграцию примордиальных половых клеток у ранних эмбрионов.Таким образом, это исследование раскрывает новый регуляторный механизм для онтогенетического контроля Crb-опосредованного морфогенеза. | Пелликка М. и Тепасс У. (2017). Уникальные биологические профили клеток сетчатки, вызывающие болезни, миссенс-мутации в белке полярности Crumbs . J Cell Sci [Epub перед печатью]. Идентификатор PubMed: 28515229 Сводка : Мутации в человеческом гомологе CRB1 Crumbs являются основной причиной заболеваний сетчатки, которые приводят к слепоте.CRB1 — это трансмембранный белок, обнаруженный во внутреннем сегменте фоторецепторных клеток (PRC) и апикальной мембране глии Мюллера. Функция внеклеточной области CRB1 плохо изучена, хотя с ней картировано более 80 вызывающих заболевание миссенс-мутаций. В этом исследовании воссозданы четыре мутации Drosophila Crumbs (Crb), которые влияют на различные внеклеточные домены. Crb регулирует полярность и рост эпителия и способствует дифференцировке и выживанию PRC. Мутантные изоформы Crb показали замечательное разнообразие в изобилии белка, внутриклеточном распределении и способности восстанавливать недостаток эндогенного Crb, вызывать фенотип увеличения функции или способствовать дегенерации PRC.Интересно, что хотя экспрессия мутантных изоформ в значительной степени устраняет дефекты развития мутантов crb , они ускоряют дегенерацию PRC по сравнению с сетчатками, в которых отсутствует Crb, что указывает на то, что функция Crb в клеточной дифференцировке и выживании клеток зависит от различных молекулярных путей. Несколько мутантных белков Crb аномально накапливались в рабдомере и влияли на доставку родопсина, что позволяет предположить, что аномальная физиология родопсина вносит вклад в Crb / CRB1-зависимую дегенерацию сетчатки. | Оливарес-Кастинейра, И. и Ллимаргас, М. (2017). EGFR контролирует удлинение трахейной трубки Drosophila посредством регуляции внутриклеточного транспорта . PLoS Genet 13 (7): e1006882. PubMed ID: 28678789 Сводка : Развитие регулируется несколькими консервативными сигнальными путями. Среди них путь EGFR многократно используется для формирования органов и тканей, и при нарушении регуляции может привести к раку и метастазированию. Учитывая его актуальность, критически важно определить его нижележащий молекулярный механизм и понять, как он управляет клеточными изменениями.Это исследование подошло к этому вопросу в дыхательной системе дрозофилы. Была идентифицирована новая роль EGFR, ограничивающего удлинение спинного ствола трахеи. Было обнаружено, что EGFR регулирует апикальный детерминант Crumbs и регулятор внеклеточного матрикса Serpentine, два фактора, ранее известных для контроля длины пробирки. EGFR регулирует организацию эндосом, в которые загружены белки Crb и Serp. Эти результаты согласуются с ролью EGFR в регулировании маршрутов рециркуляции Retromer / WASH.Кроме того, это исследование дает новое представление о торговле Crb и его рециркуляции во время формирования органов. Эта работа связывает передачу сигналов клеток, механизмы трафика и морфогенез и предполагает, что регуляция транспорта грузов может быть общим результатом активации EGFR. | Hochapfel, F., Denk, L., Mendl, G., Schulze, U., Maassen, C., Zaytseva, Y., Pavenstadt, H., Weide, T., Rachel, R., Witzgall, R. и Krahn, MP (2017). Отличительные функции Crumbs, регулирующие щелевые диафрагмы и эндоцитоз в нефроцитах дрозофилы .Cell Mol Life Sci. PubMed ID: 28717874 Сводка : Подоциты млекопитающих, ключевые детерминанты фильтрационного барьера почек, дифференцируются от столбчатых эпителиальных клеток, и было показано, что несколько ключевых детерминант апикально-базальной полярности в обычном эпителии регулируют морфогенез и функцию подоцитов. Однако мало что известно о роли Crumbs, консервативного регулятора полярности во многих эпителиях, в формировании щелевой диафрагмы и функции подоцитов.Это исследование использовало нефроциты дрозофилы в качестве модельной системы для подоцитов млекопитающих и идентифицировало консервативную функцию белков Крамбса для клеточного морфогенеза, сборки / поддержания диафрагмы нефроцитов и эндоцитоза. Специфический для нефроцитов нокдаун Crumbs приводит к нарушению сборки / поддержания диафрагмы нефроцитов и снижению эндоцитоза, который может быть устранен Drosophila Crumbs, а также человеческими Crumbs2 и Crumbs3, которые оба экспрессируются в подоцитах человека. В отличие от внеклеточного домена, который способствует сборке / поддержанию диафрагмы нефроцитов, внутриклеточный мотив взаимодействия FERM у Crumbs важен для регуляции эндоцитоза.Более того, Moesin, который связывается с FERM-связывающим доменом Crumbs, необходим для эффективного эндоцитоза. Таким образом, это исследование описывает новый механизм развития и функции нефроцитов, который, вероятно, сохраняется в подоцитах млекопитающих. | Дас, С. и Кнуст, Э. (2018). Двойная роль внеклеточного домена Drosophila Crumbs для морфогенеза эмбриональной нейроэктодермы . Биол Открытый 7 (1). PubMed ID: 29374056 Сводка : Эпителий — это ткань с высокой поляризацией, и несколько белковых комплексов с высокой консервативной полярностью служат для установления и поддержания полярности.Трансмембранный белок Crumbs (Crb), центральный компонент белкового комплекса Crb, необходим, среди прочего, для поддержания полярности в большинстве эпителиев эмбриона дрозофилы. Однако разные эпителии проявляют разную фенотипическую тяжесть при потере crb . Использование трансгеномного подхода позволило более точно определить роль crb в различных эпителиях. В частности, представлены доказательства того, что потеря целостности эпителиальной ткани в вентральном эпидермисе у мутантных эмбрионов crb происходит из-за нарушения активности актомиозина и избыточного количества нейробластов.Было продемонстрировано, что внутриклеточный домен Crb может только частично спасти этот фенотип, в то время как он способен полностью восстанавливать целостность ткани в других эпителиях. На основании этих результатов предполагается двойная роль внеклеточного домена Crb в вентральной нейроэктодерме. Во-первых, он необходим для апикального обогащения белка Crb, который, в свою очередь, регулирует активность актомиозина и тем самым обеспечивает целостность ткани; и, во-вторых, внеклеточный домен Crb стабилизирует рецептор Notch и тем самым обеспечивает правильную передачу сигналов Notch и спецификацию правильного количества нейробластов. | Баджур, А. Т., Айер, К. В. и Кнуст, Э. (2019). Цитокортекс-зависимая динамика Drosophila Crumbs контролирует стабильность и напряжение соединений во время ретракции зародышевой ленты . J Cell Sci. PubMed ID: 31300472 Сводка : Во время морфогенеза эпителий претерпевает динамические перестройки, которые требуют постоянного ремоделирования соединений и формы клеток, но в то же время механизмов, сохраняющих полярность клеток и целостность ткани.Апико-базальная полярность является ключом к локализации механизмов, которые позволяют изменять форму клеток. Эволюционно консервативный белок Drosophila Crumbs является критическим для поддержания апико-базальной полярности и целостности эпителия. Каким образом Crumbs сохраняется в динамично развивающемся эмбрионе, остается в значительной степени неизвестным. В этом исследовании были применены методы количественной флуоресценции, чтобы показать, что во время ретракции зародышевой полосы динамика Крамбса коррелирует с морфогенетической активностью эпителия. Генетические и фармакологические изменения показали, что мобильный пул Crumbs настраивается актомиозиновой корой в зависимости от стадии.Стабилизация Crumbs на плазматической мембране зависит от правильной связи с актомиозиновой корой через интактный сайт связывания FERM-домена в его внутриклеточном домене, потеря которого приводит к увеличению напряжения в соединениях и более высокому обмену DE-кадгерина, что приводит к нарушению перестройки соединений. Эти данные определяют Crumbs как посредника между полярностью и регуляцией соединений, чтобы управлять ремоделированием эпителия в ответ на изменения активности актомиозина. | Латтнер, Дж., Ленг, В., Кнут, Э., Бранкачк, М., Флорес-Бенитес, Д. (2019). Crumbs организует транспортный механизм, регулируя апикальные уровни PI (4,5) P2 у Drosophila . Elife 8. Идентификатор PubMed: 31697234 Сводка : Эффективный векторный внутриклеточный транспортный механизм зависит от хорошо установленной апико-базальной полярности и является предпосылкой для функции секреторного эпителия. Несмотря на обширные знания об отдельных путях торговли людьми, мало что известно о механизмах, координирующих их временную и пространственную регуляцию.Это исследование сообщает, что белок полярности Crumbs необходим для фосфолипидного гомеостаза апикальной плазматической мембраны и эффективной апикальной секреции. Привлекая betaHeavy-Spectrin и MyosinV к апикальной мембране, Crumbs поддерживает Rab6-, Rab11- и Rab30-зависимый транспорт и регулирует липидные фосфатазы Pten и Ocrl. Нокдаун Crumbs приводит к увеличению апикальных уровней PI (4,5) P2 и образованию нового, обогащенного Moesin и PI (4,5) P2 апикального мембранного мешочка, содержащего микроворсинки-подобные структуры.Эти результаты идентифицируют Crumbs как важный центр, необходимый для поддержания организации апикальной мембраны и физиологической активности личиночной слюнной железы. | Краут, Р. С., Кнут, Э. (2019). Изменения в эндолизосомной организации определяют пред-дегенеративное состояние у мутанта Drosophila retina крохи. PLoS One 14 (12): e0220220. Идентификатор PubMed: 31834921 Сводка : Мутации в гене crumbs (crb) полярности эпителия приводят к дегенерации сетчатки у дрозофилы и человека.Общая морфология сетчатки и ее ухудшение у мутантов Drosophila crb хорошо охарактеризованы, но клеточная биологическая природа дегенерации не совсем понятна. Дегенеративные состояния сетчатки и других участков нервной системы часто связаны с дефектами внутриклеточных путей деградации. Однако до сих пор эффекты crb на эндолизосомную систему или на пространственную организацию этих компартментов в фоторецепторных клетках не описаны.Это исследование поэтому спрашивает, проявляют ли фоторецепторы у мутантов crb изменения в эндолизосомных компартментах в преддегенеративных условиях, когда сетчатка все еще остается морфологически интактной. Данные, представленные в этом исследовании, показывают, что уже задолго до начала дегенерации Arl8, Rab7 и Atg8-несущие эндолизосомные и аутофагосомные компартменты претерпевают изменения в морфологии и позиционировании относительно друг друга в мутантных сетчатках crb . Предполагается, что эти изменения могут быть ранними признаками предрасположенного к дегенерации состояния сетчатки глаза. | Агилар-Арагон, М., Флетчер, Г. и Томпсон, Б. Дж. (2019). Моторные белки цитоскелета Dynein и MyoV направляют апикальный транспорт Crumbs . Dev Biol. Идентификатор PubMed: 31881198 Резюме : Крохи (Crb у Drosophila; CRB1-3 у млекопитающих) являются трансмембранной детерминантой полярности эпителиальных клеток и регулятором передачи сигналов Hippo. Crb обычно локализован в апикальных межклеточных контактах, чуть выше слипчивых соединений, но как апикальный транспорт Crb регулируется в эпителиальных клетках, остается неясным.В этом исследовании использовался фолликулярный эпителий дрозофилы, чтобы продемонстрировать, что поляризованный транспорт Crb опосредуется транспортом по микротрубочкам с помощью моторного белка Dynein и по актиновым филаментам с помощью моторного белка Myosin-V (MyoV). Блокирование транспорта Crb-содержащих везикул с помощью Dynein или MyoV ведет к накоплению Crb в эндосомах Rab11, а не к апикальной доставке. Заключительные этапы доставки и стабилизации Crb на плазматической мембране требуют комплекса экзоцисты и трех апикальных белков домена FERM — Merlin, Moesin и Expanded, одновременная потеря которых нарушает апикальную локализацию Crb.Соответственно, нокаутированная делеция Crb FERM-связывающего мотива (FBM) также нарушает апикальную локализацию. Наконец, избыточная экспрессия Crb бросает вызов этой системе, создавая сенсибилизированный фон для идентификации компонентов, участвующих в поляризации цитоскелета, перемещении апикальной мембраны и стабилизации Crb в апикальном домене. | Хван, Дж. Х., Вуонг, Л. Т. и Чой, К. У. (2020). Crumbs, Galla и Xpd необходимы для регуляции кинезина-5 в митозе и росте органов у Drosophila .J Cell Sci. PubMed ID: 32501288 Сводка : Xeroderma Pigmentosum D (XPD) — это многофункциональный белок, участвующий в транскрипции, репарации ДНК и сегрегации хромосом. У Drosophila Xpd взаимодействует с Crumbs (Crb) и Galla, чтобы регулировать митоз во время эмбриогенеза. Неизвестно, как эти белки связаны с митозом. Это исследование показывает, что Crb, Galla-2 и Xpd регулируют ядерное деление у синцитиального эмбриона путем взаимодействия с Klp61F, митотическим кинезином-5 дрозофилы, связанным с биполярными веретенами.Crb, Galla-2 и Xpd физически взаимодействуют с Klp61F и совместно локализуются на митотических веретенах. Нокдаун любого из этих белков приводит к аналогичным митотическим дефектам. Эти фенотипы восстанавливаются за счет сверхэкспрессии Klp61F, что позволяет предположить, что Klp61F является основным эффектором. Митотические дефекты galla-2 RNAi подавляются сверхэкспрессией Xpd, но не наоборот. Истощение Crb, Galla-2 или Xpd приводит к снижению уровней Klp61F. Снижение функции протеасом восстанавливает уровни Klp61F и подавляет митотические дефекты, вызванные нокдауном Crb, Galla-2 или Xpd.Кроме того, рост глаз регулируется Xpd и Klp61F. Следовательно, это исследование предполагает, что Crb, Galla-2 и Xpd взаимодействуют для поддержания уровня Klp61F во время митоза и роста органов. | Юнг, Дж., Йом, Э. и Чой, К. У. (2020). Ciao1 взаимодействует с Crumbs и Xpd, чтобы регулировать рост органов у Drosophila . Cell Death Dis 11 (5): 365. Идентификатор PubMed: 32404863 Сводка : Ciao1 является компонентом комплекса сборки цитозольного кластера железа-серы (CIA) вместе с MMS19 и MIP18.Xeroderma pigmentosum группа D (XPD), ДНК-геликаза, участвующая в регуляции клеточного цикла и транскрипции, является мишенью CIA для модификации железо-сера (Fe / S). Функция Ciao1 и Xpd in vivo у развивающихся животных редко изучалась. Это исследование показывает, что Ciao1 взаимодействует с Crumbs (Crb), Galla и Xpd, чтобы регулировать рост органов у дрозофилы. Аномальный рост глаза за счет сверхэкспрессии внутриклеточного домена Crb (Crb (intra)) подавляется за счет снижения уровня Ciao1. Потеря Ciao1 или Xpd вызывает аналогичное нарушение роста органов.РНКи-нокдаун как Ciao1, так и Xpd обнаруживает сходные фенотипы, как Ciao1 или только Xpd RNAi, что указывает на их функцию в пути. Дефекты роста, вызванные Ciao1 RNAi, подавляются сверхэкспрессией Xpd. Ciao1 физически взаимодействует с Crb (intra), Galla и Xpd, поддерживая их генетические взаимодействия. Примечательно, что дефекты Xpd RNAi также могут подавляться сверхэкспрессией Ciao1, что подразумевает взаимную регуляцию между двумя генами. Мутантные клоны Ciao1 в имагинальных дисках показывают пониженные уровни циклина E (CycE) и ассоциированного со смертью ингибитора апоптоза 1 (Diap1).Мутантные клоны Xpd обладают сходным восстановлением CycE и Diap1. Следовательно, нокдаун Ciao1 и Xpd с помощью РНКи показывает повышенную апоптотическую гибель клеток. Кроме того, избыточной экспрессии CycE достаточно для восстановления дефектов роста Ciao1 RNAi или Xpd RNAi. Интересно, что избыточная экспрессия Diap1 в мутантных клонах Ciao1 индуцирует экспрессию CycE, подтверждая, что снижение CycE в мутантных клетках Ciao1 является вторичным по отношению к потере Diap1. Взятые вместе, это исследование раскрывает новые роли Ciao1 и Xpd в выживании и росте клеток посредством регулирования уровня Diap1 во время развития органов. | Шард, К., Луна-Эскаланте, Дж. И Швайсгут, Ф. (2020). Тканевая координация перехода эпителия к нервным стволовым клеткам в оптической доле дрозофилы требует Neuralized . J Cell Biol 219 (11). PubMed ID: 32946560 Сводка : Многие ткани продуцируются специализированными клетками-предшественниками, происходящими из эпителия через эпителиально-мезенхимальный переход (EMT). Большинство исследований до сих пор было сосредоточено на ЭМП с участием отдельных или изолированных групп клеток.Это исследование описывает процесс, подобный EMT, который требует координации на тканевом уровне. Этот EMT-подобный процесс происходит вдоль непрерывного фронта в нейроэпителии зрительной доли Drosophila с образованием нервных стволовых клеток (NSC). Возникающие NSCs остаются эпителиальными и апикально сужаются перед асимметричным делением с образованием нейронов. Апикальное сужение связано с импульсами сократительного миозина и включает RhoGEF3 и подавление комплекса Crumbs с помощью Neuralized убиквитинлигазы E3. Анизотропия на сложных уровнях Крамбса также приводит к накоплению соединительного миозина.Нарушение регуляции Crumbs с помощью Neuralized пониженного соединительного миозина и привело к неточности интеграции возникающих NSC в переднюю часть. Таким образом, Neuralized способствует плавному прогрессированию фронта дифференцировки путем связывания ремоделирования эпителия на тканевом уровне с приобретением судьбы NSC. | Бхагаватула, С. и Кнуст, Э. (2020). Предполагаемая структура стволовых петель у Drosophila crumbs необходима для локализации мРНК в эпителии и клетках зародышевой линии .J Cell Sci. Идентификатор PubMed: 33310910 Резюме : Crumbs (Crb) представляет собой эволюционно консервативный трансмембранный белок, локализованный в апикальной мембране эпителиальных клеток. Потеря или неправильная локализация Crb часто связана с нарушением полярности апико-базальных клеток. crb мРНК также апикально обогащена эпителиальными клетками и, как показано в этом исследовании, накапливается в ооците развивающихся яичных камер. Элемент локализации (LE) мРНК crb был сужен до 47 нуклеотидов, образующих предполагаемую структуру петля-стебель, что позволяет предположить, что он распознается эгалитаристом (Egl).Мутации в консервативных нуклеотидах отменяют апикальный транспорт. На обогащение мРНК crb в ооците влияет камера мутанта egl для яиц. Геномная делеция на основе CRISPR локуса crb , которая включает в себя LE, нарушает асимметричную локализацию мРНК crb в эпителии и предотвращает ее накопление в ооците на ранних стадиях оогенеза, но не влияет на локализацию белка Crb в эмбриональном и фолликулярном эпителии. Однако у мух, лишенных LE, в кормовых клетках наблюдается эктопическая экспрессия белка Crb.Эти данные предполагают дополнительную роль 3′-UTR Drosophila в регуляции трансляции тканеспецифическим образом. |
php — Простая динамическая цепочка навигации
Я начал с кода Доминика Барнса, включил отзывы cWoDeR, и у меня все еще были проблемы с хлебными крошками на третьем уровне, когда я использовал подкаталог. Поэтому я переписал его и включил приведенный ниже код.
Обратите внимание, что я настроил структуру своего веб-сайта таким образом, что страницы, которые будут подчиняться (ссылаться) на страницу на корневом уровне, настроены следующим образом:
Создайте папку с ТОЧНЫМ именем файла (включая заглавные буквы) без суффикса, как папку на корневом уровне
поместить все подчиненные файлы / страницы в эту папку
(например, если нужно согласовать страницы для клиентов.php:
создайте папку с именем Customers на том же уровне, что и Customers.php
добавьте файл index.php в папку Customers, который перенаправляет на вызывающую страницу для папки (код см. Ниже)
Эта структура будет работать для нескольких уровней вложенных папок.
Просто убедитесь, что вы следуете описанной выше файловой структуре И вставляете файл index.php с кодом, показанным в каждой подпапке.
Код в индексе.php в каждой подпапке выглядит так:
Перенаправлено
Если вы используете корневой каталог сервера в качестве корневого веб-сайта (т.е. / var / www / html), тогда установите $ root_dir = «»: (НЕ оставляйте завершающий «/» внутри). Если вы используете подкаталог для своего веб-сайта (например, / var / www / html / web_root, тогда установите $ root_dir = «web_root /»; (замените web_root фактическим именем вашего веб-каталога) (обязательно включите завершающий /)
во всяком случае, вот мой (производный) код:
в качестве корневого каталога для этого веб-сайта
// Чтобы включить это, введите имя подкаталога, используемого в качестве корневого веб-каталога
// в переменной $ directory2 ниже
// Не забудьте включить завершающий "/" в конце имени каталога
// например, используйте $ directory2 = "this_folder /";
// НЕ используйте $ directory2 = "this_folder";
// Если вы действительно ИСПОЛЬЗУЕТЕ / var / www / html в качестве корневого каталога,
// просто установите $ directory2 = "" (пусто)
// без завершающего "/"
// Эта функция возьмет $ _SERVER ['REQUEST_URI'] и построит хлебную крошку на основе текущего пути пользователя
функция хлебные крошки ($ separator = '& raquo;', $ home = 'Home')
{
// Это устанавливает подкаталог как web_root (если вы хотите использовать подкаталог)
// Если вы не используете подкаталог web_root, установите $ directory2 = ""; (БЕЗ завершающих /)
$ directory2 = "каталог-сети /";
// Получение REQUEST_URI (/ путь / к / файлу.php), разбивает строку (используя '/') в массив, а затем отфильтровывает любые пустые значения
$ path = parse_url ($ _ SERVER ['REQUEST_URI'], PHP_URL_PATH);
$ path_array = array_filter (взрыв ('/', $ path));
// Эта строка кода позволяет использовать подпапку (/ var / www / html / ) в качестве root
// Это удаляет первый элемент в пути массива, поэтому он не повторяется
если ($ directory2! = "")
{
array_shift ($ path_array);
}
// Это создаст наш "базовый URL" ... Также учитывается HTTPS :)
$ base = ($ _SERVER ['HTTPS']? 'https': 'http').': //'. $ _SERVER ['HTTP_HOST']. '/'. $ directory2;
// Инициализируем временный массив с помощью наших хлебных крошек. (начиная с нашей домашней страницы, которая, как я предполагаю, будет базовым URL-адресом)
$ breadcrumbs = Array (" $ home ");
// Получаем индекс для последнего значения в нашем массиве путей
$ last = end ($ path_array);
// Инициализируем счетчик
$ crumb_counter = 2;
// Строим остальные панировочные сухари
foreach ($ path_array как $ crumb)
{
// Наш «заголовок» - это текст, который будет отображать имя файла без символа.суффикс
// Если нет "." в крошке это каталог
if (strpos ($ crumb, ".") == false)
{
$ title = $ крошка;
}
еще
{
$ title = substr ($ crumb, 0, strpos ($ crumb, "."));
}
// Если мы не на последнем индексе, то создаем гиперссылку
если ($ крошка! = $ последняя)
{
$ call_page_array = array_slice (array_values (array_filter (explode ('/', $ path))), 0, $ crumb_counter, false);
$ call_page_path = "/".implode ('/', $ call_page_array). ". php";
$ breadcrumbs [] = "". $ title. "";
}
// В противном случае просто отображаем заголовок
еще
{
$ breadcrumbs [] = $ title;
}
$ crumb_counter = $ crumb_counter + 1;
}
// Собираем наш временный массив (кусочки хлеба) в одну большую строку :)
return implode (разделитель $, $ панировочные сухари);
}
//
// ')? >
//
?>
Что такое хлебные крошки и как оно упрощает навигацию? (Примеры)
Разве мы все не были в тематическом парке и заблудились, пытаясь найти выход? Мы не примем этого, но есть крохотное деревце разочарования, которое вот-вот поднимет голову.
Мы отчаянно оглядываемся, спрашиваем прохожего, и они указывают нам на карту, которая говорит нам, где мы находимся в парке.
Внезапно вы знаете, где находитесь и где находятся вход и выход, в зависимости от вашего местоположения. Паника уходит! И мы пошли гав гав!
В навигации по сайту «хлебные крошки» делают нечто подобное.
«Панировочные сухари» помогут вам ориентироваться и точно показывают, где вы находитесь на веб-сайте.
Вот что вы узнаете из этого сообщения:
- Что такое навигация по хлебным крошкам?
- Наша эволюционная потребность в навигации по хлебным крошкам
- Типы панировочных сухарей для веб-сайтов
- 5 преимуществ панировочных сухарей
- 10 лучших практик использования панировочных сухарей
- Проблема с навигацией по хлебным крошкам
- Последнее слово о панировочных сухарях
«Хлебные крошки» — это вспомогательное средство навигации, которое помогает пользователям легко понять связь между их местоположением на странице (например, страница продукта) и страницами более высокого уровня (например, страница категории).Этот термин заимствован из сказки о Гензеле и Гретель, где дети бросают след из панировочных сухарей, чтобы проследить свой путь назад.
С помощью панировочных сухарей, если вы дошли до страницы, на которой не хотите быть, вы можете легко найти путь назад или вернуться на шаг или два назад и начать все сначала.
Панировочные сухари не предназначены исключительно для веб-сайтов. Microsoft представила хлебные крошки в Windows Vista, и с тех пор эта функция присутствует в каждой редакции Windows.
Сравнение Windows XP (вверху) и Windows 7 (внизу).Обратите внимание на беспорядок на пути в Windows XP, тогда как в Windows 7 отображаются хлебные крошки, которые позволяют пользователям визуализировать путь и при необходимости проследить его. Изображение предоставлено: GeekGirl’s Наша потребность в панировочных сухарях имеет эволюционную причинуПитер Пиролли обнаружил интересные параллели между тем, как животные добывают себе пищу, и тем, как люди ищут полезную информацию. По сути, все мы живем по эвристическому правилу — не тратьте больше энергии на поиски пищи, чем то, что дает еда.Рассмотрим, как мы ищем информацию в Интернете: мы что-то ищем, поисковая система выдает много результатов. Мы сканируем ссылки и не переходим по ним, если не улавливаем подсказки (заголовок SEO, мета и т. Д.) О том, что ссылка предоставит нам нужную информацию. Это мало чем отличается от зверя, охотящегося за добычей в саванне; зверь будет заряжаться только тогда, когда след запаха достаточно силен, чтобы оправдать энергию, которая должна быть потрачена на преследование.
На веб-сайте при наличии нескольких навигационных следов лучше всего, если пользователи могут четко идентифицировать след, ведущий к тому, что они хотят.Без этого они могут сдаться и отскочить.
Типы панировочных сухарей в дизайне веб-сайтов Хлебные крошки на основе местоположения или иерархииНа базовом уровне люди зарабатывают себе на хлеб, но жаждут панировочных сухарей.
Хлебные крошки на основе местоположения помогают пользователю перейти к более широкой категории (странице более высокого уровня) со страницы, на которой он находится. Представьте, что кто-то ищет в Google пару ботинок для пустыни, находит результат поиска и попадает на одну из страниц вашего продукта.Им не нравится то, что они видят, и они хотят изучить другие варианты. Как человек возвращается на главную страницу категории (мужская обувь) или страницу подкатегории (мужская обувь — обувь для пустыни)?
С помощью хлебных крошек на основе иерархии пользователь может легко увидеть, где они находятся в архитектуре вашего сайта, и легко перейти на страницу более высокого уровня. Посмотрите, как это делает Best Buy.
Панировочные сухари на основе пути или историиОни служат той же цели, что и кнопка «Назад» в браузере, позволяя пользователю вернуться на одну из предыдущих страниц, которые он посещал.Это может быть полезно, когда пользователь перешел на страницу продукта после применения нескольких фильтров на странице категории. С помощью хлебных крошек на основе истории пользователь может быстро перейти к одной из предыдущих страниц в своем путешествии, сохранив все свои выборы без изменений.
Однако наиболее распространенным применением хлебных крошек на основе истории является использование ссылки «Назад к результатам».
Панировочные сухари на основе атрибутовНавигационные цепочки на основе атрибутов просто показывают вам атрибуты, выбранные пользователем на странице.Такие хлебные крошки находят применение на страницах категорий электронной коммерции, предлагаемых большинством платформ электронной коммерции, где пользователи могут выбирать атрибуты для фильтрации результатов поиска.
5 преимуществ использования панировочных сухарейПервоначально задуманный как простое средство навигации, сегодня хлебные крошки стали влиять на многие аспекты взаимодействия с пользователем в Интернете.
Поощряет просмотр и снижает количество отказовЕсли пользователь достигает страницы продукта, которая ему не интересна, он либо откажется от нее, либо вернется на страницу категории, чтобы начать все сначала.Хлебные крошки побуждают пользователя начать все сначала и пока не откажутся. В исследовании, проведенном Hull, S.S. (2004) [1] , было обнаружено, что пользователи, получившие инструкции по использованию хлебных крошек, выполняли задачи намного быстрее, чем пользователи, которые не использовали хлебные крошки. Для пользователей, которые ежедневно пользуются Интернетом (даже в интрасети, например, в организации), использование хлебных крошек может значительно повысить производительность и сэкономить время.
Связанное сообщение : 14 способов уменьшить отказы и повысить вовлеченность на вашем сайте электронной коммерции
Повышает удобство поиска вашего веб-сайтаПитер Морвилл, один из отцов-основателей информационной архитектуры Всемирной паутины, разработал соты взаимодействия с пользователем, которые иллюстрируют семь основных аспектов хорошего взаимодействия с пользователем.Помогая пользователям легко ориентироваться и находить то, что они ищут, хлебные крошки помогают повысить удобство поиска веб-сайта.
Связанное сообщение : 15 основных элементов эффективного продукта электронной коммерции Страница
Панировочные сухари обеспечивают преимущества для SEOGoogle объявил [2] , что они заменяют URL в своих результатах поиска на имя сайта и путь навигации хлебных крошек. Это происходит сейчас и для мобильных устройств, а в ближайшие дни может появиться и для настольных компьютеров.Google считает, что использование реального имени веб-сайта поможет улучшить удобство использования, а ссылки на хлебные крошки покажут пользователям, куда они будут перенаправлены при нажатии на результат поиска.
Изображение предоставлено: Search Engine LandВеб-мастера могут использовать разметку схемы для Breadcrumbs [3] для более эффективного взаимодействия с поисковой системой. Панировочные сухари становятся для вас дополнительным способом лучше объяснить поисковым системам, о чем ваши страницы, и получить это небольшое дополнительное преимущество для SEO.Они также действуют как дополнительные ссылки в результатах поиска, чтобы дать вам больше шансов для перехода по ссылке.
Потому что нет веской причины не использовать ихЯкоб Нильсен говорит, что рекомендует панировочные сухари с 1995 года [4] . Причины просты.
Панировочные сухари никогда не вызывают проблем при пользовательском тестировании. Пользователи обращают внимание на панировочные сухари только 30% времени; когда они это делают, это помогает, и даже когда они этого не делают, это не причиняет никакого вреда.
Они занимают очень мало места на странице, поэтому недвижимость не может быть причиной, по которой вы отказываетесь от использования панировочных сухарей.
Панировочные сухари на протяжении многих лет остаются довольно неизменными по своему внешнему виду. Такое долгое знакомство делает его мгновенно узнаваемым и, следовательно, полезным.
Панировочные сухари помогают снизить беспокойство пользователей«Хлебные крошки» могут уменьшить беспокойство пользователей о том, чего ожидать, показывая им весь след процесса. Например, настройка учетной записи на VWO состоит из нескольких шагов. Используя хлебные крошки, мы можем точно сказать пользователю, сколько шагов задействовано в процессе, и позволить ему вернуться к любому из предыдущих шагов в любой момент.Это дает пользователю представление об обязательствах (времени и усилиях), требуемых от него для настройки процесса. Когда кто-то инициирует попытку, такие визуальные подсказки могут послужить сильным мотиватором для завершения процесса.
[5] ведутся споры о том, следует ли называть эту функцию «индикатором выполнения» или «навигационной цепочкой». Но народная мудрость подсказывает, что, если след позволяет пользователям вернуться к предыдущему шагу, это все равно, что быть хлебной крошкой.
10 Рекомендуемых практик использования панировочных сухарейОбычно хлебные крошки появляются в горизонтальной линии, показывающей путь от страницы самого высокого уровня (домашней) до текущей страницы, на которой находится пользователь.Ниже приведен список из 10 лучших практик, некоторые из которых рекомендованы Стивом Кругом, автором книги «Не заставляйте меня думать» и богом веб-дизайна.
Всегда показывать весь путь : Предоставьте пользователям контекст. Хорошая навигационная цепочка действует как шоры, удерживая пользователя сосредоточенным на текущей задаче.
Начать с домашней страницы : Лучше всего использовать маршруты, когда они показывают путь от одного конца до другого. Включение домашней страницы в трейл действует как сильный якорь, давая пользователям четкое ощущение ориентации.
Используйте «>» в качестве разделителя между уровнями. : Он работает, потому что работал всегда. Значок «>» быстро показывает взаимосвязь между страницами более высокого уровня и страницами более низкого уровня. Якоб Нильсен изначально рекомендовал использовать двоеточие [6] (:) в панели навигации по хлебным крошкам. После пользовательского тестирования он изменил свою рекомендацию на использование «>», потому что оно легко представляет связь между страницами более высокого уровня и страницами более низкого уровня.
Поместите крошки наверх : панели навигации обычно размещаются в самом верху веб-сайта.Поскольку хлебные крошки работают как вспомогательное средство навигации, их следует размещать над содержимым. Группа Nielsen Norman провела пользовательские тесты и обнаружила, что пользователи ожидают появления хлебных крошек в верхних [7] страницах. Хорошее место для панировочных сухарей — под основной панелью навигации и над заголовком страницы.
Покажите немного контраста : Контраст помогает хлебным крошкам выделиться как важная функция. Поскольку они занимают меньше места и в основном основаны только на тексте, контраст становится решающим фактором их успеха.
Используйте мелкий шрифт : Использование мелкого шрифта помогает сообщить пользователю об относительной важности хлебных крошек по отношению к главной панели навигации. Его никогда не следует путать с основным навигационным средством.
Полужирным шрифтом последний элемент : Последний элемент (текущая страница) выделен жирным шрифтом, чтобы выделить его и сообщить пользователю: «Это то место, где вы сейчас находитесь».
Не делайте гиперссылку на последний элемент : Зачем вам гиперссылка на текст, ведущий на ту же страницу? Это сбивает пользователя с толку.
Не используйте хлебные крошки на главной странице : домашняя страница — это то место, где начинается пользовательский путь, нет смысла отображать хлебные крошки на вашей домашней странице.
Использовать полные заголовки страниц в хлебных крошках : имеет смысл включать полные заголовки страниц в хлебные крошки, чтобы пользователи точно знали, куда ведет каждая из гиперссылок. Но из-за длинных заголовков страниц или других причин страницы некоторые предпочитают обрезать заголовки страниц, используя многоточие (см. Изображение ниже).В таких случаях лучше опускать такие фразы, как «Вы здесь», чтобы освободить место для наиболее важных элементов, которыми являются сами заголовки страниц.
URL-адрес страницы должен отражать цепочку «хлебных крошек» : URL-адрес страницы дает представление об архитектуре сайта. Важно обеспечить согласованность между URL-адресом и хлебными крошками, поскольку в противном случае пользователи могут получать несогласованные сообщения от обоих этих элементов.
Когда не следует использовать панировочные сухари?Это сложный вариант, потому что панировочные сухари в большинстве случаев безвредны.Но есть нюанс. Если ваш сайт содержит всего несколько страниц без множества уровней иерархии, хлебные крошки могут быть бесполезны. Включать панировочные сухари в этих случаях нет необходимости и они могут сбить с толку читателя.
Если вы действительно посмотрите под лесную подстилку, панировочные сухари не все идеальны. Проблема с навигацией по хлебным крошкам
Даже после более чем десятилетнего существования хлебные крошки по-прежнему не считаются лучшей практикой. На то есть много причин:
1. Люди до сих пор не уверены, стоит ли это затраченных усилий: это не функциональность, которую нужно исправить или сломать. Здесь нет споров.
2. Существует мнение, что хлебные крошки «необходимы» только в том случае, если ваша информационная архитектура находится в упадке. Если пользователи могут легко перемещаться по вашему веб-сайту, они будут знать, где они находятся, даже если вы их не показываете. По сути, эта школа мысли считает, что хлебные крошки — это запасной вариант, когда ваша архитектура испорчена. Поэтому они спрашивают: если вы собираетесь тратить время и силы, почему бы не улучшить вашу информационную иерархию, чем инвестировать в хлебные крошки?
Проблемы с хлебными крошками на основе историиДля динамического создания хлебных крошек на основе истории необходимо получить доступ к файлам cookie браузера.Это требует дополнительного кода и усилий. Дополнительные усилия будут иметь смысл, если преимущества перевешивают усилия. Но так ли это?
Панировочные сухари на основе истории просто дублируют функциональность кнопки «Назад».
Нет преимущества SEO : Поскольку хлебные крошки должны основываться на истории пользователей, боты поисковых систем никогда не увидят его и не имеют преимуществ SEO
Бесполезно для всех : Многие пользователи напрямую попадают на страницы в глубине сайта через поисковые системы; для таких людей историческая тропа не приносит пользы.
Compounds Error : Для людей, которые попадают на страницу методом проб и ошибок, будет бессмысленно иметь неоптимальную историю навигации в виде хлебных крошек.
Однако стоит отметить, что последний пункт проистекает из неправильной информационной архитектуры сайта в целом. Если пользователи не могут легко определить самый простой путь к странице, возможно, лучше инвестировать в оптимизацию архитектуры сайта, чем возиться с хлебными крошками.
Проблемы с навигационными сухарямиДля многоиерархических сайтов, таких как большинство веб-сайтов электронной коммерции нового поколения, иерархия более плоская, и каждый выбор продукта может принадлежать к нескольким категориям.Например, для кого-то, кто хочет купить мешочек с фасолью, след панировочных сухарей может выглядеть как
.Дом> Меблировка> Гостиная> Стулья> Одноместные> Сумка-мешок «The Dude Abides»
или
Домой> Меблировка> Гостиная> Стулья> Повседневные> Сумка-мешок «The Dude Abides»
Что вы выберете для отображения?
Давайте рассмотрим сначала страницу продукта .
Для страницы продукта один разумный способ — позвонить и узнать, какой тип хлебных крошек лучше всего отображать.Самый простой или наиболее используемый путь навигации — хороший соперник.
Если страница вашей категории позволяет пользователям выбирать атрибуты продукта, было бы полезно иметь наиболее подходящую навигационную цепочку навигации и ссылку «вернуться к результатам» (исторический путь) на странице продукта. Сделать выбор — дело непростое. Для пользователей, которые прошли через это, перешли на страницу продукта, не совсем удовлетворены своим выбором и хотят проверить другой продукт, соответствующий атрибуту, кнопка возврата в браузере может вызвать беспокойство.Это связано с тем, что пользователи не могут быть уверены, что браузер запомнит выбранные ими атрибуты. Опция «Вернуться к результатам» может успокоить посетителя и удержать его в воронке конверсии.
Вернуться к результатам | На главную> Мебель> Гостиная> Бин-мешки> Бин-сумка «The Dude Abides»
На странице продукта Macy отображаются хлебные крошки на основе местоположения и истории. Если пользователю не нравится продукт, он может легко вернуться на страницу и выбрать другой продукт.Теперь поговорим о странице категории .
Для страницы категории, если вы предоставляете пользователям выбор атрибутов продукта, таких как «бренд» или «размер», другой способ использования хлебных крошек — отображать как стандартный путь навигации, так и атрибуты вместе, но должным образом дифференцированно.
Начало> Меблировка> Стулья | Одноместный *
На странице категории на Flipkart отображаются хлебные крошки на основе местоположения и атрибутов. Это помогает пользователям быстро понять, как они попали сюда и какие фильтры уже были применены. Панировочные сухари: вторичный по статусу, первичный по значениюПанировочные сухари существуют уже более десяти лет.Но они никогда не привлекали особого внимания из-за своего постоянного статуса «вторичной панели навигации». Несмотря на свой статус, хлебные крошки могут быть мощным элементом для улучшения взаимодействия с пользователем и привлечения внимания. С точки зрения бизнеса, это помогает как в привлечении (подумайте о SEO), так и в конверсии (упрощает навигацию по сайту).
Чтобы пояснить суть дела, позвольте мне процитировать некоторые цифры из исследования Baymard, проведенного двухлетней давностью. После сравнительного исследования 40 сайтов электронной коммерции выяснилось, что 68% неоптимально использовали навигацию Breadcrumb.23% вообще не имели элемента «хлебные крошки». Некоторые считают хлебные крошки архаичным элементом дизайна, не имеющим необходимости или приоритетом в современном мире, в то время как для других это жемчужина, которая занимает мало места и требует небольших усилий, но вносит огромный вклад в общее впечатление пользователя.
Так стоит ли иметь на вашем сайте электронной коммерции панировочные сухари? Обращайтесь к своим пользователям за ответом, принесут ли они пользу? Я думаю, что это единственный вопрос, который стоит задать.
Часто задаваемые вопросы о панировочных сухарях
Что такое меню из хлебных крошек?Меню «хлебных крошек» на веб-сайте дает новому посетителю ориентацию и показывает ему или ему, где именно он находится на веб-сайте.
Какая польза от хлебных крошек?«Хлебные крошки» на веб-сайте помогают стимулировать просмотр и снижать показатель отказов, улучшая доступность вашего веб-сайта, обеспечивая преимущества SEO и помогая снизить беспокойство пользователей.
Продолжить чтение
.