
|
|
|
|
 Far Far |
| WinNavigator |
| Frigate |
| Norton
Commander |
| WinNC |
| Dos
Navigator |
| Servant
Salamander |
| Turbo
Browser |
|
|
| Winamp,
Skins, Plugins |
| Необходимые
Утилиты |
| Текстовые
редакторы |
| Юмор |
|
|

|
File managers and best utilites |
Реферат: Клеточное ядро. Реферат ядро
Доклад - Клеточное ядро - Биология и химия
Ядро — важнейшая составная часть клетки. Клеточное ядро содержит ДНК, т.е. гены, и, благодаря этому, выполняет две главные функции:
1)хранения и воспроизведения генетической информации
2)регуляции процессов обмена веществ, протекающих в клетке
Безъядерная клетка не может долго существовать, и ядро тоже не способно к самостоятельному_существованию, поэтому цитоплазма и ядро образуют взаимозависимую систему. Большинство клеток имеет одно ядро. Нередко можно наблюдать 2-3 ядра в одной например в клетках печени. Известны и многоядерные клетки, причем число ядер может достигать нескольких десятков. Форма ядра зависит большей частью от формы клетки, она может быть и совершенно неправильной. Различают ядра шаровидные, многолопастные. Впячивания и выросты ядерной оболочки значительно увеличивают поверхность ядра и тем самым усиливают связь ядерных и цитоплазматических структур и веществ.
Строение ядра
Ядро окружено оболочкой, которая состоит из двух мембран, имеющих типичное строение. Наружная ядерная мембрана с поверхности, обращенной в цитоплазму, покрыта рибосомами, внутренняя мембрана гладкая.
Ядерная оболочка-часть мембранной системы клетки.Выросты внешней ядерной мембраны соединяются с каналами эндоплазматической сети, образуя единую систему сообщающихся каналов.Обмен веществ между ядром и цитоплазмой осуществляется двумя основными путями.Во-первых, ядерная оболочка пронизана многочисленными порами, через которые происходит обмен молекулами между ядром и цитоплазмой.Во-вторых, вещества из ядра в цитоплазму и обратно могут попадать вследствии отшнуровывания впячиваний и выростов ядерной оболочки.Несмотря на активный обмен веществами между ядром и цитоплазмой, ядерная оболочка ограничивает ядерное содержимое от цитоплазмы, обеспечивая тем самым различия в химическом составе ядерного сока и цитоплазмы.Это необходимо для нормального функционирования ядерных структур.
Содержимое ядра подразделяют на ядерный сок, хроматин и ядрышко.
В живой клетке ядерный сок выглядит бесструктурной массой, заполняющей промежутки между структурами ядра.В состав ядерного сока входят различные белки, в том числе большенство ферментов ядра, белки хроматина и рибосомальные белки.В ядерном соке находятся также свободные нуклеотиды, необходимые для построения молекул ДНК и РНК, аминокислоты, все виды РНК, а также продукты деятельности ядрышка и хроматина, транспортируемые затем из ядра в цитоплазму.
Хроматином (то греч.chroma-окраска, цвет)называют глыбки, гранулы и сетевидные структуры ядра, интенсивно окрашивающиеся некоторыми красителями и отличаются по форме от ядрышка. Хроматин содержит ДНК и белки и представляет собой спирализованные и уплотненные участки хромосом Спирализованные участки хромосом в генетическом отношении неактивны.
Свою специфическую роль-передачу генетической информации-могут осуществлять только деспирализованные-раскрученные участки хромосом, которые в силу своей малой толщины не видны в световой микроскоп.
В делящихся клетках все хромосомысильно спирализуются, укорачиваются и приобретают компактные размеры и форму.Хромосомой называют самостоятельные ядерные структуры, имеющие плечи и первичную перетяжку.Форма хромосом зависит от положения так называемой первичной перетяжки, или центормеры,-области, к которой во время деления клетки(митоза)прикрепляются нити веретена деления. Центромера делит хромосому на два плеча. Расположение центромеры определяет три основных типа хромосом:
1)равноплечие-с плечами равной или почти равной длинны;
2)неравноплечие-с плечами неравной длинны;
3)палочковидные — с одним длинным и вторым очень коротким, иногда с трудом обнаруживаемым плечом. Выделяются еще точечные хромосомы с очень короткими плечами.
Изучение хромосом позволило установить следующие факты.
1.Во всех соматических клетках любого растительного или животного организма число хромосом одинаково.
2.Половые клетки всегда содержат двое меньше хромосом, чем соматические клетки данного вида организма.
3.У всех организмов, относящихся к одному виду, число хромосом в клетках одинаково.
Число хромосом не зависит от уровня организации и не всегда указывает на родство: одно и тоже число их может быть у очень далёких друг от друга систематических групп и может сильно отличаться у близких по происхождению видов.
Таким образом, само по себе число хромосом не является видоспецифическим признаком.Однако характеристика хромосомного набора в целом видоспецифична, т.е. свойственна только одному какому-то виду организмов растений растений или животных.
Совокупность количественных (число и размеры) и качественных (форма) признаковхромосомного набора соматической клетки называюткариотипом.
Число хромосом в кариотипе большинства видов живых организмов четное.Это объясняетя тем, что в соматических клетках находятся две одинаковые по форме и размеру хромосомы-одна из отцовского организма, вторая – из материнского. Хромосомы, одинаковые по форме и размеру и несущие одинаковые гены, называют гомологичными.
Хромосомный набор соматической клетки, в котором каждая хромосома имеет пару, носит название двойного или диплоидного и обозначается 2N. Количество ДНК, соответствующее диплоидному набору хромосом, обозначают 2C.
Из каждой пары гомологичных хромосом в половые клетки попадает только одна, и поэтому хромосомный набор гамет называют одинарным или гаплоидным. Кариотип таких клеток обозначается 2n1c.
Диплоидное число хромосом у животных и растений.
| Вид организмов | Число хромосом |
| Малярийный плазмодий | 2 |
| Сазан | 104 |
| Лошадиная аскарида | 2 |
| Человек | 46 |
| Плодовая мушка дрозофила | 8 |
| Ясень обыкновенный | 46 |
| Головная вошь | 12 |
| Шимпанзе | 48 |
| Шпинат | 12 |
| Таракан | 48 |
| Домашняя муха | 12 |
| Перец | 48 |
| Тритон | 24 |
| Домашняя овца | 54> |
| Ель, сосна | 24 |
| Домашняя собака | 78 |
| Окунь | 28 |
| Голубь | 80 |
После завершения деления клетки хромосомы диспирализуются, и в ядрах образовавшихся дочерних клеток снова становятся видимыми только тонкая сеточка и глыбки хроматина.
Третья характерная для клетки структура – ядрышко.Оно представляет собой плотное округлое тельце, погруженное в ядерный сок. В ядрах разных клеток, а также в ядре одной и той же клетки в зависимости от её функционального состояния число ядрышек может колебаться от 1 до 5-7 и более. Количество ядрышек может превышать число хромосомом в наборе; это происходит за счет избирательной редупликации генов, отвечающих за синтез р-РНК. Ядрышки есть только в неделящихся ядрах, во время митоза они исчезают вследствие спирализации хромосом и выхода всех ранее образованных рибосом в цитоплазму, а после завершения деления возникают вновь.
Ядрышко не является самостоятельной структурой ядра.Оно образуется вокруг участка хромосомы, в котором закодирована структура р-РНК. Этот участок хромосомы-ген-носит название ядрышкового организатора(ЯО), и на нем происходит синтез р-РНК.
Кроме накопления р-РНК, в ядрышке формируются субъединицы рибосом, которые потом перемещаются в цитоплазму и, объединяясь при участии катионов Ca2+, формируют целостностные рибосомы, способные принимать участие в биосинтезе белка.
Таким образом, ядрышко – это скопление р-РНК и рибосом на разных этапах формирования, в основе которого лежит участок хромосомы, несущий ген – ядрышковый организатор, заключающий наследственную информацию о структуре р –РНК.
www.ronl.ru
Реферат - Ядро - Рефераты на репетирем.ру
“Структура и функции ядра”
Говоря о клеточном ядре, мы имеем в виду собственно ядра эукариотических клеток. Их ядра построены сложным образом и довольно резко отличаются от “ядерных” образований, нуклеоидов, прокариотических организмов. У последних в состав нуклеоидов (ядроподобных структур) входит одиночная кольцевая молекула ДНК, практически лишенная белков. Иногда такую молекулу ДНК бактериальных клеток называют бактериальной хромосомой, или генофором (носителем генов). Бактериальная хромосома не отделена мембранами от основной цитоплазмы, однако собрана в компактную ядерную зону - нуклеоид, который можно видеть в световом микроскопе после специальных окрасок.
Сам термин “ядро” впервые был применен Броуном в 1833 г. Для обозначения шаровидных постоянных структур в клетках растений. Позднее такую же структуру описали во всех клетках высших организмов.
Клеточное ядро обычно одно на клетку (есть примеры многоядерных клеток), состоит из ядерной оболочки, отделяющей его от цитоплазмы, хроматина, ядрышка, кариоплазмы (или ядерного сока) (рис). Эти четыре основных компонента встречаются практически во всех неделящихся клетках эукариотических одно- и многоклеточных организмов.
Ядра имеют обычно шаровидную или яйцевидную форму; диаметр первых равен приблизительно 10 мкм, а длина вторых - 20 мкм.
Ядро необходимо для жизни клетки, поскольку именно оно регулирует всю ее активность. Связано это с тем, что ядро несет в себе генетическую (наследственную) информацию, заключенную в ДНК.
Ядерная оболочка
Эта структура характерна для всех эукариотических клеток. Ядерная оболочка состоит из внешней и внутренней мембран, разделенных перинуклеарным пространством шириной от 20 до 60 нм. В состав ядерной оболочки входят ядерные поры.
Мембраны ядерной оболочки в морфологическом отношении не отличаются от остальных внутриклеточных мембран: они имеют толщину около 7 нм и состоят из двух осмиофильных слоев.
В общем виде ядерная оболочка может быть представлена, как полый двухслойный мешок, отделяющий содержимое ядра от цитоплазмы. Из всех внутриклеточных мембранных компонентов таким типом расположения мембран обладают только ядро, митохондрии и пластиды. Однако ядерная оболочка имеет характерную особенность, отличающую ее от других мембранных структур клетки. Это наличие особых пор в оболочке ядра, которые образуются за счет многочисленных зон слияний двух ядерных мембран и представляет собой как бы округлые перфорации всей ядерной оболочки.
Строение ядерной оболочки
Внешняя мембрана ядерной оболочки, непосредственно контактирующая с цитоплазмой клетки, имеет ряд структурных особенностей, позволяющих отнести ее к собственно мембранной системе эндоплазматического ретикулума. Так, на внешней ядерной мембране обычно располагается большое количество рибосом. У большинства животных и растительных клеток внешняя мембрана ядерной оболочки не представляет собой идеально ровную поверхность - она может образовывать различной величины выпячивания или выросты в сторону цитоплазмы.
Внутренняя мембрана контактирует с хромосомным материалом ядра (см. Ниже).
Наиболее характерной и бросающейся в глаза структурой в ядерной оболочке является ядерная пора. Поры в оболочке образуются за счет слияния двух ядерных мембран в виде округлых сквозных отверстий или перфораций с диаметром 80-90 нм. Округлое сквозное отверстие в ядерной оболочке заполнено сложноорганизованными глобулярными и фибриллярными структурами. Совокупность мембранных перфораций и этих структур называют комплексом пор ядра. Тем самым подчеркивается, что ядерная пора не просто сквозная дыра в ядерной оболочке, через которую непосредственно вещества ядра и цитоплазмы могут сообщаться.
Сложный комплекс пор имеет октагональную симметрию. По границе округлого отверстия в ядерной оболочке располагаются три ряда гранул, по 8 штук в каждом: один ряд лежит со стороны ядра, другой - со стороны цитоплазмы, третий расположен в центральной части пор. Размер гранул около 25 нм. От этих гранул отходят фибриллярные отростки. Такие фибриллы, отходящие от периферических гранул, могут сходиться в центре и создавать как бы перегородку, диафрагму, поперек поры. В центре отверстия часто можно видеть так называемую центральную гранулу.
Число ядерных пор зависит от метаболической активности клеток: чем выше синтетические процессы в клетках, тем больше пор на единицу поверхности клеточного ядра.
Количество ядерных пор в различных объектах
| Объект | Число ядерных пор на мкм2 | Число пор на одно ядро |
| Ксенопус, почки | 10,05 | 3417 |
| Ксенопус, ооцит | 51,0 | 37,6*106 |
| Крыса, гепатоцит | 16,1 | 3816 |
| Мышь, лимфоцит | 3,3 | 403 |
| Человек, лимфоцит | 4,47 | 713 |
Химия ядерной оболочки
В составе ядерных оболочек обнаруживаются небольшие количества ДНК (0-8%), РНК (3-9%), но основными химическими компонентами являются липиды (13-35%) и белки (50-75%), что для всех клеточных мембран.
Состав липидов сходен с таковым в мембранах микросом или мембранах эндоплазматической сети. Ядерные оболочки характеризуются относительно низким содержанием холестерина и высоким - фосфолипидов, обогащенных насыщенными жирными кислотами.
Белковый состав мембранных фракций очень сложен. Среди белков обнаружен ряд ферментов, общих с ЭР (например, глюкозо-6-фосфатаза, Mg-зависимая АТФаза, глютамат-дегидрогеназа и др.) не обнаружена РНК-полимераза. Тут выявлены активности многих окислительных ферментов (цитохромоксидазы, НАДН-цитохром-с-редуктазы) и различных цитохромов.
Среди белковых фракций ядерных мембран встречаются основные белки типа гистонов, что объясняется связью участков хроматина с ядерной оболочкой.
Ядерная оболочка и ядерно-цитоплазматический обмен
Ядерная оболочка - система, разграничивающая два основных клеточных отсека: цитоплазму и ядро. Ядерные оболочки полностью проницаемы для ионов, для веществ малого молекулярного веса, таких, как сахара, аминокислоты, нуклеотиды. Считается, что белки молекулярного веса до 70 тыс. И размером не больше 4,5 нм могут свободно диффундировать через оболочку.
Известен и обратный процесс - перенос веществ из ядра в цитоплазму. Это в первую очередь касается транспорта РНК синтезируещегося исключительно в ядре.
Еще один путь транспорта веществ из ядра в цитоплазму связан с образованием выростов ядерной оболочки, которые могут отделяться от ядра в виде вакуолей, содержимое их затем изливается или выбрасывается в цитоплазму.
Таким образом, из многочисленных свойств и функциональных нагрузок ядерной оболочки следует подчеркнуть ее роль как барьера, отделяющего содержимое ядра от цитоплазмы, ограничивающего свободный доступ в ядро крупных агрегатов биополимеров, барьера, активно регулирующего транспорт макромолекул между ядром и цитоплазмой.
Одной из основных функций ядерной оболочки следует считать также ее участие в создании внутриядерного порядка, в фиксации хромосомного материала в трехмерном пространстве ядра.
Ядерный матрикс
Этот комплекс не представляет собой какую-то чистую фракцию, сюда входят компоненты и ядерной оболочки, и ядрышка, и кариоплазмы. С ядерным матриксом оказались связаны как гетерогенная РНК, так и часть ДНК. Эти наблюдения дали основание считать, что матрикс ядра играет важную роль не только в поддержании общей структуры интерфазного ядра, но и может участвовать в регуляции синтеза нуклеиновых кислот.
Хроматин
При наблюдении некоторых живых клеток, особенно растительных или же клеток после фиксации и окраски, внутри ядра выявляются зоны плотного вещества. В состав хроматина входит ДНК в комплексе с белком. В интерфазных клетках хроматин может равномерно заполнять объем ядра или же располагаться отдельными сгустками (хромоцентры). Часто он особенно четко выявляется на периферии ядра (пристеночный, примембранный хроматин) или образует внутри ядра переплетения довольно толстых (около 0.3 мкм) и длинных тяжей, образующих подобие внутриядерной цепи.
Хроматин интерфазных ядер представляет собой несущие ДНК тельца (хромосомы), которые теряют в это время свою компактную форму, разрыхляются, деконденсируются. Степень такой деконденсации хромосом может быть различной в ядрах разных клеток. Когда хромосома или ее участок полностью деконденсирован, тогда эти зоны называют диффузным хроматином. При неполном разрыхлении хромосом в интерфазном ядре видны участки конденсированного хроматина (иногда называемого гетерохроматин). Показано, что степень деконденсации хромосомного материала в интерфазе может отражать функциональную нагрузку этой структуры. Чем более диффузен хроматин интерфазного ядра, тем выше в нем синтетические процессы. Падение синтеза РНК в клетках обычно сопровождается увеличением зон конденсированного хроматина.
Максимально конденсирован хроматин во время митотического деления клеток, когда он обнаруживается в виде плотных телец - хромосом. В этот период хромосомы не несут никаких синтетических нагрузок, в них не происходит включение предшественников ДНК и РНК.
Исходя из этого можно считать, что хромосомы клеток могут находиться в двух структурно-функциональных состояниях:
в рабочем, частично или полностью деконденсированном, когда с их участием в интерфазном ядре происходят процессы транскрипции и редупликации;
в неактивном - в состоянии метаболического покоя при максимальной их конденсированности, когда они выполняют функцию распределения и перенося генетического материала в дочерние клетки.
В химическом отношении препараты хроматина представляют собой сложные комплексы дезоксирибонуклеопротеидов, в состав которых входит ДНК и специальные хромосомные белки - гистоны. В составе хроматина обнаружено также РНК. В количественном отношении ДНК, белок и РНК находятся как 1:1,3:0,2. О значении РНК в составе хроматина еще нет достаточно однозначных данных. Возможно, что эта РНК представляет собой сопутствующую препарату функцию синтезирующейся РНК и поэтому частично связанной с ДНК или это особый вид РНК, характерный для структуры хроматина.
ДНК хроматина
В препарате хроматина на долю ДНК приходится обычно 30-40%. Эта ДНК представляет собой двухцепочечную спиральную молекулу. ДНК хроматина обладает молекулярной массой 7-9*106. Такую сравнительно малую массу ДНК из препаратов можно объяснить механическими повреждениями ДНК в процессе выделения хроматина.
Общее количество ДНК, входящее в ядерные структуры клеток, в геном организмов, колеблется от вида к виду. Сравнивая количество ДНК на клетку у эукариотических организмов, трудно уловить какие-либо корреляции между степенью сложности организма и количеством ДНК на ядро. Примерно одинаковое количество ДНК имеют различные организмы, как лен, морской еж, окунь (1,4-1,9 пг) или рыба голец и бык (6,4 и 7 пг).
У некоторых амфибий в ядрах количество ДНК больше, чем в ядрах человека, в 10-30 раз, хотя генетическая конституция человека несравненно сложнее, чем у лягушек. Следовательно, можно предполагать, что “избыточное” количество ДНК у более низко организованных организмов либо не связано с выполнением генетической роли, либо число генов повторяется то или иное число раз.
Сателлитная ДНК, или фракция ДНК с часто повторяющимися последовательностями, может участвовать в узнавании гомологичных районов хромосом при мейозе. По другим предположениям, эти участки играют роль разделителей (спейсеров) между различными функциональными единицами хромосомной ДНК.
Как оказалось, фракция умеренно повторяющихся (от 102 до 105 раз) последовательностей принадлежит к пестрому классу участков ДНК, играющих важную роль в обменных процессах. В эту фракцию входят гены рибосомных ДНК, многократно повторенные участки для синтеза всех тРНК. Более того, некоторые структурные гены, ответственные за синтез определенных белков, также могут быть многократно повторены, представлены многими копиями (гены для белков хроматина - гистонов).
Итак, ДНК эукариотических клеток гетерогенна по составу, содержит несколько классов последовательностей нуклеотидов:
часто повторяющиеся последовательности (>106 раз), входящие во фракцию сателитной ДНК и не транскрибирующиеся;
фракция умеренно повторяющихся последовательностей (102-105), представляющих блоки истинных генов, а также короткие последовательности, разбросанные по всему геному;
фракция уникальных последовательностей, несущая информацию для большинства белков клетки.
ДНК прокариотического организма представляет собой одну гигантскую циклическую молекулу. ДНК эукариотических хромосом представляет собой линейные молекулы, состоящие из тандемно (друг за другом) расположенных репликонов разного размера. Средний размер репликона около 30 мкм. Тем самым в составе генома человека должно встречаться более 50 000 репликонов, участков ДНК, которые синтезируются как независимые единицы. Эти репликоны имеют начальную и терминальную точки синтеза ДНК.
Представим себе, что у эукариотических клеток каждая из хромосомных ДНК, как и у бактерий, является одним репликоном. В этом случае при скорости синтеза 0,5 мкм в минуту (для человека) редупликация первой хромосомы с длиной ДНК около 7 см должна занять 140 000 минут, или около трех месяцев. На самом же деле благодаря полирепликонному строению молекул ДНК весь процесс занимает 7-12 ч.
Белки хроматина
К ним относятся гистоны и негистоновые белки.
Гистоны - сильноосновные белки. Их щелочность связана с их обогащенностью основными аминокислотами (главным образом лизином и аргинином). Эти белки не содержат триптофана. Препарат суммарных гистонов можно разделить на 5 фракций:
Н1 (от английского histone) - богатый лизином гистон, мол. Масса 2100;
Н2а - умеренно богатый лизином гистон, масса 13 700;
Н2б - умеренно богатый лизином гистон, масса 14 500;
Н4 - богатый аргинином гистон, масса 11 300;
Н3 - богатый аргинином гистон, масса 15 300.
В препаратах хроматина эти фракции гистонов обнаруживаются в приблизительно равных количествах, кроме Н1, которого примерно в 2 раза меньше любой из других фракций.
Для молекул гистонов характерно неравномерное распределение основных аминокислот в цепи: обогащенные положительно заряженными аминогруппами наблюдается на концах белковых цепей. Эти участки гистонов связываются с фосфатными группировками на ДНК, в то время как сравнительно менее заряженные центральные участки молекул обеспечивают их взаимодействие между собой. Таким образом, взаимодействие между гистонами и ДНК, приводящее к образованию дезоксирибонуклеопротеинового комплекса, носит ионный характер.
Гистоны синтезируются на полисомах в цитоплазме, этот синтез начинается несколько раньше редупликации ДНК. Синтезированные гистоны мигрируют из цитоплазмы в ядро, где и связываются с участками ДНК.
Функциональная роль гистонов не вполне ясна. Одно время считалось, что гистоны являются специфическими регуляторами активности ДНК хроматина, но одинаковость строения основной массы гистонов говорит о малой вероятности этого. Более очевидна структурная роль гистонов, которая обеспечивает не только специфическую укладку хромосомной ДНК, но и играет роль в регуляции транскрипции.
Негистоновые белки - наиболее плохо охарактеризованная фракция хроматина. Кроме ферментов, непосредственно связанных с хроматином (ферменты, ответственные за репарацию, редубликацию, транскрипцию и модификации ДНК, ферменты модификации гистонов и других белков), в эту фракцию входит множество других белков. Весьма вероятно, что часть негистоновых белков представляет собой специфические белки - регуляторы, узнающие определенные нуклеотидные последовательности в ДНК.
РНК хроматина составляет от 0,2 до 0,5% от содержания ДНК. Эта РНК представляет собой все известные клеточные типы РНК, находящиеся в процессе синтеза или созревания в связи с ДНК хроматина.
В составе хроматина могут быть обнаружены липиды до 1 % от весового содержания ДНК, их роль в структуре и функционировании хромосом остается неясной.
Хромосомы
Первичная степень укладки молекул ДНК - хромосомная фибрилла. Наблюдения за структурой хроматина с помощью электронного микроскопа показали, что в составе ядра на ультратонких срезах всегда видны фибриллярные элементы. Впервые их обнаружил Х. Рис (1957), который и дал им название элементарных хромосомных фибрилл.
Морфология хромосом
Морфологию хромосом лучше всего изучать в момент их наибольшей конденсации, в метафазе и в начале анафазы. Хромосомы животных и растений в этом состоянии представляют собой палочковидные структуры разной длины с довольно постоянной толщиной, у большей части хромосом удается легко найти зону первичной перетяжки, которая делит хромосому на два плеча. Хромосомы с равными или почти равными плечами называют метацентрическими, с плечами неодинаковой длины - субметацентрическими. Палочковидные хромосомы с очень коротким, почти незаметным вторым плечом - акроцентрические.
В области первичной перетяжки расположена центромера, или кинетохор. Это пластинчатая структура, имеющая форму диска. Она связана тонкими фибриллами с телом хромосомы в области перетяжки. От него отрастают пучки микротрубочки митотического веретена, идущие в направлении к центриолям. Они принимают участие в движении хромосом к полюсам клетки при митозе.
Обычно одна хромосома имеет только одну центромеру (моноцентрические хромосомы), но могут встречаться хромосомы дицентрические и полицентрические.
Некоторые хромосомы имеют вторичную перетяжку. Последняя обычно расположена вблизи дистального конца хромосомы и отделяет маленький участок, спутник. Вторичные перетяжки называют, кроме того, ядрышковыми организаторами, так как именно на этих участках хромосом в интерфазе происходит образование ядрышка. Здесь же локализована ДНК, ответственная за синтез рРНК.
Плечи хромосом оканчиваются теломерами, конечными участками. Теломерные концы хромосом не способны соединяться с другими хромосомами или их фрагментами, в отличие от концов хромосом, лишенных теломерных участков, которые могут присоединяться к таким же разорванным концам других хромосом.
Размеры хромосом у разных организмов варьируют в широких пределах. Так, длина хромосом может колебаться от 0,2 до 50 мкм. Самые мелкие хромосомы обнаруживаются у некоторых простейших, грибов. Наиболее длинные - у некоторых прямокрылых насекомых, у амфибий и у лилейных. Длина хромосом человека находится в пределах 1,5-10 мкм.
Число хромосом у различных объектов тоже значительно колеблется, но характерно для каждого вида. У некоторых радиолярий число хромосом достигает 1000-1600. Рекордсменом среди растений по числу хромосом (около 500) является папоротник ужовник, 308 хромосом у тутового дерева, у речного рака 196 хромосом. Наименьшее количество хромосом (2 на диплоидный набор) наблюдается у одной из рас аскариды, у сложноцветного Haplopappus gracilic - всего 4 хромосомы (2 пары).
Совокупность числа, величины, величины и морфологии хромосом называется кариотипом данного вида. Даже у близких видов хромосомные наборы отличаются друг от друга или по числу хромосом, или по величине хотя бы одной или нескольких хромосом. Следовательно, структура кариотипа может быть таксономическим признаком.
Ядрышко
Практически во всех живых клетках эукариотических организмов в ядре видно одно или несколько обычно округлой формы тельц, сильно преломляющих свет, - это ядрышки, или нуклеолы.
Ядрышко - не самостоятельная структура или органоид. Оно - производное хромосомы, один из ее локусов, активно функционирующий в интерфазе.
В процессах синтеза клеточных белков ядрышко клетки является местом образования рибосомных РНК и рибосом, на которых происходит синтез полипептидных цепей.
Количество ядрышек в клетке
Начиная с зеленых водорослей, грибов и низших простейших и кончая высшими организмами, все клетки имеют обязательные внутриядерные структуры - ядрышки. Это правило имеет большое число исключений, которые только подчеркивают важность и необходимость ядрышка в жизненном цикле клетки. К таким исключениям относятся клетки дробящихся яиц, где ядрышки отсутствуют на ранних этапах эмбриогенеза, или клетки закончившие развитие и необратимо специализировавшиеся, например, некоторые клетки крови.
Количество ядрышек в клетке может меняться, однако их число на ядро зависит от генного баланса клетки. Было найдено, что в образовании ядрышек участвуют определенные места некоторых хромосом, связь которых с ядрышком можно хорошо проследить в телофазе и профазе. Такие хромосомы, как правило, имеют вторичные перетяжки, зоны которых представляют собой места, где идет развитие ядрышек. Мак Клинток (1934) назвал эти участки хромосом “ядрышковыми организаторами”.
Места вторичных перетяжек особенно характерны для расположения ядрышковых организаторов, но последние иногда могут находиться на концах хромосом или в нескольких местах по длине хромосомы.
Общее число ядрышек на ядро определяется числом ядрышковых организаторов и увеличивается согласно плоидности ядра. Однако часто количество ядрышек на ядро бывает меньше числа ядрышковых организаторов. Было показано, что ядрышки могут сливаться; кроме того, в образовании одного ядрышка иногда участвует несколько организаторов.
Еще в работах М.С.Навашина (1934) было показано, что хромосомный локус, который в нормальных условиях образует крупное ядрышко, становится неактивным, когда после гибридизации в ядре появляется более “сильный” локус на другой хромосоме. Тот факт, что в определенных условиях может подавляться активность одних ядрышковых организаторов или же повышаться активность других, бывших до этого в латентном, скрытом состоянии, указывает на то, что в клетках поддерживается определенный баланс количества ядрышкового материала или, другими словами, регулируется “валовая” продукция, выдаваемая ядрышками.
Исходя из перечисленных выше фактов, можно сделать следующие заключения:
образования ядрышек и их число связаны с активностью определенных участков хромосом - ядрышковых организаторов, которые расположены большей частью в зонах вторичных перетяжек;
изменения в числе ядрышек в клетках данного типа могут происходить за счет слияния ядрышек или за счет сдвигов в хромосомном балансе клетки.
Физиология и химия ядрышка
Ядрышко по сравнению с другими компонентами клетки характеризуется как самая плотная структура с наиболее высокой концентрацией РНК, с чрезвычайно высокой активностью в отношении синтеза РНК.
Концентрация РНК в ядрышках всегда выше концентрации РНК в других компонентах клетки, так концентрация РНК в ядрышке может быть в 2-8 раз выше, чем в ядре, и в 1-3 раза выше, чем в цитоплазме. Отношение концентрации РНК в ядре, ядрышке и цитоплазме клеток печени мыши составляет 1:7,3:4,1, в клетках поджелудочной железы - 1:9,6:6,6.
В ядрышке не обнаруживается ДНК, но все же при исследовании фиксированных клеток вокруг ядрышка всегда выделяется зона хроматина. Этот околоядрышковый хроматин, по данным электронной микроскопии, представляется, как интегральная часть сложной структуры ядрышка.
Ядрышко - одно из самых активных мест в клетке по включению предшественников в РНК. Ядрышковая РНК является предшественником цитоплазматической РНК.
Цитоплазматическая РНК синтезируется в ядрышке.
РНК ядрышек
Оценивая общее содержание в ядрышковых фракциях белков, РНК и ДНК, можно видеть, что на долю РНК приходится около 10% всей массы ядрышка.
Содержание РНК, ДНК и белка в изолированных ядрышках (сухой вес в %)
| Объект | РНК | ДНК | Белок | РНК/ДНК |
| Печень крысы | 11,0 | 8,0 | 78,0 | 1,4 |
| Регенерирующая печень (6 ч) | 7,6 | 4,6 | 87,8 | 1,7 |
| Регенерирующая печень (18 ч) | 15,5 | 5,4 | 79,1 | 2,9 |
| Печень морской свинки | 4,1 | 9,5 | 86,4 | 0,43 |
| Стебель гороха (4 дня) | 15,11 | 10,6 | 74,0 | 1,5 |
| Проростки гороха (36 ч) | 16,7 | 6,4 | 76,9 | 2,6 |
Так как основную массу цитоплазматической РНК составляет рибосомная РНК, то можно сказать, что ядрышковая РНК принадлежит к этому классу.
Подтверждением представлений того, что именно ядрышко является местом синтеза рРНК и образования рибосом, послужило то, что из ядрышковых препаратов были выделены РНП-частицы, которые как по составу РНК (по седиментационным свойствам), так и по размеру можно охарактиризовать как рибосомы или их предшественники с различными коэффициентами седиментации.
ДНК ядрышек
Биохимическими исследованиями обнаружено в выделенных ядрышках определенное количество ДНК, которую можно отождествить с околоядрышковым хроматином или с ядрышковыми организаторами хромосом. Содержание ДНК в выделенных ядрышках - 5-12% от сухого веса и 6-17% от всей ДНК ядра.
ДНК ядрышкового организатора - это та самая ДНК, на которой происходит синтез ядрышковой, т.е. рибосомной, РНК.
Таким образом из биохимических работ появились представления о том, что в ядрышке на ДНК локализованы многочисленные одинаковые гены для синтеза рРНК. Синтез рРНК идет путем образования огромного предшественника и дальнейшего его превращения (созревания) в более короткие молекулы РНК для большой и малой субъедениц рибосом.
Изучая ядрышки ооцитов тритонов, исследователи столкнулись с интересным явлением - сверхчисленностью ядрышек. У X. laevis во время роста ооцита появляется до 1000 мелких ядрышек, не связанных с хромосомами. Именно эти ядрышки выделил О.Миллер. вместе с этим на ядро ооцита увеличивается количество рДНК. Это явление получило название амплификации. Оно заключается в том, что происходит сверхрепликация зоны ядрышкового организатора, многочисленные копии отходят от хромосом и становятся дополнительно работающими ядрышками. Такой процесс необходим для накопления огромного (1012) количества рибосом на яйцевую клетку, что обеспечит в будущем развитие эмбриона на ранних стадиях даже при отсутствии синтеза новых рибосом. Сверхчисленные ядрышки после созревания яйцевой клетки исчезают.
Ультраструктура ядрышек
При изучении большого числа различных клеток животных и растений отмечена волокнистая или сетчатая структура ядрышек, заключенная в более или менее плотную диффузную массу. Были предложены названия для этих частей: волокнистая часть - нуклеонема и диффузная, гомогенная часть - аморфное вещество, или аморфная часть. Сделанные почти одновременно с этим электронно-микроскопичес-кие исследования также выявили волокнисто-нитчатое строение ядрышек.
Однако такое нитчатое строение ядрышка не всегда четко выражено. У некоторых клеток отдельные нити нуклеонем сливаются, и ядрышки могут быть совершенно однородными.
При более пристальном изучении ядрышка можно заметить, что основные структурные компоненты ядрышка - плотные гранулы диаметром около 15 нм и тонкие фибриллы толщиной 4-8 нм. Во многих случаях (ооциты рыб и амфибий, меристематические клетки растений) фибриллярный компонент собран в плотную центральную зону (сердцевина), лишенную гранул, а гранулы занимают переферическую зону ядрышка. В ряде случаев (например, клетки корешков растений) в этой гранулярной зоне не наблюдается никакой дополнительной структуризации.
Было найдено, что аморфные участки ядрышек неоднородны. В их структуре выявляются малоокрашенные зоны - фибриллярные центры - и окружающие их более темные участки, тоже имеющие фибриллярное строение.
Кроме этих двух компонентов ядрышек в последнее время большое внимание уделялось строению околоядрышкового хроматина. Этот хроматин и внутриядрышковая сеть ДНК являются единой системой и представляют собой интегральный компонент ядрышка.
Гранулы и фибриллярная часть состоят из рибонуклеопротеидов.
Показано, что именно светлые фибриллярные центры содержат рДНК.
Судьба ядрышка при делении клеток
Известно, что ядрышко исчезает в профазе и появляется вновь в средней телофазе.
По мере затухания синтеза рРНК в средней профазе происходит разрыхление ядрышка и выход готовых рибосом в кариоплазму, а затем и в цитоплазму. При конденсации профазных хромосом фибриллярный компонент ядрышка и часть гранул тесно ассоциируют с их поверхностью, образуя основу матрикса митотических хромосом. Этот фибриллярно-гранулярный материал, синтезированный до митоза, переносится хромосомами в дочерние клетки.
В ранней телофазе по мере деконденсации хромосом происходит высвобождение компонентов матрикса. Его фибриллярная часть начинает собираться в мелкие многочисленные ассоциаты - предъядрышки, которые могут объединяться друг с другом. По мере возобновления синтеза РНК предъядрышки претерпевают перестройку, что выражается в появлении в их структуре гранул РНК, а затем в становлении дефинитивной формы нормально функционирующего ядрышка.
Роль ядра.
Ядро осуществляет две группы общих функций: одну, связанную собственно с хранением генетической информации, другую - с ее реализацией, с обеспечением синтеза белка.
В первую группу входят процессы, связанные с поддержанием наследственной информации в виде неизменной структуры ДНК. Эти процессы связаны с наличием так называемых репарационных ферментов, ликвидирующих спонтанные повреждения молекулы ДНК (разрыв одной из цепей ДНК, часть радиационных повреждений), что сохраняет строение молекул ДНК практически неизменным в ряду поколений клеток или организмов. Далее, в ядре происходит воспроизведение или редупликация молекул ДНК, что дает возможность двум клеткам получить совершенно одинаковые и в качественном и в количественном смысле объемы генетической информации. В ядрах происходят процессы изменения и рекомбинации генетического материала, что наблюдается во время мейоза (кроссинговер). Наконец, ядра непосредственно участвуют в процессах распределения молекул ДНК при делении клеток.
Другой группой клеточных процессов, обеспечивающихся активностью ядра, является создание собственно аппарата белкового синтеза. Это не только синтез, транскрипция на молекулах ДНК разных информационных РНК и рибосомных РНК. В ядре эукариотов происходит также образование субъедениц рибосом путем комплексирования синтезированных в ядрышке рибосомных РНК с рибосомными белками, которые синтезируются в цитоплазме и переносятся в ядро.
Таким образом, ядро представляет собой не только вместилище генетического материала, но и место, где этот материал функционирует и воспроизводится. Поэтому выпадение лил нарушение любой из перечисленных выше функций губительно для клетки в целом. Так нарушение репарационных процессов будет приводить к изменению первичной структуры ДНК и автоматически к изменению структуры белков, что непременно скажется на их специфической активности, которая может просто исчезнуть или измениться так, что не будет обеспечивать клеточные функции, в результате чего клетка погибает. Нарушения редупликации ДНК приведут к остановке размножения клеток или к появлению клеток с неполноценным набором генетической информации, что также губительно для клеток. К такому же результату приведет нарушение процессов распределения генетического материала (молекул ДНК) при делении клеток. Выпадение в результате поражения ядра или в случае нарушений каких-либо регуляторных процессов синтеза любой формы РНК автоматически приведет к остановке синтеза белка в клетке или к грубым его нарушениям.
Значение ядра как хранилища генетического материала и его главная роль в определении фенотипических признаков были установлены давно. Немецкий биолог Хаммерлинг одним из первых продемонстрировал важнейшую роль ядра. Он выбрал в качестве объекта своих экспериментов необычайно крупную одноклеточную (или неклеточную) морскую водоросль Acetabularia. Существует два близко родственных вида A. medierranea и A. crenulata, различающихся только по форме “шляпки”.
В ряде экспериментов, в том числе таких, в которых “шляпку” отделяли от нижней части “стебелька” (где находится ядро), Хаммерлинг показал, что для нормального развития шляпки необходимо ядро. В дальнейших экспериментах, в которых соединяли нижнюю часть, содержащую ядро одного вида с лишенным ядра стебельком другого вида, у таких химер всегда развивалась шляпка, типичная для того вида, которому принадлежит ядро.
При оценке этой модели ядерного контроля следует, однако, учитывать примитивность организма, использованного в качестве объекта. Метод пересадок был применен позднее в экспериментах, проведенных в 1952 г. двумя американскими исследователями, Бриггсом и Кингом, с клетками лягушки Rana pipenis. Эти авторы удаляли из неоплодотворенных яйцеклеток ядра и заменяли их ядрами из клеток поздней бластулы, уже проявлявших признаки дифференцировки. Во многих случаях из яиц реципиентов развивались нормальные взрослые лягушки.
Литература:
Свенсон К., Уэбстер П. “Клетка”. М., Мир, 1980.
Де Робертис Э. Новинский В., Саэс Ф. “Биология Клетки”. М., Мир, 1971
Ченцов Ю.С., Поляков В.Ю. “Ультраструктура клеточного ядра”. М., Наука, 1974
Зегнбуш П. “Молекулярная и клеточная биология”. М., Мир, т.1,2, 1982
ref.repetiruem.ru
Реферат Ядро клетки
скачатьРеферат на тему:
План:
- Введение
- 1 Тонкая структура клеточного ядра
- 1.1 Хроматин
- 1.2 Ядерная оболочка, ядерная ламина и ядерные поры (кариолемма)
- 1.3 Ядрышко
- 1.4 Ядерный матрикс
- 2 Эволюционное значение клеточного ядра
Введение
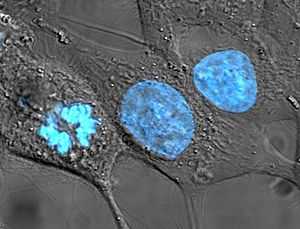
Клетки HeLa, ДНК которых окрашена голубым красителем Хёхста 33258. Центральная и правая клетки находятся в интерфазе, поэтому окрашено всё ядро. Клетка слева находится в состоянии митоза (анафаза), поэтому её ядро не видно, а ДНК сконденсирована так, что видны хромосомы
Ядро (лат. nucleus) — это один из структурных компонентов эукариотической клетки, содержащий генетическую информацию (молекулы ДНК). В ядре происходит репликация — удвоение молекул ДНК, а также транскрипция — синтез молекул РНК на молекуле ДНК. В ядре же синтезированные молекулы РНК подвергаются ряду модификаций, после чего выходят в цитоплазму. Образование субъединиц рибосом также происходит в ядре в специальных образованиях - ядрышках.
1. Тонкая структура клеточного ядра
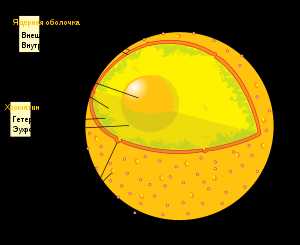
Схема строения клеточного ядра.
1.1. Хроматин
Огромная длина молекул ДНК эукариот предопределила появление специальных механизмов хранения, репликации и реализации генетического материала. Хроматином называют молекулы хромосомной ДНК в комплексе со специфическими белками, необходимыми для осуществления этих процессов. Основную массу составляют «белки хранения», так называемые гистоны. Из этих белков построены нуклеосомы, структуры на которые намотаны нити молекул ДНК. Нуклеосомы располагаются довольно регулярно, так что образующаяся структура напоминает бусы. Нуклеосома состоит из белков четырех типов: h3A, h3B, h4 и h5. В одну нуклеосому входят по два белка каждого типа — всего восемь белков. Гистон h2, более крупный чем другие гистоны, связывается с ДНК в месте ее входа на нуклеосому. Нуклеосома вместе с h2 называется хроматосомой.
Схема, показывающая цитоплазму, вместе с ее компонентами (или органеллами), в типичной животной клетке. Органеллы:(1) Ядрышко(2) Ядро(3) рибосома (маленькие точки)(4) Везикула(5) шероховатый эндоплазматический ретикулум (ER)(6) Аппарат Гольджи(7) Цитоскелет(8) Гладкий эндоплазматический ретикулум(9) Митохондрия(10) Вакуоль(11) Цитоплазма(12) Лизосома(13) Центриоль и Центросома
Нить ДНК с нуклеосомами образует нерегулярную соленоид-подобную структуру толщиной около 30 нанометров, так называемую 30 нм фибриллу. Дальнейшая упаковка этой фибриллы может иметь различную плотность. Если хроматин упакован плотно, его называют конденсированным или гетерохроматином, он хорошо видим под микроскопом. ДНК, находящаяся в гетерохроматине, не транскрибируется, обычно это состояние характерно для незначащих или молчащих участков. В интерфазе гетерохроматин обычно располагается по периферии ядра (пристеночный гетерохроматин). Полная конденсация хромосом происходит перед делением клетки. Если хроматин упакован неплотно, его называют эу- или интерхроматином. Этот вид хроматина гораздо менее плотный при наблюдении под микроскопом и обычно характеризуется наличием транскрипционной активности. Плотность упаковки хроматина во многом определяется модификациями гистонов — ацетилированием и фосфориллированием.
Считается, что в ядре существуют так называемые функциональные домены хроматина(ДНК одного домена содержит приблизительно 30 тысяч пар оснований), то есть каждый участок хромосомы имеет собственную «территорию». К сожалению, вопрос пространственного распределения хроматина в ядре изучен пока недостаточно. Известно, что теломерные (концевые) и центромерные (отвечающие за связывание сестринских хроматид в митозе) участки хромосом закреплены на белках ядерной ламины.
1.2. Ядерная оболочка, ядерная ламина и ядерные поры (кариолемма)
От цитоплазмы ядро отделено ядерной оболочкой, образованной за счёт расширения и слияния друг с другом цистерн эндоплазматической сети таким образом, что у ядра образовались двойные стенки за счёт окружающих его узких компартментов. Полость ядерной оболочки называется люменом или перинуклеарным пространством. Внутренняя поверхность ядерной оболочки подстилается ядерной ламиной, жёсткой белковой структурой, образованной белками-ламинами, к которой прикреплены нити хромосомной ДНК. Ламины прикрепляются к внутренней мембране ядерной оболочки при помощи заякоренных в ней трансмембранных белков — рецепторов ламинов. В некоторых местах внутренняя и внешняя мембраны ядерной оболочки сливаются и образуют так называемые ядерные поры, через которые происходит материальный обмен между ядром и цитоплазмой. Пора не является дыркой в ядре, а имеет сложную структуру, организованную несколькими десятками специализированных белков — нуклеопоринов. Под электронным микроскопом она видна как восемь связанных между собой белковых гранул с внешней и столько же с внутренней стороны ядерной оболочки.
Различные структуры клеточного ядра видны из-за накопления в них зелёного флуоресцентного белка
1.3. Ядрышко
Ядрышко находится внутри ядра, и не имеет собственной мембранной оболочки, однако хорошо различимо под световым и электронным микроскопом. Основной функцией ядрышка является синтез рибосом. В геноме клетки имеются специальные участки, так называемые ядрышковые организаторы, содержащие гены рибосомной РНК (рРНК), вокруг которых и формируются ядрышки. В ядрышке происходит синтез рРНК РНК полимеразой I, ее созревание, сборка рибосомных субчастиц. В ядрышке локализуются белки, принимающие участие в этих процессах. Некоторые из этих белков имеют специальную последовательность — сигнал ядрышковой локализации (NoLS, от англ. Nucleolus Localization Signal). Следует отметить, самая высокая концентрация белка в клетке наблюдается именно в ядрышке. В этих структурах было локализовано около 600 видов различных белков, причем считается, что лишь небольшая их часть действительно необходима для осуществления ядрышковых функций, а остальные попадают туда неспецифически.
Под электронным микроскопом в ядрышке выделяют несколько субкомпартментов. Так называемые Фибриллярные центры окружены участками плотного фибриллярного компонента, где и происходит синтез рРНК. Снаружи от плотного фибриллярного компонента расположен гранулярный компонент, представляющий собой скопление созревающих рибосомных субчастиц.
1.4. Ядерный матрикс
Ядерным матриксом некоторые исследователи называют нерастворимый внутриядерный каркас. Считается, что матрикс построен преимущественно из негистоновых белков, формирующих сложную разветвленную сеть, сообщающуюся с ядерной ламиной. Возможно, ядерный матрикс принимает участие в формировании функциональных доменов хроматина. В геноме клетки имеются специальные незначащие А-Т-богатые участки прикрепления к ядерному матриксу (англ. S/MAR — Matrix/Scaffold Attachment Regions), служащие, как предполагается, для заякоривания петель хроматина на белках ядерного матрикса. Впрочем, не все исследователи признают существование ядерного матрикса.

Принципиальная схема реализации генетической информации у про- и эукариот.ПРОКАРИОТЫ. У прокариот синтез белка рибосомой (трансляция) пространственно не отделен от транскрипции и может происходить ещё до завершения синтеза мРНК РНК-полимеразой. Прокариотические мРНК часто полицистронные, то есть содержат несколько независимых генов.ЭУКАРИОТЫ. мРНК эукариот синтезируется в виде предшественника, пре-мРНК, претерпевающего затем сложное стадийное созревание — процессинг, включающий присоединение кэп-структуры к 5'-концу молекулы, присоединение нескольких десятков остатков аденина к ее 3'-концу (полиаденилирование), выщепление незначащих участков — интронов и соединение друг с другом значащих участков — экзонов (сплайсинг). При этом соединение экзонов одной и той же пре-мРНК может проходить разными способами, приводя к образованию разных зрелых мРНК, и в конечном итоге разных вариантов белка (альтернативный сплайсинг). Только мРНК, успешно прошедшая процессинг, экспортируется из ядра в цитоплазму и вовлекается в трансляцию.
2. Эволюционное значение клеточного ядра
Основное функциональное отличие клеток эукариот от клеток прокариот заключается в пространственном разграничении процессов транскрипции (синтеза матричной РНК) и трансляции (синтеза белка рибосомой), что дает в распоряжение эукариотической клетки новые инструменты регуляции биосинтеза и контроля качества мРНК.
В то время, как у прокариот мРНК начинает транслироваться еще до завершения ее синтеза РНК-полимеразой, мРНК эукариот претерпевает значительные модификации (так называемый процессинг), после чего экспортируется через ядерные поры в цитоплазму, и только после этого может вступить в трансляцию. Процессинг мРНК включает несколько элементов.
Из предшественника мРНК (пре-мРНК) в ходе процесса, называемого сплайсингом вырезаются интроны — незначащие участки, а значащие участки — экзоны соединяются друг с другом. Причем экзоны одной и той же пре-мРНК могут быть соединены несколькими разными способами (альтернативный сплайсинг), так что один предшественник может превращаться в зрелые мРНК нескольких разных видов. Таким образом, один ген может кодировать сразу несколько белков.
Кроме того, интрон-экзонная структура генома, практически невозможная у прокариот (так как рибосомы смогут транслировать незрелые мРНК), дает эукариотам определенную эволюционную мобильность. Учитывая протяженность интронных участков, рекомбинация между двумя генами зачастую сводится к обмену экзонами. Благодаря тому, что экзоны часто соответствуют функциональным доменам белка, участки получившегося в результате рекомбинации «гибрида», зачастую сохраняют свои функции. В то же время у прокариот рекомбинация между генами невозможна без разрыва в значащей части, что безусловно уменьшает шансы на то, что получившийся белок будет функционален.
Модификациям подвергаются концы молекулы мРНК. К 5' -концу молекулы прикрепляется 7-метилгуанин (так называемый кэп). К 3'-концу нематрично присоединяются несколько десятков остатков аденина (полиаденирование).
Процессинг мРНК тесно сопряжен с синтезом этих молекул и необходим для контроля качества. Непроцессированная или не полностью процессированная мРНК не сможет выйти из ядра в цитоплазму или будет нестабильна и быстро деградирует. У прокариот нет таких механизмов контроля качества, и из-за этого прокариотические мРНК имеют меньший срок жизни — нельзя допустить, чтобы неправильно синтезированная молекула мРНК, если такая появится, транслировалась в течение долгого времени.
wreferat.baza-referat.ru
Реферат - Клеточное ядро - Производство
Ядро — важнейшая составная часть клетки. Клеточное ядро содержит ДНК, т.е. гены, и, благодаря этому, выполняет две главные функции:
1)хранения и воспроизведения генетической информации
2)регуляции процессов обмена веществ, протекающих в клетке
Безъядерная клетка не может долго существовать, и ядро тоже не способно к самостоятельному_существованию, поэтому цитоплазма и ядро образуют взаимозависимую систему. Большинство клеток имеет одно ядро. Нередко можно наблюдать 2-3 ядра в одной например в клетках печени. Известны и многоядерные клетки, причем число ядер может достигать нескольких десятков. Форма ядра зависит большей частью от формы клетки, она может быть и совершенно неправильной. Различают ядра шаровидные, многолопастные. Впячивания и выросты ядерной оболочки значительно увеличивают поверхность ядра и тем самым усиливают связь ядерных и цитоплазматических структур и веществ.
Строение ядраЯдро окружено оболочкой, которая состоит из двух мембран, имеющих типичное строение. Наружная ядерная мембрана с поверхности, обращенной в цитоплазму, покрыта рибосомами, внутренняя мембрана гладкая.
Ядерная оболочка-часть мембранной системы клетки.Выросты внешней ядерной мембраны соединяются с каналами эндоплазматической сети, образуя единую систему сообщающихся каналов.Обмен веществ между ядром и цитоплазмой осуществляется двумя основными путями.Во-первых, ядерная оболочка пронизана многочисленными порами, через которые происходит обмен молекулами между ядром и цитоплазмой.Во-вторых, вещества из ядра в цитоплазму и обратно могут попадать вследствии отшнуровывания впячиваний и выростов ядерной оболочки.Несмотря на активный обмен веществами между ядром и цитоплазмой, ядерная оболочка ограничивает ядерное содержимое от цитоплазмы, обеспечивая тем самым различия в химическом составе ядерного сока и цитоплазмы.Это необходимо для нормального функционирования ядерных структур.
Содержимое ядра подразделяют на ядерный сок, хроматин и ядрышко.
В живой клетке ядерный сок выглядит бесструктурной массой, заполняющей промежутки между структурами ядра.В состав ядерного сока входят различные белки, в том числе большенство ферментов ядра, белки хроматина и рибосомальные белки.В ядерном соке находятся также свободные нуклеотиды, необходимые для построения молекул ДНК и РНК, аминокислоты, все виды РНК, а также продукты деятельности ядрышка и хроматина, транспортируемые затем из ядра в цитоплазму.
Хроматином (то греч.chroma-окраска, цвет)называют глыбки, гранулы и сетевидные структуры ядра, интенсивно окрашивающиеся некоторыми красителями и отличаются по форме от ядрышка. Хроматин содержит ДНК и белки и представляет собой спирализованные и уплотненные участки хромосом Спирализованные участки хромосом в генетическом отношении неактивны.
Свою специфическую роль-передачу генетической информации-могут осуществлять только деспирализованные-раскрученные участки хромосом, которые в силу своей малой толщины не видны в световой микроскоп.
В делящихся клетках все хромосомысильно спирализуются, укорачиваются и приобретают компактные размеры и форму.Хромосомой называют самостоятельные ядерные структуры, имеющие плечи и первичную перетяжку.Форма хромосом зависит от положения так называемой первичной перетяжки, или центормеры,-области, к которой во время деления клетки(митоза)прикрепляются нити веретена деления. Центромера делит хромосому на два плеча. Расположение центромеры определяет три основных типа хромосом:
1)равноплечие-с плечами равной или почти равной длинны;
2)неравноплечие-с плечами неравной длинны;
3)палочковидные — с одним длинным и вторым очень коротким, иногда с трудом обнаруживаемым плечом. Выделяются еще точечные хромосомы с очень короткими плечами.
Изучение хромосом позволило установить следующие факты.
1.Во всех соматических клетках любого растительного или животного организма число хромосом одинаково.
2.Половые клетки всегда содержат двое меньше хромосом, чем соматические клетки данного вида организма.
3.У всех организмов, относящихся к одному виду, число хромосом в клетках одинаково.
Число хромосом не зависит от уровня организации и не всегда указывает на родство: одно и тоже число их может быть у очень далёких друг от друга систематических групп и может сильно отличаться у близких по происхождению видов.
Таким образом, само по себе число хромосом не является видоспецифическим признаком.Однако характеристика хромосомного набора в целом видоспецифична, т.е. свойственна только одному какому-то виду организмов растений растений или животных.
Совокупность количественных (число и размеры) и качественных (форма) признаковхромосомного набора соматической клетки называюткариотипом.
Число хромосом в кариотипе большинства видов живых организмов четное.Это объясняетя тем, что в соматических клетках находятся две одинаковые по форме и размеру хромосомы-одна из отцовского организма, вторая – из материнского. Хромосомы, одинаковые по форме и размеру и несущие одинаковые гены, называют гомологичными.
Хромосомный набор соматической клетки, в котором каждая хромосома имеет пару, носит название двойного или диплоидного и обозначается 2N. Количество ДНК, соответствующее диплоидному набору хромосом, обозначают 2C.
Из каждой пары гомологичных хромосом в половые клетки попадает только одна, и поэтому хромосомный набор гамет называют одинарным или гаплоидным. Кариотип таких клеток обозначается 2n1c.
Диплоидное число хромосом у животных и растений.
Вид организмов Число хромосом Малярийный плазмодий 2 Сазан 104 Лошадиная аскарида 2 Человек 46 Плодовая мушка дрозофила 8 Ясень обыкновенный 46 Головная вошь 12 Шимпанзе 48 Шпинат 12 Таракан 48 Домашняя муха 12 Перец 48 Тритон 24 Домашняя овца 54> Ель, сосна 24 Домашняя собака 78 Окунь 28 Голубь 80После завершения деления клетки хромосомы диспирализуются, и в ядрах образовавшихся дочерних клеток снова становятся видимыми только тонкая сеточка и глыбки хроматина.
Третья характерная для клетки структура – ядрышко.Оно представляет собой плотное округлое тельце, погруженное в ядерный сок. В ядрах разных клеток, а также в ядре одной и той же клетки в зависимости от её функционального состояния число ядрышек может колебаться от 1 до 5-7 и более. Количество ядрышек может превышать число хромосомом в наборе; это происходит за счет избирательной редупликации генов, отвечающих за синтез р-РНК. Ядрышки есть только в неделящихся ядрах, во время митоза они исчезают вследствие спирализации хромосом и выхода всех ранее образованных рибосом в цитоплазму, а после завершения деления возникают вновь.
Ядрышко не является самостоятельной структурой ядра.Оно образуется вокруг участка хромосомы, в котором закодирована структура р-РНК. Этот участок хромосомы-ген-носит название ядрышкового организатора(ЯО), и на нем происходит синтез р-РНК.
Кроме накопления р-РНК, в ядрышке формируются субъединицы рибосом, которые потом перемещаются в цитоплазму и, объединяясь при участии катионов Ca2+, формируют целостностные рибосомы, способные принимать участие в биосинтезе белка.
Таким образом, ядрышко – это скопление р-РНК и рибосом на разных этапах формирования, в основе которого лежит участок хромосомы, несущий ген – ядрышковый организатор, заключающий наследственную информацию о структуре р –РНК.
www.ronl.ru
Реферат на тему Клеточное ядро
Ядро - важнейшая составная часть клетки. Клеточное ядро содержит ДНК, т.е. гены, и, благодаря этому ,выполняет две главные функции:
1)хранения и воспроизведения генетической информации
2)регуляции процессов обмена веществ, протекающих в клетке
Безъядерная клетка не может долго существовать, и ядро тоже не способно к самостоятельному_существованию, поэтому цитоплазма и ядро образуют взаимозависимую систему. Большинство клеток имеет одно ядро. Нередко можно наблюдать 2-3 ядра в одной например в клетках печени. Известны и многоядерные клетки, причем число ядер может достигать нескольких десятков. Форма ядра зависит большей частью от формы клетки, она может быть и совершенно неправильной. Различают ядра шаровидные, многолопастные. Впячивания и выросты ядерной оболочки значительно увеличивают поверхность ядра и тем самым усиливают связь ядерных и цитоплазматических структур и веществ.
Строение ядра
Ядро окружено оболочкой,которая состоит из двух мембран, имеющих типичное строение. Наружная ядерная мембрана с поверхности,обращенной в цитоплазму, покрыта рибосомами, внутренняя мембрана гладкая.
Ядерная оболочка-часть мембранной системы клетки.Выросты внешней ядерной мембраны соединяются с каналами эндоплазматической сети,образуя единую систему сообщающихся каналов.Обмен веществ между ядром и цитоплазмой осуществляется двумя основными путями.Во-первых,ядерная оболочка пронизана многочисленными порами,через которые происходит обмен молекулами между ядром и цитоплазмой.Во-вторых, вещества из ядра в цитоплазму и обратно могут попадать вследствии отшнуровывания впячиваний и выростов ядерной оболочки.Несмотря на активный обмен веществами между ядром и цитоплазмой, ядерная оболочка ограничивает ядерное содержимое от цитоплазмы,обеспечивая тем самым различия в химическом составе ядерного сока и цитоплазмы.Это необходимо для нормального функционирования ядерных структур.
Содержимое ядра подразделяют на ядерный сок,хроматин и ядрышко.
В живой клетке ядерный сок выглядит бесструктурной массой,заполняющей промежутки между структурами ядра.В состав ядерного сока входят различные белки ,в том числе большенство ферментов ядра, белки хроматина и рибосомальные белки.В ядерном соке находятся также свободные нуклеотиды,необходимые для построения молекул ДНК и РНК ,аминокислоты,все виды РНК, а также продукты деятельности ядрышка и хроматина,транспортируемые затем из ядра в цитоплазму.
Хроматином (то греч.chroma-окраска,цвет)называют глыбки, гранулы и сетевидные структуры ядра, интенсивно окрашивающиеся некоторыми красителями и отличаются по форме от ядрышка. Хроматин содержит ДНК и белки и представляет собой спирализованные и уплотненные участки хромосом Спирализованные участки хромосом в генетическом отношении неактивны.
Свою специфическую роль-передачу генетической информации-могут осуществлять только деспирализованные-раскрученные участки хромосом, которые в силу своей малой толщины не видны в световой микроскоп.
В делящихся клетках все хромосомысильно спирализуются, укорачиваются и приобретают компактные размеры и форму.Хромосомой называют самостоятельные ядерные структуры,имеющие плечи и первичную перетяжку.Форма хромосом зависит от положения так называемой первичной перетяжки, или центормеры,-области,к которой во время деления клетки(митоза)прикрепляются нити веретена деления. Центромера делит хромосому на два плеча. Расположение центромеры определяет три основных типа хромосом:
1)равноплечие-с плечами равной или почти равной длинны;
2)неравноплечие-с плечами неравной длинны;
3)палочковидные - с одним длинным и вторым очень коротким, иногда с трудом обнаруживаемым плечом. Выделяются еще точечные хромосомы с очень короткими плечами.
Изучение хромосом позволило установить следующие факты.
1.Во всех соматических клетках любого растительного или животного организма число хромосом одинаково.
2.Половые клетки всегда содержат двое меньше хромосом, чем соматические клетки данного вида организма.
3.У всех организмов, относящихся к одному виду, число хромосом в клетках одинаково.
Число хромосом не зависит от уровня организации и не всегда указывает на родство:одно и тоже число их может быть у очень далёких друг от друга систематических групп и может сильно отличаться у близких по происхождению видов.
Таким образом,само по себе число хромосом не является видоспецифическим признаком.Однако характеристика хромосомного набора в целом видоспецифична, т.е. свойственна только одному какому-то виду организмов растений растений или животных.
Совокупность количественных (число и размеры) и качественных (форма) признаковхромосомного набора соматической клетки называюткариотипом.
Число хромосом в кариотипе большинства видов живых организмов четное.Это объясняетя тем, что в соматических клетках находятся две одинаковые по форме и размеру хромосомы-одна из отцовского организма , вторая – из материнского. Хромосомы, одинаковые по форме и размеру и несущие одинаковые гены , называют гомологичными.
Хромосомный набор соматической клетки , в котором каждая хромосома имеет пару,носит название двойного или диплоидного и обозначается 2N. Количество ДНК, соответствующее диплоидному набору хромосом , обозначают 2C.
Из каждой пары гомологичных хромосом в половые клетки попадает только одна, и поэтому хромосомный набор гамет называют одинарным или гаплоидным. Кариотип таких клеток обозначается 2n1c.
Диплоидное число хромосом у животных и растений.
| Вид организмов | Число хромосом |
| Малярийный плазмодий | 2 |
| Сазан | 104 |
| Лошадиная аскарида | 2 |
| Человек | 46 |
| Плодовая мушка дрозофила | 8 |
| Ясень обыкновенный | 46 |
| Головная вошь | 12 |
| Шимпанзе | 48 |
| Шпинат | 12 |
| Таракан | 48 |
| Домашняя муха | 12 |
| Перец | 48 |
| Тритон | 24 |
| Домашняя овца | 54> |
| Ель,сосна | 24 |
| Домашняя собака | 78 |
| Окунь | 28 |
| Голубь | 80 |
После завершения деления клетки хромосомы диспирализуются, и в ядрах образовавшихся дочерних клеток снова становятся видимыми только тонкая сеточка и глыбки хроматина.
Третья характерная для клетки структура – ядрышко.Оно представляет собой плотное округлое тельце, погруженное в ядерный сок. В ядрах разных клеток, а также в ядре одной и той же клетки в зависимости от её функционального состояния число ядрышек может колебаться от 1 до 5-7 и более. Количество ядрышек может превышать число хромосомом в наборе; это происходит за счет избирательной редупликации генов, отвечающих за синтез р-РНК. Ядрышки есть только в неделящихся ядрах, во время митоза они исчезают вследствие спирализации хромосом и выхода всех ранее образованных рибосом в цитоплазму, а после завершения деления возникают вновь.
Ядрышко не является самостоятельной структурой ядра.Оно образуется вокруг участка хромосомы, в котором закодирована структура р-РНК. Этот участок хромосомы-ген-носит название ядрышкового организатора(ЯО), и на нем происходит синтез р-РНК.
Кроме накопления р-РНК, в ядрышке формируются субъединицы рибосом, которые потом перемещаются в цитоплазму и, объединяясь при участии катионов Ca2+, формируют целостностные рибосомы, способные принимать участие в биосинтезе белка.
Таким образом, ядрышко – это скопление р-РНК и рибосом на разных этапах формирования, в основе которого лежит участок хромосомы, несущий ген – ядрышковый организатор, заключающий наследственную информацию о структуре р –РНК.
alive-inter.net
Реферат "Структура и функции ядра"
РЕФЕРАТ
“Структура и функции ядра”
Студента Биологического Факультета
группы БХ-21
Михайличенко Михаила.
г. Харьков
1996
Говоря о клеточном ядре, мы имеем в виду собственно ядра эукариотических клеток. Их ядра построены сложным образом и довольно резко отличаются от “ядерных” образований, нуклеоидов, прокариотических организмов. У последних в состав нуклеоидов (ядроподобных структур) входит одиночная кольцевая молекула ДНК, практически лишенная белков. Иногда такую молекулу ДНК бактериальных клеток называют бактериальной хромосомой, или генофором (носителем генов). Бактериальная хромосома не отделена мембранами от основной цитоплазмы, однако собрана в компактную ядерную зону - нуклеоид, который можно видеть в световом микроскопе после специальных окрасок.
Сам термин “ядро” впервые был применен Броуном в 1833 г. Для обозначения шаровидных постоянных структур в клетках растений. Позднее такую же структуру описали во всех клетках высших организмов.
Клеточное ядро обычно одно на клетку (есть примеры многоядерных клеток), состоит из ядерной оболочки, отделяющей его от цитоплазмы, хроматина, ядрышка, кариоплазмы (или ядерного сока) (рис). Эти четыре основных компонента встречаются практически во всех неделящихся клетках эукариоти-ческих одно- и многоклеточных организмов.
Ядра имеют обычно шаровидную или яйцевидную форму; диаметр первых равен приблизительно 10 мкм, а длина вторых - 20 мкм.
Ядро необходимо для жизни клетки, поскольку именно оно регулирует всю ее активность. Связано это с тем, что ядро несет в себе генетическую (наследственную) информацию, заключенную в ДНК.
Ядерная оболочка
Эта структура характерна для всех эукариотических клеток. Ядерная оболочка состоит из внешней и внутренней мембран, разделенных перинуклеарным пространством шириной от 20 до 60 нм. В состав ядерной оболочки входят ядерные поры.
Мембраны ядерной оболочки в морфологическом отношении не отличаются от остальных внутриклеточных мембран: они имеют толщину около 7 нм и состоят из двух осмиофильных слоев.
В общем виде ядерная оболочка может быть представлена, как полый двухслойный мешок, отделяющий содержимое ядра от цитоплазмы. Из всех внутриклеточных мембранных компонентов таким типом расположения мембран обладают только ядро, митохондрии и пластиды. Однако ядерная оболочка имеет характерную особенность, отличающую ее от других мембранных структур клетки. Это наличие особых пор в оболочке ядра, которые образуются за счет многочисленных зон слияний двух ядерных мембран и представляет собой как бы округлые перфорации всей ядерной оболочки.
Строение ядерной оболочки
Внешняя мембрана ядерной оболочки, непосредственно контактирующая с цитоплазмой клетки, имеет ряд сруктурных особенностей, позволяющих отнести ее к собственно мембранной системе эндоплазматического ретикулума. Так, на внешней ядерной мембране обычно располагается большое количество рибосом. У большинства животных и растительных клеток внешняя мембрана ядерной оболочки не представляет собой идеально ровную поверхность - она может образовывать различной величины выпячивания или выросты в сторону цитоплазмы.
Внутренняя мембрана контактирует с хромосомным материалом ядра (см. Ниже).
Наиболее характерной и бросающейся в глаза структурой в ядерной оболочке является ядерная пора. Поры в оболочке образуются за счет слияния двух ядерных мембран в виде округлых сквозных отверстий или перфораций с диаметром 80-90 нм. Округлое сквозное отверстие в ядерной оболочке заполнено сложноорганизованными глобулярными и фибриллярными структурами. Совокупность мембранных перфораций и этих структур называют комплексом пор ядра. Тем самым подчеркивается, что ядерная пора не просто сквозная дыра в ядерной оболочке, через которую непосредственно вещества ядра и цитоплазмы могут сообщаться.
Сложный комплекс пор имеет октагональную симметрию. По границе округлого отверстия в ядерной оболочке располагаются три ряда гранул, по 8 штук в каждом: один ряд лежит со стороны ядра, другой - со стороны цитоплазмы, третий расположен в центральной части пор. Размер гранул около 25 нм. От этих гранул отходят фибриллярные отростки. Такие фибриллы, отходящие от периферических гранул, могут сходиться в центре и создавать как бы перегородку, диафрагму, поперек поры. В центре отверстия часто можно видеть так называемую центральную гранулу.
Число ядерных пор зависит от метаболической активности клеток: чем выше синтетические процессы в клетках, тем больше пор на единицу поверхности клеточного ядра.
Количество ядерных пор в различных объектах
| Объект | Число ядерных пор на мкм2 | Число пор на одно ядро |
| Ксенопус, почки | 10,05 | 3417 |
| Ксенопус, ооцит | 51,0 | 37,6*106 |
| Крыса, гепатоцит | 16,1 | 3816 |
| Мышь, лимфоцит | 3,3 | 403 |
| Человек, лимфоцит | 4,47 | 713 |
Химия ядерной оболочки
В составе ядерных оболочек обнаруживаются небольшие количества ДНК (0-8%), РНК (3-9%), но основными химическими компонентами являются липиды (13-35%) и белки (50-75%), что для всех клеточных мембран.
Состав липидов сходен с таковым в мембранах микросом или мембранах эндоплазматической сети. Ядерные оболочки характеризуются относительно низким содержанием холестерина и высоким - фосфолипидов, обогащенных насыщенными жирными кислотами.
Белковый состав мембранных фракций очень сложен. Среди белков обнаружен ряд ферментов, общих с ЭР (например, глюкозо-6-фосфатаза, Mg-зависимая АТФаза, глютамат-дегидрогеназа и др.) не обнаружена РНК-полимераза. Тут выявлены активности многих окислительных ферментов (цитохромоксидазы, НАДН-цитохром-с-редуктазы) и различных цитохромов.
Среди белковых фракций ядерных мембран встречаются основные белки типа гистонов, что объясняется связью участков хроматина с ядерной оболочкой.
Ядерная оболочка и ядерно-цитоплазматический обмен
Ядерная оболочка - система, разграничивающая два основных клеточных отсека: цитоплазму и ядро. Ядерные оболочки полностью проницаемы для ионов, для веществ малого молекулярного веса, таких, как сахара, аминокислоты, нуклеотиды. Считается, что белки молекулярного веса до 70 тыс. И размером не больше 4,5 нм могут свободно диффундировать через оболочку.
Известен и обратный процесс - перенос веществ из ядра в цитоплазму. Это в первую очередь касается транспорта РНК синтезируещегося исключительно в ядре.
Еще один путь транспорта веществ из ядра в цитоплазму связан с образованием выростов ядерной оболочки, которые могут отделяться от ядра в виде вакуолей, содержимое их затем изливается или выбрасывается в цитоплазму.
* * *
Таким образом, из многочисленных свойств и функциональных нагрузок ядерной оболочки следует подчеркнуть ее роль как барьера, отделяющего содержимое ядра от цитоплазмы, ограничивающего свободный доступ в ядро крупных агрегатов биополимеров, барьера, активно регулирующего транспорт макромолекул между ядром и цитоплазмой.
Одной из основных функций ядерной оболочки следует считать также ее участие в создании внутриядерного порядка, в фиксации хромосомного материала в трехмерном пространстве ядра.
Ядерный матрикс
Этот комплекс не представляет собой какую-то чистую фракцию, сюда входят компоненты и ядерной оболочки, и ядрышка, и кариоплазмы. С ядерным матриксом оказались связаны как гетерогенная РНК, так и часть ДНК. Эти наблюдения дали основание считать, что матрикс ядра играет важную роль не только в поддержании общей структуры интерфазного ядра, но и может участвовать в регуляции синтеза нуклеиновых кислот.
Хроматин
При наблюдении некоторых живых клеток, особенно растительных или же клеток после фиксации и окраски, внутри ядра выявляются зоны плотного вещества. В состав хроматина входит ДНК в комплексе с белком. В интерфазных клетках хроматин может равномерно заполнять объем ядра или же располагаться отдельными сгустками (хромоцентры). Часто он особенно четко выявляется на периферии ядра (пристеночный, примембранный хроматин) или образует внутри ядра переплетения довольно толстых (около 0.3 мкм) и длинных тяжей, образующих подобие внутриядерной цепи.
Хроматин интерфазных ядер представляет собой несущие ДНК тельца (хромосомы), которые теряют в это время свою компактную форму, разрыхляются, деконденсируются. Степень такой деконденсации хромосом может быть различной в ядрах разных клеток. Когда хромосома или ее участок полностью деконденсирован, тогда эти зоны называют диффузным хроматином. При неполном разрыхлении хромосом в интерфазном ядре видны участки конденсированного хроматина (иногда называемого гетерохроматин). Показано, что степень деконденсации хромосомного материала в интерфазе может отражать функциональную нагрузку этой структуры. Чем более диффузен хроматин интерфазного ядра, тем выше в нем синтетические процессы. Падение синтеза РНК в клетках обычно сопровождается увеличением зон конденсированного хроматина.
Максимально конденсирован хроматин во время митотического деления клеток, когда он обнаруживается в виде плотных телец - хромосом. В этот период хромосомы не несут никаких синтетических нагрузок, в них не происходит включение предшественников ДНК и РНК.
Исходя из этого можно считать, что хромосомы клеток могут находиться в двух структурно-функциональных состояниях:
в рабочем, частично или полностью деконденсированном, когда с их участием в интерфазном ядре происходят процессы транскрипции и редупликации;
в неактивном - в состоянии метаболического покоя при максимальной их конденсированности, когда они выполняют функцию распределения и перенося генетического материала в дочерние клетки.
В химическом отношении препараты хроматина представляют собой сложные комплексы дезоксирибонуклеопротеидов, в состав которых входит ДНК и специальные хромосомные белки - гистоны. В составе хроматина обнаружено также РНК. В количественном отношении ДНК, белок и РНК находятся как 1:1,3:0,2. О значении РНК в составе хроматина еще нет достаточно однозначных данных. Возможно, что эта РНК представляет собой сопутствующую препарату функцию синтезирующейся РНК и поэтому частично связанной с ДНК или это особый вид РНК, характерный для структуры хроматина.
ДНК хроматина
В препарате хроматина на долю ДНК приходится обычно 30-40%. Эта ДНК представляет собой двухцепочечную спиральную молекулу. ДНК хроматина обладает молекулярной массой 7-9*106. Такую сравнительно малую массу ДНК из препаратов можно объяснить механическими повреждениями ДНК в процессе выделения хроматина.
Общее количество ДНК, входящее в ядерные структуры клеток, в геном организмов, колеблется от вида к виду. Сравнивая количество ДНК на клетку у эукариотических организмов, трудно уловить какие-либо корреляции между степенью сложности организма и количеством ДНК на ядро. Примерно одинаковое количество ДНК имеют различные организмы, как лен, морской еж, окунь (1,4-1,9 пг) или рыба голец и бык (6,4 и 7 пг).
У некоторых амфибий в ядрах количество ДНК больше, чем в ядрах человека, в 10-30 раз, хотя генетическая конституция человека несравненно сложнее, чем у лягушек. Следовательно, можно предполагать, что “избыточное” количество ДНК у более низко организованных организмов либо не связано с выполнением генетической роли, либо число генов повторяется то или иное число раз.
Сателлитная ДНК, или фракция ДНК с часто повторяющимися последовательностями, может участвовать в узнавании гомологичных районов хромосом при мейозе. По другим предположениям, эти участки играют роль разделителей (спейсеров) между различными функциональными единицами хромосомной ДНК.
Как оказалось, фракция умеренно повторяющихся (от 102 до 105 раз) последовательностей принадлежит к пестрому классу участков ДНК, играющих важную роль в обменных процессах. В эту фракцию входят гены рибосомных ДНК, многократно повторенные участки для синтеза всех тРНК. Более того, некоторые структурные гены, ответственные за синтез определенных белков, также могут быть многократно повторены, представлены многими копиями (гены для белков хроматина - гистонов).
Итак, ДНК эукариотических клеток гетерогенна по составу, содержит несколько классов последовательностей нуклеотидов:
часто повторяющиеся последовательности (>106 раз), входящие во фракцию сателитной ДНК и не транскрибирующиеся;
фракция умеренно повторяющихся последовательностей (102-105), представляющих блоки истинных генов, а также короткие последовательности, разбросанные по всему геному;
фракция уникальных последовательностей, несущая информацию для большинства белков клетки.
ДНК прокариотического организма представляет собой одну гигантскую циклическую молекулу. ДНК эукариотических хромосом представляет собой линейные молекулы, состоящие из тандемно (друг за другом) расположенных репликонов разного размера. Средний размер репликона около 30 мкм. Тем самым в составе генома человека должно встречаться более 50 000 репликонов, участков ДНК, которые синтезируются как независимые единицы. Эти репликоны имеют начальную и терминальную точки синтеза ДНК.
Представим себе, что у эукариотических клеток каждая из хромосомных ДНК, как и у бактерий, является одним репликоном. В этом случае при скорости синтеза 0,5 мкм в минуту (для человека) редупликация первой хромосомы с длиной ДНК около 7 см должна занять 140 000 минут, или около трех месяцев. На самом же деле благодаря полирепликонному строению молекул ДНК весь процесс занимает 7-12 ч.
Белки хроматина
К ним относятся гистоны и негистоновые белки.
Гистоны - сильноосновные белки. Их щелочность связана с их обогащенностью основными аминокислотами (главным образом лизином и аргинином). Эти белки не содержат триптофана. Препарат суммарных гистонов можно разделить на 5 фракций:
Н1 (от английского histone) - богатый лизином гистон, мол. Масса 2100;
Н2а - умеренно богатый лизином гистон, масса 13 700;
Н2б - умеренно богатый лизином гистон, масса 14 500;
Н4 - богатый аргинином гистон, масса 11 300;
Н3 - богатый аргинином гистон, масса 15 300.
В препаратах хроматина эти фракции гистонов обнаруживаются в приблизительно равных количествах, кроме Н1, которого примерно в 2 раза меньше любой из других фракций.
Для молекул гистонов характерно неравномерное распределение основных аминокислот в цепи: обогащенные положительно заряженными аминогруппами наблюдается на концах белковых цепей. Эти участки гистонов связываются с фосфатными группировками на ДНК, в то время как сравнительно менее заряженные центральные участки молекул обеспечивают их взаимодействие между собой. Таким образом, взаимодействие между гистонами и ДНК, приводящее к образованию дезоксирибонуклеопротеинового комплекса, носит ионный характер.
Гистоны синтезируются на полисомах в цитоплазме, этот синтез начинается несколько раньше редупликации ДНК. Синтезированные гистоны мигрируют из цитоплазмы в ядро, где и связываются с участками ДНК.
Функциональная роль гистонов не вполне ясна. Одно время считалось, что гистоны являются специфическими регуляторами активности ДНК хроматина, но одинаковость строения основной массы гистонов говорит о малой вероятности этого. Более очевидна структурная роль гистонов, которая обеспечивает не только специфическую укладку хромосомной ДНК, но и играет роль в регуляции транскрипции.
Негистоновые белки - наиболее плохо охарактеризованная фракция хроматина. Кроме ферментов, непосредственно связанных с хроматином (ферменты, ответственные за репарацию, редубликацию, транскрипцию и модификации ДНК, ферменты модификации гистонов и других белков), в эту фракцию входит множество других белков. Весьма вероятно, что часть негистоновых белков представляет собой специфические белки - регуляторы, узнающие определенные нуклеотидные последовательности в ДНК.
РНК хроматина составляет от 0,2 до 0,5% от содержания ДНК. Эта РНК представляет собой все известные клеточные типы РНК, находящиеся в процессе синтеза или созревания в связи с ДНК хроматина.
В составе хроматина могут быть обнаружены липиды до 1 % от весового содержания ДНК, их роль в структуре и функционировании хромосом остается неясной.
Хромосомы
Первичная степень укладки молекул ДНК - хромосомная фибрилла. Наблюдения за структурой хроматина с помощью электронного микроскопа показали, что в составе ядра на ультратонких срезах всегда видны фибриллярные элементы. Впервые их обнаружил Х. Рис (1957), который и дал им название элементарных хромосомных фибрилл.
Морфология хромосом
Морфологию хромосом лучше всего изучать в момент их наибольшей конденсации, в метафазе и в начале анафазы. Хромосомы животных и растений в этом состоянии представляют собой палочковидные структуры разной длины с довольно постоянной толщиной, у большей части хромосом удается легко найти зону первичной перетяжки, которая делит хромосому на два плеча (рис). Хромосомы с равными или почти равными плечами называют метацентрическими, с плечами неодинаковой длины - субметацентрическими. Палочковидные хромосомы с очень коротким, почти незаметным вторым плечом - акроцентрические.
В области первичной перетяжки расположена центромера, или кинетохор. Это пластинчатая структура, имеющая форму диска. Она связана тонкими фибриллами с телом хромосомы в области перетяжки. От него отрастают пучки микротрубочки митотического веретена, идущие в направлении к центриолям. Они принимают участие в движении хромосом к полюсам клетки при митозе.
Обычно одна хромосома имеет только одну центромеру (моноцентрические хромосомы), но могут встречаться хромосомы дицентрические и полицентрические.
Некоторые хромосомы имеют вторичную перетяжку. Последняя обычно расположена вблизи дистального конца хромосомы и отделяет маленький участок, спутник. Вторичные перетяжки называют, кроме того, ядрышковыми организаторами, так как именно на этих участках хромосом в интерфазе происходит образование ядрышка. Здесь же локализована ДНК, ответственная за синтез рРНК.
Плечи хромосом оканчиваются теломерами, конечными участками. Теломерные концы хромосом не способны соединяться с другими хромосомами или их фрагментами, в отличие от концов хромосом, лишенных теломерных участков, которые могут присоединяться к таким же разорванным концам других хромосом.
Размеры хромосом у разных организмов варьируют в широких пределах. Так, длина хромосом может колебаться от 0,2 до 50 мкм. Самые мелкие хромосомы обнаруживаются у некоторых простейших, грибов. Наиболее длинные - у некоторых прямокрылых насекомых, у амфибий и у лилейных. Длина хромосом человека находится в пределах 1,5-10 мкм.
Число хромосом у различных объектов тоже значительно колеблется, но характерно для каждого вида. У некоторых радиолярий число хромосом достигает 1000-1600. Рекордсменом среди растений по числу хромосом (около 500) является папоротник ужовник, 308 хромосом у тутового дерева, у речного рака 196 хромосом. Наименьшее количество хромосом (2 на диплоидный набор) наблюдается у одной из рас аскариды, у сложноцветного Haplopappus gracilic - всего 4 хромосомы (2 пары).
Совокупность числа, величины, величины и морфологии хромосом называется кариотипом данного вида. Даже у близких видов хромосомные наборы отличаются друг от друга или по числу хромосом, или по величине хотя бы одной или нескольких хромосом. Следовательно, структура кариотипа может быть таксономическим признаком.
Ядрышко
Практически во всех живых клетках эукариотических организмов в ядре видно одно или несколько обычно округлой формы тельц, сильно преломляющих свет, - это ядрышки, или нуклеолы.
Ядрышко - не самостоятельная структура или органоид. Оно - производное хромосомы, один из ее локусов, активно функционирующий в интерфазе.
В процессах синтеза клеточных белков ядрышко клетки является местом образования рибосомных РНК и рибосом, на которых происходит синтез полипептидных цепей.
Количество ядрышек в клетке
Начиная с зеленых водорослей, грибов и низших простейших и кончая высшими организмами, все клетки имеют обязательные внутриядерные структуры - ядрышки. Это правило имеет большое число исключений, которые только подчеркивают важность и необходимость ядрышка в жизненном цикле клетки. К таким исключениям относятся клетки дробящихся яиц, где ядрышки отсутствуют на ранних этапах эмбриогенеза, или клетки закончившие развитие и необратимо специализировавшиеся, например, некоторые клетки крови.
Количество ядрышек в клетке может меняться, однако их число на ядро зависит от генного баланса клетки. Было найдено, что в образовании ядрышек участвуют определенные места некоторых хромосом, связь которых с ядрышком можно хорошо проследить в телофазе и профазе. Такие хромосомы, как правило, имеют вторичные перетяжки, зоны которых представляют собой места, где идет развитие ядрышек. Мак Клинток (1934) назвал эти участки хромосом “ядрышковыми организаторами”.
Места вторичных перетяжек особенно характерны для расположения ядрышковых организаторов, но последние иногда могут находиться на концах хромосом или в нескольких местах по длине хромосомы.
Общее число ядрышек на ядро определяется числом ядрышковых организаторов и увеличивается согласно плоидности ядра. Однако часто количество ядрышек на ядро бывает меньше числа ядрышковых организаторов. Было показано, что ядрышки могут сливаться; кроме того, в образовании одного ядрышка иногда участвует несколько организаторов.
Еще в работах М.С.Навашина (1934) было показано, что хромосомный локус, который в нормальных условиях образует крупное ядрышко, становится неактивным, когда после гибридизации в ядре появляется более “сильный” локус на другой хромосоме. Тот факт, что в определенных условиях может подавляться активность одних ядрышковых организаторов или же повышаться активность других, бывших до этого в латентном, скрытом состоянии, указывает на то, что в клетках поддерживается определенный баланс количества ядрышкового материала или, другими словами, регулируется “валовая” продукция, выдаваемая ядрышками.
Исходя из перечисленных выше фактов, можно сделать следующие заключения:
образования ядрышек и их число связаны с активностью определенных участков хромосом - ядрышковых организаторов, которые расположены большей частью в зонах вторичных перетяжек;
изменения в числе ядрышек в клетках данного типа могут происходить за счет слияния ядрышек или за счет сдвигов в хромосомном балансе клетки.
Физиология и химия ядрышка
Ядрышко по сравнению с другими компонентами клетки характеризуется как самая плотная структура с наиболее высокой концентрацией РНК, с чрезвычайно высокой активностью в отношении синтеза РНК.
Концентрация РНК в ядрышках всегда выше концентрации РНК в других компонентах клетки, так концентрация РНК в ядрышке может быть в 2-8 раз выше, чем в ядре, и в 1-3 раза выше, чем в цитоплазме. Отношение концентрации РНК в ядре, ядрышке и цитоплазме клеток печени мыши составляет 1:7,3:4,1, в клетках поджелудочной железы - 1:9,6:6,6.
В ядрышке не обнаруживается ДНК, но все же при исследовании фиксированных клеток вокруг ядрышка всегда выделяется зона хроматина. Этот околоядрышковый хроматин, по данным электронной микроскопии, представляется, как интегральная часть сложной структуры ядрышка.
Ядрышко - одно из самых активных мест в клетке по включению предшественников в РНК. Ядрышковая РНК является предшественником цитоплазматической РНК.
Цитоплазматическая РНК синтезируется в ядрышке.
РНК ядрышек
Оценивая общее содержание в ядрышковых фракциях белков, РНК и ДНК, можно видеть, что на долю РНК приходится около 10% всей массы ядрышка.
Содержание РНК, ДНК и белка в изолированных ядрышках (сухой вес в %)
| Объект | РНК | ДНК | Белок | РНК/ДНК |
| Печень крысы | 11,0 | 8,0 | 78,0 | 1,4 |
| Регенерирующая печень (6 ч) | 7,6 | 4,6 | 87,8 | 1,7 |
| Регенерирующая печень (18 ч) | 15,5 | 5,4 | 79,1 | 2,9 |
| Печень морской свинки | 4,1 | 9,5 | 86,4 | 0,43 |
| Стебель гороха (4 дня) | 15,11 | 10,6 | 74,0 | 1,5 |
| Проростки гороха (36 ч) | 16,7 | 6,4 | 76,9 | 2,6 |
Так как основную массу цитоплазматической РНК составляет рибосомная РНК, то можно сказать, что ядрышковая РНК принадлежит к этому классу.
Подтверждением представлений того, что именно ядрышко является местом синтеза рРНК и образования рибосом, послужило то, что из ядрышковых препаратов были выделены РНП-частицы, которые как по составу РНК (по седиментационным свойствам), так и по размеру можно охарактиризовать как рибосомы или их предшественники с различными коэффициентами седиментации.
ДНК ядрышек
Биохимическими исследованиями обнаружено в выделенных ядрышках определенное количество ДНК, которую можно отождествить с околоядрышковым хроматином или с ядрышковыми организаторами хромосом. Содержание ДНК в выделенных ядрышках - 5-12% от сухого веса и 6-17% от всей ДНК ядра.
ДНК ядрышкового организатора - это та самая ДНК, на которой происходит синтез ядрышковой, т.е. рибосомной, РНК.
Таким образом из биохимических работ появились представления о том, что в ядрышке на ДНК локализованы многочисленные одинаковые гены для синтеза рРНК. Синтез рРНК идет путем образования огромного предшественника и дальнейшего его превращения (созревания) в более короткие молекулы РНК для большой и малой субъедениц рибосом.
Изучая ядрышки ооцитов тритонов, исследователи столкнулись с интересным явлением - сверхчисленностью ядрышек. У X. laevis во время роста ооцита появляется до 1000 мелких ядрышек, не связанных с хромосомами. Именно эти ядрышки выделил О.Миллер. вместе с этим на ядро ооцита увеличивается количество рДНК. Это явление получило название амплификации. Оно заключается в том, что происходит сверхрепликация зоны ядрышкового организатора, многочисленные копии отходят от хромосом и становятся дополнительно работающими ядрышками. Такой процесс необходим для накопления огромного (1012) количества рибосом на яйцевую клетку, что обеспечит в будущем развитие эмбриона на ранних стадиях даже при отсутствии синтеза новых рибосом. Сверхчисленные ядрышки после созревания яйцевой клетки исчезают.
Ультраструктура ядрышек
При изучении большого числа различных клеток животных и растений отмечена волокнистая или сетчатая структура ядрышек, заключенная в более или менее плотную диффузную массу. Были предложены названия для этих частей: волокнистая часть - нуклеонема и диффузная, гомогенная часть - аморфное вещество, или аморфная часть. Сделанные почти одновременно с этим электронно-микроскопичес-кие исследования также выявили волокнисто-нитчатое строение ядрышек.
Однако такое нитчатое строение ядрышка не всегда четко выражено. У некоторых клеток отдельные нити нуклеонем сливаются, и ядрышки могут быть совершенно однородными.
При более пристальном изучении ядрышка можно заметить, что основные структурные компоненты ядрышка - плотные гранулы диаметром около 15 нм и тонкие фибриллы толщиной 4-8 нм. Во многих случаях (ооциты рыб и амфибий, меристематические клетки растений) фибриллярный компонент собран в плотную центральную зону (сердцевина), лишенную гранул, а гранулы занимают переферическую зону ядрышка. В ряде случаев (например, клетки корешков растений) в этой гранулярной зоне не наблюдается никакой дополнительной структуризации.
Было найдено, что аморфные участки ядрышек неоднородны. В их структуре выявляются малоокрашенные зоны - фибриллярные центры - и окружающие их более темные участки, тоже имеющие фибриллярное строение.
Кроме этих двух компонентов ядрышек в последнее время большое внимание уделялось строению околоядрышкового хроматина. Этот хроматин и внутриядрышковая сеть ДНК являются единой системой и представляют собой интегральный компонент ядрышка.
Гранулы и фибриллярная часть состоят из рибонуклеопротеидов.
Показано, что именно светлые фибриллярные центры содержат рДНК.
Судьба ядрышка при делении клеток
Известно, что ядрышко исчезает в профазе и появляется вновь в средней телофазе.
По мере затухания синтеза рРНК в средней профазе происходит разрыхление ядрышка и выход готовых рибосом в кариоплазму, а затем и в цитоплазму. При конденсации профазных хромосом фибриллярный компонент ядрышка и часть гранул тесно ассоциируют с их поверхностью, образуя основу матрикса митотических хромосом. Этот фибриллярно-гранулярный материал, синтезированный до митоза, переносится хромосомами в дочерние клетки.
В ранней телофазе по мере деконденсации хромосом происходит высвобождение компонентов матрикса. Его фибриллярная часть начинает собираться в мелкие многочисленные ассоциаты - предъядрышки, которые могут объединяться друг с другом. По мере возобновления синтеза РНК предъядрышки претерпевают перестройку, что выражается в появлении в их структуре гранул РНК, а затем в становлении дефинитивной формы нормально функционирующего ядрышка.
Роль ядра.
Ядро осуществляет две группы общих функций: одну, связанную собственно с хранением генетической информации, другую - с ее реализацией, с обеспечением синтеза белка.
В первую группу входят процессы, связанные с поддержанием наследственной информации в виде неизменной структуры ДНК. Эти процессы связаны с наличием так называемых репарационных ферментов, ликвидирующих спонтанные повреждения молекулы ДНК (разрыв одной из цепей ДНК, часть радиационных повреждений), что сохраняет строение молекул ДНК практически неизменным в ряду поколений клеток или организмов. Далее, в ядре происходит воспроизведение или редупликация молекул ДНК, что дает возможность двум клеткам получить совершенно одинаковые и в качественном и в количественном смысле объемы генетической информации. В ядрах происходят процессы изменения и рекомбинации генетического материала, что наблюдается во время мейоза (кроссинговер). Наконец, ядра непосредственно участвуют в процессах распределения молекул ДНК при делении клеток.
Другой группой клеточных процессов, обеспечивающихся активностью ядра, является создание собственно аппарата белкового синтеза. Это не только синтез, транскрипция на молекулах ДНК разных информационных РНК и рибосомных РНК. В ядре эукариотов происходит также образование субъедениц рибосом путем комплексирования синтезированных в ядрышке рибосомных РНК с рибосомными белками, которые синтезируются в цитоплазме и переносятся в ядро.
Таким образом, ядро представляет собой не только вместилище генетического материала, но и место, где этот материал функционирует и воспроизводится. Поэтому выпадение лил нарушение любой из перечисленных выше функций губительно для клетки в целом. Так нарушение репарационных процессов будет приводить к изменению первичной структуры ДНК и автоматически к изменению структуры белков, что непременно скажется на их специфической активности, которая может просто исчезнуть или измениться так, что не будет обеспечивать клеточные функции, в результате чего клетка погибает. Нарушения редупликации ДНК приведут к остановке размножения клеток или к появлению клеток с неполноценным набором генетической информации, что также губительно для клеток. К такому же результату приведет нарушение процессов распределения генетического материала (молекул ДНК) при делении клеток. Выпадение в результате поражения ядра или в случае нарушений каких-либо регуляторных процессов синтеза любой формы РНК автоматически приведет к остановке синтеза белка в клетке или к грубым его нарушениям.
Значение ядра как хранилища генетического материала и его главная роль в определении фенотипических признаков были установлены давно. Немецкий биолог Хаммерлинг одним из первых продемонстрировал важнейшую роль ядра. Он выбрал в качестве объекта своих экспериментов необычайно крупную одноклеточную (или неклеточную) морскую водоросль Acetabularia. Существует два близко родственных вида A. medierranea и A. crenulata, различающихся только по форме “шляпки”.
В ряде экспериментов, в том числе таких, в которых “шляпку” отделяли от нижней части “стебелька” (где находится ядро), Хаммерлинг показал, что для нормального развития шляпки необходимо ядро. В дальнейших экспериментах, в которых соединяли нижнюю часть, содержащую ядро одного вида с лишенным ядра стебельком другого вида, у таких химер всегда развивалась шляпка, типичная для того вида, которому принадлежит ядро.
При оценке этой модели ядерного контроля следует, однако, учитывать примитивность организма, использованного в качестве объекта. Метод пересадок был применен позднее в экспериментах, проведенных в 1952 г. двумя американскими исследователями, Бриггсом и Кингом, с клетками лягушки Rana pipenis. Эти авторы удаляли из неоплодотворенных яйцеклеток ядра и заменяли их ядрами из клеток поздней бластулы, уже проявлявших признаки дифференцировки. Во многих случаях из яиц реципиентов развивались нормальные взрослые лягушки.
Литература:
Свенсон К., Уэбстер П. “Клетка”. М., Мир, 1980.
Де Робертис Э. Новинский В., Саэс Ф. “Биология Клетки”. М., Мир, 1971
Ченцов Ю.С., Поляков В.Ю. “Ультраструктура клеточного ядра”. М., Наука, 1974
Зегнбуш П. “Молекулярная и клеточная биология”. М., Мир, т.1,2, 1982
При подготовке реферата были использованы материалы, полученные из Всемирной Биологической Сети (BIOSCI) посредством сети Internet.
refdb.ru
Реферат - Атомное ядро - Рефераты на репетирем.ру
Реферат по физикена тему:“Атомное ядро”
Москва 1996
Оглавление
1 История открытий в области строения атомного ядра
1.1 Модели атома до Бора
1.2 Открытие атомного ядра
1.3 Атом Бора
1.4 Расщепление ядра
1.5 Протонно-нейтронная модель ядра
1.6 Искусственная радиоактивность
2 Строение и важнейшие свойства атомных ядер
2.1 Основные свойства и строение ядра
2.2 Энергия связи ядер. Дефект массы
2.3 Ядерные силы
2.4 Радиоактивность, g-излучение, a и b-распад
Литература
1 История открытий в области строения атомного ядра
Изучение атомного ядра вынуждает заниматься элементарными частицами. Причина этого ясна: в ядрах атомов частиц так мало, что свойства каждой из них в отдельности не усредняются, а, напротив, играют определяющую роль.
1.1 Модели атома до Бора
Развитие исследований радиоактивного излучения, с одной стороны, и квантовой теории - с другой, привели к созданию квантовой модели атома Резерфорда - Бора. Но созданию этой модели предшествовали попытки построить модель атома на основе представлений классической электродинамики и механики. В 1904 году появились публикации о строении атома, одни из которых принадлежали японскому физику Хантаро Нагаока, другие - английскому физику Д.Д. Томсону.
Нагаока представил строение атома аналогичным строению солнечной системы: роль Солнца играет положительно заряженная центральная часть атома, вокруг которой по установленным кольцеобразным орбитам движутся “планеты” - электроны. При незначительных смещениях электроны возбуждают электромагнитные волны.
В атоме Томсона положительное электричество “распределено” по сфере, в которую вкраплены электроны. В простейшем атоме водорода электрон находится в центре положительно заряженной сферы. В многоэлектронных атомах электроны располагаются по устойчивым конфигурациям, рассчитанным Томсоном. Томсон считал каждую такую конфигурацию определяющей химические свойства атомов. Он предпринял попытку теоретически объяснить периодическую систему элементов Д.И. Менделеева. Позднее Бор указал, что со времени этой попытки идея о разделении электронов в атоме на группы сделалась исходным пунктом.
Но вскоре оказалось, что новые опытные факты опровергают модель Томсона и, наоборот, свидетельствуют в пользу планетарной модели. Эти факты были открыты Резерфордом. В первую очередь следует отметить открытие ядерного строения атома.
1.2 Открытие атомного ядра
Уподобление атома планетной системе делалось еще в начале XX века. Но эту модель было трудно совместить с моделями электродинамики, и она была оставлена, уступив место модели Томсона. Однако в 1904 году начались исследования, приведшие к утверждению планетарной модели.
При изучении a-частиц Резерфорд, исходя из модели Томсона, подсчитал, что рассеивание a-частиц не может давать больших углов отклонений даже при многих столкновениях с частицей. И здесь Резерфорд обратился к планетарной модели.
7 марта 1911 года Резерфорд сделал в философском обществе в Манчестере доклад “Рассеяние a и b-лучей и строение атома”. В докладе он, в частности, говорил: “Рассеяние заряженных частиц может быть объяснено, если предположить такой атом, который состоит из центрального электрического заряда, сосредоточенного в точке и окруженного однородным сферическим распределением противоположного электричества равной величины. При таком устройстве a и b-частицы, когда они проходят на близком расстоянии от центра атома, испытывают большие отклонения, хотя вероятность такого отклонения мала”.
Важным следствием теории Резерфорда было указание на заряд атомного центра, который Резерфорд положил равным ±Ne. Заряд оказался пропорциональным атомному весу. “Точное значение заряда центрального ядра не было определено,- писал Резерфорд, - но для атома золота оно приблизительно равно 100 единицам заряда”.
Из последующих исследований и экспериментов Гейгера и Мардсена, предпринявших проверку формул Резерфорда, возникло представление о ядре как устойчивой части атома, несущей в себе почти всю массу атома и обладающей положительным (Резерфорд считал знак заряда неопределенным) зарядом. При этом число элементарных зарядов оказалось пропорциональным атомному весу.
Заряд ядра оказался важнейшей характеристикой атома. В 1913 году было показано, что заряд ядра совпадает с номером элемента в таблице Менделеева. Бор писал: ”С самого начала было ясно, что благодаря большой массе ядра и его малой протяженности в пространстве сравнительно с размерами всего атома строение электронной системы должно зависеть почти исключительно от полного электрического заряда ядра. Такие рассуждения сразу наводили на мысль о том, что вся совокупность физических и химических свойств каждого элемента может определяться одним целым числом...”
После знакомства с Резерфордом Бор, отказавшись от изучения электронной модели, начал работу в его группе. Обратившись к планетарной модели, Бор создал на ее основе теорию атома Резерфорда-Бора. Резерфорд понял революционный характер идей Бора и обсудил с ним основы этой теории, высказал критические замечания, после чего статьи Бора были опубликованы.
Во время Первой Мировой войны Бор продолжает работать в лаборатории Резерфорда. В 1915 году он опубликовал работы “О сериальном спектре водорода” и “О квантовой теории излучения в структуре атома”. В 1916 году была опубликована статья Зоммерфельда, где он рассмотрел движение электрона по эллиптическим орбитам и обобщил правила квантования Бора. Бор с восторгом отозвался об этой статье. Теория атома после открытий Зоммерфельда стала называться теорией Бора - Зоммерфельда.
В 1936 году Бор выступил со статьей “Захват нейтрона и строение ядра”, в которой предложил капельную модель ядра и механизм захвата нейтрона ядром. Странно, но ни Бор, ни другие не могли сразу предсказать деление ядра, подсказываемое капельной моделью, пока в начале 1939 г. не было открыто деление урана.
1.3 Атом Бора
Бор, как и Томсон до него, ищет такое расположение электронов в атоме, которое объяснило бы его физические и химические свойства. Бор берет за основу модель Резерфорда. Ему также известно, что заряд ядра и число электронов в нем, равное числу единиц заряда, определяется местом элемента в периодической системе элементов Менделеева. Таким образом, это важный шаг в понимании физико-химических свойств элемента. Но остаются непонятными две вещи: необычайная устойчивость атомов, несовместимая с представлением о движении электронов по замкнутым орбитам, и происхождение их спектров, состоящих из вполне определенных линий. Такая определенность спектра, его ярко выраженная химическая индивидуальность, очевидно, как-то связана со структурой атома. Все это трудно увязать с универсальностью электрона, заряд и масса которого не зависят от природы атома, в состав которого они входят. Устойчивость атома в целом противоречит законам электродинамики, согласно которым электроны, совершая периодические движения, должны непрерывно излучать энергию и, теряя ее, “падать” на ядро. К тому же и характер движения электрона, объясняемый законами электродинамики, не может приводить к таким характерным линейчатым спектрам, которые наблюдаются на самом деле. Линии спектра группируются в серии, они сгущаются в коротковолновом “хвосте” серии, частоты линий соответствующих серий подчинены странным арифметическим законам.
“Основным результатом тщательного анализа видимой серии линейчатых спектров и их взаимоотношений, - писал Бор, - было установление того факта, что частота u каждой линии спектра данного элемента может быть представлена с необыкновенной точностью формулой u = Tґ - Tґґ, где Tґ и Tґґ - какие-то два члена из множества спектральных элементов Т, характеризующих элемент”.
Бору удалось найти объяснение этого основного закона спектроскопии. Но для этого ему пришлось ввести в физику атома представления о стационарных состояниях атомов, находясь в которых электрон не излучает, хотя и совершает периодическое движение по круговой орбите.
1.4 Расщепление ядра
В 1919 году Резерфордом было сделано новое сенсационное открытие - расщепление ядра.
Резерфорд изучал столкновение a-частиц с легкими атомами. Столкновения a-частицы с ядрами таких атомов должны их ускорять. Так, при ударе a-частицы о ядро водорода оно увеличивает свою скорость в 1,6 раза, и ядро отбирает у a-частицы 64% ее энергии.
Прибор, применявшийся Резерфордом для излучения таких столкновений, представлял собой латунную камеру длиной 18 см, высотой 6 см и шириной 2 см. Источником a-частиц служил металлический диск, покрытый активным веществом. Диск помещался внутри камеры и мог устанавливаться на разных расстояниях от экрана из сернистого цинка. Камера могла заполняться различными газами. В частности, ее заполняли азотом.
С помощью многочисленных опытов Резерфорд показал, что в результате таких столкновений получаются частицы с максимальным пробегом, таким же, как у Н-атомов. “Из полученных до сих пор результатов, - писал Резерфорд, - трудно избежать заключения, что атомы с большим пробегом, возникающие при столкновении a-частиц с азотом, являются не атомами азота, но, по всей вероятности, атомами водорода или атомами с массой 2. Если это так, то мы должны заключить, что атом азота распадается вследствие громадных сил, развивающихся при столкновении с быстрой a-частицей, и что освобождающийся водородный атом образует составную часть атома”.
Так было открыто явление расщепления ядер азота при ударах быстрых a-частиц и впервые высказана мысль, что ядра водорода представляют собой составную часть ядер атомов. Впоследствии Резерфорд предложил термин “протон” для этой составной части ядра. Резерфорд заканчивал свою статью словами: “Результаты в целом указывают на то, что если a-частицы или подобные им быстро движущиеся частицы со значительно большей энергией могли бы применяться для опытов, то можно было бы обнаружить разрушение ядерных структур многих легких атомов”.
В 1920 году Резерфорд в лекции “Нуклеарное строение атома” делает предположение о том, что существуют ядра с массой 3 и 2 и ядра с массой ядра водорода, но с нулевым зарядом. При этом он исходил из гипотезы, высказанной впервые М. Склодовской-Кюри, что в состав ядра входят электроны.
Резерфорд пишет, что ему “кажется весьма правдоподобным, что один электрон может связать два Н-ядра и, возможно, даже и одно Н-ядро. Если справедливо первое предположение, то оно указывает на возможность существования атома с массой около 2 и с одним зарядом. Такое вещество нужно рассматривать как изотоп водорода. Второе предположение заключает в себе мысль о возможности существования атома с массой 1 и нуклеарным зарядом, равным нулю. Подобные образования представляются вполне возможными”. Так была высказана гипотеза о существовании нейтрона и тяжелого изотопа водорода.
1.5 Протонно-нейтронная модель ядра
В 1932 году Д.Д. Иваненко опубликовал заметку, в которой высказал предположение, что нейтрон является наряду с протоном структурным элементом ядра. Однако протонно-нейтронная модель ядра была встречена большинством физиков скептически. Даже Резерфорд полагал, что нейтрон - это лишь сложное образование протона и электрона.
В 1933 году Иваненко на конференции в Ленинграде сделал доклад о модели ядра, в котором он защищал протонно-нейтронную модель, сформулировав основной тезис: в ядре имеются только тяжелые частицы. Иваненко отверг идеи о сложной структуре нейтрона и протона. По его мнению, обе частицы должны обладать одинаковой степенью элементарности, т.е. и нейтрон, и протон могут переходить друг в друга. В дальнейшем протон и нейтрон стали рассматриваться как два состояния одной частицы - нуклона, и идея Иваненко стала общепринятой, а в 1932 году в составе космических лучей была открыта еще одна элементарная частица - позитрон.
1.6 Искусственная радиоактивность
В 1934 году Фредерик Жолио и Ирен Кюри сообщили о б открытии ими нового вида радиоактивности. Им удалось доказать методом камеры Вильсона, что некоторые легкие элементы (бериллий, бор, алюминий) испускают положительные электроны при бомбардировке их a-частицами полония. Жолио и Кюри, исследуя это явление, показали, что в этом случае возникает новый этап радиоактивности, сопровождаемый испусканием положительных электронов. Они впервые искусственно вызвали радиоактивность, создав новые радиоактивные изотопы, не наблюдаемые до этого в природе и были награждены за это выдающееся открытие Нобелевской премией.
На сегодняшний день теория атомного ядра получила дальнейшее развитие, и в следующей главе рассматривается ее актуальное состояние.
2 Строение и важнейшие свойства атомных ядер
2.1 Основные свойства и строение ядра
1. Ядром называется центральная часть атома, в которой сосредоточена практически вся масса атома и его положительный электрический заряд. Все атомные ядра состоят из элементарных частиц: протонов и нейтронов, которые считаются двумя зарядовыми состояниями одной частицы - нуклона. Протон имеет положительный электрический заряд, равный по абсолютной величине заряду электрона. Нейтрон не имеет электрического заряда.
2. Зарядом ядра называется величина Ze, где е - величина заряда протона, Z - порядковый номер химического элемента в периодической системе Менделеева, равный числу протонов в ядре. В настоящее время известны ядра с Z от Z=1 до Z=107. Для всех ядер, кроме и некоторых других нейтронодефицитных ядер NіZ, где N - число нейтронов в ядре. Для легких ядер N/Z”1; для ядер химических элементов, расположенных в конце периодической системы, N/Z”1,6.
3. Число нуклонов в ядре A=N+Z называется массовым числом. Нуклонам (протону и нейтрону) приписывается массовое число, равное единице, электрону - нулевое значение А.
Ядра с одинаковыми Z, но различными А называются изотопами. Ядра, которые при одинаковом А имеют различные Z, называются изобарами. Ядро химического элемента X обозначается , где Х - символ химического элемента.
Всего известно около 300 устойчивых изотопов химических элементов и более 2000 естественных и искусственно полученных радиоактивных изотопов.
4. Размер ядра характеризуется радиусом ядра, имеющим условный смысл ввиду размытости границы ядра. Эмпирическая формула для радиуса ядра м, может быть истолкована как пропорциональность объема ядра числу нуклонов в нем.
Плотность ядерного вещества составляет по порядку величины 1017 кг/м3 и постоянна для всех ядер. Она значительно превосходит плотности самых плотных обычных веществ.
5. Ядерные частицы имеют собственные магнитные моменты, которыми определяется магнитный момент ядра Рmяд в целом. Единицей измерения магнитных моментов ядер служит ядерный магнетон mяд:
(в СИ)
(в СГС).
Здесь е - абсолютная величина заряда электрона, mp - масса протона, с - электродинамическая постоянная. Ядерный магнетон в раз меньше магнетона Бора, откуда следует, что магнитные свойства атомов определяются магнитными свойствами его электронов.
6. Распределение электрического заряда протонов по ядру в общем случае несимметрично. Мерой отклонения этого распределения от сферически симметричного является квадрупольный электрический момент ядра Q. Если плотность заряда считается везде одинаковой, то Q определяется только формой ядра.
2.2 Энергия связи ядер. Дефект массы
1. Нуклоны в ядрах находятся в состояниях, существенно отличающихся от их свободных состояний. За исключением ядра обычного водорода во всех ядрах имеется не менее двух нуклонов, между которыми существует особое ядерное сильное взаимодействие - притяжение - обеспечивающее устойчивость ядер, несмотря на отталкивание одноименно заряженных протонов.
2. Энергией связи нуклона в ядре называется физическая величина, равная той работе, которую нужно совершить для удаления нуклона из ядра без сообщения ему кинетической энергии.
Энергия связи ядра определяется величиной той работы, которую нужно совершить, чтобы расщепить ядро на составляющие его нуклоны без придания им кинетической энергии. Из закона сохранения энергии следует, что при образовании ядра должна выделяться такая же энергия, какую нужно затратить при расщеплении ядра на составляющие его нуклоны. Энергия связи ядра является разностью между энергией всех свободных нуклонов, составляющих ядро, и их энергией в ядре.
3. При образовании ядра происходит уменьшение его массы: масса ядра меньше, чем сумма масс составляющих его нуклонов. Уменьшение массы ядра при его образовании объясняется выделением энергии связи. Если Wсв - величина энергии, выделяющейся при образовании ядра, то соответствующая ей масса Dm, равная
называется дефектом массы и характеризует уменьшение суммарной массы при образовании ядра из составляющих его нуклонов. Если ядро с массой Mяд образовано из Z протонов с массой mp и из (A-Z) нейтронов с массой mn, то
Dm=Zmp+(A-Z)mn-Mяд.
Вместо массы ядра Мяд величину Dm можно выразить через атомную массу Мат:
Dm=ZmН+(A-Z)mn-Mат,
где mH - масса водородного атома.
При практическом вычислении Dm массы всех частиц и атомов выражаются в атомных единицах массы.
Дефект массы служит мерой энергии связи ядра:
Wсв=Dmс2=[Zmp+(A-Z)mn-Mяд]с2
Одной атомной единице массы соответствует атомная единица энергии (а.е.э.): а.е.э.=931,5016 МэВ.
4. Удельной энергией связи ядра wсв называется энергия связи, приходящаяся на один нуклон: wсв=. Величина wсв составляет в среднем 8 МэВ/нуклон. По мере увеличения числа нуклонов в ядре удельная энергия связи убывает.
5. Критерием устойчивости атомных ядер является соотношение между числом протонов и нейтронов в устойчивом ядре для данных изобаров. (А=const).
2.3 Ядерные силы
1. Ядерное взаимодействие свидетельствует о том, что в ядрах существуют особые ядерные силы, не сводящиеся ни к одному из типов сил, известных в классической физике (гравитационных и электромагнитных).
2. Ядерные силы являются короткодействующими силами. Они проявляются лишь на весьма малых расстояниях между нуклонами в ядре порядка 10-15 м. Длина (1,5ј2,2)10-15 м называется радиусом действия ядерных сил.
3. Ядерные силы обнаруживают зарядовую независимость: притяжение между двумя нуклонами одинаково независимо от зарядового состояния нуклонов - протонного или нуклонного. Зарядовая независимость ядерных сил видна из сравнения энергий связи в зеркальных ядрах. Так называются ядра, в которых одинаково общее число нуклонов, но число протонов в одном равно числу нейтронов в другом. Например, ядра гелия тяжелого водорода трития - .
4. Ядерные силы обладают свойством насыщения, которое проявляется в том, что нуклон в ядре взаимодействует лишь с ограниченным числом ближайших к нему соседних нуклонов. Именно поэтому наблюдается линейная зависимость энергий связи ядер от их массовых чисел А. Практически полное насыщение ядерных сил достигается у a-частицы, которая является очень устойчивым образованием.
2.4 Радиоактивность, g-излучение, a и b-распад
1. Радиоактивностью называется превращение неустойчивых изотопов одного химического элемента в изотопы другого элемента, сопровождающееся испусканием некоторых частиц.
Естественной радиоактивностью называется радиоактивность, наблюдающаяся у существующих в природе неустойчивых изотопов.
Искусственной радиоактивностью называется радиоактивность изотопов, полученных в результате ядерных реакций.
2. Обычно все типы радиоактивности сопровождаются испусканием гамма-излучения - жесткого, коротковолнового электроволнового излучения. Гамма-излучение является основной формой уменьшения энергии возбужденных продуктов радиоактивных превращений. Ядро, испытывающее радиоактивный распад, называется материнским; возникающее дочернее ядро, как правило, оказывается возбужденным, и его переход в основное состояние сопровождается испусканием g-фотона.
3. Альфа-распадом называется испускание ядрами некоторых химических элементов a-частиц. Альфа-распад является свойством тяжелых ядер с массовыми числами А>200 и зарядами ядер Ze>82. Внутри таких ядер происходит образование обособленных a-частиц, состоящих каждая из двух протонов и двух нейтронов.
4. Термином бета-распад обозначают три типа ядерных превращений: электронный (b-) и позитронный (b+) распады, а также электронный захват. Первые два типа превращения состоят в том, что ядро испускает электрон (позитрон) и электронное антинейтрино (электронное нейтрино). Эти процессы происходят путем превращения одного вида нуклона в ядре в другой: нейтрона в протон или протона в нейтрон. В случае электронного захвата превращение заключается в том, что исчезает один из электронов в ближайшем к ядру слое. Протон, превращаясь в нейтрон, как бы “захватывает” электрон; отсюда произошел термин ”электронный захват”. Электронный захват в отличие от b± -захвата сопровождается характеристическим рентгеновским излучением.
5. b--распад происходит у естественно-радиоактивных, а также искусственно-радиоактивных ядер; b+-распад характерен только для явления искусственной радиоактивности.
Литература
1. Григорьев В.И., Мякишев Г.Я. Силы в природе.// М., Наука, 1983 г.
2. Кудрявцев П.С. Курс истории физики.// М., Просвещение, 1982 г.
3. Яворский Б.М., Детлаф А.А. Справочник по физике.// М., Наука, 1990 г.
ref.repetiruem.ru



|
|
..:::Счетчики:::.. |
|
|
|
|
|
|
|
|