
Элла Пуркалина
В последние годы было сделано удивительное открытие, что именно комбинация витаминов А, С, Е является самой главной для здоровья человека. (витамин А в форме бета-каротина, присутствующего в ярко окрашенных фруктах и овощах).
Эти витамины защищают наш организм от повреждений, вызванных избытком свободных радикалов — заряженных, чрезвычайно агрессивных частиц, которые постоянно образуются внутри тела.
Пока ты читашь это предложение, уже образовались тысячи свободных радикалов.
Витамины A,C,E способны нейтрализовать эти частицы, прежде чем они смогут повредить всем клеткам организма.
Большое количество свободных радикалов в организме в высшей степени вредны, хотя небольшое их количество эффективно борется с бактериями и вирусами.
Проблема возникает тогда, когда множество образовавшихся радикалов выходит из под контроля и начинает атаковать само тело.
Свободные радикалы — фрагменты молекул, имеющие неспаренный электрон. От этого они становятся очень неустойчивыми.
Из-за того, что электроны предпочитают соединяться в пары, они пытаются «ухватить» электрон любой ближайшей молекулы для равновесия.
Свободный радикал имеет свободный электрон, ищущий пару, чтобы «успокоиться».
К несчастью, в поисках партнера он разбивает другие пары и создает в этом процессе множество других неустойчивых соединений.
Короче говоря, в организме проходит цепная реакция и повреждение клеток.
Кроме витаминов A,C,E есть еще ряд элементов, которые помогают бороться со свободными радикалами.
Надо признать, что в организме первая линия обороны в борьбе с повреждениями, вызванными свободными радикалами, проходит через группу ферментов, которые содержат ионы металлов: марганца, меди, цинка и селена.
Один из них — супероксиддисмутаза (СОД) — находится на первом плане в исследованиях заболеваний сердца и онкологии.
Другой, тесно связанный с антиоксидантом фермент — глютатионпероксидаза (ГП) также рассматривается, как многообещающее средство предотвращения онкологических заболеваний.
Когда у нас нет селена, то нет и глутатион-пероксидазы — мощного антиокислительного фермента.
Его отсутствие оставляет громадную брешь в нашей обороне от болезней, связанных с окислением, в том числе атеросклероза, сердечно — сосудистых заболеваний, ревматоидного артрита и катаракта.
Ферменты контролируют все химические изменения, которые происходят в клетках, включая образование и высвобождение энергии, поддерживающей жизнедеятельность организма и хорошее самочувствие.
Ферменты — это катализаторы, они ускоряют химические процессы в клетках нашего организма.
Но для такой работы им нужна поддержка, требуются ионы металлов, которые усиливают действие различных ферментов.
Нет сомнений, что всем нам необходимо защищать себя от вредоносного действия свободных радикалов, потребляя с пищей побольше антиоксидантов.
Вред, наносимый клеткам свободными радикалами, ощущает весь наш организм, и сейчас считают, что именно из-за этого возникают сердечные заболевания.
Свободные радикалы бывают причиной заболевания диабетом, катаракты, артрита, морщинистой кожи и даже самого процесса старения организма.
Антиоксиданты — идеальное оружие для нашего организма, чтобы бороться с повреждениями, которые наносят огромные количества свободных радикалов.
Съедать большие количества продуктов, богатых бета — каротином, витамином С и витамином Е — вот, что необходимо для поддержания крепкого здоровья, хотя нужное количество их просто ошеломляет.
Например, для того, чтобы получить 15 мг бета — каротина, нам нужно съесть почти килограмм абрикосов, чтобы получить 100 мг витамина С — больше килограмма картошки, чтобы получить 75 мг витамина Е — почти четыре килограмма сливочного масла.
Для получения дневной нормы селена надо съедать более 7 килограммов моркови ежедневно.
Из этого можно заключить, что нам катастрофически не хватает важнейших антиоксидантов ежедневно. (Потом мы удивляемся, откуда берутся все наши болезни).
Поэтому прием суточной дозы препаратов — наиболее легкий путь подкрепить себя антиоксидантами.
Это особенно важно, если вы подвергаетесь воздействию факторов, способствующих образованию свободных радикалов: ультрафиолетовому излучению; воздуху, содержащему табачный дым, загрязненному выхлопными газами автомобилей и растворителями; воде, которую большинство из нас пьет из водопровода.
Мясо животных, которое мы покупаем в магазине и даже выращиваемых в домашних условиях, тоже содержит какое то количество солей тяжелых металлов, накапливаемых в мясе в течении жизни из употребляемой ими пищи.
Но, к сожалению, среди множества предлагаемых препаратов на рынке много подделок с низким качеством, не соответствующим международным требованиям, не прошедших хороших клинических испытаний.
Поэтому старайся не гнаться за дешевизной, что бы еще не усугубить существующего положения в организме, а брать качественные препараты, рекомендованные мной или консультантами нашей компании.
www.ronl.ru
Федеральное агентство по образованию
ФГАОУ ВПО «Уральский федеральный университет
имени первого Президента России Б.Н.Ельцина»
Курсовой проект
по курсу химии окружающей среды
Роль свободных радикалов в природной среде
Екатеринбург 2010
РЕФЕРАТ
СВОБОДНЫЕ РАДИКАЛЫ ОКИСЛЕНИЕ АТМОСФЕРА ПЕРОКСИД ГИДРОКСИЛ ОКСИДЫ АЗОТА ПРОБА КИСЛОТЫ ФЕРМЕНТЫ КАТАЛИЗАТОРЫ
В работе рассказывается об общем строении свободных радикалов, их свойствах и значение в природной среде. Отдельное внимание уделяется рассмотрению химических реакций с участием радикалов в атмосфере и живых организмах. Описываются процессы, происходящие при участии свободных радикалов в различных средах и их роль.
ОГЛАВЛЕНИЕ
ВВЕДЕНИЕ
1.ОБЩАЯ ХАРАКТЕРИСТИКА И НОМЕНКЛАТУРА СВОБОДНЫХ РАДИКАЛОВ
1.1Что такое свободные радикалы
1.2Номенклатура радикалов
1.3Классификация радикалов
1.3.1Первичные радикалы и реактивные молекулы
1.3.2Вторичные и третичные радикалы
2.РЕАКЦИИ С УЧАСТИЕМ СВОБОДНЫХ РАДИКАЛОВ В РЕЧНЫХ СИСТЕМАХ
2.1Наблюдения и эксперименты
2.2Результаты исследования
2.3Анализ результатов
3.СВОБОДНОРАДИКАЛЬНЫЕ РЕАКЦИИ В АТМОСФЕРЕ
3.1Реакции в стратосфере
3.2Химические процессы в тропосфере
3.2.1Роль углеводородов в тропосферных фотохимических процессах
4.СВОБОДНОРАДИКАЛЬНЫЕ РЕАКЦИИ В БИОЛОГИЧЕСКИХ СИСТЕМАХ
ЗАКЛЮЧЕНИЕ
СПИСОК ЛИТЕРАТУРЫ
ВВЕДЕНИЕ
Вся природа, окружающая нас, состоит из химических веществ и их соединений различной структуры. Их все можно разделить на различные группы по самым разнообразным признакам: состав, строение, роль в биологических системах и так далее. В своей работе я рассматриваю такую группу веществ, как свободные радикалы. Мой интерес вызван их широчайшей дифференциацией и повсеместным нахождением во всех компонентах биосферы.
Высокая реакционная способность свободных радикалов обуславливает большое количество химических реакций, которые происходят с различными элементами окружающей среды. Все они играют определенную роль в природных процессах.
В этой работе рассматриваются химические реакции с участием свободных радикалов и их участие в антропогенных и биологических циклах отдельные компонентов окружающей среды.
1. ОБЩАЯ ХАРАКТЕРИСТИКА И НОМЕНКЛАТУРА СВОБОДНЫХ РАДИКАЛОВ
1.1 Что такое свободные радикалы
Хорошо известно, что в органических молекулах (включая те, из которых состоит наш организм) электроны на внешней электронной оболочке располагаются парами: одна пара на каждой орбитали (рис. 1)
Свободные радикалы отличаются от обычных молекул тем, что у них на внешней электронной оболочке имеется неспаренный (одиночный) электрон (рис. 2 и 3).
Рисунок 1.1. Электронная структура молекулы метанол

Рисунок 1.2. Электронная структура радикала метанола
Неспаренный электрон в радикалах принято обозначать точкой. Например, радикал гидроксила обозначают как HO·, радикал перекиси водорода как HOO·, радикал супероксида как ·OO- или O2·-. Ниже даны формулы трех радикалов этилового спирта: Ch4Ch3O·; Ch4·CHOH; Ch4Ch3O·
Итак:
· Свободным радикалом называется частица — атом или молекула, имеющая на внешней оболочке один или несколько неспаренных электронов.
· Это делает радикалы химически активными, поскольку радикал стремится либо вернуть себе недостающий электрон, отняв его от окружающих молекул, либо избавиться от «лишнего» электрона, отдавая его другим молекулам.
· В особом положении оказалась молекула кислорода (диоксигена), которая содержит на внешней оболочке целых два неспаренных электрона. Таким образом, диоксиген — это бирадикал и, подобно другим радикалам, обладает высокой реакционной способностью.

Рисунок 1.3. Электронная структура некоторых молекул и радикалов
Важно подчеркнуть, что неспаренные электроны должны находиться на внешней оболочке атома или молекулы. В понятие свободного радикала не включаются ионы металлов переменной валентности, неспаренные электроны в которых находятся на внутренних оболочках. Поскольку и радикалы и ионы таких металлов как железо, медь или марганец (так же как комплексы этих металлов) дают сигналы электронного парамагнитного резонанса (ЭПР), эти частицы в совокаупности часто называют парамагнитными центрами.
Образование радикалов из устойчивых молекул обусловлено, таким образом, появлением на свободной, валентной орбитали нового электрона или наоборот — удалением одного электрона из электронной пары. Эти процессы обычно происходят в результате реакций одноэлектронного окисления или восстановления. В таких реакциях обычно участвует, наряду с молекулой, из которой радиукал образуется, ион металла переменной валентности, который ка раз и служит донором или акцептором одного электрона (а не двух сразу, как это бывает в реакиях между двумя органическими молекулами или между органической молекулой и кислородом). Типичный пример реакции, в которой образуется радикал — это реакция Фентон: взаимодействие пероксида водорода с ионом двухвалентного железа:
Fe2+ + h3O2 => Fe3+ + OH- + ·OH (радикал гидроксила)
При высоких температурах или под действием ультрафиолетового излучения радикалы могут образовываться также в результате разрыва химической связи (гомолитическое расщепление). В обычных условиях такие реакции в нормальных живых клетках практически не имеют места.
1.2 Номенклатура радикалов
Относительно недавно Комиссия по Номенклатуре Неорганической химии сформулировала основные правила номенклатуры радикалов [1] (табл. 1).
Таблица 1. Названия некоторых радикалов и молекул согласно рекомендациям Комиссии по Номенклатуре Неорганической Химии (1990).
| Формула | Структурная формула | Название радикала |
| O·- | ·O- | Оксид (1-), [оксид] |
| О2 | ·ОО· | [Диоксиген] |
| О2·- | ·ОО- | Диоксид (1-), супероксид, [диоксид] |
| O3 |  | Триоксиген, [озон] |
| °O3·- | ·OOO- | Триоксид (1-), озонид |
| HO· | HO· или ·OH | [Гидроксил] |
| HO2· | HOO· | Гидродиоксид, [гидродиоксил] |
| Н2 02 | HOOH | [Перекись водорода] |
| RO· | RO· | [Алкоксил] |
| C2 H5 O· | Ch4 Ch3 O· | [Этоксил] |
| R02· | ROO· | [Алкилдиоксил] |
| RO2 H | ROOH | [Апкилгидропероксид] |
Остановимся на некоторых из этих рекомендаций. Прежде всего нет необходимости писать «свободный» перед словом радикал [533]. О радикальной природе рассматриваемой частицы говорит окончание «ил». Так радикалы RO· и НО· имеют наименование, соответственно «алкоксил» и «гидроксил».
Существенно новым можно считать рекомендацию не злоупотреблять производными от «пероксид» и «гидропероксид». Группа из двух связанных между собой атомов кислорода называется «диоксид». В соответствии с этим радикал ROO· рекомендуется называть «алкилдиоксилом» {Koppenol, 1990 #7}. Допускается сохранение и альтернативного названия «алкилпероксил», но это хуже {Koppenol, 1990 #7}. Молекулярный кислород называется «диоксигеном», а озон — «триоксигеном».
Наименование с окончанием «ил» весьма удобно, но ничего не горит о том, каков заряд частицы. Поэтому в необходимых случаях рекомендуется использовать систематическое название радикала, где после названия группы дается в скобках заряд. Например радикал O2·- имеет наименование «диоксид (1-)». В этой работе мы будем использовать более краткое название «диоксид».При написании формул радикалов в суперскрипте сначала ставится точка, указывающая на наличие неспаренного электрона у данного атома, а затем знак заряда иона. Например «O2·- ». В структурных формулах точка должна стоять именно у того атома, где локализован неспаренный электрон. Например, чтобы подчеркнуть, что диоксиген имеет два неспаренных электрона, можно написать его формулу таким образом «О2 ». В таблице 1 приведен список рекомендуемых названий радикалов; в квадратных скобках даны названия, которые будут преимущественно использованы в данной книге.
1.3 Классификация радикалов
1.3.1 Первичные радикалы и реактивные молекулы
Все радикалы, образующиеся в человеческом организме, можно разделить на природные и чужеродные. В свою очередь природные радикалы можно разделить на первичные, вторичные и третичные (См. схему на рис. 4).

Рисунок 1.4. Классификация свободных радикалов, образующихся в нашем организме
Первичными можно назвать радикалы, образование которых осуществляется при участии определенных ферментных систем. Прежде всего к ним относятся радикалы (семихиноны), образующиеся в реакциях таких переносчиков электронов, как коэнзим Q (обозначим радикал как Q·) и флавопротеины. Два других радикала — супероксид (·OO- ) и монооксид азота (·NO) также выполняют полезные для организма функции, которые будут подробнее рассмотрены в соответствующих разделах.
Из первичного радикала — супероксида, а также в результате других реакций, в организме образуются весьма активные молекулярные соединения: перекись водорода, гипохлорит и гидроперекиси липидов (см. рис. 5). Такие молекулы, наряду с радикалами, получили в англоязычной литературе название «reactive species», что в русской литературе чаще всего переводится как «активные формы». Чтобы провести водораздел между радикалами и молекулярными продуктами, мы предлагаем называть последние «реактивными молекулами». Таким образом, предлагается такая терминология:
Активные формы = свободные радикалы + реактивные молекулы
Halliwell предлагает термины активные формы кислорода, азота и хлора {Halliwell, 1998 #9}. Как видно из схемы на рис. 5, к активным формам кислорода относятся супероксид, радикал гидроксила, перекись водорода и синглетный кислород. Окись азота и результат ее взаимодействия с супероксидом — пероксинитрит предлагается называть активными формами азота. Активной формой хлора можно назвать гипохлорит, образуемый в реакции перекиси водорода с ионом хлорида, которую катализирует фермент миелопероксидаза.

Рисунок 1.5. Активные формы (Reactive species) кислорода, азота и хлора
В складывающейся в настоящее время терминологии нужно найти место радикалам и гидроперекисям полиненасыщенных жирных кислот, которые образуются в очень важной реакции цепного окисления липидов. С химической точки зрения — это неоднородная группа. При отрыве атома водорода от молекулы полиненасыщенной жирной кислоты образуется алкоксильный радикал, в котором неспаренный электрон локализован у углеродного атома. Это как бы «активная форма углерода». Но при дальнейшем взаимодействии алкильного радикала с диоксигеном (молекулярным кислородом) образуется диоксид-радикал с локализацией неспаренного электрона на атоме кислорода. По структуре, и отчасти по свойствам, такой радикал напоминает супероксид, и его можно отнести к активным формам кислорода, что и делают некоторые авторы. Образующиеся при перекисном окислении липидов гидроперекиси ненасыщенных жирных кислот также можно отнести к этой категории активных форм, по аналогии с пероксидом водорода. В эту же категорию попадают тогда и алкоксильные радикалы липидов, образующиеся при одноэлектронном восстановлении гидроперекисей, например, ионами Fe2+; по сути, это гомологи гидроксильного радикала.
Несмотря на все сказанное, мы предлагаем объединить все перечисленные продукты (и реагенты) цепного окисления липидов одним термином: активные формы липидов. Для биолога и врача все же важнее, не у какого именно атома локализован непарный электрон, а какая молекула при этом становится химически агрессивной, т. е. приобретает черты свободного радикала или его реактивного предшественника. Итак, к активным формам липидов мы отнесем алкильные, алкоксильные и диоксид-радикалы. а также гидроперекиси полиненасыщенных жирных кислот и соответствующих цепей фосфолипидов, триглицеридов или холестерина (см.рис. 5).
1.3.2 Вторичные и третичные радикалы
Реактивные молекулы: перекись водорода, гидроперекиси липидов, пероксинитрит, — образуются в реакциях, одним из участников которых в большинстве случаев является радикал, а иногда — диоксиген, который, впрочем, тоже имеет неспаренные электроны на внешней электронной оболочке. В свою очередь, эти молекулы, а наряду с ними — гипохлорит, охотно образуют радикалы в присутствии ионов металлов переменной валентности, в первую очередь — ионов двухвалентного железа. Такие радикалы мы будем называть вторичными; сюда относятся радикал гидроксила и радикалы липидов. Вторичные радикалы, в отличие от первичных, образуются в неферментативных реакциях и, насколько известно в настоящее время, не выполняют физиологически-полезных функций. Напротив, они обладают разрушительным действием на клеточные структуры и с полным основанием могут быть названы вредными радикалами. Именно образование вторичных радикалов (а не радикалов вообще) приводит к развитию патологических состояний и лежит в основе канцерогенеза, атеросклероза, хронических воспалений и нервных дегенеративных болезней. Впрочем, реактивные молекулы также обладают цитотоксическим действием, причем не только благодаря образованию из них свободных радикалов, но и непосредственно, как это доказано для пероксинитрита и гипохлорита, а в некоторых ситуациях — и для перекиси водорода.
Для защиты от повреждающего действия вторичных радикалов в организме используется большая группа веществ, называемых антиоксидантами, к числу которых принадлежат ловушки, или перехватчики свободных радикалов. Примером последних служат альфа-токоферол, тироксин, восстановленный убихинон (Qh3 ) и женские стероидные гормоны. Реагируя с липидными радикалами, эти вещества сами превращаются в радикалы антиоксидантов, которые можно рассматривать как третичные радикалы (см. рис. 1.3).
Наряду с этими радикалами, постоянно образующимися в том или ином количестве в клетках и тканях нашего организма, разрушительное действие могут оказывать радикалы, появляющиеся при таких воздействиях, как ионизирующее излучение, ультрафиолетовое облучение или даже освещение интенсивным видимым светом, например, светом лазера. Такие радикалы можно назвать чужеродными. К ним принадлежат также радикалы, образующиеся из попавших в организм посторонних соединений, ксенобиотиков, многие из которых оказывают токсическое действие именно благодаря свободным радикалам, образующимся при метаболизме этих соединений (рис. 1.3).
Реакции с участием свободных радикалов, в особенности реакции цепного окисления, отличаются большой сложностью и протекают через ряд последовательных стадий. В изучении механизма цепных реакций основную роль сыграло исследование кинетики процессов; при этом измерение кинетики хемилюминесценции позволяет непосредственно видеть изменение во времени концентрации радикалов, например радикалов липидов. Параллельное измерение хемилюминесценции, окисления ионов двухвалентного железа и накопления продуктов реакции в суспензиях митохондрий и фосфолипидных везикул (липосом) позволило экспериментально определить константы скоростей основных реакций свободнорадикального цепного окисления липидов. [2]
2. РЕАКЦИИ С УЧАСТИЕМ СВОБОДНЫХ РАДИКАЛОВ В РЕЧНЫХ СИСТЕМАХ
Одно из областей деятельности человека каким-либо образом связана с реками – оросительные системы, строительство водохранилищ или же постройка электростанций. Все это так или иначе приводит к изменению гидрологического режима рек, что не происходит бесследно для природы и приводит, как правило, к замедлению скорости потока воды и уменьшению аэрированности. Это не может не сказываться на способности речной воды к самоочищению. В этой связи следует упомянуть изменение условий протекания фотохимических реакций окисления водных органических веществ, коагуляции и седиментации, а также создание благоприятной среды для развития микроорганизмов и накопления продуктов их жизнедеятельности.
В этом разделе работы приведены эксперименты и наблюдения, в ходе которых были созданы условия, практически исключавшие для речной воды возможность обмена веществом с внешней средой (кроме незначительного газообмена с атмосферой), что позволило выявить действие отдельных факторов самоочищения.
2.1 Наблюдения и эксперименты
Основные этапы круговорота биогенных элементов в водной среде — разложение и синтез органического вещества сопровождаются сдвигом углекислотного равновесия, что приводит к изменению рН среды. Поэтому обобщенным показателем динамики неравновесных процессов в воде в проведенных исследованиях выбрано изменение значений водородного показателя. Измерения рН осуществлялись при помощи ионометра И-130М при комнатной температуре. Основные опыты были проведены на воде из р.Волхов, а также на модельных растворах (200 мг/л  в дистиллированной воде).
в дистиллированной воде).
2.2 Результаты исследования
В ходе проводившихся экспериментов установлено, что в любой отдельно взятой пробе природной воды наблюдается естественное увеличение водородного показателя во времени. Эта тенденция проявляется независимо от того, была ли проба оставлена на свету или в темноте. В случае предварительного кипячения пробы в течение 20 минут значительное возрастание рН начинается примерно через три недели после начала эксперимента, а если при этом дополнительно вводился обеззараживающий агент (перекись водорода в количестве 500 мг/л), то после начального подъема рН оставался стабильным в течение всего времени наблюдений (см. рис. 2.1). Уменьшение рН на кривой 2 в первые дни проведения опыта объясняется постепенным растворением углекислоты, удаленной в результате кипячения, а начальное увеличение рН на кривой 3 будет подробнее рассмотрено ниже.
Величина подщелачивания практически не зависит от седиментации дисперсных частиц — повышение рН происходило равномерно по высоте сосуда, незначительные отклонения наблюдались лишь у самого дна. Параллельно подщелачиванию в опытах без добавления  зафиксировано также некоторое возрастание оптической плотности раствора (1-2%). Возможное объяснение: возрастание доли ионизированных органических молекул. После стабилизации рН появлялась тенденция к снижению оптической плотности раствора, возможно вследствие окисления двойных связей растворенных фульвокислот. Во время проведения каждого опыта с речной водой выпадал бурый осадок гуминовых соединений, даже в случае предварительного фильтрования пробы через плотную фильтровальную бумагу для тонкодисперсных примесей.
зафиксировано также некоторое возрастание оптической плотности раствора (1-2%). Возможное объяснение: возрастание доли ионизированных органических молекул. После стабилизации рН появлялась тенденция к снижению оптической плотности раствора, возможно вследствие окисления двойных связей растворенных фульвокислот. Во время проведения каждого опыта с речной водой выпадал бурый осадок гуминовых соединений, даже в случае предварительного фильтрования пробы через плотную фильтровальную бумагу для тонкодисперсных примесей.

Рис. 2.1. Изменение рН в пробах речной воды: 1 — речная вода без воздействий, 2 — кипяченая речная вода, 3 — кипяченая речная вода с добавлением 

Рис. 2.2. Влияние добавок  на изменение рН в водных системах: 4 — речная вода на свету, 5 — речная вода в темноте, 6 — модельный раствор без добавления
на изменение рН в водных системах: 4 — речная вода на свету, 5 — речная вода в темноте, 6 — модельный раствор без добавления 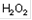 , 7 — модельный раствор с добавлением
, 7 — модельный раствор с добавлением 
На повышение рН оказывает некоторое влияние выщелачивание стеклянной посуды, но контрольные опыты с дистиллированной кипяченой водой в закрытом сосуде показали, что изменение рН в этом случае начинает фиксироваться ионометром лишь через 20-25 дней после начала опыта и по абсолютной величине соответствует изменению концентрации ионов водорода  моль/л. За такой же период времени концентрация ионов водорода изменилась в пробе речной воды примерно на
моль/л. За такой же период времени концентрация ионов водорода изменилась в пробе речной воды примерно на 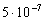 моль/л, а в ряде случаев в системах с добавлением перекиси водорода произошли изменения порядка
моль/л, а в ряде случаев в системах с добавлением перекиси водорода произошли изменения порядка  моль/л.
моль/л.
В случае добавления перекиси водорода эффект возрастания рН проявлялся значительно быстрее, причем отмечено увеличение рН после добавления перекиси водорода в пробы со стабилизировавшимся водородным показателем. В качестве обеззараживающей добавки перекись водорода выбрана не случайно. Этот окислитель в незначительных количествах постоянно присутствует в речной воде и активно участвует в процессах самоочищения. Существует несколько путей поступления  в природные воды. Перекись водорода является одним из промежуточных продуктов метаболизма водных микроводорослей. Фотохимические реакции получения
в природные воды. Перекись водорода является одним из промежуточных продуктов метаболизма водных микроводорослей. Фотохимические реакции получения  и гидроксильных радикалов будут рассмотрены ниже. В ходе опытов вводились высокие концентрации
и гидроксильных радикалов будут рассмотрены ниже. В ходе опытов вводились высокие концентрации  — порядка 500 мг/л — в связи с тем, что перекись водорода активно участвовала в реакциях окисления водной органики (расход составлял примерно 23-28% за неделю наблюдений), а стерильность раствора необходимо было обеспечить в течение всего опыта.
— порядка 500 мг/л — в связи с тем, что перекись водорода активно участвовала в реакциях окисления водной органики (расход составлял примерно 23-28% за неделю наблюдений), а стерильность раствора необходимо было обеспечить в течение всего опыта.
Были обнаружены значительные расхождения в действии  , на свету и в темноте (см. рис. 2.2). Как известно, световое излучение ускоряет разложение перекисных соединений. Особенности изменения рН на кривой 5 можно объяснить особенностями гетерогенного окисления перекисью водорода коллоидной органики.
, на свету и в темноте (см. рис. 2.2). Как известно, световое излучение ускоряет разложение перекисных соединений. Особенности изменения рН на кривой 5 можно объяснить особенностями гетерогенного окисления перекисью водорода коллоидной органики.
2.3 Анализ результатов
Предположительно, источником подщелачивания являлись окислительно-восстановительные реакции с участием перекиси водорода. Разложение перекиси водорода в водных растворах происходит непрерывно под влиянием разнообразных физико-химических воздействий: тепла, света, кавитации и т.п. Восстановление  может происходить либо через диссоциацию с образованием гидроксил-иона и гидроксильного радикала, либо через протонирование и распад на воду и гидроксильный радикал.
может происходить либо через диссоциацию с образованием гидроксил-иона и гидроксильного радикала, либо через протонирование и распад на воду и гидроксильный радикал.
Известно, что гидроксильный радикал является самым реакционноспособным окислителем в природной среде и способен реагировать практически со всеми растворенными в воде органическими и неорганическими веществами. Основные пути превращения гидроксильных радикалов в водных растворах — это отрыв водорода от органических соединений с образованием воды и органического радикала, гидроксилирование ненасыщенных органических соединений и реакции одноэлектронного переноса с анионами. В результате подобных радикальных перегруппировок образуются более стабильные и долгоживущие радикалы. Рекомбинация углеродсодержащих радикалов приводит в конечном итоге к появлению органических соединений, и это служило причиной образования осадка в проводившихся опытах. Образование подобного осадка рыжевато-коричневого цвета имело место как в чистых растворах гидрокарбонатов, так и в водопроводной воде при длительном отстаивании.
В речной воде образующийся осадок, как правило, более темный, что связано с включением в процессы радикальных превращений фрагментов гумусовых соединений, присутствующих в значительных количествах в природных водах.
Если в водных растворах преобладают карбонаты и гидрокарбонаты, то восстановление гидроксильных радикалов наиболее вероятно по следующей схеме:

Лимитирующим фактором реакции (1) является активность гидроксильных радикалов, концентрация которых в природных водах составляет  моль/л. В случае добавления перекиси водорода концентрация гидроксил-ионов должна увеличиться, и эффект подщелачивания в результате реакции (1) станет более заметным. Факторы, ускоряющие процесс разложения
моль/л. В случае добавления перекиси водорода концентрация гидроксил-ионов должна увеличиться, и эффект подщелачивания в результате реакции (1) станет более заметным. Факторы, ускоряющие процесс разложения  , в том числе солнечный свет, должны усиливать подщелачивание. Именно такое поведение растворов и наблюдалось в проводившихся опытах (см. кривые 1, 4, 5).
, в том числе солнечный свет, должны усиливать подщелачивание. Именно такое поведение растворов и наблюдалось в проводившихся опытах (см. кривые 1, 4, 5).
В качестве восстановителей и одновременно катализаторов процесса могут выступать металлы переменной валентности, в том числе железо (общая концентрация ионов железа в речной воде составляет 0,2-1,2 мг/л). Полная схема катализа ионами железа распада перекиси водорода представлена ниже. Логарифмы скоростей реакций приведены для температуры 298 К.
свободный радикал стратосфера тропосфера

В растворе без доступа света преобладают наиболее быстрые реакции (2) и (3), что и является причиной подщелачивания среды (см. кривую 5). Под действием света происходит фотодиссоциация перекиси водорода, концентрация гидроксильных и гидроперекисных радикалов возрастает — и начинает заметно сказываться влияние реакций (4)-(6), что существенно ускоряет ход процесса (см. кривую 4). Реакции (7), (8), способствующие подкислению среды, по-видимому, являются второстепенными, хотя с повышением рН их роль может усиливаться.
Окислительно-восстановительные системы подобной структуры помимо переходных металлов образуют и органические соединения, в частности гидрохинон. Установлено, что гидрохинон является экзометаболитом некоторых видов бактерий, так что его присутствие в речной воде и участие в процессах каталитического разложения  вполне закономерно.
вполне закономерно.
Генерация свободных радикалов в природных водах может происходить и без участия перекиси водорода. Причем в роли инициаторов могут активно выступать органические соединения, характерные для речной воды. Такие процессы происходят в гумусовых веществах в результате фотовозбуждения карбонильных групп до так называемых n- p-триплетных состояний. Возможен и вариант фотоэлектронного переноса, когда супероксид-ион образуется непосредственно из растворенного в воде кислорода. Подобные реакции часто осуществляются в молекулах красителей и фульвокислот.
Реальна также и непосредственная активация молекулярного кислорода ионами металлов переменной валентности, возбужденных солнечным светом. Образующиеся металл-кислородные комплексы распадаются с выделением либо гидропероксид-иона, либо супероксид-иона.
Металлы в природных водах присутствуют либо в форме комплексов с водными органическими соединениями, либо в коллоидной форме гидроокисей. Перенос электрона при взаимодействиях ионов металлов с  осуществляется через образование промежуточных металл-перекисных комплексов. Такие комплексы образуются при наличии одного или двух вакантных мест в координационной сфере иона металла. Соответствующие изменения геометрической структуры координационной сферы или числа лигандов могут происходить без больших затрат энергии извне даже для самых прочных координационных связей. Каталитическая активность проявляется, таким образом, для ионов металлов и в комплексной, и в коллоидной форме. Вторичные реакции образующегося супероксид-иона могут приводить как к образованию гидроксил-иона, так и к выделению кислорода. Предполагается, что они и являются причиной подщелачивания в изолированной речной воде (см. кривую 1 на рис.2.1.).
осуществляется через образование промежуточных металл-перекисных комплексов. Такие комплексы образуются при наличии одного или двух вакантных мест в координационной сфере иона металла. Соответствующие изменения геометрической структуры координационной сферы или числа лигандов могут происходить без больших затрат энергии извне даже для самых прочных координационных связей. Каталитическая активность проявляется, таким образом, для ионов металлов и в комплексной, и в коллоидной форме. Вторичные реакции образующегося супероксид-иона могут приводить как к образованию гидроксил-иона, так и к выделению кислорода. Предполагается, что они и являются причиной подщелачивания в изолированной речной воде (см. кривую 1 на рис.2.1.).
Деятельность микроорганизмов в водной среде не приводит к значительным разрушениям водных органических соединений. В противном случае это сопровождалось бы понижением рН в речной воде. По этой же причине биохимические превращения гидрокарбонат-ионов незначительны. Эффект увеличения рН в модельных растворах легко можно было бы объяснить выделением части углекислоты в атмосферу в ходе установления равновесия (при рН < 8,35 в водных растворах всегда содержится определенное количество свободных молекул  ):
):

Именно такое объяснение наиболее приемлемо для чистых растворов гидрокарбонатов (см. кривую 6). Однако сходство поведения речной воды и гидрокарбонатных модельных растворов с добавлением  свидетельствует об участии растворенной углекислоты в окислительно-восстановительных процессах (см. кривую 7). Удаление из раствора углекислоты (через кипячение) при наличии высокой активности гидроксорадикалов приводит к быстрой стабилизации рН (см. кривую 3). Отсюда можно предположить, что гидрокарбонат-ионы играют роль своеобразного аккумулятора свободных радикалов, существенно замедляя дальнейшие радикальные реакции. Конкретный механизм участия гидрокарбонатов в окислительно-восстановительных реакциях требует проверки в ходе дальнейших исследований. [3]
свидетельствует об участии растворенной углекислоты в окислительно-восстановительных процессах (см. кривую 7). Удаление из раствора углекислоты (через кипячение) при наличии высокой активности гидроксорадикалов приводит к быстрой стабилизации рН (см. кривую 3). Отсюда можно предположить, что гидрокарбонат-ионы играют роль своеобразного аккумулятора свободных радикалов, существенно замедляя дальнейшие радикальные реакции. Конкретный механизм участия гидрокарбонатов в окислительно-восстановительных реакциях требует проверки в ходе дальнейших исследований. [3]
3. СВОБОДНО-РАДИКАЛЬНЫЕ РЕАКЦИИ В АТМОСФЕРЕ
Атмосфера — газовая оболочка Земли. Ее масса составляет около 5,9·1015 тонн. Во многих отношения она напоминает слой жидкой воды, покрывающей в виде морей и океанов три четверти земной поверхности. Знания об атмосфере необходимо рассматривать воедино с позиций и географии, и физики, и химии. Атмосфера имеет слоистое строение и состоит из нескольких сфер, между которыми располагаются переходные слои — «паузы». В сферах изменяется количество воздуха и его температура. Области минимума и максимума температур — «паузы», а промежуточные области — сферы. Так, тропопауза отделяет тропосферу от стратосферы; стратопауза — стратосферу от мезосферы и т.д. Атмосфера представляет собой чрезвычайно сложную систему. Ее пронизывает излучение Солнца и испускаемые им частицы высокой энергии, а также космическое излучение. Этот поток энергии оказывает заметное химическое воздействие на атмосферу. Кроме того, под воздействием земного притяжения более тяжелые атомы и молекулы опускаются в нижнюю часть атмосферы, а в верхней ее части остаются более легкие. В результате состав атмосферы оказывается непостоянным. [4]
3.1 Реакции в стратосфере
Каталитические циклы разрушения стратосферного озона
Наиболее важными из катализаторов разрушение озона:
· Гидроксильный радикал ОН – наиболее распространенная частица в атмосфере, вызывающая множество разнообразных реакций
· Монооксид азота
· Атомарный хлор
Разрушение озона при участии этих катализаторов происходит следующим образом:
1) Гидроксильный радикал

Цикл может разрушиться в результате гибели (рекомбинации) радикалов:

Но до гибели ОН упевает разрушить до 100 молекул О3
2) Монооксид азота

3) Атомарный хлор

Каждый из катализаторов [в цикле ] регенерируется и вновь вступает в реакции, т.е. развивается циклический процесс, когда 1 частица разрушает большое число молекул О3 ОН, NO, Cl в атмосфере могут появится в результате естественных процессов и в результате антропогенных загрязнений.
Гидроксильный радикал. Их источником являются в основном природные процессы. ОН-радикал
а) является продуктом фотодиссоциации воды

Но этот процесс происходит только в верхних слоях атмосферы, т.к. требуется коротковолновое излучение
б) образуется (как отмечалось) при взаимодействии атомарного O находящегося в возбужденном О(1 d), который выделяется при фотолизе О2 в верхних слоях стратосферы при фотолизе О3 :
 это гораздо более значимый процесс
это гораздо более значимый процесс
в) к образованию ОН приводит еще ряд стратосферных процессов, вносящих несколько меньший вклад:

Оксид азота NO
Источники NO в стратосфере преимущественно природные. Главным источником NO считается проникающий в атмосферу оксид N2 O. Закись азота образуется на уровне Земли. Она выделяется в атмосферу:
· В результате микробиологической активности в почвах
· При горении биомассы
· При сжигании ископаемого и моторного топлива (в результате высокотемпературного окисления N2 воздуха)
N2 O чрезвычайно химически инертен, мало растворим в воде. Таким образом он равномерно распределяется во всей толще тропосферы и имеет продолжительность жизни в тропосфере ~100 лет… Благодаря инертности, устойчивости он легко проникает (переносится)в стратосферу где:
а) Может претерпевать фотодиссоциацию

б) Или взаимодействует с ат O(1 d):

— этот процесс основной, приводящий к образованию NO стратосфере.
Кроме того в качестве источника NO рассматривается звуковая высотная авиация – NO содержится в отработавших газах авиационных турбин
В принципе роль NO в химии стратосферного озона – двойная. В нижней части стратосферы (близи термопаузы) NO включается в реакции, аналогичные тем, что протекают у земной поверхности, которые сопровождаются образованием озона.
Но на высоте около 20 км NO включается в цикл разрушения озона.
Атомарный хлор
Главный источник атомарного хлора – антропогенный. Атомарных хлор образуется в результате фотохимического разрушения фреонов (фторхлорметанов или ФХУ), применяющихся в качестве хладоагентов (их др.название хладоны) в холодильных установках, кондиционерах и в качестве пропеллентов (средств для образования аэрозолей) в аэрозольных баллончиках, пенных огнетушителях.
Наиболее широко распространенными фреонами является CF2 Cl2 (фреон 12) и CFCl3 (фреон 11). (применительно к производству фреон 12 составляет 85% всей мировой продукции F-содержащих соединений)
В молекулы некоторых фреонов так же включен Br наряду с ат. Cl или вместо него – эти вещества называются галлоны (вместо фреонов), например CF2 ClBr – гало 1211, CF3 Br – галлон 1301
Фреоны химически инертны и очень устойчивы с нижних слоях атмосферы время их жизни в тропосферы ~ 80 лет, таким образом постепенно они (как и N2 O) м.б. перенесены в стратосферу.
В стратосфере ( в средних слоях) ХФУ фотохимических разлатапется под действием света с l<214 нм.
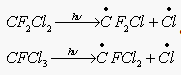
Наряду с фреонами (антропогенного происхождения) в стратосферу спосолен проникать и давать ат Cl природных Cl-содержащий компонент Ch4 Cl – он выделяется в больших количествах из морской воды, при вулканической дятельности, при горении биомассы ( в результате рекомбинации  )
)
Потенциальная роль отдельных частиц, участвующих разрушении озона, видна из сравнения значений констант скоростей их реакций с озоном (при одной температуре)
| K, см3 /молекул×с | |
 | 8×10-12 |
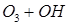 | 6,7×10-14 |
 | 2,5×10-15 |
 | 5×10-16 |
Таким образом наиболее активен в этом процессе ат Cl – его константа скорости на 2-3 порядка выше константы скорости других катализаторов.
Систематические наблюдения, проводимые с середины 70-х годов, указывают на существенное изменение в озоновом слое – снижение концентрации озорна. Учитывая отмеченные факты в качестве гл причины рассматривают поступление в стратосферу фреонов.
Образующиеся при фотолизе фреонов атом Cl имеет «сток» из стратосферы (процесс удаления) по реакции с Ch5 :

Исследования отмечают увеличение концентрации HCl в стратосфере, что свидетельствует в пользу ведущей роли фреонов в разрушении озона. Реакция Cl с Ch5 разрушает «хлорный» цикл, поскольку HCl неактивен по отношению к озону, но он может быть источником активной формы по реакции с гидроксилом:

Поэтому HCl относят к так называемым «резервуарным» газам. К образованию «резервуарных» газов приводят и реакции ClO – промежуточная компонента хлорного цикла:

Их фотохимическое разложение генерирует ClO и атомарный хлор – активные к озону частицы.

Сходным образом с фреонами ведут себя галлоны – Cl, F, Br, углероды. Они столь же инертны, имеют время жизни в тропосфере примерно 70 лет и могут проникать в стратосферу. Под действием УФ-излучения они разлагаются с выделением Br.

Эти реакции протекают уже в нижней части стратосферы вблизи тропопаузы, таким образом роль атома Br особенно велика в этом слое. Br действует аналогично Cl, разрушая озон в «бромном» цикле, но существенно активнее Cl – один атом Br может разрушить до 100 тыс. молекул озона. Это объясняется малыми скоростями реакции атома Br и BrO, приводящих к неактивным «резервуарным» газам.
Обобщая, можно сделать вывод, что разрушение стратосферного озона происходит с участием различных частиц, предшественниками которых являются и природные компоненты и антропогенные загрязнители. При этом отдельные циклы действуют не изолированно, а испытывают взаимное влияние, например, дезактивация ClO по реакции с NO связывает азотные и хлорные циклы.
Тем не менее, главную опасность, по общему мнению, несут ХФУ. Учитывая это, во многих странах законодательно введены ограничения на использование фторхлорметанов. Это же стимулировало работы по замене ХФУ на вещества с близкими потребительскими свойствами, не представляющими опасность для озона.
В качестве таких заменителей предложены F-Cl углеводороды CHFCl2 (фреон 21), CHF2 Cl (фреон 22) и другие. Их отличие от ФХУ заключается в возможности разрушения в тропосфере (тропосферный сток) по реакции с ОН (на примере фреона 22):

Существование такого стока значительно уменьшает их время жизни в тропосфере и делает их менее опасными для стратосферы. Вместе с тем, тропосферный сток фреонов-заменителей зависит от концентрации ОН, который участвует в реакциях с у/в и с СО. Поэтому увеличение поступления у/в и СО в атмосферу в результате антропогенной деятельности может привести снижению скорости разложения фреонов.
Еще одна проблема разложения F, Cl-углеводородов — образование токсичных веществ в ходе превращения радикалов, образующихся по реакции фреонов с ОН, например:
а) при взаимодействии с молекулярным кислородом
 — образование пероксидного радикала
— образование пероксидного радикала
б) его взаимодействие с NO
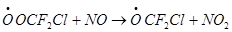
в) образование дифторфосгена – чрезвычайно токсичный и опасный для всех живых организмов, поэтому замена ХФУ F,Cl углеводороды в общем-то не устраняет совсем угрозы озонового слоя и порождает новую экологическую проблему.
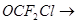 О=СF2
О=СF2

3.2 Химические процессы в тропосфере
Тропосферы достигает УФ излучение достаточно низкой энергии с λ>300 нм, поскольку более коротковолновые практически полностью поглощаются в более высших слоях в процессах фотодиссоциации О2 и О3 .
УФ-излучение низкой энергии не вызывает фотохимических реакций основных компонентов, т.е. О2 и N2. Реакции с участием основных компонентов могут протекать например при газовых разрядах


И затем доокисляя

Но в фотохимических реакциях в тропосфере участвуют ряд примесей, средняя концентрация которых в атмосфере мала, но локальная может быть значительной в результате активной хозяйственной деятельности – это прежде всего NOx, у/в, озон.
В тропосфере образуется и накапливается сильный окислитель озон, но по механизму, отличающемуся от стратосферного, где к образованию озона приводит атом О(3 р), образующийся при фотодиссоциации О2 .
Озон в тропосфере образуется в фотолитическом цикле диоксида азота.
NO2 поступает в атмосферу в значительных количествах от стационарных и передвижных источников, сжигающих органическое топливо непосредственно при сжигании образуется NOx, который постепенно доокисляется до NO2 [ ]
]
NO2 легко диссоциирует под действием УФ-излучения с λ<380 нм
(1)
– это одна из наиболее важных фотохиических реакций в тропосфере, приводящая к образованию активного атома О
Далее по известной реакции образуется озон:
(2) 
Затем озон окисляет NO в NO2 и цикл замыкается
(3) 
NO2 может выводится из цикла по разным реакциям, например, окисляясь в азотную кислоту в капельной фазе атмосферной влаги

Либо гидролизуясь в газовой фазе:
 — это реакция равновесная
— это реакция равновесная
Три реакции (1-3) фотолитического цикла NO2 протекают очень быстро и их комбинация должна определять некоторый постоянный уровень концентрации озона в нижних слоях атмосферы.
Однако измерения показывают, что фактическая концентрация О3 в атмосфере городов могут примерно на порядок превышать те, что следуют из фотолитичского цикла NO2, т.е. очевидно есть еще какой то альтернативный механизм окисления NO в NO2 .
3.2.1 Роль углеводородов в тропосферных фотохимических процессах
В атмосферу поступают разнообразные по строению и молекулярной массе углеводороды. Прежде всего это СН4, выделяющийся в естественных процессах (микробиологическая активность в почвах, и антропогенного происхождения. С продуктами сгорания топлив в ДВС, стационарных установках в атмосферу выбрасывается большой набор разных по строению веществ – алканы, алкены, ароматические углеводороды.
У/в в атмосфере окисляются активными компонентами – атомарным О, О3 и гидроксильным радикалом, который играет исключительно важную роль в химических превращениях загрязняющих веществ в тропосфере.
Окисление у/в протекает по радикальному механизму через образование на одной из стадий пероксидного радикала 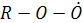 способного окислять NO:
способного окислять NO:
 (алкоксильный радикал)
(алкоксильный радикал)
Эта реакция ускоряет образование NO2 и включение его в фотолитический цикл. При этом скорость данной реакции значительно больше, чем скорость реакции, в которой расходуется окислитель озон ( )
)
Это и приводит к накоплению озона.
Окислители у/в – атомарный О и О3 образуются в рассмотренном фотолитическом цикле NO2. Атомарный О в основном участвует в образовании озона, но частично может расходоваться на реакции с у/в
Гидроксильный радикал образуется:
1) главным образом по реакции с Н2 О атомарного О(1 Д), выделяющегося при фотолизе озона (в основном эта реакция идет в верхних слоях тропосферы, куда проникает излучение с λ<300 нм)

2) дополнительное количество ОН радикала в тропосфере дает реакция О3 с НО2, который образуется по нескольким реакциям (о них ниже)

3) и кроме того ОН радикалы, хотя и начинают процесс окисления у/в, т.е. в начале расходуются, но в этих же процессах и накапливаются в условиях загрязненной антропогенными выбросами атмосферы
ОН-радикал наиболее важный окислитель в тропосфере, он начинает большинство многостадийных процессов окисления у/в и других примесей.
Окисление метана и его гомологов
Окисление метана инициируется ОН-радикалом. В последующем в процесс включается молекулярный О2. Сопряжено с окислением СН4 идет окисление NO (т.е. NO включается в цепочку реакций на одной из стадий)
Начальная стадия

Взаимодействие алкильного (метильного) радикала с О2 дает пероксильный радикал:
 ,
,
который как отмечалось определяет альтернативный механизм окисления NO в NO2 (вместо окислителя О3 )
Т.е.

Взаимодействие радикала  с О2 приводит к образованию формальдегида и гидропероксидного радикала
с О2 приводит к образованию формальдегида и гидропероксидного радикала

Образующийся NO2 включается в фотолитический цикл


Что приводит к образованию озона
Гидропероксидный радикал окисляет NO ( как и  )
)
 ,
,
генерируя ОН радикал
Таким образом процесс окисления СН4 (и углеводородов вообще) – совокупность реакций, инициируемых солнечным излучение с λ=300-400 нм (которые приводят к О, ОН, НО2 ), протекающий при участии NO и приводящий к накоплению окислителей О3, ОН
Окисление у/в в этом процесс сопровождается также вторичным загрязнением атмосферы оксидом углерода, которые образуется при превращении СН2 О (что по масштабности сопоставимо с выбросами СО при сжигании топлива)

Окисление

НО2 также дает другая реакции

Формальную схему суммарной реакции окисления СН4 можно записать в виде
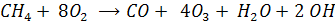
Таким образом в воздухе накапливается озон и гидроксорадикал, Причем скорость образования О3 зависит от содержания в атмосфере NO – она тем больше чем выше концентрация NO
Озон в тропосфере уже выполняет не защитную функцию как в стратосфере, а губительную вследствие сильных губительных свойств.
По подобной схеме окисляются и другие у/в алканового ряда. При этом скорость взаимодействия у/в с ОН радикалом сильно зависит от строения молекулы алкана (от стабильности образующегося у/в радикала) Очевидно скорость (как стабильность R•) возрастает с увеличением длины цепи и разветвленности алкана, Поэтому например скорость взаимодействия бутана с ОН радикалом на 3 порядка выше, чем с СН4 (к скорости соответственно 2,6·10-12 и 8·10-15 ).
При окислительных превращениях гомологов метана возможен еще один очень важный путь развития процесса, связанный с реакцией карбонильного радикала с О2
В случае с метаном простейший карбонильный радикал —  , образовавшийся из формальдегида, приводит к СО (по рассматриваемым реакциям)
, образовавшийся из формальдегида, приводит к СО (по рассматриваемым реакциям)
В случае с другими углеводородами карбонильный радикал (со структурой, определяемой структурой исходного углеводорода) присоединяет О2

Давая ацилпероксидный радикал, который в конечном итоге приводит к образованию важнейших (с точки зрения воздействия на ОС) продуктов веществ группы ПАН – пероксиацилнитратов.
ПАН- вещества общей формулы  (R – углеводородный радикал) являются наиболее опасными компонентами фотохимического смога. Из них наиболее известен пероксиацетил нитрат, т.е.
(R – углеводородный радикал) являются наиболее опасными компонентами фотохимического смога. Из них наиболее известен пероксиацетил нитрат, т.е. 
Таким образом фотохимические и окислительные превращения углеводородов с участие NОx являются главной причиной образования фотохимического смога – смеси газообразных веществ в сильными окислительными свойствами.
Образование фотохимического смога – обычное явление для крупных городов в большим количество автотранспорта. Выхлопные газы ДВС содержат в сове составе NОx и у/в, которые распространяются в фактически в зоне дыхания.
Обобщим все что говорилось об окислении у/в в единую схему, которой принято описывать образование фотохимического смога

Реакционноспособоные у/в (с двойными связями) также легко окисляются озоном, образуя при это альдегид (либо кетон – в зависимости от строения алкена) и кислоту – продукт последующего окисления по общей схеме:

Значительную долю у/в в атмосфере составляют ароматические у/в (в городах 30-40% от всех органических соединений). Они окисляются гидроскорадикалом по различны механизмам. Основным направление окисление является раскрытие цикла
C образованием перокидного радикала, который окисляется О2 с участием NO через ряд промежуточных стадий, включающих внутримолекулярную циклизацию пероксидного радикала) с раскрытием цикла с образованием дикарбонильных соединений.
Гомологи бензола дают большое число различных продуктов. Например при окислении толуола зарегистрировано более 40 (47) соединений, половина из которых диальдегиды и альдегиды дикарбонильных соединений легко фотохимически разлагаются с образованием различных радикалов.
Таким образом, фотохимический смог – смесь разнообразных продуктов фотохимических и окислительных реакций озона, альдегидов, кислот, пероксидных соединений, свободных радикалов, ПАН, главным из которых является окислители – озон и вещества группы ПАН. ПАН отличаются высокой токсичностью для человека, а также подавляют процесс фотосинтеза. Кроме того компоненты фотохимического смога будучи сильными окислителями, оказывают разрушительное действие на конструкционные материалы – усиливают коррозию металлических конструкций, разрушение строительных соединений.
Вредное воздействие смога на живые организмы обусловлено также присутствием альдегидов – весьма токсичных веществ.
Интересно проследить за изменением концентрации основных компонентов, участвующих в образовании фотохимического смога (например для города с активными физико-химическими условиями) в течение суток

График 3.1. изменение концентраций компонентов смога в городе в течение суток
Как видно из графиков – содержание NO достигает максимум быстрее по мере увеличения интенсивности движения транспорта, чем содержание NO2 (со смещением в несколько часов).
Концентрация окислителей достигает максимума только после нескольких часов воздействия солнечного света.
Образование фотохимического смога – один из процессов, в котором участвуют гидроксильные радикалы, окисляя у/в.
4. СВОБОДНО-РАДИКАЛЬНЫЕ РЕАКЦИИ В БИОЛОГИЧЕСКИХ СИСТЕМАХ
Радикалы свободные – атомы или группы химически связанных атомов, обладающие свободными валентностями, т.е. неспаренными (нескомпенсированными) электронами на внешней (валентной) орбитали. Наличие неспаренных электронов определяет высокую химическую реакционную способность и электронный спиновый магнетизм свободных радикалов.
Фундаментальные закономерности реакций с участием свободных радикалов были установлены Н.Н. Семеновым и его учениками и послужили основой для создания нового раздела физики — химической физики. Значение свободных радикалов в биологических процессах начали изучать в 30-х гг. 20 века.
Многочисленные данные свидетельствуют об участии свободных радикалов в нормальном функционировании живых клеток и тканей, а также в развитии некоторых патологических состояний. Установлено, что процесс старения сопровождается появлением и накоплением в тканях аномальных количеств свободных радикалов и перекисей. Свободные радикалы обладают выраженным мутагенным эффектом. Предполагают, что свободнорадикальные процессы играют существенную роль в онкогенезе. Антибактериальное действие некоторых антибиотиков объясняют их способностью образовывать свободные радикалы, обладающие цитолитическим эффектом в отношении бактериальных клеток.
Стабильные свободные радикалы используют в качестве меток и зондов при изучении конформации белков и нуклеиновых кислот, а также при исследовании механизма взаимодействия субстрата с ферментом, антигена с антителом, свойств биологических мембран и т.п.
В живых организмах свободные радикалы образуются в результате реакций одноэлектронного окисления или восстановления молекул соответствующими донорами или акцепторами электрона, например кислородом или металлами переменной валентности, а также непосредственно под действием ионизирующего или ультрафиолетового излучения.

Рисунок 3.1. Механизм пероксидного окисления липидов
Одним из механизмов управления внутриклеточного метаболизма является перекисное окисление липидов. Усиление этого процесса ведет к образованию избыточного количества свободных радикалов, что нарушает состояние клеточных мембран и коллоидное состояние протоплазмы. Ведущую роль в запуске перекисного окисления липидов играют первичные свободные радикалы (кислород и его активированные формы). При перекисном окислении липидов окислительным превращениям подвергаются полиненасыщенные жирнокислотные фосфолипиды, нейтральные жиры и холестерин, которые являются основными компонентами клеточных мембран. Поэтому при стимуляции перекисного окисления липидов в мембранах уменьшается содержание липидов, а также меняются их микровязкость и электростатический заряд. При более глубоком окислении фосфолипидов нарушается структура липидного бислоя и появляются дефектные зоны в мембранах клеток, а это нарушает функциональную активность. [5]
Перекисное окисление мембранных липидов – спутник рака, лучевых поражений, старения, некоторых отравлений; и во всех случаях применяют универсальный метод борьбы с этим злом. Суть этого метода довольно проста… Даже незначительное снижение стационарной концентрации активных частиц приводит к тому, что окисление перестает быть цепным. Это общее свойство всех цепных реакций, и останавливают их почти всегда одинаково, подменяя активные частицы менее реакционноспособными с помощью ловушек радикалов. Ловушками обычно служат соединения, при одноэлектронном окислении которых активными радикалами образуются тоже радикалы, но более инертным. В медицине в качестве таких ловушек (их принято называть антиоксидантами) используют и синтетические препараты (например, дибунол), и вещества природного происхождения (токоферол, аскорбиновая кислота).

Рисунок 4.2. Структура антиоксидантов, применяемая для борьбы с раковыми опухолями
В отличие от природных антиоксидантов — витаминов С и Е — дибунол является синтетическим препаратом. Антиоксиданты защищают липиды, белки и нуклеиновые кислоты от от окислительных повреждений, являющихся спутниками и причиной многих заболеваний.[6]
Одноэлектронное восстановление кислорода может происходить в клетках и тканях при участии ряда ферментов, таких как ксантиноксидаза, глюкозооксидаза и др. Действие некоторых антибиотиков основано на том, что они обеспечивают одноэлектронное восстановление молекулярного кислорода, отводя поток электронов от терминальных оксидаз бактериальной клетки. В результате образуются супероксидные, а значит, и гидроксильные радикалы, вызывающие в конечном счете гибель такой клетки.
При действии ионизирующего и ультрафиолетового излучений на аминокислоты, белки, нуклеотиды, нуклеиновые кислоты, жирные кислоты и липиды в результате отрыва электрона или разрыва химической связи образуются различные Р. с., а также первичные продукты фотолиза — сольватированный (т. е. захваченный молекулами среды, в основном воды) электрон, атом водорода и органические радикалы.
При затраченной энергии ионизирующего излучения в 100 эВ образуется 2—4 свободных радикала, при поглощении каждых 100 квантов света возникает всего несколько свободных радикалов.
В результате реакции с участием свободных радикалов в облученных белках и нуклеиновых кислотах происходит химическая модификация макромолекул (разрывы пептидных или нуклеиновых связей, образование «сшивок», химические изменения различных аминокислотных остатков, нуклеотидов и др.). Химическая модификация приводит к изменению структуры макромолекулы, ее формы и биохимических свойств, появлению точковых мутаций, к инактивации ферментов, разрушению биологических мембран и т.д.
Полагают, что функционально самой важной и универсальной по распространению группой свободных радикалов, в живых клетках являются семихиноны — анион-радикалы, постоянно образующиеся в ходе обмена веществ и энергии, а именно при окислительно-восстановительных превращениях переносчиков электронов в митохондриях, хлоропластах, мембранах бактериальных клеток и внутриклеточных мембранах эукариотов.
Большое значение для нормальной жизнедеятельности клетки, а также при развитии ряда патологическмх процессов имеют свободные радикалы, образующиеся при окислении липидов молекулярным кислородом, в первую очередь при окислении полиненасыщенных жирных кислот и жирных кислот фосфолипидов, входящих в состав липопротеидов и биологических мембран.
Как показал Б.Н. Тарусов (1954), механизм свободнорадикального окисления липидов в тканях и мембранах соответствует общим законам ценного окисления. Процесс цепного окисления начинается со стадии инициирования, причем в роли инициатора может выступать ОН-радикал, способный отнимать атом водорода у органических соединений (RH) с образованием воды и активного органического свободного радикала, участвующего в цепи последующих реакции.
Свободные радикалы участвуют в процессах окисления, окислительного фосфорилирования и тканевого дыхания. Установлено, что в норме в клетках постоянно протекают процессы свободнорадикального окисления липидов. Фагоцитоз микроорганизмов и вирусов сопровождается активацией свободнорадикального окисления.
Важную роль играют свободные радикалы в патологических процессах. Усиление свободнорадикалького окисления липидов может привести к нарушению нормальной жизнедеятельности организма и создать условия для развития ряда заболеваний. Признаками участия свободнорадикального окисления липидов в развитии того или иного заболевания, помимо активации свободнорадикального окисления, являются нарастание клинической симптоматики, а также улучшение состояния больного или его полное излечение в результате торможения свободнорадикального окисления липидов при терапии антиоксидантами.
Об активации процесса свободнорадикального окисления судят обычно по увеличению содержания в тканях и крови больных свободных радикалов, липидных гидроперекисей, альдегидов, в частности малонового диальдегида, а также по снижению содержания липидных антиоксидантов. Разработан метод регистрации уровня свободнорадикального окисления в организме больных в клинических условиях по содержанию пентана в выдыхаемом воздухе. Усиление свободнорадикального окисления липидов было обнаружено в печени при отравлении четыреххлористым углеродом, алкоголем, солями меди, озоном, кислородом; в коже после УФ-облучения; в очагах гипоксии и воспаления и при ожогах; в сетчатке глаза при чрезмерном освещении; во всех органах и тканях при развитии лучевой болезни и на определенных стадиях онкогенеза; при некоторых инфекционных болезнях, авитаминозах, воспалительных процессах: в мозге животных усиление свободнорадикального окисления липидов было выявлено при экспериментальной эпилепсии и т.д. Однако патогенетическая роль свободнорадикального окисления липидов во всех этих случаях пока не ясна.
Состояние больных или животных (в условиях эксперимента) почти всегда значительно улучшается после терапии биоантиоксидантами: например, уменьшается эритема, вызванная УФ-облучением кожи, снижается токсическое действие на организм четыреххлористого углерода, купируются эпилептические припадки (в эксперименте), увеличиваются сроки консервации клеток и органов. Описано успешное применение антиоксидантов при печении ожогов и ишемической болезни сердца, связанной с атеросклерозом.
Большое внимание исследователи уделяют роли свободных радикалов в онкогенезе, Обнаружена корреляция между способностью ряда онкогенов к образованию свободных радикалов и их онкогенной активностью. Как правило, по мере развития опухоли концентрация свободных радикалов в тканях снижается в 2—6 раз по сравнению с контролем, а интенсивность свободнорадикального окисления в других тканях организма обычно повышается, особенно на терминальных стадиях болезни, что, возможно, связано с перераспределением антиоксидантов между тканью злокачественной опухоли и другими тканями.[7]
ЗАКЛЮЧЕНИЕ
В своей работе я рассмотрела, как проявляют себя свободные радикалы в реакциях, происходящих в природной среде. Среди них есть множество жизненно важных для организмов, как например реакции окисления в клетках, значимых для окружающей среды, таких как реакции с озоном и оксидами азота в атмосфере или процессы в водоемах.
Здесь я постаралась коротко описать наиболее значимые и интересные аспекты этого вопроса, поскольку в общем тема очень обширная и по сути каждый радикал заслуживает отдельного подробного изучения.
СПИСОК ЛИТЕРАТУРЫ
1. Koppenol, 1990 #7
2. www.cmjournal.com/rp218.htm
3. admin.novsu.ac.ru/uni/scpapers.nsf
4. Чибисова Н.В., Долгань Е.К. Экологическая химия: Учебное пособие / Калинингр. ун-т. — Калининград, 1998. — 113 с.
5. moikompas.ru/compas/lipid_oxidation
6. vivovoco.rsl.ru/HOME/PAPERS/TEXT/DRUGS/DRUGS_3.HTM
7. www.curemed.ru/medarticle/articles/34450.htm
www.ronl.ru
Перечисленные в таблицах 1 и 2 радикалы можно считать природными, поскольку они в определенном количестве всегда образуются в наших клетках. Наряду с этими радикалами разрушительное действие могут оказывать радикалы, появляющиеся при таких воздействиях, как ионизирующее излучение, ультрафиолетовое облучение или даже освещение интенсивным видимым светом, например, светом лазера. Такие радикалы можно назвать чужеродными. К ним принадлежат также радикалы, образующиеся из попавших в организм посторонних соединений, ксенобиотиков, многие из которых оказывают токсическое действие именно благодаря свободным радикалам, образующимся при метаболизме этих соединений.
Свободнорадикальные процессы в облученных организмах в разных тканях облученных животных не одинаковы. Так, в одной группе тканей и клеток (головной мозг, скелетные мышцы, эритроциты) после облучения животных в дозах 6-15 Гр не обнаружены пострадиационные изменения содержания свободных радикалов; в другой (печень и почки) происходит усиление процесса сополимеризации на первые-вторые сутки после облучения; в третьей группе (плазма крови, селезенка) наблюдаются фазовые изменения количества свободных радикалов в ткани. В плазме крови в течение двух суток после облучения происходит непрерывное увеличение концентрации свободных радикалов, интенсивность накопления которых зависит от дозы излучения в широком диапазоне. Еще ранее было показано, что основное количество активных оксипродуктов («радиотоксинов») возникает в тканевых ненасыщенных липидах, количество токсических продуктов возрастает в зависимости от величины дозы и времени, прошедшего после облучения.
Фундаментальные исследования, экспериментально подтвердившие важнейшую роль свободных радикалов и антиоксиданов, регулирующих их уровень, в развитии лучевого поражения клетки и организма проводились в радиобиологических школах Н. М. Эмануэля и Б. Н. Тарусова. Получены многочисленные данные, указывающие на связь между количеством индуцируемых излучением свободных радикалов и степенью лучевого поражения клеток. Большинство таких данных получено с использованием методов, позволяющих регулировать уровень и дальнейшее превращение свободных радикалов в облученной клетке.
Активные кислород-содержащие соединения — АКСС — принимают непосредственное участие в зарождении и развитии нарушений системы окислительно-восстановительного равновесия в облученной клетке.
Система окислительно-восстановительного гомеостаза клетки и ее изменения после облучения
Известно, что система окислительно-восстановительных процессов составляет основу жизнедеятельности клетки и организма, а кислород играет ключевую роль в энергетике дыхания нормальных аэробных клеток, являясь единственным акцептором электронов.
Основное количество молекулярного кислорода в клетке — 90-98% расходуется в реакциях тканевого дыхания и микросомального окисления, катализируемых соответственно митохондриальной цитохромоксидазой, а также и другими оксидазами, в результате чего в дыхательных цепях происходит запасение энергии и осуществляется четырехэлектронное восстановление О2 с образованием двух молекул воды:
Значительно реже под действием ферментов — оксигеназ — происходит включение одного из атомов кислорода в биоорганические молекулы:
О2 + М + 2Н МО + Н2О.
Так, важную роль в окислении ненасыщенных липидов играют ферментные реакции, катализируемые липоксигеназами и циклооксигеназами — первыми ферментами на путях образования специфических регуляторов метаболизма — эйкозаноидов.
Около 2% поглощаемого кислорода восстанавливается самопроизвольным, неферментативным путем. Эти реакции всегда начинаются с присоединения к молекуле О2 одного электрона и образования аниона-радикала (содержащего неспаренный электрон) — супероксида :
Донорами электронов в клетке служат Fe2+, Cu+, семихинон и другие промежуточные участники процессов тканевого дыхания, а под воздействием ионизирующей радиации, как известно, электроны генерируются вследствие ионизации воды и биоорганических молекул.
В основном состоянии молекулярный кислород парамагнитен — представляет собой триплет — «бирадикал», т. е. имеет два неспаренных электрона (с параллельными спинами) на различных орбиталях. По этой причине прямые, неферментативные реакции с кислородом могут осуществляться по свободнорадикальным механизмам. Большинство органических молекул синглетны (электроны с антипараллельными спинами), поэтому, вследствие различий с триплетным кислородом в направлении спинов, их реакции с молекулярным кислородом из-за спиновых запретов затруднены и протекают очень медленно. Кинетические барьеры одноэлектронного восстановления кислорода в клетке достаточно высоки, поэтому образование супероксида процесс медленный, однако он очень важен для формирования системы окислительно-восстановительного гомеостаза в клетке, так как J участвует в генерации многих окислителей — активных кислород-содержащих соединений — АКСС.
Активные формы кислорода
К активным формам кислорода (АФК) относятся свободные радикалы — супероксидный анион-радикал (супероксид), гидроксид- и гидропероксид (пергидроксид)-радикалы и нейтральные молекулы — пероксид водорода и синглетный кислород .
АФК обнаруживаются в клеточных органеллах. Главными местами их образования являются митохондриальная, ядерная, плазматическая и микросомальная мембраны у животных, хлоропласты и хроматофоры у растений.
В клетке существуют ферментный и неферментный пути образования АФК. Метаболические, ферментные пути образования АФК, участвующие в нормальной жизнедеятельности клетки, представлены в схеме Бурлаковой-Михайлова-Мазурика.
В ферментной генерации АФК участвуют: дыхательная цепь митохондрий и система микросомального окисления, системы оксидаз (ксантин-, флавин-, НАДФ.Н*-оксидазы) и цитохромов, глутатионпероксидаза и супероксиддисмутаза (СОД).
Неферментативное образование основных АФК и их превращения происходят по схеме, изображенной на рис. 1, в следующих реакциях
Рис.1. Схема последовательности образования АФК в клетке в цепи неферментного одноэлектронного восстановления кислорода до воды
Синглетный кислород, содержащийся в хлоропластах растений и в зрительных рецепторах животных, занимает особое место среди АФК. Образование его здесь связано не только с реакцией, а преимущественно с фотохимическими процессами. Поглощая квант света, пигмент-фотосенсибилизатор переходит в синглетное, а затем в триплетное возбужденные состояния. В обоих возбужденных состояниях молекулы пигмента взаимодействуют с молекулярным кислородом, передают ему свою энергию и превращают его в активный окислитель.
Супероксид при взаимодействии с органическими биомолекулами обладает слабой реакционной способностью, так как имеет невысокий окислительно-восстановительный потенциал. Однако он выполняет важную роль, продолжая дальнейшую цепь одноэлектронного восстановления кислорода путем образования молекулы пероксида (перекиси) водорода Н2О2.
Пероксид водорода — сильный окислитель и образуется при реакции супероксида с электроном и двумя протонами:
Экспериментально показано, что гидроксид-радикал относится к наиболее эффективным окислителям АКСС.
Биологическая роль АФК может осуществляться в жизненно необходимых реакциях иммунитета и воспаления: АФК способствуют образованию цитокинов и иммунных рецепторов, миграции лейкоцитов в «аварийные ткани», выполняют бактерицидную функцию в фагоцитозе.
Известно, что возрастающий уровень 1О2 может быть причиной обновления «отработанных» субклеточных компонентов фоторецепторов, а также фотодинамического действия пигментов в клетках, не имеющих фоторецепторных систем.
В конце XX в. получены данные о способности АФК выполнять сигнальную функцию в клеточной регуляции. Показано, что АФК стимулируют накопление цАМФ и цГМФ, ионов Са2+ в цитозоле, активацию протеинкиназ, протеинтирозинкиназ и подавление активности протеинфосфатаз. Помимо стимуляции процессов фосфорилирования белков, АФК активируют и белок Ras, участвующий в передаче сигналов в ядро клетки.
Вопрос о биологической роли АФК находится также в связи с инициированием ими значимых процессов в клетке — перекисного окисления липидов.
В настоящее время окончательно установлено, что АФК способны инициировать цепные реакции липопероксидации в норме, а их избыток — усиливать цепные процессы при лучевом поражении и других физических и химических воздействиях.
Непрямой эффект радиации при радиолизе воды в облученной клетке был изучен давно. Однако биологическое действие ОН·и других АКСС в норме и в механизме усиления радиационных повреждений более полно стало раскрываться только в последние 10-20 лет.
Гидроксид-радикал — чрезвычайно активный окислитель и способен разрушать фактически любую находящуюся рядом с ним молекулу в клетке. При взаимодействии с нуклеиновыми кислотами он вступает в реакции с азотистыми основаниями, образуя продукты их повреждения, а также разрушает углеводные мостики между нуклео-тидами и вызывает разрывы цепей ДНК и РНК. Благодаря этому ОН· способен вызывать мутации и гибель клеток. Взаимодействуя с другой мишенью — биологическими мембранами (БМ), гидроксид-радикал внедряется в липидный слой, инициирует цепные реакции липопероксидации, приводит тем самым к нарушениям структуры и функции мембран, инициируя процессы гибели клеток. При облучении клетки в больших дозах, действуя на тиольные белковые молекулы, ОН· денатурирует их и инактивирует ферменты Все же следует оговориться, что время жизни ОН· небольшое и гидроксид-радикал успевает диффундировать лишь на один-два молекулярных диаметра, взаимодействуя лишь с молекулами близлежащих компонентов клетки. Поэтому вероятность прямого взаимодействия его с ядерной ДНК малая. Диффундирующей «скрытой» формой гидроксид-радикала является, пероксид водорода — более длительно живущая АФК, содержащаяся в клетке в относительно больших количествах. Молекула Н2О2 (вступая в реакцию с ионом металла) «доставляет» ОН· в мембранные структуры и ядро клетки.
Важнейшую роль в протекании окислительно-восстановительных реакций, к продуктам которых относятся АФК-прооксиданты, играет и недавно открытая система NО, выполняющая функции регулятора многих ключевых физиологических и биохимических процессов.
Активные соединения азота
В последние два десятилетия проведены фундаментальные исследования, открывшие новую страницу в молекулярной биологии: установлено, что, казалось бы, простое химическое соединение — моноксид азота NO, образующееся в организме из L-аргинина ферментативным путем , — выполняет функции одного из наиболее универсальных регуляторов метаболизма.
Суммарное уравнение ферментной реакции образования NО, включающее пятиэлектронное окисление атома азота аргинина сопряженное с окислением НАДФ.Н, имеет вид:
2 Арг+3 НАДФ.Н+4О2+3 Н+ = 2 Цит+2 NО+3 НАДФ++4Н2О.
Ферментная реакция генерации моноксида азота обеспечивается изоформами эндотелиальной и нейрональной NO-синтаз.
Обнаружено, что активные соединения NО выполняют многообразные физиологические регуляторные функции в организме, выступая в качестве антагониста адренергической нервной системы, регулирующего по эфферентным нервам деятельность дыхательной, мочеполовой, мышечной, секреторной, сосудистой и других систем. Установлено активирующее действие NО на растворимую форму важнейшего внутриклеточного регуляторного белка — гуанилатциклазы — фермента, ответственного за синтез одного из вторичных мессенджеров — цГМФ.
Малые размеры двухатомной газообразной молекулы N0 и отсутствие заряда позволяют ей легко проникать через плазматические и внутриклеточные мембраны. Молекулы моноксида азота легко диффундируют в биологических средах и являются относительно долгоживущими.
Наряду с нормальными регуляторными функциями, NО, образуясь в фагоцитах в больших концентрациях, проявляет цитотоксическую активность, обнаруживая эффекторные свойства системы клеточного иммунитета.
Установлено также, что длительная генерация NО способна вызывать патологию различного генеза. Так, наряду с другими АКСС, оксид азота и его производные являются ключевыми факторами воспаления, инфекции, канцерогенеза, а также развития радиационных, стрессорных и адаптивных ответов клеток и организма на соответствующие воздействия.
Показано, что цитотоксические и цитогенетические эффекты обусловлены образованием чрезвычайно активного окислителя — пе-роксинитрита, возникающего в реакции взаимодействия оксида азота с супероксидным анион-радикалом:
turboreferat.ru
Как видно, пероксинитрит представляет собой связующее звено двух высокоактивных низкомолекулярных кислород- и азотсодержащих систем: АФК и АСА (активные соединения азота). Пероксинитрит может разлагаться с образованием
Протонированный пероксинитрит так же, как и пероксид водорода, выполняет функции диффундирующей «скрытой» формы гидроксид-радикала.
Установлено, что пероксинитрит и его протонированная форма, диффундируя в клетке и проникая через мембраны с помощью переносчиков анионов, имеют уникальный спектр химической реактивности: разрывы цепочек и окисление оснований ДНК, нитрование гуанина и белков, окисление липидов биологических мембран и т.д. Результатом многих таких реакций могут быть цитогенетические эффекты и мутагенез.
NО является источником не только пероксинитрита и гидроксид-радикала, но и синглетного кислорода, который образуется при реакции моноксида азота с пероксидом водорода.
Сильные окислители ·NO и ONO2-, обладая высокой реакционной способностью, могут влиять на цепные свободнорадикальные реакции. Их оксидазная активность во многом связана со способностью АСА модифицировать уровень продуктов перекисного окисления липидов.
Продукты цепных реакций перекисного окисления липидов
Перекисное окисление липидов исследовано не только в модельных системах, но и на всех уровнях организации живого и имеет свои особенности в норме и патологии клетки.
Известно, что биологические мембраны способны подвергаться интенсивной окислительной деградации, так как ненасыщенные участки жирных кислот (линоленовой, арахидоновой и др.) фосфолипидов чрезвычайно чувствительны к окислению.
Многочисленные данные молекулярной биологии показали, что при различных патологиях, в том числе и при лучевом поражении, одной из первых клеточных структур повреждается мембрана митохондрий.
Лучевые оксидативные изменения структуры и функций БМ (повреждения фосфолипидного барьера) приводят к дальнейшему нарушению регуляции окислительно-восстановительного равновесия, возрастанию мембранной проницаемости, дезинтеграции внутриклеточных процессов, выравниванию градиентов с фатальным для клетки исходом.
Большое содержание полиненасыщенных жирных кислот в фосфолипидах определяет высокую способность БМ к неферментным цепным реакциям окисления и образованию инициаторов и продуктов окисления, обладающих оксидазной активностью. В результате цепных реакций липопероксидации образуются свободные радикалы, гидроперокси-ды — LOOH и другие молекулярные ППОЛ.
Первичная роль в зарождении неферментивных цепных процессов ПОЛ в клетке, как и in vitro, принадлежит свободным электронам, донорами которых могут служить Fe2+, Cu2+, семихиноны; сигнально-пусковыми продуктами являются, как известно, , Н2О2, ОН· и NО·. В инициировании цепных реакций могут принимать участие и другие продукты АКСС, проникающие в липиды БМ.
Биологическая значимость цепного процесса, инициированного в клетке, заключается в том, что он способствует накоплению токсических продуктов окисления в связи с их многократным циклическим воспроизведением.
Цепной процесс липопероксидации в клетке протекает в такой последовательности:
1. Молекулы ненасыщенных жирных кислот липидов LH окисляются преимущественно радикалом ОН' (или другими АКСС):
2. Образовавшиеся радикалы липида L· претерпевают в клетке дальнейшее окисление с помощью молекулярного кислорода или с большей вероятностью — АКСС (АФК, NO-системами). Появляются пероксил- и алкоксил-LO·-радикалы
3. Далее происходят реакции
В процесс вовлекаются все новые и новые молекулы липидов LH и активных форм кислорода. В результате накапливаются гидропероксиды LOOH. Число радикалов L·, LO· (или LО2) не изменяется (принцип неуничтожимости свободной валентности, «бессмертия свободных радикалов»), хотя структурно радикалы могут отличаться друг от друга.
4. Образующиеся липидные гидропероксиды вступают в реакциюс радикалами ОН· (которые могут мигрировать в клетке на большие расстояния с помощью пероксида водорода или пероксинитрита):
или с радикалом LO·:
и осуществляют вторичное инициирование цепи реакции и протекание разветвленных свободнорадикальных цепных процессов.
5. При окислительном распаде липидных гидропероксидов образуются высоко лабильные ППОЛ, представляющие собой жирнокислотные фрагменты, содержащие пероксидные, эпоксидные,альдегидные и кетонные группы. Лишь в последние годы, бла годаря применению высокочувствительных аналитических методов, удалось идентифицировать некоторые из ППОЛ и показать исключительно важную их роль в биологии клетки.
Радиационно-биофизические исследования цепных свободно-радикальных механизмов вызвали резкий приток работ, посвященных роли липопероксидации в патогенезе очень многих, самых разнообразных клеточных патологий и заболеваний организма. Однако не менее важными являются и другие работы, на основе которых сформировались представления и о биологической роли ППОЛ в условиях физиологической нормы.
Процессы ПОЛ широко распространены в нормальной жизнедеятельности клетки. Биологическая роль ППОЛ и их связь с активными формами кислорода и NО подтверждается работами, в которых показано, что синтез тканевых гормонов — простагландинов, лейкотриенов и др. — нуждается в образовании перекисей ненасыщенных жирных кислот.
В настоящее время хорошо известна важная биологическая роль взаимного влияния активности ППОЛ и физико-химических свойств липидного бислоя БМ. Так, ППОЛ оказывают значительное влияние на функционирование плазматических и внутриклеточных БМ, обладая способностью включаться в механизмы формирования, разборки и обновления их структуры, являясь физиологическими регуляторами важнейших функций и универсальными модификаторами структуры БМ
Установлена зависимость интенсивности реакций липопероксидации от степени ненасыщенности жирных кислот и состояния структурной организации биологических мембран: молекулярной подвижности липидов, прочности липид-липидных и белок-липидных взаимодействий Интенсивность свободнорадикальных процессов перекисного окисления липидов, находящихся под контролем АКСС, эндогенных антиокислителей и активности ферментов, взаимосвязана с составом и физическим состоянием фосфолипидов биологических мембран (текучестью, окисляемостью и др.), с чувствительностью их рецепторов к сигналам лигандов. Активация ПОЛ ведет к ускорению выхода легко окисляемых липидов из БМ и обогащению их липидов резистентными к окислению фракциями. Это в свою очередь замедляет скорость ЛПО и возвращает этот процесс к норме.
Окислительные процессы в облученной клетке
Радиационные изменения окислительно-восстановительного равновесия в живых системах начинаются уже сразу, с момента передачи энергии ионизирующих излучений биоорганическим молекулам. В процессе ионизации происходит образование электронов, которые представляют особый интерес с биологической точки зрения, так как вызывают поляризацию и окисление молекул. Электроны в водной среде стабилизируются до гидратированного состояния, превращаясь в е-гидр , способные диффундировать на значительные молекулярные расстояния и эффективно взаимодействовать с молекулярным кислородом, повышая уровень содержания супероксида О2 (и других окислительных радикалов и продуктов) не только в водных растворах, но и в облученной клетке. Другая часть электронов окисляет биоорганические молекулы, первоначально превращая их в катионы и радикалы.
Известно, что ионизирующие излучения могут индуцировать в клетке образование пероксинитрита. Уже первые работы (В.Д.Микоян и др., 1994) показали, что его содержание в облученной клетке возрастает за счет усиления активности NO-синтазы. Накапливающиеся в облученной клетке оксипродукты ОН·, Н2О2 и высоко эффективны и могут быть отнесены к радиотоксинам (АКСС-радиотоксины — АКССрт). Важным свойством АКССрт является их «пусковая» способность — инициировать и усиливать цепной процесс перекисного окисления и инициировать образование сильных и относительно долгоживущих окислителей — ППОЛ. В облученной клетке избыточно накопленные ППОЛ известны под названием «липидных радиотоксинов» (ЛРТ). Они образуются, в основном, при окислении полиненасыщенных жирных кислот фосфолипидов БМ.
Обе группы радиотоксинов АКССрт и ЛРТ объединяются под общим названием оксирадиотоксины (ОРТ). Они вызывают в клетках и тканях облученного организма повреждения, приводящие к так называемому лучевому токсическому эффекту.
В начальный период после облучения в клетке происходит нарушение баланса между образованием ОРТ и антиокислителями. Эти изменения неспецифичны для ионизирующих излучений и известны под названием оксидативного стресса. Вначале при этом защитные антиокислительные ресурсы клетки активируются и сдерживают выход окислителей из стационарного режима. Затем, по мере развития лучевого поражения, содержание антиокислителей постепенно исчерпывается, и накопление радиотоксинов приобретает бесконтрольный характер.
Известно, что содержание АКССрт и ЛРТ может увеличиваться не только в липидах, но и в водной фазе клетки. Поэтому эти продукты способны взаимодействовать с обоими радиационными клеточными мишенями — БМ и ДНК. Результат этих взаимодействий — дальнейшее изменение системы окислительно-восстановительного гомеостаза, перестройка физико-химической регуляции метаболизма в клетке, нарушение барьерно-матричных свойств БМ, образование аддуктов [ОРТ ДНК]. Участвуя в непрямом действии излучений, ОРТ, и в частности ЛРТ, вызывают хромосомные аберрации, мутации, вовлекаются в формирование радиационно-индуцируемой нестабильности генома.
В экстремальной стадии оксидативных повреждений происходят многоэтапные и значительные цитогенетические повреждения мишеней, завершающиеся гибелью облученной клетки.
Радиотоксины
Понятие о радиотоксинах возникло еще в начале прошлого столетия. Ряд немецких радиобиологов-биохимиков вели поиски эндогенных «клеточных ядов», образующихся под действием излучений. Предполагалось, что эти «яды» могли быть гистамином, холином, какими-то иными низкомолекулярными продуктами распада белков. Строились гипотезы о роли этих и других веществ в зарождении и формировании лучевой болезни.
Однако достаточно глубоко рассматривать возникающие гипотезы не представлялось возможным ввиду отсутствия в то время знаний о механизмах действия ионизирующих излучений и данных о химической идентификации веществ, выделяемых из облученных организмов. Позднее делались попытки найти сходство в эффектах облучения и действия бактериальных экзотоксинов или синтезированных препаратов — иприта, эмбихина, мильерана и их аналогов, имитирующих действие ионизирующей радиации (радиомиметиков), а также эндогенных хиноноподобных веществ и липидов.
Радиобиологи ряда лабораторий мира часто обращались к мысли, что многие проявления лучевого поражения можно объяснить образующимися при облучении токсическими веществами. На роль радиотоксинов в развитии лучевого поражения указывали многочисленные исследования по введению экстрактов, выделенных из облученных организмов, интактным биологическим объектам; по дистанционному действию излучений; эксперименты с парабионтами .Все больше укреплялось предположение, что токсичность является результатом появления в облученных клетках избыточных концентраций продуктов обычного метаболизма, но в условиях выхода его из нормального режима, а не появления новых, чуждых для организма веществ.
Подводя итог этим исследованиям, Б.Н.Тарусов еще в 1954 г. отмечал, что анализ развития лучевого поражения во времени позволяет говорить о механизмах усиления незначительных первичных радиационных повреждений — образования активных «первичных токсинов» (тогда еще не идентифицированных), действие которых является пусковым для автокаталитической цепной реакции.
Б. Н. Тарусов выдвинул и развил теорию свободнорадикальных цепных процессов лучевого поражения клетки и организма, сводящуюся в основном к следующим трем положениям.
turboreferat.ru
