
|
|
|
|
 Far Far |
| WinNavigator |
| Frigate |
| Norton
Commander |
| WinNC |
| Dos
Navigator |
| Servant
Salamander |
| Turbo
Browser |
|
|
| Winamp,
Skins, Plugins |
| Необходимые
Утилиты |
| Текстовые
редакторы |
| Юмор |
|
|

|
File managers and best utilites |
Доклад: Белки и аминокислоты. Реферат по химии белки
Доклад - Химия белка - Химия
Биохимия — это наука о химических и физико-химических процессах, которые протекают в живых организмах и лежат в основе всех проявлений жизнедеятельности. Биохимия возникла на стыке органической химии и физиологии в конце прошлого века.
СТРОЕНИЕ И СВОЙСТВА БЕЛКОВ
БЕЛКИ или ПРОТЕИНЫ — это высокомолекулярные азотсодержащие органические вещества, линейные гетерополимеры, структурным компонентом которых являются аминокислоты, связанные пептидными связями.
Кроме понятия «белок», в химии встречается термины «ПЕПТИД» и «ПОЛИПЕПТИД». Пептидом обычно называют олигомер, состоящий не более чем из 10 аминокислот. Но встречаются и молекулы, содержащие от 10 до 100 аминокислот – они относятся к группе небольших ПОЛИПЕПТИДОВ, крупные же полипептиды могут содержать и более 100 аминокислот. Столько же аминокислот могут содержать и некоторые небольшие белки. Поэтому граница по количеству аминокислотных остатков, а, стало быть, и по молекулярной массе, между белками и полипептидами, весьма условна.
В природе встречаются десятки тысяч различных белков. И все они отличаются друг от друга по пяти основным признаком.
Основные различия в строении белковых молекул
По количеству аминокислот
По соотношению количества различных аминокислот. Например, в белке соединительной ткани коллагене 33% от общего количества аминокислот составляет глицин, а в молекуле белкового гормона инсулина, вырабатываемого в поджелудочной железе, содержание глицина гораздо меньше – всего 8%.
Различная последовательность чередования аминокислот. Это означает, что даже при одинаковом соотношении разных аминокислот в каких-нибудь двух белках порядок их расположения этих аминокислот различен, то это будут разные белки.
Количество полипептидных цепей в различных белках может варьировать от 1 до 12, но если больше единицы, то обычно четное (2, 4, 6 и т.п.)
По наличию небелкового компонента, который называется «ПРОСТЕТИЧЕСКАЯ ГРУППА». Если ее нет, то это – простой белок, если есть – сложный белок
В природе встречается около 150 аминокислот. Для построения белков используются только 20 из них, хотя в метаболизме организма человека участвует большее количество аминокислот. Эти 20 аминокислот имеют несколько общих признаков строения (общие свойства аминокислот):
1. Все они являются альфа-аминокислотами. Аминогруппа общей части всех аминокислот присоединена к альфа-углеродному атому.

2. По стереохимической конфигурации альфауглеродного атома все они принадлежат к L-ряду.
Следовательно, все эти 20 аминокислот имеют совершенно одинаковый фрагмент молекулы. Различаются они по строению радикалов.
Молекула воды обладает полярными свойствами.
Атом кислорода сильнее притягивает электроны, чем атомы водорода, поэтому электронное облако смещено в сторону кислорода. Степень полярности определяется величиной частичных зарядов и расстоянием между центрами тяжести этих зарядов. Таким образом, молекула воды является диполем.


Молекулы воды структурированы и образуют кластеры.
В эти кластерные структуры хорошо встраиваются молекулы, которые сами являются полярными, потому что полярные вещества хорошо растворимы в воде. Полярными являются все те молекулы, которые содержат электроотрицательные атомы. В молекулах белков электроотрицательными атомами являются O (кислород), N (азот) и S (сера).
Высокая полярность обеспечивает остальные общие свойства аминокислот:
3. Хорошая растворимость в воде благодаря наличию общего фрагмента молекулы. Общий фрагмент обладает полярными свойствами, потому что содержит карбоксильную группу –COOH (при физиологическом значении pH эта группа заряжена отрицательно), и аминогруппы -Nh3 (при физиологическом значении pH заряжена положительно).
4. Способность к электролитической диссоциации. Аминокислоты существуют в водном растворе в виде амфионов (биполярных ионов). В целом такая молекула при нейтральном значении pH (при pH=7) электронейтральна.

5. Наличие ИЗОЭЛЕКТРИЧЕСКОЙ ТОЧКИ (ИЭТ, pI). (ИЭТ) — это значение pH среды, при котором молекула амфотерного вещества (например, аминокислоты) находится в электронейтральном состоянии .
РАЗЛИЧИЯ В СТРОЕНИИ АМИНОКИСЛОТ
Радикалы аминокислот могут значительно отличаться друг от друга по строению.
Если есть дополнительные карбоксильные группы в радикале, то заряд молекулы в нейтральной среде отрицателен, а ИЭТ такой молекулы находится в кислой среде.
Аминокислота, в радикале которой есть дополнительная аминогруппа (Nh3 -группа), в нейтральной среде заряжена положительно. ИЭТ такой аминокислоты находится в щелочной среде (pI>7). К таким аминокислотам относятся лизин, аргинин и гистидин.
Аминокислота, в радикале которой есть дополнительная карбоксильная группа (COOH-группа), в нейтральной среде заряжена отрицательно. ИЭТ такой аминокислоты находится в кислой среде (pI<7). К ним относятся аспарагиновая кислота и глутаминовая кислота.
С помощью значения рН внешней среды характеризуют соотношение -СООН и -Nh3 групп. Это относится и к пептидам, и к белкам.
Различные функциональные группы, содержащиеся в радикалах аминокислот, придают им способность к взаимодействию с образованием разных типов соединений. Приведем примеры таких взаимодействий.






В связи с различиями в строении радикалов различны и физико-химические свойства аминокислот.
КЛАССИФИКАЦИЯ АМИНОКИСЛОТ
Существуют три типа классификации:
ФИЗИКО-ХИМИЧЕСКАЯ – основана на различиях в физико-химических свойствах аминокислот.
Гидрофобные аминокислоты (неполярные). Компоненты радикалов содержат обычно углеводородные группы, где равномерно распределена электронная плотность и нет никаких зарядов и полюсов. В их составе могут присутствовать и электроотрицательные элементы, но все они находятся в углеводородном окружении. Например, в радикале метионина сера окружена углеводородными группировками, которые не позволяют этому элементу проявлять своих электроотрицательных свойств: -(Ch3 )2 -S-Ch4. Аналогичная ситуация наблюдается, например, и в отношении азота, находящегося в составе радикала триптофана.
Гидрофильные незаряженные (полярные) аминокислоты. Радикалы таких аминокислот содержат в своем составе полярные группировки:

Эти группы взаимодействуют с дипольными молекулами воды, которые ориентируются вокруг них.
Отрицательно заряженные аминокислоты. Сюда относятся аспарагиновая и глутаминовая кислоты. Имеют дополнительную СООН-группу в радикале — в нейтральной среде приобретают отрицательный заряд.

Все они гидрофильны.
Положительно заряженные аминокислоты: аргинин, лизин и гистидин. Имеют дополнительную Nh3 -группу (или имидазольное кольцо, как гистидин) в радикале — в нейтральной среде приобретают положительный заряд.

Все они также являются гидрофильными.

Такие свойства характерны для свободных аминокислот. В белке же ионогенные группы общей части аминокислот участвуют в образовании пептидной связи, и все свойства белка определяются только свойствами радикалов аминокислот.
Не все аминокислоты, принимающие участие в построении белков человеческого тела, способны синтезироваться в нашем организме. На этом основана еще одна классификация аминокислот — биологическая.
II. Биологическая классификация.
а) Незаменимые аминокислоты, их еще называют «эссенциальные». Они не могут синтезироваться в организме человека и должны обязательно поступать с пищей. Их 8 и еще 2 аминокислоты относятся к частично незаменимым.
Незаменимые: метионин, треонин, лизин, лейцин, изолейцин, валин, триптофан, фенилаланин.
Частично незаменимые: аргинин, гистидин.
а) Заменимые (могут синтезироваться в организме человека). Их 10: глутаминовая кислота, глутамин, пролин, аланин, аспарагиновая кислота, аспарагин, тирозин, цистеин, серин и глицин.
III. Химическая классификация — в соответствии с химической структурой радикала аминокислоты (алифатические, ароматические).
Белки синтезируются на рибосомах, не из свободных аминокислот, а из их соединений с транспортными РНК (т-РНК).
Этот комплекс называется «аминоацил-т-РНК».
ТИПЫ СВЯЗЕЙ МЕЖДУ АМИНОКИСЛОТАМИ В МОЛЕКУЛЕ БЕЛКА
2 группы:
1. КОВАЛЕНТНЫЕ СВЯЗИ — обычные прочные химические связи.
а) пептидная связь
б) дисульфидная связь
2. НЕКОВАЛЕНТНЫЕ (СЛАБЫЕ) ТИПЫ СВЯЗЕЙ — физико-химические взаимодействия родственных структур. В десятки раз слабее обычной химической связи. Очень чувствительны к физико-химическим условиям среды. Они неспецифичны, то есть соединяются друг с другом не строго определенные химические группировки, а самые разнообразные химические группы, но отвечающие определенным требованиям.
а) Водородная связь
б) Ионная связь
в) Гидрофобное взаимодействие
ПЕПТИДНАЯ СВЯЗЬ.
Формируется за счет COOH-группы одной аминокислоты и Nh3 -группы соседней аминокислоты. В названии пептида окончания названий всех аминокислот, кроме последней, находящейся на «С»-конце молекулы меняются на «ил»
Тетрапептид: валил-аспарагил-лизил-серин


ПЕПТИДНАЯ СВЯЗЬ формируется ТОЛЬКО ЗА СЧЕТ АЛЬФА-АМИНОГРУППЫ И СОСЕДНЕЙ COOH-ГРУППЫ ОБЩЕГО ДЛЯ ВСЕХ АМИНОКИСЛОТ ФРАГМЕНТА МОЛЕКУЛЫ!!! Если карбоксильные и аминогруппы входят в состав радикала, то они никогда(!) не участвуют в формировании пептидной связи в молекуле белка.
Любой белок — это длинная неразветвленная полипептидная цепь, содержащая десятки, сотни, а иногда более тысячи аминокислотных остатков. Но какой бы длины ни была полипептидная цепь, всегда в основе ее — стержень молекулы, абсолютно одинаковый у всех белков. Каждая полипептидная цепь имеет N-конец, на котором находится свободная концевая аминогруппа и С-конец, образованный концевой свободной карбоксильной группой. На этом стержне сидят как боковые веточки радикалы аминокислот. Числом, соотношением и чередованием этих радикалов один белок отличается от другого. Сама пептидная связь является частично двойной в силу лактим-лактамной таутомерии. Поэтому вокруг нее невозможно вращение, а сама она по прочности в полтора раза превосходит обычную ковалентную связь. На рисунке видно, что из каждых трех ковалентных связей в стержне молекулы пептида или белка две являются простыми и допускают вращение, поэтому стержень (вся полипептидная цепь) может изгибаться в пространстве.
Хотя пептидная связь довольно прочная, ее сравнительно легко можно разрушить химическим путем – кипячением белка в крепком растворе кислоты или щелочи в течении 1-3 суток.
К ковалентным связям в молекуле белка помимо пептидной, относится также ДИСУЛЬФИДНАЯ СВЯЗЬ.
Цистеин — аминокислота, которая в радикале имеет SH-группу, за счет которой и образуются дисульфидные связи.

Дисульфидная связь — это ковалентная связь. Однако биологически она гораздо менее устойчива, чем пептидная связь. Это объясняется тем, что в организме интенсивно протекают окислительно-восстановительные процессы. Дисульфидная связь может возникать между разными участками одной и той же полипептидной цепи, тогда она удерживает эту цепь в изогнутом состоянии. Если дисульфидная связь возникает между двумя полипептидами, то она объединяет их в одну молекулу.
СЛАБЫЕ ТИПЫ СВЯЗЕЙ
В десятки раз слабее ковалентных связей. Это не определенные типы связей, а неспецифическое взаимодействие, которое возникает между разными химическими группировками, имеющими высокое сродство друг к другу (сродство – это способность к взаимодействию). Например: противоположно заряженные радикалы.
Таким образом, слабые типы связей — это физико-химические взаимодействия. Поэтому они очень чувствительны к изменениям условий среды (температуры, pH среды, ионной силы раствора и так далее).
ВОДОРОДНАЯ СВЯЗЬ — это связь, возникающая между двумя электроотрицательными атомами за счет атома водорода, который соединен с одним из электроотрицательных атомов ковалентно (см. рисунок).

Водородная связь примерно в 10 раз слабее, чем ковалентная. Если водородные связи повторяются многократно, то они удерживают полипептидные цепочки с высокой прочностью. Водородные связи очень чувствительны к условиям внешней среды и присутствию в ней веществ, которые сами способны образовывать такие связи (например, мочевина).
ИОННАЯ СВЯЗЬ — возникает между положительно и отрицательно заряженными группировками (дополнительные карбоксильные и аминогруппы), которые встречаются в радикалах лизина, аргинина, гистидина, аспарагиновой и глутаминовой кислот.

ГИДРОФОБНОЕ ВЗАИМОДЕЙСТВИЕ — неспецифическое притяжение, возникающее в молекуле белка между радикалами гидрофобных аминокислот — вызывается силами Ван-дер-Ваальса и дополняется выталкивающей силой воды. Гидрофобное взаимодействие ослабевает или разрывается в присутствии различных органических растворителей и некоторых детергентов. Например, некоторые последствия действия этилового спирта при проникновении его внутрь организма обусловлены тем, что под его влиянием ослабляются гидрофобные взаимодействия в молекулах белков.
ПРОСТРАНСТВЕННАЯ ОРГАНИЗАЦИЯ БЕЛКОВОЙ МОЛЕКУЛЫ
В основе каждого белка лежит полипептидная цепь. Она не просто вытянута в пространстве, а организована в трехмерную структуру. Поэтому существует понятие о 4-х уровнях пространственной организации белка, а именно — первичной, вторичной, третичной и четвертичной структурах белковых молекул.
ПЕРВИЧНАЯ СТРУКТУРА
Первичная структура белка — последовательность аминокислотных фрагментов, прочно (и в течение всего периода существования белка) соединенных пептидными связями. Существует период полужизни белковых молекул — для большинства белков около 2-х недель. Если произошел разрыв хотя бы одной пептидной связи, то образуется уже другой белок.
ВТОРИЧНАЯ СТРУКТУРА
Вторичная структура — это пространственная организация стержня полипептидной цепи. Существуют 3 главнейших типа вторичной структуры:
1) Альфа-спираль — имеет определенные характеристики: ширину, расстояние между двумя витками спирали. Для белков характерна правозакрученная спираль. В этой спирали на 10 витков приходится 36 аминокислотных остатков. У всех пептидов, уложенных в такую спираль, эта спираль абсолютно одинакова. Фиксируется альфа-спираль с помощью водородных связей между NH-группами одного витка спирали и С=О группами соседнего витка. Эти водородные связи расположены параллельно оси спирали и многократно повторяются, поэтому прочно удерживают спиралеобразную структуру. Более того, удерживают в несколько напряженном состоянии (как сжатую пружину).


Бета-складчатая структура — или структура складчатого листа. Фиксируется также водородными связями между С=О и NH-группами. Фиксирует два участка полипептидной цепи. Эти цепи могут быть параллельны или антипараллельны. Если такие связи образуются в пределах одного пептида, то они всегда антипараллельны, а если между разными полипептидами, то параллельны.
3) Нерегулярная структура — тип вторичной структуры, в котором расположение различных участков полипептидной цепи относительно друг друга не имеет регулярного (постоянного) характера, поэтому нерегулярные структуры могут иметь различную конформацию.
ТРЕТИЧНАЯ СТРУКТУРА
Это трехмерная архитектура полипептидной цепи – особое взаимное расположение в пространстве спиралеобразных, складчатых и нерегулярных участков полипептидной цепи. У разных белков третичной структуры различна. В формировании третичной структуры участвуют дисульфидные связи и все слабые типы связей.
Выделяют два общих типа третичной структуры:
1) В фибриллярных белках (например, коллаген, эластин ) молекулы которых имеют вытянутую форму и обычно формируют волокнистые структуры тканей, третичная структура представлена либотройной альфа-спиралью (например, в коллагене), либо бета-складчатыми структурами.
2) В глобулярных белках , молекулы которых имеют форму шара или эллипса (латинское название: GLOBULA — шар), встречается сочетание всех трех типов структур: всегда есть нерегулярные участки, есть бета-складчатые структуры и альфа-спирали.
Обычно в глобулярных белках гидрофобные участки молекулы находятся в глубине молекулы. Соединяясь между собой, гидрофобные радикалы образуют гидрофобные кластеры (центры). Формирование гидрофобного кластера вынуждает молекулу соответствующим образом изгибаться в пространстве. Обычно в молекуле глобулярного белка бывает несколько гидрофобных кластеров в глубине молекулы. Это является проявлением двойственности свойств белковой молекулы: на поверхности молекулы — гидрофильные группировки, поэтому молекула в целом — гидрофильная, а в глубине молекулы — спрятаны гидрофобные радикалы.
ЧЕТВЕРТИЧНАЯ СТРУКТУРА
Встречается не у всех белков, а только у тех, которые состоят из двух или более полипептидных цепей. Каждая такая цепь называется СУБЪЕДИНИЦЕЙ данной молекулы (или ПРОТОМЕРОМ ). Поэтому белки, обладающие четвертичной структурой, называют ОЛИГОМЕРНЫМИ белками. В состав белковой молекулы могут входить одинаковые или разные субъединицы. Например, молекула гемоглобина «А» состоит из двух субъединиц одного типа и двух субъединиц другого типа, то есть является тетрамером. Фиксируются четвертичные структуры белков всеми типами слабых связей, а иногда еще и дисульфидными связями.
МЕТОДЫ ОПРЕДЕЛЕНИЯ ПЕРВИЧНОЙ СТРУКТУРЫ БЕЛКА
1)Деградация по Эдмону
К раствору белка добавляют реактив Эдмона, содержащий фенилизотиоцианат.
Фенилизотиоцианат взаимодействует с альфа-аминогруппой первой (N-концевой) аминокислоты, а затем происходит ее отщепление от полипептидной цепи путем гидролиза:

После этого идентифицируют первую аминокислоту. Затем процесс повторяется.
В настоящее время процесс автоматизирован.
2) Секвенирование ДНК
Первичная структура любой белковой молекулы напрямую зависит от структуры ДНК-генома. Поэтому сначала выделяют ген, в котором закодирована структура белка. Далее определяют последовательность азотистых оснований в ДНК. Каждая аминокислота в белковой молекуле закодирована сочетанием трех азотистых оснований — триплетом (кодоном) в молекуле ДНК. Например, сочетание трех оснований аденина (ААА) кодирует аминокислоту фенилаланин, а последовательность из трех оснований цитозина – глицин. Это дает возможность получить информацию о первичной структуре белковой молекуле, а, значит, прогнозировать строение всей молекулы в целом, поскольку именно первичная структура определяет строение всех высших уровней организации – и вторичной, и третичной, а, иногда и четвертичной структур.
Для проверки предположений о строении высших структур используется еще один метод:
3) Рентгеноструктурный анализ
Схема, поясняющая принцип этого метода, представлена на рисунке:

В результате облучения на фотопленке фиксируется карта электронной плотности (похожа на географическую карту). Далее производится компьютерный анализ полученного изображения, в результате чего строится пространственная модель белковой молекулы.
Электронная микроскопия
Может быть использована для выяснения структуры белковых молекул с большой молекулярной массой – от 500.000 до 1.000.000 Да (дальтон). Дальтон (Да) и килодальтон (кДа) – единицы измерения массы белков. 1кДа=103 Да. 1 дальтон равен 1/16 массы атома кислорода (кислородная единица массы).
КОНФИГУРАЦИЯ И КОНФОРМАЦИЯ БЕЛКОВОЙ МОЛЕКУЛЫ
Из всего сказанного можно заключить, что пространственная организация белков очень сложна. В химии существует понятие — пространственная КОНФИГУРАЦИЯ — жестко закрепленное ковалентными связями пространственное взаимное расположение частей молекулы (например: принадлежность к L-ряду стереоизомеров или к D-ряду).
Для белков также используется понятие КОНФОРМАЦИЯ белковой молекулы — определенное, но не застывшее, не неизменное взаимное расположение частей молекулы. Так как конформация белковой молекулы формируется при участии слабых типов связей, то она является подвижной (способной к изменениям), и белок может изменять свою структуру. В зависимости от условий внешней среды молекула может существовать в разных конформационных состояниях, которые легко переходят друг в друга. Энергетически выгодными для реальных условий являются только одно или несколько конформационных состояний, между которыми существует равновесие. Переходы из одного конформационного состояния в другое обеспечивают функционирование белковой молекулы. Это обратимые конформационные изменения (встречаются в организме, например, при проведении нервного импульса, при переносе кислорода гемоглобином). При изменении конформации часть слабых связей разрушается, и образуются новые связи слабого типа.
ЛИГАНДЫ
Взаимодействие белка с каким-нибудь веществом иногда приводит к связыванию молекулы этого вещества молекулой белка. Этот явление известно как «сорбция» (связывание). Обратный же процесс — освобождение другой молекулы от белковой называется «десорбция» .
Если для какой-нибудь пары молекул процесс сорбции преобладает над десорбцией, то это уже специфическая сорбция, а вещество, которое сорбируется, называется «лиганд» .
Виды лигандов:
1) Лиганд белка-фермента – субстрат.
2) Лиганд траспортного белка – транспортируемое вещество.
3) Лиганд антитела (иммуноглобулина) – антиген.
4) Лиганд рецептора гормона или нейромедиатора – гормон или нейромедиатор.
Белок может изменять свою конформацию не только при взаимодействии с лигандом, но и в результате любого химического взаимодействия. Примером такого взаимодействия может служить присоединение остатка фосфорной кислоты.
В природных условиях белки имеют несколько термодинамически выгодных конформационных состояний. Это нативные состояния (природные). Natura (лат.) – природа.
НАТИВНОСТЬ БЕЛКОВОЙ МОЛЕКУЛЫ
НАТИВНОСТЬ — это уникальный комплекс физических, физико-химических, химических и биологических свойств белковой молекулы, который принадлежит ей, когда молекула белка находится в естественном, природном (нативном) состоянии.
Например: белок хрусталика глаза — кристаллин — обладает высокой прозрачностью только в нативном состоянии).
ДЕНАТУРАЦИЯ БЕЛКА
Для обозначения процесса, при котором нативные свойства белка теряются, используют термин ДЕНАТУРАЦИЯ.
ДЕНАТУРАЦИЯ — это лишение белка его природных, нативных свойств, сопровождающееся разрушением четвертичной (если она была), третичной, а иногда и вторичной структуры белковой молекулы, которое возникает при разрушении дисульфидных и слабых типов связей, участвующих в образовании этих структур. Первичная структура при этом сохраняется, потому что она сформирована прочными ковалентными связями. Разрушение первичной структуры может произойти только в результате гидролиза белковой молекулы длительным кипячением в растворе кислоты или щелочи.
ФАКТОРЫ, ВЫЗЫВАЮЩИЕ ДЕНАТУРАЦИЮ БЕЛКОВ
Факторы, которые вызывают денатурацию белков, можно разделить на физические и химические .
Физические факторы
1. Высокие температуры. Для разных белков характерна различная чувствительность к тепловому воздействию. Часть белков подвергается денатурации уже при 40-500С. Такие белки называют термолабильными. Другие белки денатурируют при гораздо более высоких температурах, они являются термостабильными .
2. Ультрафиолетовое облучение
3. Рентгеновское и радиоактивное облучение
4. Ультразвук
5. Механическое воздействие (например, вибрация).
Химические факторы
1. Концентрированные кислоты и щелочи. Например, трихлоруксусная кислота (органическая), азотная кислота (неорганическая).
2. Соли тяжелых металлов (например, CuSO4 ).
3. Органические растворители (этиловый спирт, ацетон)
4. Растительные алкалоиды.
5. Мочевина в высоких концентрациях

5. Другие вещества, способные нарушать слабые типы связей в молекулах белков.
Воздействие факторами денатурации применяют для стерилизации оборудования и инструментов, а также как антисептики.
Обратимость денатурации
В пробирке (in vitro) чаще всего это – необратимый процесс. Если же денатурированный белок поместить в условия, близкие к нативным, то он может ренатурировать, но очень медленно, и такое явление характерно не для всех белков.
In vivo, в организме, возможна быстрая ренатурация. Это связано с выработкой в живом организме специфических белков, которые «узнают» структуру денатурированного белка, присоединяются к нему с помощью слабых типов связи и создают оптимальные условия для ренатурации. Такие специфические белки известны как «белки теплового шока » или «белки стресса ».
Белки стресса
Существует несколько семейств этих белков, они отличаются по молекулярной массе.
Например, известен белок hsp 70 – heatshock protein массой 70 kDa.
Такие белки есть во всех клетках организма. Они выполняют также функцию траспорта полипептидных цепей через биологические мембраны и участвуют в формировании третичной и четвертичной структур белковых молекул. Перечисленные функции белков стресса называются шаперонными. При различных видах стресса происходит индукция синтеза таких белков: при перегреве организма (40-440С), при вирусных заболеваниях, отравлениях солями тяжелых металлов, этанолом и др.
В организме южных народов установлено повышенное содержание белков стресса, по сравнению с северной расой.
Молекула белка теплового шока состоит из двух компактных глобул, соединенных свободной цепью:

Разные белки теплового шока имеют общий план построения. Все они содержат контактные домены.
Разные белки с различными функциями могут содержать одинаковые домены. Например, различные кальций-связывающие белки имеют одинаковый для всех них домен, отвечающий за связывание Ca+2 .
Роль доменной структуры заключается в том, что она предоставляет белку большие возможности для выполнения своей функции благодаря перемещениям одного домена по отношению к другому. Участки соединения двух доменов – самое слабое в структурном отношении место в молекуле таких белков. Именно здесь чаще всего происходит гидролиз связей, и белок разрушается.
ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА БЕЛКОВ. РАСТВОРИМОСТЬ БЕЛКОВ В ВОДЕ
Большинство белков гидрофильны. Однако белковые молекулы имеют очень большие размеры, поэтому белки не могут образовывать истинных растворов, а только коллоидные. Внешнее проявление этого — это эффект Тиндаля (или конус Тиндаля). Эффект Тиндаля вызывается рассеянием тонкого пучка света при прохождении через белковый раствор. Несмотря на большую величину, многие белковые молекулы не осаждаются в водных растворах. Осаждению белковых молекул препятствуют факторы стабилизации белкового раствора.
ФАКТОРЫ СТАБИЛИЗАЦИИ БЕЛКА В РАСТВОРЕ
ГИДРАТНАЯ ОБОЛОЧКА — это слой молекул воды, определенным образом ориентированных на поверхности белковой молекулы. Поверхность большинства белковых молекул заряжена отрицательно, и диполи молекул воды притягиваются к ней своими положительно заряженными полюсами (смотрите рисунок).
Чем больше гидрофильных свойств у белковой молекулы, чем больше в ее составе и на ее поверхности аминокислот с полярными (гидрофильными) радикалами, тем сильнее выражена и прочнее удерживается гидратная оболочка и тем больше в ней слоев. Вода гидратной оболочки обладает особыми свойствами: она не является свободной, а связана с белковой молекулой. Это — “связанная” вода. Она принадлежит белку, и поэтому имеет особые свойства.

Свойства воды гидратной оболочки
а) Температура кипения выше 1000С.
б) Температура замерзания ниже 0О С.
в) В воде гидратной оболочки не растворяются различные соли и другие гидрофильные вещества.
г) Окружая каждую молекулу белка, гидратная оболочка не дает этим белковым молекулам сблизиться, соединиться и выпасть в осадок.
2) ЗАРЯД БЕЛКОВОЙ МОЛЕКУЛЫ. Поверхность большинства белковых молекул заряжена потому, что в каждой молекуле белка есть свободные заряженные СОО- и Nh4+ группы. Изоэлектрическая точка (ИЭТ) большинства белков организма находится в слабокислой среде. Это означает, что у таких белков количество кислотных (СООН) групп больше количества основных групп (Nh4 ). рН плазмы крови около 7,36 — это выше ИЭТ большинства белков, поэтому в плазме крови белки имеют отрицательный заряд.
СПОСОБЫ ОСАЖДЕНИЯ БЕЛКОВ
Делятся на две группы:
1) Способы осаждения нативного белка
2)Способы осаждения денатурированного белка
Чтобы осадить белок из раствора, надо лишить его обоих факторов стабилизации: и заряда, и гидратной оболочки.
ОСАЖДЕНИЕ НАТИВНЫХ БЕЛКОВ
Чтобы сохранить нативность белковой молекулы, ее заряд можно устранить только одним способом: приблизить рН среды к изоэлектрической точке белка (ИЭТ), а для большинства белков нашего организма ИЭТ находится в слабокислой среде. Другой фактор стабилизации — гидратную оболочку можно устранить разными способами.
Наиболее типичным примером осаждения нативного белка является ВЫСАЛИВАНИЕ.
а) ВЫСАЛИВАНИЕ — это осаждение белков высокими концентрациями нейтральных солей щелочных и щелочноземельных металлов, поскольку такие соли очень гидрофильны и обладают в высоких концентрациях водоотнимающими свойствами. Чаще это NaCl, Na2 SO4, (Nh5 )2 SO4, CaCl2. По мере добавления таких солей к раствору белка они сначала растворяюся в свободной воде, а затем, при дальнейшем повышении концентрации соли, конкурируют с белком за обладание водой, которая входит в состав гидратных оболочек. Белки менее гидрофильные, которые плохо удерживают воду гидратной оболочки, теряют ее раньше. Более гидрофильные белки требуют большей концентрации соли для высаливания. Поэтому с помощью высаливания можно разделить белки с разной степенью гидрофильности. Таким способом, например, можно разделить альбумины и глобулины плазмы крови.
При высаливании сохраняется нативность белковых молекул. Если осадить белки с помощью высаливания, а затем уменьшить концентрацию солей, например, методом диализа, то белок опять растворится .
Осаждения белков без потери ими нативности можно достичь также с помощью водоотнимающих средств.
б) ПРИМЕНЕНИЕ ВОДООТНИМАЮЩИХ СРЕДСТВ . Такими средствами являются растворители, которые смешиваются с водой в любых соотношениях. Чаще всего это ацетон, этиловый спирт. Эти вещества отнимают гидратные оболочки белков, и белки выпадают в осадок, если они лишены заряда. Но, в отличие от высаливания, осадок сразу (немедленно!) должен быть отделен от растворителя. Если растворитель и белок будут длительно находиться в контакте, то могут произойти необратимые изменения структуры белковой молекулы (денатурация).
ОСАЖДЕНИЕ ДЕНАТУРИРОВАННЫХ БЕЛКОВ
а) ДЕЙСТВИЕ СОЛЕЙ ТЯЖЕЛЫХ МЕТАЛЛОВ . Образуют соединения с SH-группами белков. Ядовиты для человека и животных. В медицинской практике применяются способы детоксикации при отравлениях тяжелыми металлами. В этих случаях для обезвреживания этих металлов дают внутрь молоко или другие белковые растворы.
б) КИПЯЧЕНИЕ (или просто нагревание до высоких температур) — усиливается тепловое движение молекул, ослабляются слабые типы связей, теряется нативность, белковая молекула “разворачивается”, гидрофобные структуры выходят наружу. Это приводит к потере гидратной оболочки, молекулы сближаются и взаимодействуют друг с другом. Это приводит к тому, что белок выпадает в осадок. При охлаждении нативность не восстанавливается.
При кипячении белок не всегда выпадает в осадок. Если нагревать белок в любой среде (сильно кислой, сильно щелочной или нейтральной средах), то денатурация белка происходит обязательно, белковые молекулы теряют гидратную оболочку. Но в сильно кислой или в сильно щелочной средах молекулы белка в осадок не выпадают, потому что у них остается один из факторов стабилизации — заряд. Сохранение заряда не позволяет молекулам белка сблизиться друг с другом — агрегация полипептидных цепей не происходит. Даже если раствор белка охладить — осадок все равно не выпадает — это будет коллоидный раствор денатурированного белка. Если приблизить затем рН среды к изоэлектрической точке белка (например, добавлением кислоты или щелочи), то белок выпадет в осадок, потому что будет лишен обоих факторов стабильности в растворе — и заряда, и гидратной оболочки.
www.ronl.ru
Реферат - Белки - Химия
БЕЛКИ
Любое соединение, которое содержитодновременно карбоксильную и аминогруппу, является аминокислотой.Однако, чаще этот термин применяется для обозначения карбоновых кислот,аминогруппа которых находится в a-положении к карбоксильной группе.
/>
Аминокислоты, как правило, входят всостав полимеров — белков. В природе встречается свыше 70 аминокислот,но только 20 играют важную роль в живых организмах. Незаменимыминазываются аминокислоты, которые не могут быть синтезированы организмом извеществ, поступающих с пищей, в количествах, достаточных для того, чтобыудовлетворить физиологические потребности организма. Незаменимые аминокислотыприводятся в табл. 1. Для больных фенилкетонурией незаменимой аминокислотойявляется также тирозин (см. табл. 1).
Таблица 1.Незаменимыеаминокислоты R-CHNh3COOH
Название (сокращение) R изолейцин (ile, ileu)Ch4Ch3CH(CH)3-
лейцин (leu)(Ch4)2CHCh3-
лизин (lys)Nh3Ch3Ch3Ch3Ch3-
метионин (met)Ch4SCh3Ch3-
фенилаланин (phe)C6H5Ch3-
треонин (thr)Ch4CH(OH)-
триптофан (try)/>
валин (val)(Ch4)2CH-
тирозин (tyr)Аминокислоты называют обычно какзамещенные соответствующих карбоновых кислот, обозначая положение аминогруппыбуквами греческого алфавита. Для простейших аминокислот обычно применяютсятривиальные названия (глицин, аланин, изолейцин и т.д.). Изомерия аминокислотсвязана с расположением функциональных групп и со строением углеводородногоскелета. Молекула аминокислоты моет содержать одну или несколько карбоксильныхгрупп и в соответствии с этим аминокислоты различаются по основности. Также вмолекуле аминокислоты может находиться разное количество аминогрупп.
Аминокислоты способны кполиконденсации, в результате которой образуется полиамид. Полиамиды, состоящиеиз a-аминокислот, называются пептидамиили полипептидами. Амидная связь в таких полимерах называется пептиднойсвязью. Полипептиды с молекулярной массой не меньше 5000 называют белками.В состав белков входит около 25 различных аминокислот. При гидролизе данногобелка могут образовываться все эти аминокислоты или некоторые из них вопределенных пропорциях, характерных для отдельного белка.
Уникальная последовательностьаминокислотных остатков в цепи, присущая данному белку, называется первичнойструктурой белка. Особенности скручивания цепей белковых молекул(взаимное расположение фрагментов в пространстве) называются вторичнойструктурой белков. Полипептидные цепи белков могут соединяться междусобой с образованием амидных, дисульфидных, водородных и иных связей за счетбоковых цепей аминокислот. В результате этого происходит закручивание спирали вклубок. Эта особенность строения называется третичной структурой белка.Для проявления биологической активности некоторые белки должны сначалаобразовать макрокомплекс (олигопротеин), состоящий из несколькихполноценных белковых субъединиц. Четвертичная структураопределяет степень ассоциации таких мономеров в биологически активномматериале.
Белки делятся на две большие группы — фибриллярные (отношение длины молекулы к ширине больше 10) и глобулярные(отношение меньше 10). К фибриллярным белкам относится коллаген,наиболее распространенный белок позвоночных; на его долю приходится почти 50%сухого веса хрящей и около 30% твердого вещества кости. В большинстверегуляторных систем растений и животных катализ осуществляется глобулярнымибелками, которые носят название ферментов.
Для полипептидов существуетспециальная номенклатура. Аминокислота со свободной аминогруппой на концеполипептидной цепи называется N-концевой аминокислотой, а аминокислота со свободной карбоксильной группой надругом конце этой молекулы — С-концевой аминокислотой.Аминокислотные остатки в полипептидной цепи нумеруются по порядку, начиная с N-концевой аминокислоты, котораяобозначается цифрой 1. Полипептиды называются как производные С-концевойаминокислоты, причем первой пишут название N-концевой аминокислоты, а последней — С-концевуюаминокислоту. Так, глицилаланин — это дипептид, состоящий из глицина (N-концевая аминокислота) и аланина(С-концевая аминокислота). В сокращенном виде название пишут gly-ala. Аланилглицин (ala-gly) — это изомерный дипептид, в котором N-концевой аминокислотой является аланин, а С-концевой — глицин.
АНАЛИЗБЕЛКОВ
Белки, как и прочие амидыгидролизуются под действием кислот и щелочей. После полного гидролизаможно установить качественный и количественный аминокислотный состав, но неточную последовательность аминокислот. Если перед гидролизом обработатьполипептид реактивом Сэнгера, то можно будет затем идентифицировать N-концевую аминокислоту, так как онадаст устойчивое окрашенное производное анилина, которое не разрушается пригидролизе.
/>
В результате неполногогидролиза образуются небольшие пептидные фрагменты. Меняя условиягидролиза, можно разбивать полипептид на различные фрагменты, которыеперекрываются по составляющим их аминокислотным остаткам. Для воссоздания первичнойструктуры необходимо идентифицировать аминокислоты, которые входят в составкаждого из фрагментов, и определить последовательность, в которой этиаминокислоты соединяются друг с другом в исходном полипептиде. Для этого можнопровести полный гидролиз фрагментов, идентифицировать аминокислоты, а затемосуществить химический синтез фрагментов. Существует и другой путь — избирательныйгидролиз. В этом случае от фрагмента отщепляют по одной аминокислоте накаждом этапе, чаще всего при помощи ферментов поджелудочной железы (карбоксипептидаз).Эти ферменты способны гидролизовать только С-концевые аминокислоты и,следовательно, разрушать пептидный фрагмент с С-конца.
N-концевую кислоту можно удалить припомощи фенилизотиоцианата. В результате следующая аминокислота становится N-концевой, ее также можно отщепитьдействием фенилизотиоцианата и т.д. (деградация по Эдману).
/>
В результате реакциифенилизотиоцианата с N-концевойаминокислотой образуется аддукт (тиомочевина), который циклизуется в уксуснойкислоте. При кислотном гидролизе аддукт расщепляется, давая N-концевую аминокислоту и неустойчивоепромежуточное соединение, которое изомеризуется в тиогидантоин. Специфическийтиогидантоин служит для идентификации первой N-концевой аминокислоты. Проводя во второй раз всюпоследовательность реакций, можно определить следующую аминокислоту, так какона стала N-концевой в результате деградации поЭдману.
СИНТЕЗБЕЛКОВ
Реакции, протекающие при синтезе белка- см. Реакции аминокислот. Вышеприведенная схема допускает некоторые изменения- например, для активации карбоксильной группы вместо этилхлорформиатаиспользуют дициклогексилкарбодиимид. В любом случае, синтез белка по этой схеметребует нескольких стадий, что и служит ограничением использования этогометода.
Для синтеза белков стали применятьметод, впервые предложенный Р. Меррифилдом и получивший название твердофазногосинтеза пептидов (ТФСП). На первой стадии синтеза аминокислоту,которая будет стоять на С-конце будущего белка, присоединяют через еекарбоксильную группу к полимеру. При этом нередко используют образованиеэфирной связи между этой группой и хлорметильной группой полимера:
/>
Затем проводят ацилирование свободнойаминогруппы этой аминокислоты путем реакции с дициклогексилкарбодиимидом идругой аминокислотой, которая содержит защищенную аминогруппу. В результатеобразуется первая пептидная связь:
/>
После удаления трет-бутоксикарбонильнойгруппы и нейтрализации полимер, несущий пептид, готов для следующей стадииацилирования:
/>
Вслед за присоединением последнегоаминокислотного остатка аддукт полипептида и полимера обрабатывают смесьюбромистого водорода и трифторуксусной кислоты. В результате полипептидосвобождается от полимера, а с N-концевойаминокислоты снимается защитная группа.
Преимущество этого метода посравнению с классическими методами синтеза состоит в том, что ни на одной изстадий не требуется выделения полипептидной цепи. В силу чрезвычайно низкойрастворимости аддукта пептида и полимера легко отмывается после каждой реакцииот побочных продуктов, растворителей и избытка реагентов без потери пептида,после чего аддукт готов к следующей реакции. В настоящее время методавтоматизирован, и запрограммированные аминокислотные синтезаторы могутбез труда присоединить шесть аминокислот к растущей полипептидной цепи за 24 ч.Эти приборы добавляют реактивы в надлежащей последовательности, меняют условияреакций, обеспечивают необходимое время реакций, отмывают побочные продукты,после чего начинают операцию снова. С помощью метода ТФСП былисинтезированы инсулин и фермент рибонуклеаза, состоящий из124 аминокислот.
www.ronl.ru
Реферат - Белки, их строение и состав
|
оглавление
ОГЛАВЛЕНИЕ_________________________________________________________ 1
БЕЛКИ________________________________________________________________ 3
СТРУКТУРА БЕЛКОВ__________________________________________________ 5
ФИЗИЧЕСКИЕ СВОЙСТВА_____________________________________________ 7
ХИМИЧЕСКИЕ СВОЙСТВА____________________________________________ 7
СИНТЕЗ БЕЛКОВ______________________________________________________ 8
Литература: ___________________________________________________________ 8
белки
ЕЛКИ — это азотсодержащие высокомолекулярные органические вещества со сложным составом и строением молекул.
Белок можно рассматривать как сложный полимер аминокислот.
Белки входят в состав всех живых организмов, но особо важную роль они играют в животных организмах, которые состоят из тех или иных форм белков (мышцы, покровные ткани, внутренние органы, хрящи, кровь).
Растения синтезируют белки (и их составные части a-аминокислоты) из углекислого газа СО2 и воды Н2 О за счет фотосинтеза, усваивая остальные элементы белков (азот N, фосфор Р, серу S, железо Fe, магний Mg) из растворимых солей, находящихся в почве.
Животные организмы в основном получают готовые аминокислоты с пищей и на их базе строят белки своей организма. Ряд аминокислот (заменимые аминокислоты) могут синтезироваться непосредственно животными организмами.
Характерной особенностью белков является их многообразие, связанное с количеством, свойствами и способах соединения входящих в их молекулу аминокислот. Белки выполняют функцию биокатализаторов — ферментов, регулирующих скорость и направление химических реакций в организме. В комплексе с нуклеиновыми кислотами обеспечивают функции роста и передачи наследственных признаков, являются структурной основой мышц и осуществляют мышечное сокращение.
В молекулах белков содержатся повторяющиеся амидные связи С(0)—NH, названные пептидными (теория русского биохимика А.Я.Данилевского).
Таким образом, белок представляет собой полипептид, содержащий сотни или тысячи аминокислотных звеньев.
Структура белков
 |
собый характер белка каждого вида связан не только с длиной, составом и строением входящих в его молекулу полипептидных цепей, но и с тем, как эти цепи ориентируются.
В структуре любого белка существует несколько степеней организации:
Первичная структура белка — специфическая последовательность аминокислот в полипептидной цепи.
|

|

|
|
дисульфидный мостик (-S-S-) между атомами серы,
сложноэфирный мостик – между карбоксильной группой (-СО-) и гидроксильной (-ОН),
солевой мостик — между карбоксильной (-СО-) и аминогруппами (Nh3 ).
|

Четвертичная структура белка — тип взаимодействия между несколькими полипептидными цепями.
|
Физические свойства
елки имеют большую молекулярную массу ( 104 —107 ), многие белки растворимы в воде, но образуют, как правило, коллоидные растворы, из которых выпадают при увеличении концентрации неорганических солей, добавлении солей тяжелых металлов, органических растворителей или при нагревании (денатурация).
Химические свойства
Денатурация — разрушение вторичной и третичной структуры белка.
Качественные реакции на белок:
биуретовая реакция: фиолетовое окрашивание при обработке солями меди в щелочной среде (дают все белки),
ксантопротеиновая реакция: желтое окрашивание при действии концентрированной азотной кислоты, переходящее в оранжевое под действием аммиака (дают не все белки),
выпадение черного осадка (содержащего серу) при добавлении ацетата свинца (II), гидроксида натрия и нагревании.
Гидролиз белков — при нагревании в щелочном или кислом растворе с образованием аминокислот.
Синтез белков
Белок — сложная молекула, и синтез его представляется трудной задачей. В настоящее время разработано много методов прекращения [ГМВ1] a-аминокислот в пептиды и синтезированы простейшие природные белки — инсулин, рибонуклеаза и др.
Большая заслуга в создании микробиологической промышленности по производству искусственных пищевых продуктов принадлежит советскому ученому А.Н.Несмеянову.
Литература:
Справочник школьника
“ХИМИЯ” М.,”СЛОВО” 1995.
Г.Е.Рудзитис, Ф.Г.Фельдман
“Химия 11. Органическая химия”
М., “Просвещение”,1993.
А.И.Артеменко, И.В. Тикунова
“Химия 10-11. Органическая химия”
М., “Просвещение” 1993.
[ГМВ1]
www.ronl.ru
Реферат - Белки - Химия
Белковые вещества составляют громадный класс органических, то есть углеродистых, а именно углеродисто азотистых соединений, неизбежно встречаемых в каждом организме. Роль белков в организме огромна. Прежде всего необходимо сказать об обмене белков в организме.
В процессах обмена веществ между внешней средой и организмом ведущее место занимает обмен белков. Белки поступают в организм человека и животных с различными пищевыми продуктами, в которых содержание белка колеблется в широких пределах. Приведем таблицу, дающую представление о содержании белка в некоторых продуктах питания.
Название продукта | содержание белка | Название продукта | содержание белка |
| мясо | 18-22% | горох | 26% |
| рыба | 17-20% | картофель | 1,5-2% |
| сыр | 20-36% | ржаной хлеб | 7,8% |
| яйца | 13% | яблоки | 0,3-0,4% |
| молоко | 3,5% | капуста | 1,1-1,6% |
| рис | 8% | морковь | 0,8-1% |
| свекла | 1,6% | макароны | 9-13% |
| пшено | 10% | гречневая крупа | 11% |
Белки, распадаясь в организме, являются, так же как углеводы и жиры, источником энергии. Энергия, получаемая при распаде белков, может быть без всякого ущерба для организма компенсирована энергией распада жиров и углеводов. Однако очень важно, что организм человека и животных не может обходиться без регулярного поступления белков извне.
Опыт показывает, что даже довольно длительное выключение жиров или углеводов из питания животного не вызывает тяжелых расстройств здоровья. Но прием в течение нескольких дней пищи, не содержащей белков, приводит к серьезным нарушениям, а продолжительное безбелковое питание неизбежно кончается смертью животного. Все это имеет место даже при обильном питании углеводами и жирами.
Отсюда очевидно, что роль и значение белков в процессах обмена веществ отнюдь не исчерпывается их энергетической ценностью.
Действительно, в процессах жизнедеятельности организма на первый план выступают совсем другие, несравненно более важные специфические свойства и функции белков.
По степени важности в процессах обмена веществ пластическая роль белков неизмеримо превосходит их как источника энергии. Более того, пластическая функция белков не только велика, но и незаменима, так как белки в этом отношении нельзя заменить ни жирами, ни углеводами, ни какими-либо другими веществами, входящими в состав живой материи или поступающими в организм из внешней среды.
Без белков или их составных частей – аминокислот – не может быть обеспечено воспроизводство основных структурных элементов органов и тканей, а также образование ряда важнейших веществ, как, например, ферментов и гормонов.
Какое же количество белка необходимо в питании человека, чтобы обеспечить сохранение его здоровья и работоспособности? «Коэффициент изнашивания» у взрослого человека составляет около 23 граммов белка. Казалось бы, нужно 23 грамма белка, чтобы покрыть расход белка при распаде его в органах и тканях. Однако оказалось, что азотистое равновесие устанавливается при приеме более высоких количеств белка в пище, чем того требует так называемый «коэффициент изнашивания».
Разные исследователи получали различные величины содержания белка в пище, при котором поддерживается азотистое равновесие. Эти величины колеблются в зависимости от состава пищи и от того, какие белковые продукты принимаются. Но в среднем азотистое равновесие устанавливается у человека при потреблении 30-45 граммов белка в сутки. Этот минимум белка, необходимый для того, чтобы поддерживать азотистое равновесие на рационе, полностью покрывающем энергетические потребности организма получил название «физиологического минимума белка». Азотистое равновесие у человека и животных, таким образом, возможно получить при приеме с пищей белка в количестве примерно вдвое большем, чем это необходимо по «коэффициенту изнашивания».
Содержание белка в пищевом рационе взрослого человека,
необходимое для поддержания азотистого равновесия.
Источники белка | Потребность в белках (г/сутки) | Источники белка | Потребность в белках (г/сутки) |
| яичный белок | 19,9 | картофель | 30,0 |
| говядина | 26,0 | хлеб из пшеничной муки | 67,0 |
| молоко | 27,6 |
Ученые пришли к выводу, что взрослый человек должен потреблять ежедневно при трате энергии в 1500 ккал не менее 100 грамм, а в жарком климате – не менее 120 грамм белка. Эти нормы соответствуют умственному труду или труду физическому, полностью механизированному. При расходовании большего количества энергии, то есть при физическом труде, недостаточно механизированном, необходимо добавочно 10 грамм белка на каждые 500 ккал. Таким образом, при физическом труде с тратой энергии в 4000 ккал требуется 130-150 грамм белка в сутки.
Растущий организм испытывает потребность в белке в зависимости от возраста. Нормы белка для детей приведены в следующей таблице:
| Возраст в годах | Количество белка |
| 1 – 3 года | 55г |
| 4 – 6 лет | 72г |
| 7 – 9 лет | 89г |
| 10 – 12 лет | 100г |
Для удовлетворения потребностей организма существенным является не только количество, но и качество белков в пище. Различные белки отличаются друг от друга процентным содержанием аминокислот. В зависимости от аминокислотного состава, организму требуется одного белка больше, а другого меньше. В этом смысле можно говорить о различной биологической ценности белков. Биологическая ценность белка определяется также степенью усвоения его организмом. Белки, находящиеся в продуктах питания, потребляемых человеком, содержат в тех или иных количествах все аминокислоты.
В тесной связи с вопросом о биологической ценности белка находятся представления о так называемых незаменимых аминокислотах. Изучение азотистого обмена у взрослых людей позволило сделать вывод, что для удовлетворительного самочувствия необходимо восемь незаменимых аминокислот и источники азота.
Необходимо особо подчеркнуть, что установленные «оптимальные уровни» аминокислот в питании человека не являются постоянными при любых условиях. Они могут значительно возрастать, причем неравномерно для различных аминокислот, при некоторых физиологических или патологических состояниях.
Белки пищи прежде, чем быть использованы для построения тканей тела, предварительно расщепляются. Организмом используется для питания не сам пищевой белок, а его структурные элементы – аминокислоты и, может быть, частично простейшие пептиды, из которых затем в клетках синтезируются специфические для данного вида организма белковые вещества.
Каждый вид организма, каждый орган и каждая ткань содержат свои характерные белки, и при усвоении чужеродных белков пищи организм прежде всего лишает их видовой специфичности. Перед тем, как быть усвоенными белки должны быть разложены на индифферентный материал. Разложение белковых веществ на более простые, лишенные видовой специфичности соединения, способные всасываться в кровь через стенки кишечника, осуществляется в пищеварительных органов человека и животных путем последовательного действия ряда ферментов.
В полости рта белки никаким изменениям не подвергаются, так как в состав слюны необходимые для этого протеолитические ферменты не входят. Переваривание белков начинается в желудке. В этом процессе решающую роль играют два фактора: сильно кислая реакция желудочного сока и присутствие в нем протеолитического фермента, действующего на белки.
При переваривании белков в желудке роль протеолитического фермента играет пепсин, содержащийся в желудочном соке. Клетки слизистой желудка выделяют неактивный пепсиноген, который под влиянием соляной кислоты желудочного сока превращается в активный пепсин.
Одни белки поддаются действию пепсина весьма легко, другие – труднее, а третьи совсем не перевариваются. Легко расщепляются пепсином альбумины и глобулины как животного, так и растительного происхождения. Пепсин действует преимущественно на внутренние пептидные связи, довольно далеко расположенные от концов полипептидной цепи. Дробление белковой молекулы пепсином происходит так, что при этом образуются близкие по величине частицы. Однако под влиянием пепсина разрываются также и некоторые пептидные связи, находящиеся на конце полипептидной цепи. Пепсин более быстро расщепляет пептидные связи, образованные аминогруппами ароматических аминокислот. Из каждого белка получается соответствующий ему пептон: мясной (мышцы), яичный, рыбный и т. д.
Особый интерес представляют те изменения, которые претерпевают в желудке важнейшие белки молока, в частности казеиноген. Казеиноген молока относится к группе сложных белков – фосфопротеидов, в состав которых входит довольно значительное количество ортофосфорной кислоты. Под влиянием желудочного сока казеиноген молока превращается в казеин.
Установлено, что наиболее важную роль в стимуляции секреции желудочного сока высокой кислотности играет гастрин – полипептид, причисляемый к группе так называемых «пищеварительных гормонов». Гастрин синтезируется в слизистой оболочке привратника (отдела желудка). Образование гастрина и поступление его в кровь резко усиливается под влиянием пищи, вводимой в желудок. Гастрин построен из семнадцати аминокислотных остатков, расположенных в следующем порядке: гли-гли-про-три-мет-глю-глю-глю-глю-глю-ала-тир-гли-три-мет-асп-фен-Nh3 .
Пептоны, образовавшиеся из белков под влиянием пепсина представляют собой смеси еще достаточно высокомолекулярных сложных соединений, которые в желудке не всасываются и поэтому поступают при очередном опорожнении желудка вместе с пищевой кашицей в двенадцатиперстную кишку.
Здесь они подвергаются действию группы протеолитических ферментов, гидролизующих как белки, так и пептоны. Пищеварительный сок в кишечнике, действующий на белки, представляет собой смесь секрета панкреатической железы и слизистой кишечника. В кишечнике белки пищи подвергаются воздействию трипсина, химотрипсина и пептидаз. Трипсин содержится в поджелудочном соке в недеятельной форме в виде триписиногена. Под влиянием другого фермента глюкопротеидной природы – энтерокиназы – трипсиноген превращается в трипсин. Процесс превращения трипсиногена в трипсин сводится к отщепления небольшого пептида (гексопептида) с α-аминного конца полипептидной цепи. Выделение трипсина в недеятельной форме имеет большое биологическое значение. Поджелудочный сок содержит ряд других ферментов, например липазу и амилазу, представляющих собой, как и все ферменты белковые вещества. Присутствие в одном с ними растворе протеолитического фермента – трипсина – в активной форме могло бы привести к их перевариванию и разрушению еще в панкреатической железе.
Трипсин гидролитически разрушает как белки, не изменившееся в желудке под влиянием пепсина, так и высокомолекулярные продукты распада белков, полипептиды типа пептонов. Оптимум РН для трипсина равен 7,8. Трипсин и пепсин действуют на различные пептидные связи в молекуле белка. Трипсин особенно легко расщепляет связи, в образовании которых участвуют карбоксильные группы аргинина или лизина. Трипсин производит сравнительно неглубокий гидролиз белка. Только около одной трети всех пептидных связей в белковой молекуле расщепляется трипсином. Основными продуктами триптического гидролиза белка являются полипептиды. Следует отметить, что под влиянием трипсина в процессе гидролиза белка могут освобождаться в небольшом количестве и свободные аминокислоты. Природы продуктов триптического гидролиза зависит от состава и строения исходного субстрата гидролиза (белка, пептона и т.п.).
Другим протолитическим ферментом, действующим в кишечнике во время пищеварения, является химотрипсин. Он содержится в поджелудочной железе в неактивном состоянии, в виде двух зимогенов – химотрипсиногена А и химотрипсиногена В, которые под действием трипсина переходят в кишечнике в активный химотрипсин.
Химотрипсиноген А состоит из полипептидной цепи, содержащей 246 аминокислот. В зависимости от того, происходит ли активирование химотрипсиногена в результате действия трипсина или химотрипсина, образуются различные смеси химотрипсинов.
Кристаллический химотрипсин подобно трипсину гидролизует как белки, так и пептоны с образованием относительно низкомолекулярных пептидов. Он расщепляет по преимуществу те пептидные связи, на которые трипсин не действует. Если на казеин воздействовать трипсином, а затем по окончании триптического гидролиза добавит химотрипсин, то гидролиз белка продолжается. Равным образом казеин, предварительно гидролизованный химотрипсином, гидролизуется дальше добавленным трипсином. В некоторых случаях химотрипсин производит даже более глубокий гидролиз белка, чем трипсин, и при этом расщепляется почти половина пептидных связей в белковой молекуле.
Следует особо подчеркнуть, что трипсин и химотрипсин гидролизуют в кишечнике также и такие белки, которые почему-либо не подверглись предварительному расщеплению пепсином в желудке.
Полипептиды, образовавшиеся в результате действия на белки пепсина, а затем трипсина и химотрипсина, подвергаются дальнейшему расщеплению в кишечнике, которое осуществляется под влиянием пептидаз. Как трипсин и химотрипсин пепсидазы выделяются железистыми клетками слизистой тонкой кишки в недеятельной или малоактивной форме. Активация их происходит под действием трипсина. В поджелудочном соке имеются две карбоксипептидазы – А и В, а в кишечном соке – аминопептидаза и ряд дипептидаз. Карбосксипептидазы расщепляют полипептиды с того конца цепи, где имеется свободная карбоксильная группа, причем карбоксипептидаза А – при наличии концевых ароматических аминокислот, а карбоксипептидаза В – при расположении на концах цепи основных аминокислот. Карбоксипептидаза, как и другие протеолитические ферменты поджелудочного сока, выделяются в виде неактивного зимогена.
Аминопептидаза расщепляет полипептиды с того конца, где имеется свободная аминогруппа.
Дипептидаза расщепляет дипептиды на свободные аминокислоты. В результате ферментативного гидролиза полипептидов пептидазами в кишечнике образуются свободные аминокислоты.
Отсюда следует важный вывод о том, что под влиянием совместного действия группы протеолитических ферментов белки пищи распадаются в желудочно-кишечном тракте до аминокислот.
Следует иметь в виду, что плохая перевариваемость различных пищевых белков может быть обусловлена также присутствием в них ингибиторов протеаз. Так, например, соевые бобы содержат мощный ингибитор трипсина; в яичном белке обнаружен мукопротеид также сильно угнетающий действие трипсина.
Помимо перечисленных выше протеолитических ферментов, в содержимом кишечника обнаруживаются также ферменты эластаза, действующая на эластин эластических связок и коллагеназа, гидролизующая коллаген костной и хрящевой ткани. Эти ферменты поступают в кишечник в составе поджелудочного сока.
Расщепление пептонов, так же как и жиров и углеводов, под влиянием соответствующих гидролаз особенно энергично проходит на поверхности слизистой оболочки кишечника (пристеночное пищеварение).
Увеличению скорости пищеварения непосредственно на поверхности слизистой кишечника в немалой степени способствует протекание тесно связанных с перевариванием пищи процессов всасывания.
Здесь надо также напомнить, что кишечный сок в отличие от желудочного и поджелудочного секретов содержит, помимо жидкой части, плотную часть, состоящую из отторгнутых клеток слизистой оболочки кишечника. Эти клетки очень богаты ферментами. Концентрация ферментов в жидкой части сок значительно ниже. Особенно высоко содержание ферментов в только что отторгнутых от стенки кишечника клеточных элементах.
Картина превращения пищевых белков в желудочно-кишечном тракте была бы не полной, если бы мы прошли мимо тех изменений, которые претерпевают белки (аминокислоты) в кишечнике под действием разнообразных микроорганизмов, населяющих в огромном количестве этот участок пищеварительной трубки. Часть аминокислот в кишечнике до их всасывания используются микробами в качестве источника питания.
Расщепление микробами аминокислот приводит к превращению их в амины, жирные кислоты, спирты, фенолы, индол, скатол, сероводород и ряд других соединений. Этот процесс носит название гниения белков в кишечнике. Прежде чем останавливаться на частностях, рассмотрим направление этих реакций.
1. При декарбоксилировании аминокислот возможно получение соответствующих, нередко ядовитых аминов.
2. При дезаминировании аминокислот микробами, когда происходит отщепление аминогруппы в виде аммиака, в зависимости от условий, возникают различные продукты, среди них насыщенные и не насыщенные кислоты кетокислоты и оксикислоты.
Гниение белков в органах пищеварения с образованием ядовитых продуктов происходит в более или менее значительных размерах лишь в нижних отделах кишечника. В полости рта и желудке условий для развития гнилостных бактерий обычно нет.
Амины, получающиеся при декарбоксилировании аминокислот, представляют собой фармакологически активные вещества, а некоторые из них являются даже сильными ядами. Из отдельных протеиногенных аминов, то есть аминов, образующихся из аминокислот под влиянием микробов кишечника необходимо назвать путресцин, кадаверин, фенилэтиламин и индолэтиламин.
Путресцин получается при декарбоксилировании аминокислоты орнитина:
При всасывании путресцина из кишечника в кровь этот диамин выделяется с мочой.
Кадаверин получается совершенно аналогичным образом при декарбоксилировании диаминокислоты лизина (α,ε-диаминокапроновой кислоты):
Если кадаверин всасывается из кишечника в кровь, то часть его, так же как и путресцина выводится из организма через почки в неизмененном виде. Часть образующихся аминов может обезвреживаться в стенках кишечника и в других тканях под действием фермента аминксидазы.
Кадаверин, так же как и путресцин, относят к группе трупных ядов, или птомаинов, так как они образуются и при разложении трупов; ядовитость этих диаминов, однако, незначительна.
Совершенно аналогичным образом из фенилаланина в кишечнике при гниении белков получается фенилэтиламин, а из триптофана – индолилэтиламин:
Сероводород (h3 S), метилмеркаптан (Ch4 SH) и другие содержащие серу соединения получаются при глубоком разрушении кишечными бактериями аминокислот цистина, цистеина и метионина. Из ядовитых продуктов гниения белков следует назвать фенол, паракрезол, скатол и индол.
Фенол и крезол могут образовываться из аминокислоты тирозина
Микробы разрушают боковую цепь аминокислоты, постепенно укорачивая ее.
Образовавшиеся в кишечнике под действием бактерий ядовитые продукты распада тирозина после всасывания обезвреживаются в печени, в которую оттекающая от кишечника кровь попадает через систему воротной вены. Обезвреживание фенола и крезола может происходить двояким путем: либо посредством связывания их серной кислотой, либо путем соединения их с глюкуроновой кислотой.
Список использованной литературы
1) Аврансон Л.А., Гуткевич Н.В. Обмен белков. ― М.: Красный крест, 1994. ― 127 с.
2) Баранова Т. А. Правильное питание. ― М.: Интербук, 1991. ― 141 с.
3) Башлов В., Гуржин И. Биологическая химия. ― Самара:, 1992. ― 517 с.
4) Велобова Е. Н. Переваривание белков. ― Киев: Гродынец, 1993. ― 29 с.
5) Герундов И. Н., Когосов П.Р. Рассказы о питании. ― Минск: Наука и техника, 1986. ― 89 с.
6) Зеродич., Лучинков. Справочник по химии. — Лениздат. — 594.
7) Збарский Б.И., Иванов И.И., Мардашев С.Р. Биологическая химия. – Л.: Медицина, 1972. ― 583 с.
8) Погудов И. П. Что химия знает о нас? ― М.: Политиздат, 1990. ― 287 с.
9) Яковлев В. В., Яковлев Д.В. Биологическая химия ― Минск: Вышэйш. шк., 1985. ― 494 с.
www.ronl.ru
Доклад - Аминокислоты, белки - Химия
Содержание.
1. Классификация аминокислот.
2. Синтезы  ,
,  ,
,  — аминокислот.
— аминокислот.
3. Свойства аминокислот: амфотерность, реакция по аминогруппе и карбоксилу.
4.  — аминокислоты, их роль в природе.
— аминокислоты, их роль в природе.
5. Синтез пептидов.
Белковые вещества:
1. Классификация.
2. Строение. Первичная структура, понятие о вторичной, третичной и четвертичной структурах.
3. Понятие о ферментах.
Классификация аминокислот.
Аминокислотами называются органические кислоты, содержащие одну или несколько аминогрупп. В зависимости от природы кислотной функции аминокислоты подразделяют на аминокарбоновые, например h3 N(Ch3 )5 COOH, аминосульфоновые, например h3 N(Ch3 )2 SO3 H, аминофосфоновые, h3 NCH[P(O)(OH)2 ]2, аминоарсиновые, например, h3 NC6 h5 AsO3 h3 .
Согласно правилам ИЮПАК название аминокислот производят от названия соответствующей кислоты; взаимное расположение в углеродной цепи карбоксильной и аминной групп обозначают обычно цифрами, в некоторых случаях — греческими буквами. Однако, как правило, пользуются тривиальными названиями аминокислот. ( см. таблицу 1.).
В зависимости от положения аминогруппы по отношению к карбоксилу различают  ,
,  и
и 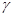 — аминокислоты:
— аминокислоты:
|
|
|
 -аминомасляная
-аминомасляная -аминопропионовая
-аминопропионовая  -аминомасляная
-аминомаслянаяВсе  — аминокислоты, кроме аминоуксусной (глицина), имеют асимметрический
— аминокислоты, кроме аминоуксусной (глицина), имеют асимметрический  — углеродный атом и существуют в виде двух энантиомеров. За редким исключением, природные
— углеродный атом и существуют в виде двух энантиомеров. За редким исключением, природные  - аминокислоты относятся к L- ряду (S-конфигурация) и имеют следующее пространственное строение:
- аминокислоты относятся к L- ряду (S-конфигурация) и имеют следующее пространственное строение:
 По физическим и ряду химических свойств аминокислоты резко отличаются от соответствующих кислот и оснований. Они лучше растворяются в воде, чем в органических растворителях; хорошо кристаллизуются; имеют высокую плотность и исключительно высокие температуры плавления. Эти свойства указывают на взаимодействие аминных и кислотных групп, вследствие чего аминокислоты в твёрдом состоянии и в растворе (в широком интервале pH) находятся в цвиттер-ионной форме (т.е. как внутренние соли). Взаимное влияние групп особенно ярко проявляется у
По физическим и ряду химических свойств аминокислоты резко отличаются от соответствующих кислот и оснований. Они лучше растворяются в воде, чем в органических растворителях; хорошо кристаллизуются; имеют высокую плотность и исключительно высокие температуры плавления. Эти свойства указывают на взаимодействие аминных и кислотных групп, вследствие чего аминокислоты в твёрдом состоянии и в растворе (в широком интервале pH) находятся в цвиттер-ионной форме (т.е. как внутренние соли). Взаимное влияние групп особенно ярко проявляется у 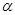 - аминокислот, где обе группы находятся в непосредственной близости.
- аминокислот, где обе группы находятся в непосредственной близости.
 |
Цвиттер-ионная структура аминокислот подтверждается их большим дипольным моментом (не менее 50×10-30 Кл × м), а также полосой поглощения в ИК- спектре твердой аминокислоты или её раствора.
Таблица 1. Важнейшие аминокислоты.
| Тривиальное название | Сокр.название ос- татка ами нок-ты | Формула | Температура плавления, 0С. | Растворимость в воде при 250С, г/100г. |
| Моноаминомонокарбоновые кислоты | ||||
| Гликокол или глицин | Gly | h3 NCh3 COOH | 262 | 25 |
| Аланин | Ala | h3 NCH(Ch4 ) COOH | 297 | 16,6 |
| Валин | Val | h3 NCHCOOH ï CH(Ch4 )2 | 315 | 8,85 |
| Лейцин | Leu | h3 NCHCOOH ï Ch3 CH(Ch4 )2 | 337 | 2,2 |
| Изолейцин | He | h3 NCHCOOH ï Ch4 ─ CH ─ C2 H5 | 284 | 4,12 |
| Фенилаланин | Phe | h3 NCHCOOH ï Ch3 C6 H5 | 283 (разл.) | ― |
| Моноаминодикарбоновые кислоты и их амиды | ||||
| Аспарагиновая кислота | Asp(D) | h3 NCHCOOH ï Ch3 COOH | 270 | 0,5 |
| Аспарагин | Asn(N) | h3 NCHCOOH ï Ch3 CONh3 | 236 | 2,5 |
| Глутаминовая кислота | Glu(E) | h3 NCHCOOH ï Ch3 Ch3 COOH | 249 | 0,84 |
| Глутамин | Gln(Q) | h3 NCHCOOH ï Ch3 Ch3 CONh3 | 185 | 4,2 |
| Диаминомонокарбоновые кислоты | ||||
| Орнитин(+) | Orn | h3 NCHCOOH ï Ch3 Ch3 Ch3 Nh3 | 140 | ― |
| Лизин | Lys(K) | h3 NCHCOOH ï Ch3 Ch3 Ch3 Ch3 Nh3 | 224 | Хорошо растворим |
| Аминокислоты | ||||
| Аргинин | Arg® | h3 NCHCOOH ï Ch3 ï Ch3 Ch3 NH ─ C ─ Nh3 ║ NH | 238 | 15 |
| Гидроксиаминокислоты | ||||
| Серин | Ser(S) | h3 NCHCOOH ï Ch3 OH | 228 | 5 |
| Треонин | Tre(T) | h3 NCHCOOH ï Ch3 (OH)Ch4 | 253 | 20,5 |
| Тирозин | Tyr(Y) | h3 NCHCOOH ï Ch3 C6 h5 OH-n | 344 | ― |
| Тиоаминокислоты | ||||
| Метионин | Met(M) | h3 NCHCOOH ï Ch3 Ch3 SCh4 | 283 | 3,5 |
 Цистин Цистин | (Cys)2 | 2 | 260 | 0,011 |
| Цистеин | Cys© | h3 NCHCOOH ï Ch3 SH | 178 | Хорошо растворим |
| Гетероциклические аминокислоты | ||||
         Триптофан Триптофан | Try(W) | h3 NCHCOOH ï h3 C NH | 382 | 1,14 |
   Пролин Пролин | Pro(P) | h3 C Ch3 ïï h3 C CHCOOH NH | 299 | 16,2 |
   Оксипролин Оксипролин | Opr | HOHC Ch3 ïï h3 C CHCOOH NH | 270 | 36,1 |
     Гистидин Гистидин | His(H) | Nh3 CHCOOH ï h3 C ―C ―― CH ïï N NH CH | 277 | 4,3 |
Синтезы  ,
,  ,
,  — аминокислот.
— аминокислот.
 - аминокислоты получают галогенированием карбоновых кислот или эфиров в
- аминокислоты получают галогенированием карбоновых кислот или эфиров в  -положение с последующей заменой галогена на аминогруппу при обработке амином, аммиаком или фталимидом калия (по Габриэлю).
-положение с последующей заменой галогена на аминогруппу при обработке амином, аммиаком или фталимидом калия (по Габриэлю).
|
 -аминоуксусная кислота
-аминоуксусная кислотаПо Штрекеру – Зелинскому  - аминокислоты получают из альдегидов:
- аминокислоты получают из альдегидов:
|
Этот метод позволяет также получать нитрилы и амиды соответствующих  - аминокислот. По сходному механизму протекает образование
- аминокислот. По сходному механизму протекает образование  - аминофосфоновых кислот по реакции Кабачника- Филдса, например:
- аминофосфоновых кислот по реакции Кабачника- Филдса, например:
|
В этой реакции вместо альдегидов могут быть использованы кетоны, а вместо диалкилфосфитов- диалкилтиофосфиты, кислые эфиры алкилфосфонистых кислот RP(OH)OR и диарилфосфиноксиды Ar2 HPO.
Альдегиды и кетоны или их более активные производные – кетали служат для синтеза  — аминокислот с увеличением числа углеродных атомов на две единицы. Для этого их конденсируют с циклическими производными аминоуксусной кислоты – азалактонами, гидантоинами, тиогидантоинами, 2,5-пиперазиндионами или с её медными или кобальтовыми хелатами, например:
— аминокислот с увеличением числа углеродных атомов на две единицы. Для этого их конденсируют с циклическими производными аминоуксусной кислоты – азалактонами, гидантоинами, тиогидантоинами, 2,5-пиперазиндионами или с её медными или кобальтовыми хелатами, например:
 |
Удобные предшественники  - аминокислот — аминомалоновый эфир и нитроуксусный эфир. К их
- аминокислот — аминомалоновый эфир и нитроуксусный эфир. К их  - углеродным атомам можно предварительно ввести желаемые радикалы методами алкилирования или конденсации.
- углеродным атомам можно предварительно ввести желаемые радикалы методами алкилирования или конденсации.  - кетокислоты превращают в
- кетокислоты превращают в  - аминокислоты гидрированием в присутствии Nh4 или гидрированием их оксимов, гидразонов и фенилгидразонов.
- аминокислоты гидрированием в присутствии Nh4 или гидрированием их оксимов, гидразонов и фенилгидразонов.
 |

Можно получать  - аминокислоты также непосредственно из
- аминокислоты также непосредственно из  - кетонокислот, действуя на них аммиаком и водородом над никелевым катализатором:
- кетонокислот, действуя на них аммиаком и водородом над никелевым катализатором:

Некоторые L- - аминокислоты ввиду сложности синтеза и разделения оптических изомеров получают микробиологическим способом (лизин, триптофан, треонин) или выделяют из гидролизатов природных белковых продуктов (пролин, цистин, аргинин, гистидин).
- аминокислоты ввиду сложности синтеза и разделения оптических изомеров получают микробиологическим способом (лизин, триптофан, треонин) или выделяют из гидролизатов природных белковых продуктов (пролин, цистин, аргинин, гистидин).
 — аминосульфоновые кислоты получают при обработке аммиаком продуктов присоединения NaHSO3 к альдегидам:
— аминосульфоновые кислоты получают при обработке аммиаком продуктов присоединения NaHSO3 к альдегидам:
RCHO + NaHSO3 ® RCH(OH)SO3 Na ® RCH(Nh3 )SO3 Na
|
 - аминокислоты синтезируют присоединением Nh4 или аминов к
- аминокислоты синтезируют присоединением Nh4 или аминов к  ,
,  - ненасыщенным кислотам:
- ненасыщенным кислотам:В.М.Родионов предложил метод, в котором совмещаются в одной операции получение  ,
,  -непредельной кислоты конденсацией альдегида с малоновой кислотой и присоединение аммиака:
-непредельной кислоты конденсацией альдегида с малоновой кислотой и присоединение аммиака:
 |


 -аминокислоты получают гидролизом соответствующих лактамов, которые образуются в результате перегруппировки Бекмана из оксимов циклических кетонов под действием h3 SO4.
-аминокислоты получают гидролизом соответствующих лактамов, которые образуются в результате перегруппировки Бекмана из оксимов циклических кетонов под действием h3 SO4.  -аминоэтановую и
-аминоэтановую и  -аминоундекановую кислоты синтезируют из
-аминоундекановую кислоты синтезируют из  ,
, ,
, ,
, -тетрахлоралканов путем их гидролиза конц. h3 SO4 до -хлоралкановых кислот с последующим аммонолизом:
-тетрахлоралканов путем их гидролиза конц. h3 SO4 до -хлоралкановых кислот с последующим аммонолизом:
Cℓ(Ch3 Ch3 )n CCℓ3 → Cℓ(Ch3 Ch3 )n COOH → h3 N(Ch3 Ch3 )n COOH
Исходные тетрахлоралканы получают теломеризацией этилена с CCℓ4.
Бекмановская перегруппировка оксимов циклических кетонов. Наибольшей практический интерес представляет перегруппировка оксима циклогексанона:

Получаемый этим путем капролактам полимеризуют в высокомолекулярный поликапромид



из которого изготовляют капроновое волокно.
Свойства аминокислот: амфотерность, реакция по аминогруппе и карбоксилу.
1. Большинство аминокислот – бесцветные кристаллические вещества, обычно хорошо растворимы в воде, часто сладковаты на вкус.
2. В молекулах аминокислот содержатся две группы с прямо противоположными свойствами: карбоксильная группа-кислотная, и аминогруппа с основными свойствами. Поэтому они обладают одновременно и кислотными и основными свойствами. Как кислоты, аминокислоты образуют со спиртами сложные эфиры, а с металлами и основаниями-соли:
 |
Для аминокислот особенно характерно образование медных солей, обладающих специфической синей окраской. Эти вещества являются внутренними комплексными солями; в них атом меди связан не только с атомами кислорода, но и с атомами азота аминогрупп:
|


Связь между атомом меди и азота осуществляется дополнительными валентностями( за счет свободной пары электронов азота аминогруппы). Как видно, при этом возникают кольчатые структуры, состоящие из пятичленных циклов. На легкость образования подобных пяти- и шестичленных циклов обратил внимание в 1906г. Л.А. Чугаев и отметил их значительную устойчивость. Медь(и другие металлы) в таких внутрикомплексных соединениях не имеют ионного характера. Водные растворы подобных соединений не проводят в заметной степени электрический ток.
При действии едких щелочей на медные соли аминокислот не происходит выпадания гидрата окиси меди. Однако при действии сероводорода происходит разрушение внутрикомплексного соединения и выпадает труднорастворимая в воде сернистая медью
3. Кислотные свойства в моноаминокислотах выражены весьма слобо-аминокислоты почти не изменяют окраски лакмуса. Таким образом, кислотные свойства карбоксила в них значительно ослаблены.
4. Как амины, аминокислоты образуют соли с кислотами, например:
HCℓ∙Nh3 Ch3 COOH
Но эти соли весьма непрочны и легко разлагаются. Таким образом, основные свойтва аминогруппы в аминокислотах также значительно ослаблены.
5. При действии азотистой кислоты на аминокислоты образуются оксикислоты:
 Nh3 Ch3 COOH + NHO2 HOCh3 COOH + N2 + h3 O
Nh3 Ch3 COOH + NHO2 HOCh3 COOH + N2 + h3 O
Эта реакция совершенно аналогична реакции образования спиртов при действии азотистой кислоты на первичные амины.
6. С галоидангидритами кислот аминокислоты образуют вещества, которые одновременно являются и аминоксилотами и амидами кислот. Так, при действии хлористого ацетила на аминоуксусную кислоту образуется ацетиламиноуксусная килослота:
 Ch4 COСℓ + Nh3 Ch3 COOH Сh3 CONHСh3 COOH + HCℓ
Ch4 COСℓ + Nh3 Ch3 COOH Сh3 CONHСh3 COOH + HCℓ
ацетиламиноуксусная килослота
Ацетиламиноуксусную кислоту можно рассматривать и как производное аминоуксусной кислоты, в молекуле которой атом водорода аминогруппы замещен ацетилом Ch4 CO- и как ацетамид, в молекуле которого атом водорода аминогруппы замещен остатком уксусной кислоты -Ch3 COOH.
7.  - Аминокислоты принагревании легко отщепляют воду,
- Аминокислоты принагревании легко отщепляют воду,
причем из двух молекул аминокислоты выделяются две молекулы воды и образуются дикетопиперазины:
 |
 |

|
Дикетопиперазины-циклические соединения, кольцо которых образовано четырьмя атомами углерода и двумя атомами азота. Дикетопиперазины — твердые, хорошо кристаллизующиеся вещества.
|
 — Аминопропиновая кислота акриловая кислота
— Аминопропиновая кислота акриловая кислота — Аминокслоты при нагревании теряют аммиак, переходя в непредельные кислоты:
— Аминокслоты при нагревании теряют аммиак, переходя в непредельные кислоты:
 |
 — Аминокслоты легко отщепляют воду, образуя лактамы:
— Аминокслоты легко отщепляют воду, образуя лактамы:




|
Лактамы можно рассматривать как внутренние амиды.
8. Аминокислоты образуют сложные эфиры при действии хлористого водорода на них спиртовые растворы. При этом, разумеется, образуется солянокислые соли эфиров, из которых свободные эфиры можно получить, удаляя хлористый водород окисью серебра, окисью свинца или триэтиламином:



 + +
+ +
 Nh4 —Ch3 —C—O– + C2 H5 OH + HCℓ Nh4 —Ch3 —C—OC2 H5 Cℓ–
Nh4 —Ch3 —C—O– + C2 H5 OH + HCℓ Nh4 —Ch3 —C—OC2 H5 Cℓ–
║ ║

 O O
O O
+




 2 Nh4 —Ch3 —C—OC2 H5 Cℓ– + Ag2 O Nh3 —Ch3 —C—OC2 H5 + 2AgCℓ– + h3 O
2 Nh4 —Ch3 —C—OC2 H5 Cℓ– + Ag2 O Nh3 —Ch3 —C—OC2 H5 + 2AgCℓ– + h3 O
║ ║

 OO
OO
Эфиры обычных аминокислот — жидкости, перегоняющиеся в вакууме. Именно этерификацией суммы аминокислот, получающихся в результате гидролиза белка, разгонкой в вакууме и последующим гидролизом Э.Фишер выделил индивидуальные аминокислоты и дал способ установления аминокислотного состава белков.
9. При действии пятихлористого фосфора на аминокислоты образуются солянокислые соли хлорангидридов аминокислот, довольно неустойчивые соединения, при отщеплении HCℓ образующие совсем неустойчивые свободные хлорангидриды:



 + +
+ +
 Nh4 —Ch3 —C—O– + PCℓ5 Nh4 —Ch3 —C—Cℓ Cℓ– + POCℓ3
Nh4 —Ch3 —C—O– + PCℓ5 Nh4 —Ch3 —C—Cℓ Cℓ– + POCℓ3
║ ║

 OO
OO
10. Аминокислоты ацилируются по аминогруппе:

 +
+
 Nh3 —Ch3 —C—O– + Ch3 —C O Ch4 —C—NH—Ch3 —C—OH + Ch4 —C—OH
Nh3 —Ch3 —C—O– + Ch3 —C O Ch4 —C—NH—Ch3 —C—OH + Ch4 —C—OH
║ ║ ║ ║ ║
O O 2 O O O
ацетилгликокол
+
 Nh3 —Ch3 —C—O– + C6 H5 —C—Сℓ C6 H5 —C—NH—Ch3 —C—OH + HСℓ
Nh3 —Ch3 —C—O– + C6 H5 —C—Сℓ C6 H5 —C—NH—Ch3 —C—OH + HСℓ
║ ║ ║ ║
O O O O
бензоилгликокол
(гиппуровая кислота)
Образующаяся в последней из написанных реакций гиппуровая кислота – вещество, в виде которого травоядные животные выделяют с мочой небезвредную бензойную кислоту, попадающую в организм с пищей.
11. Аминокислоты можно алкилировать по аминогруппе. Алкилированием глицина получается метиламиноуксосная кислота-саркозин
+ +
 Nh4 —Ch3 —C—O– + Ch3 I Ch4 Nh3 —Ch3 —C—O– + HI
Nh4 —Ch3 —C—O– + Ch3 I Ch4 Nh3 —Ch3 —C—O– + HI
║ ║
OO
саркозин
которая в связанном виде содержится в некоторых белках
При избытке иодистого метила образуется замещенная на четвертичноаммониевую группировку уксусная кислота



 + +
+ +
 Nh4 —Ch3 —C—O– + 3Ch4 I (Ch4 )3 N—Ch3 —C—OH I– + 2HI
Nh4 —Ch3 —C—O– + 3Ch4 I (Ch4 )3 N—Ch3 —C—OH I– + 2HI
║ ║

 OO
OO
от которой можно отщепить HI и получить бетаин, лучше синтезируемый из триметиламина и хлоруксусной кислоты:
+
 (Сh4 )3 N + CℓCh3 —C—ONa (Ch4 )3 N—Ch3 —C—O– + NaCℓ
(Сh4 )3 N + CℓCh3 —C—ONa (Ch4 )3 N—Ch3 —C—O– + NaCℓ
║ ║
OO
бетаин
Бетаин, получивший свое название от свеклы(Betavulgaris), в соке которой он находится, дал название и всему классу внутренних солей, в которых анион и катион связаны внутри одной молекулы. В этом смысле говорят о бетаинобразной структуре самих аминокислот:
+ +
h4 N—Ch3 —C—O– (Ch4 )3 N—Ch3 —C—O–
║ ║
OO
Бетаины обладают большим дипольным моментом и солеобразны (тверды, нелетучи, водорастворимы).
 — аминокислоты, их роль в природе.
— аминокислоты, их роль в природе.
В молекулах большинства  - аминокислот содержится асимметрический атом углерода; природные аминокислоты существуют в виде оптических антиподов. Те из антиподов, конфигурация которых аналогична конфигурации правовращающего глицеринового альдегида, обозначаются буквой D; буквой L обозначаются антиподы, конфигурация которых соответствует конфигурации левовращающего глицеринового альдегида:
- аминокислот содержится асимметрический атом углерода; природные аминокислоты существуют в виде оптических антиподов. Те из антиподов, конфигурация которых аналогична конфигурации правовращающего глицеринового альдегида, обозначаются буквой D; буквой L обозначаются антиподы, конфигурация которых соответствует конфигурации левовращающего глицеринового альдегида:
|
Конфигурация  - асимметрического атома углерода у всех природных
- асимметрического атома углерода у всех природных  -аминокислот одинаковая, все они относятся к L-ряду.
-аминокислот одинаковая, все они относятся к L-ряду.
Наибольший интерес представляют 20  - аминокислот, входящих в состав белковых молекул.
- аминокислот, входящих в состав белковых молекул.
Принадлежность природных аминокислот к  - аминокислотам легко устанавливается по их способности образовывать дикетопиперазины и характерные внутрикомплексные медные соли. Азотистая кислота превращает их в
- аминокислотам легко устанавливается по их способности образовывать дикетопиперазины и характерные внутрикомплексные медные соли. Азотистая кислота превращает их в  - оксикислоты, из которых многие известные уже со второй половины XVIII столетия. Если при этом получается неизвестная оксикислота, то аминогруппу можно заменить на хлор действием хлористого нитролиза
- оксикислоты, из которых многие известные уже со второй половины XVIII столетия. Если при этом получается неизвестная оксикислота, то аминогруппу можно заменить на хлор действием хлористого нитролиза
RR
+ │ │
 Nh4 —CH—C—O– + NOCℓ Cℓ—CH—C—OH + N2 + h3 O
Nh4 —CH—C—O– + NOCℓ Cℓ—CH—C—OH + N2 + h3 O
║ ║
OO
а затем, заменив действием цинка в кислой среде хлор на водород, получить известную карбоновую кислоту(в случае аминодикарбоновых кислот-двухосновную). Таким путем, например, из валина образуется изовалериановая кислота, из лейцина – изокапроновая, из глутаминовой – глутаровая, которые легко идентифицировать по каким-либо твердым производным (например по анилидам).
Синтез пептидов.
Дипептидами называются  - амино-N-ациламинокислоты
- амино-N-ациламинокислоты
R—CH—C—NH—CH—C—O–
│+║ │ ║
Nh4 O R’ O
Полипептиды построены по тому же амидному принципу из нескольких одинаковых или разных аминокислот. Они называются по числу участвующих остатков аминокислот ди-, три- и т.д. полипептидами.
Дипетиды с одинаковыми  - аминокислотными остатками можно получить гидролитическим размыканием дикетопиперазинов. Дипетиды с любыми
- аминокислотными остатками можно получить гидролитическим размыканием дикетопиперазинов. Дипетиды с любыми  - аминокислотными остатками были получены Э. Фишером путем ацилирования аминокислоты по аминогруппе хлорангидритом
- аминокислотными остатками были получены Э. Фишером путем ацилирования аминокислоты по аминогруппе хлорангидритом  - галоидзамещенной кислоты и последующей заменой
- галоидзамещенной кислоты и последующей заменой  - галоида на аминогруппу действием аммиака:
- галоида на аминогруппу действием аммиака:
 Сℓ—CH—C—Сℓ h3 N—CH—C—OH Сℓ—CH—C—NH—CH—C—OH + HСℓ
Сℓ—CH—C—Сℓ h3 N—CH—C—OH Сℓ—CH—C—NH—CH—C—OH + HСℓ
│║ │ ║│║ │ ║
ROR’ OROR’ O
 Сℓ—CH—C—NH—CH—C—OH + 2Nh4 h3 N—CH—C—NH—CH—C—O– + Nh5 Сℓ
Сℓ—CH—C—NH—CH—C—OH + 2Nh4 h3 N—CH—C—NH—CH—C—O– + Nh5 Сℓ
│║ │ ║ │║ │ ║
R O R’ O R O R’ O
Подобная же последовательность реакций, примененная к полученному дипетиту, приведет к трипетиду и т.д. Э. Фишер получил таким путем октадекапептид, состоящий из 18 остатков аминокислот.
В более новых методах синтеза полипептидов исходят из хлорангидридов аминокислот(или из иных функциональных производных аминокислот с резко выраженной ацилирующей способностью) с защищенной аминогруппой. Такая защита необходима, чтобы хлорангидрид первой аминокислоты не проацилировал себе подобную молекулу, а осуществил связь со второй аминокислотой. Защита аминогруппы ацетилированием мало удобна, так как условия удаления ацетильной группы гидролизом таковы, что сам ди или полипептид будет гидролизоваться, распадаясь на аминокислоты. Поэтому аминогруппу кислоты, предназначенной в качестве ацилирующего агента и превращаемой для этого в хлорангидрид, защищают, в водя в аминогруппу такую группировку, которую можно удалить из дипептида гидролизом в очень мягких условиях или каким-либо другим методом. Например группу CF3 CO- можно удалить обработкой слабой щелочью или гидрогенолизом; группу C6 H5 Ch3 OCO- легко удалить гидрированием над палладиевым катализатором, восстановлением раствора натрия в жидком аммиаке или действием гидрозина; фталильная группа под действием гидразина отщепляется в виде
 CO
CO
 NH
NH
C6 h5 │

 NH
NH
CO
Что касается того, в форме какого функционального производного должен находится карбоксил защищенной описанным способом аминокислоты, то чаще чем хлорангидриды, применяют легко ацитилирующие эфиры или смешанные ангидриды, например:
 |
 |
HN—CH—C—O— —NO2 или HN—CH—C— —O—C—OR’
 │ │ ║ │ │ ║ ║
│ │ ║ │ │ ║ ║
X R O X R O O
где X- защищающий аминогруппу заместитель, R’- остаток пространственно затрудненного алифатического спирта, например (Ch4 )2 CH—Ch3 OH, что обеспечивает разрыв ацилирующей молекулы по линии, намеченной пунктиром.
Совершенно иное дело-получение полипептидов, даже с высоким молекулярным весом, из остатков одной кислоты. Для этой цели выработан следующий метод(Лейхс), рассмотренный на примере глицина (R=H):
R
│
 C6 H5 Ch3 —O—C—Cℓ + h3 N—CH—C—OH
C6 H5 Ch3 —O—C—Cℓ + h3 N—CH—C—OH
║ ║
O O
R
│

 C6 H5 Ch3 —O—C—NH—CH—C—OH
C6 H5 Ch3 —O—C—NH—CH—C—OH
║ ║
 O O O
O O O

R R—CH—C

 │ нагревание
│ нагревание

 C6 H5 Ch3 —O—C—NH—CH—C—Cℓ O
C6 H5 Ch3 —O—C—NH—CH—C—Cℓ O
 ║ ║
║ ║
 O O HN — C
O O HN — C
 |
O
Такие циклические внутренние смешанные ангидриды при нагревании распадаются с выделением CO2 и образованием высокомолекулярных полипетидов:
 O
O

R—CH—C R R

 │ │
│ │
 n O nCO2 + —HN—CH—C—(HN—CH—C—)n-1
n O nCO2 + —HN—CH—C—(HN—CH—C—)n-1
 ║ ║
║ ║
 HN — C O O
HN — C O O
 |
O
В химии белков и полипептидов для сокращения принято писать формулы, обозначая остатки аминокислот буквами. Например, Glu- обозначение глутаминовой кислоты, Cys- цистеина, Gly- глицина и т.д. Сокращенная формула глутатиона будет в таком изображении
Gly—Cys—Gly
Белковые вещества. Классификация.
Белки ― высокомолекулярные природные полимеры, построенные из остатков аминокислот, соединенных амидной (пептидной) связью ―CO―NH―.
Каждый Б. характеризуется специфической аминокислотной последовательностью и индивидуальной пространственной структурой (конформацией). На долю белков приходится не менее 50% сухой массы органических соединений животной клетки. Функционирование белка лежит в основе важнейших процессов жизнедеятельности организма. Обмен веществ (пищеварение, дыхание и др.), мышечное сокращение, нервная проводимость и жизнь клетки в целом неразрывно связаны с активностью ферментов — высокоспецифичных катализаторов биохимических реакций, являющихся белками. Основу костной и соединительной тканей, шерсти, роговых образований составляют структурные белки. Они же формируют остов клеточных органелл (митохондрий, мембран и др.). Расхождение хромосом при делении клетки, движение жгутиков, работа мышц животных и человека осуществляются по единому механизму при посредстве белка сократительной системы (напр., Актин, Миозин). Важную группу составляют регуляторные белки, контролирующие биосинтез белка и нуклеиновых кислот. К регуляторным белкам относятся также пептидно-белковые гормоны, которые секретируются эндокринными железами. Информация о состоянии внешней среды, различные регуляторные сигналы (в т. ч. гормональные) воспринимаются клеткой с помощью спец. рецепторных белков, располагающихся на наружной поверхности плазматической мембраны. Эти белки играют важную роль в передаче нервного возбуждения и в ориентированном движении клетки (хемотаксисе). В активном транспорте ионов, липидов, сахаров и аминокислот через биологические мембраны участвуют транспортные белки, или белки-переносчики. К последним относятся также гемоглобин и миоглобин, осуществляющие перенос кислорода. Преобразование и утилизация энергии, поступающей в организм с питанием, а также энергии солнечного излучения происходят при участии белков биоэнергетической системы (напр., родопсин, цитохромы). Большое значение имеют пищевые и запасные белки ( напр., Казеин, Проламины), играющие важную роль в развитии и функционировании организмов. Защитные системы высших организмов формируются защитными белками, к которым относятся иммуноглобулины (ответственны за иммунитет), белки комплемента (ответственны за лизис чужеродных клеток и активацию иммунологической функции), белки системы свертывания крови ( напр. Тромбин, Фибрин) и противовирусный белок интерферон.
По составу белки делят на простые, состоящие только из аминокислотных остатков (протеины), и сложные (протеиды). Сложные могут включать ионы металла (металлопротеиды) или пигмент (хромопротеиды), образовывать прочные комплексы с липидами (липопротеины), нуклеиновыми кислотами (нуклеопротеиды), а также ковалентно связывать остаток фосфорной кислоты (фосфопротеиды), углевода (гликопротеины) или нуклеиновой кислоты (геномы некоторых вирусов).
По ряду характерных свойств протеины можно разделить на несколько подгрупп:
Альбумины. Они растворимы в воде, свёртываются при нагревании, нейтральны, сравнительно трудно осаждаются растворами солей. Примерами их могут служить: альбумин белка куриного яйца, альбумин кровяной сыворотки, альбумин мускульной ткани, молочный альбумин.
Глобулины. Они нерастворимы в воде, но растворяются в очень слабых растворах солей. Более концентрированными растворами солей они вновь осаждаются; осаждение происходит при меньшей концентрации, чем та, которая необходима для осаждения альбуминов. Эти белки являются очень слабыми кислотами. Примерами глобулинов могут служить: фибриноген, глобулин кровяной сыворотки, глобулин мускульной ткани, глобулин белка куриного яйца.
Гистоны. Белки основного характера. Находятся в виде нуклеопротеидов в лейкоцитах и красных кровяных шариках.
Протамины. Не содержат серы, обладают сравнительно сильными основными свойствами, дают кристаллические соли; содержатся (в виде нуклеопротеинов) в сперматозоидах рыб.
Проламины. Находятся в зернах различных хлебных злаков. Замечательной их особенностью является растворимость в 80% -ном спирте. Представителем этих белков может служить глиадин, составляющий главную часть клейковины.
Склеропротеины. Нерастворимые белки, которые составляют наружный покров тела животного и находятся в скелете и в соединительной ткани. К ним относятся кератин, коллагены, эластин, фиброин.
Керотин является главной составной частью волос, рогов, копыт, ногтей, перьев и верхнего слоя кожи. Скорлупа куриного яйца состоит из извести и кератина. Если растворить известь скорлупы яйца в кислоте, то останется мягкая кожа, состоящая из кератина; из кератина состоит кожица, которая следует за скорлупой яйца. По химическому составу кератин богат серой.
Коллагены. Чрезвычайно распространены в живых организмах. Из коллагенов состоит соединительная ткань; они находятся в хрящах. Кости позвоночных животных состоят из неорганических веществ (фосфорнокислого и углекислого кальция), жира и коллагенов.
При кипячении с водой или при действии перегретого водяного пара коллагены образуют клей. Если из костей извлечь жир и потом, обработав их кислотой, растворить фосфорнокислый кальций, то останется белковое вещество-оссеин. При обработке оссеина перегретым водяным паром он переходит в клей. Чистый костяной клей называется желатиной. Особенно чистая желатина получается из рыбьего пузыря кипячением с водой.
Эластин входит в состав жил и других эластичных веществ соединительной ткани.
Нити сырого шелка состоят из белкового вещества-фиброина, покрытого другим белковым веществом, играющим роль шелкового клея,-серицином. При кипячении с водой шелк освобождается от клея который при этом переходит в раствор.
Протеиды также можно разделить на несколько групп: фосфоропротеиды содержат в своем составе фосфор. Они, в противоположность протаминам, обладающим, как указано выше, основными свойствами, имеют определенно выраженный кислотный характер.
Главнейшим представителем фосфоропротеидов является казеин молока. Он обладает настолько ясно выраженным кислотным характером, что разлагает углекислые соли с выделением углекислого газа. Казеин растворяется в слабых растворах щелочей, образуя с ними соли. Соли казеина называются казеинатами.
При нагревании казеин не свертывается. При действии кислот на соли казеина он выделяется в свободном виде. Этим объясняется свертывание молока при прокисании.
Казеин применяется для изготовления твердой, напоминающей рог пластмассы-галалита).Для получения галалита казеин смешивают с водой, красками и наполнителями, прессуют под давлением, и полученные пластины обрабатывают формалином. Казеин содержит фосфор в виде сложного эфира фосфорной кислоты.
Из других фосфоропротеинов следует отметить вителлин, который находится в желтке куриного яйца.
Нуклеопротеиды находятся в клеточных ядрах. При осторожном гидролизе они расщепляются на белок и нуклеиновую кислоту.
Нуклеиновые кислоты являются весьма сложными веществами, расщепляющимися при гидролизе на фосфорную кислоту, углеводы и азотосодержащие органические вещества группы пиримидина и группы пурина.
Хромопротеиды. Под этим названием известны протеиды, которые представляют собой сочетание белков с окрашенными веществами. Из хромопротеидов наиболее изучен гемоглобин красящее вещество красных кровяных шариков. Гемоглобин, соединяясь с кислородом,
превращается в оксигемоглобин, который, отдавая свой кислород другим веществам, снова превращается в гемоглобин. Значение гемоглобина в жизни человека и животных очень велико. Он играет роль переносчика кислорода от легких к тканям. Образовавшийся в легких оксигемоглабин кровью разносится по телу и, отдавая свой кислород, способствует протекание в организме окислительных процессов. Кроме того, гемоглобин вместе с плазмой крови осуществляет регуляцию величины pH крови и перенос углекислоты в организме.
Характерной особенностью гемоглобина является его способность соединятся с окисью углерода, после чего он теряет способность соединяться с кислородом. Этим объясняется ядовитое действии окиси углерода.
Гемоглобин представляет собой соединение белка глобина с красящим началом гемохромогеном. Вне организма гемоглобин, при действии воздуха, превращается в метгемоглобин, который отличается от оксигемоглобына прочностью связи с кислородом. При обработке ледяной уксусной кислотой метгемоглобин расщепляется с образованием глобина и гематина C34 h42 O4 N4 Fe(OH). Обработкой метгемоглобина тем же реактивом, но в присутствии NaCℓ, получается хлористая соль гематина, называемая гемином, C34 h42 O4 N4 FeCℓ. Гемин образует характерные красно-коричневые таблички, которые дают возможность открыть присутствие крови в пятнах даже через несколько лет. Гематин очень близок к гемохромогену, но все же от него отличается.
Глюкопротеиды. Некоторые белки этой группы встречаются в слизистых соединениях животных организмов и обусловливаются свойства этих выделений тянуться в нити даже при сравнительно большом разбавлении. Эти белки образуются в подчелюстной железе(подчелюстная железа-одна из слюнных желез), печени, железах желудка и кишечника. Другие глюкопротеиды находятся в хрящах, яичном белке, стекловидном теле глаза и т.д. Исследованныепредставители глюкопротеидов являются сочетанием белков с веществами, содержащими остатки некоторых производных углеводов, серной и уксусной кислот.
Строение белковых молекул.
 Практически все белки построены из 20
Практически все белки построены из 20  -аминокислот, принадлежащих, за исключением глицина, к L-ряду. Аминокислоты соединены между собой пептидными связями, образованными карбоксильной и
-аминокислот, принадлежащих, за исключением глицина, к L-ряду. Аминокислоты соединены между собой пептидными связями, образованными карбоксильной и  -аминогруппами соседних аминокислотных остатков Белковая молекула может состоять из одной или нескольких цепей, содержащих от 50 до нескольких сотен (иногда –более тысячи) аминокислотных остатков. Молекулы, содержащие менее 50 остатков часто относят к пептидам. В состав многих молекул входят остатки цистина, дисульфидные связи которых ковалентно связывают участки одной или нескольких цепей. В нативном состоянии макромолекулы белка обладают специфичной конформацией. Характерная для данного белка конформация определяется последовательностью аминокислотных остатков и стабилизируется водородными связями между пептидными и боковыми группами аминокислотных остатков, а также гидрофобными и электростатическими взаимодействиями.
-аминогруппами соседних аминокислотных остатков Белковая молекула может состоять из одной или нескольких цепей, содержащих от 50 до нескольких сотен (иногда –более тысячи) аминокислотных остатков. Молекулы, содержащие менее 50 остатков часто относят к пептидам. В состав многих молекул входят остатки цистина, дисульфидные связи которых ковалентно связывают участки одной или нескольких цепей. В нативном состоянии макромолекулы белка обладают специфичной конформацией. Характерная для данного белка конформация определяется последовательностью аминокислотных остатков и стабилизируется водородными связями между пептидными и боковыми группами аминокислотных остатков, а также гидрофобными и электростатическими взаимодействиями.
Различают четыре уровня организации белковых молекул.
Первичной структурой называют последовательность аминокислотных остатков в полипептидной цепи. Все белки различаются по первичной структуре, потенциально их возможное число практически неограничено.
Вторичная структура белка – это  -спираль, которая образуется в результате скручивания полипептидной цепи за счет водородных связей между группами: ―C― и ―N―
-спираль, которая образуется в результате скручивания полипептидной цепи за счет водородных связей между группами: ―C― и ―N― .
.
║ ║
OH
В одном витке спирали обычно содержится 3,6 аминокислотных остатка, шаг спирали – 0,544 нм.
Под третичной структурой белка понимают расположение его полипептидной цепи в пространстве. Существенное влияние на формирование третичной структуры оказывают размер, форма и полярность аминокислотных остатков. Третичная структура многих белков составляется из нескольких компактных глобул, называемых доменами. Между собой домены обычно бывают связаны тонкими перемычками – вытянутыми полипептидными цепями.
Термин четвертичная структура относится к макромолекулам, в состав которых входит несколько полипептидных цепей (субъединиц), не связанных между собой ковалентно. Между собой эти субъединицы соединяются водородными, ионными, гидрофобными и другими связями. Примером может служить макромолекула гемоглобина.

Понятие о ферментах.
Катализ многочисленных биохимических реакций, протекающих в растительных и животных организмах и их клетках, так же как и в одноклеточных микроорганизмах, совершается ферментами (энзимами).
Ферменты представляют, собой вещества или чисто белковые структуры, или протеиды-белки, связанные с небелковой простетической группой. Число уже известных ферментов очень велико. Считают, что одна клетка бактерии использует до 1000 разных ферментов. Однако лишь для не многих установлено строение. Примерами чисто белковых ферментов могут служить протеолитические ферменты пищеварения, такие как пепсин и трипсин. Известны случаи, когда один и тот же белок несет в организме и структурную и ферментную функцию. Примером служит белок мышц миозин, каталитически разлагающий аденозинтрифосфат – реакция, в данном случае дающая энергию сокращения мышцы.
Витамины – группы веществ обычно довольно сложной структуры, часто очень далеких химически друг от друга и объединяемых только по биологическому признаку. Это – требуемые в микроколичествах, но совершенно необходимые составные части пищи, недостаток которых вызывает болезнь, а отсутствие – гибель организма. Используя витамины(но не только их), организм человека и животных и строит свои ферменты. Одно и тоже вещество, служащее для организмов одного вида витамином, для других организмов может и не быть витамином по двум причинам: либо он может его синтезировать сам. Так аскорбиновая кислота – витамин для человека, но для крысы она не является витамином, так как организм крысы создает ее из глюкозы. Некоторые витамины имеют весьма универсальный характер и необходимы всем организмам – от одноклеточных(дрожжей и бактерий) до человека. Зеленые растения способны производить все свои вещества из минеральных исходных веществ и CO2 и, следовательно, не нуждаются в витаминах.
Для действия ряда белковых ферментов в организме необходимо присутствие микроколичеств ионов некоторых металлов-активаторов (Mg, Zn, Mo, Mn, Cu), которые витаминами не считаются. В состав некоторых коферментов и соответствующих витаминов входят металлы (железо, кобальт). Очень существенна роль микроколичеств металлов (микроэлементов) для развития растений; по-видимому, эти микроэлементы используются для постороения ферментов и служат как бы «витаминами растений».
Ферменты являются катализатороми, но отличаются от обычных катализаторов. Так им свойственны:
1. Гораздо большая специфичность по отношению к структуре катализируемого объекта и по отношению к реакции.
2. Полная стереохимическая специфичность.
3. Гораздо большая скорость протекания ферментных реакций по сравнению с теми же реакциями, катализируемыми обычнами катализаторами.
4. Невысокие оптимумы температуры их действия(обычно активность ферментов резко падает к 50О С и в интервале 50-100О С действие их полностью прекращается).
5. Денатурируемость- необратимая потеря каталитической активности при нагревании до 50-100О С. Существуют, однако, ферменты (трипсин, рибонуклеаза), активность которых восстанавливается по охлаждении даже после кипячения.
6. Существование оптимума кислотной среды для действия каждого фермента.
Высокая скорость ферментативных реакций объясняется, с одной стороны, как всегда при катализе, сильным снижением энергии активации реакции. Так, при катализе, сильным снижением энергии активации реакции. Так, при гидролизе казеина кислотой энергии активации 20,6 ккал/моль, а при гидролизе трипсином-только 12 ккал/моль. Гидролиз сахарозы кислотой требует энергии активации 25,5 ккал/моль, а ферментативный (сахарозой)-лишь 12-13 ккал/моль. С другой стороны, в ферментативных реакциях не меньшую роль играет предэкспоненциальный множитель уравнения Арреннуса, так как величина этого множителя, как правило, на много порядков выше, чем в реакциях обычного типа. Есть доказательство того, что ферменты содержат центры(«карманы»), фиксирующие субстрат на поверхности их молекул, и вторые центры, осуществляющие реакцию. Фермент может быть активен в том смысле, что он подтягивает активный центр к месту действия, несколько изменяя свою вторичную или третичную структуру.
Список используемой литературы:
1. А.Н.Несмеянов, Н.А.Несмеянов «Начала органической химии», т.1, 2.
2. Б.А.Павлов, А.П.Терентьев «Курс органической химии».
3. Химическая энциклопедия, т. 1.
4. Дж.Робертс, М.Касерио «Основы органической химии».
5. Шабаров Ю.С. «Органическая химия»
Саратовский Государственный Университет им. Н.Г.Чернышевского.
Реферат
Тема: Аминокислоты. Белки.
Выполнила: Чих Татьяна
гр. 212.
1999г.
www.ronl.ru
Доклад - Белки - Химия
БЕЛКИ
Любое соединение, которое содержитодновременно карбоксильную и аминогруппу, является аминокислотой.Однако, чаще этот термин применяется для обозначения карбоновых кислот,аминогруппа которых находится в a-положении к карбоксильной группе.
/>
Аминокислоты, как правило, входят всостав полимеров — белков. В природе встречается свыше 70 аминокислот,но только 20 играют важную роль в живых организмах. Незаменимыминазываются аминокислоты, которые не могут быть синтезированы организмом извеществ, поступающих с пищей, в количествах, достаточных для того, чтобыудовлетворить физиологические потребности организма. Незаменимые аминокислотыприводятся в табл. 1. Для больных фенилкетонурией незаменимой аминокислотойявляется также тирозин (см. табл. 1).
Таблица 1.Незаменимыеаминокислоты R-CHNh3COOH
Название (сокращение) R изолейцин (ile, ileu)Ch4Ch3CH(CH)3-
лейцин (leu)(Ch4)2CHCh3-
лизин (lys)Nh3Ch3Ch3Ch3Ch3-
метионин (met)Ch4SCh3Ch3-
фенилаланин (phe)C6H5Ch3-
треонин (thr)Ch4CH(OH)-
триптофан (try)/>
валин (val)(Ch4)2CH-
тирозин (tyr)Аминокислоты называют обычно какзамещенные соответствующих карбоновых кислот, обозначая положение аминогруппыбуквами греческого алфавита. Для простейших аминокислот обычно применяютсятривиальные названия (глицин, аланин, изолейцин и т.д.). Изомерия аминокислотсвязана с расположением функциональных групп и со строением углеводородногоскелета. Молекула аминокислоты моет содержать одну или несколько карбоксильныхгрупп и в соответствии с этим аминокислоты различаются по основности. Также вмолекуле аминокислоты может находиться разное количество аминогрупп.
Аминокислоты способны кполиконденсации, в результате которой образуется полиамид. Полиамиды, состоящиеиз a-аминокислот, называются пептидамиили полипептидами. Амидная связь в таких полимерах называется пептиднойсвязью. Полипептиды с молекулярной массой не меньше 5000 называют белками.В состав белков входит около 25 различных аминокислот. При гидролизе данногобелка могут образовываться все эти аминокислоты или некоторые из них вопределенных пропорциях, характерных для отдельного белка.
Уникальная последовательностьаминокислотных остатков в цепи, присущая данному белку, называется первичнойструктурой белка. Особенности скручивания цепей белковых молекул(взаимное расположение фрагментов в пространстве) называются вторичнойструктурой белков. Полипептидные цепи белков могут соединяться междусобой с образованием амидных, дисульфидных, водородных и иных связей за счетбоковых цепей аминокислот. В результате этого происходит закручивание спирали вклубок. Эта особенность строения называется третичной структурой белка.Для проявления биологической активности некоторые белки должны сначалаобразовать макрокомплекс (олигопротеин), состоящий из несколькихполноценных белковых субъединиц. Четвертичная структураопределяет степень ассоциации таких мономеров в биологически активномматериале.
Белки делятся на две большие группы — фибриллярные (отношение длины молекулы к ширине больше 10) и глобулярные(отношение меньше 10). К фибриллярным белкам относится коллаген,наиболее распространенный белок позвоночных; на его долю приходится почти 50%сухого веса хрящей и около 30% твердого вещества кости. В большинстверегуляторных систем растений и животных катализ осуществляется глобулярнымибелками, которые носят название ферментов.
Для полипептидов существуетспециальная номенклатура. Аминокислота со свободной аминогруппой на концеполипептидной цепи называется N-концевой аминокислотой, а аминокислота со свободной карбоксильной группой надругом конце этой молекулы — С-концевой аминокислотой.Аминокислотные остатки в полипептидной цепи нумеруются по порядку, начиная с N-концевой аминокислоты, котораяобозначается цифрой 1. Полипептиды называются как производные С-концевойаминокислоты, причем первой пишут название N-концевой аминокислоты, а последней — С-концевуюаминокислоту. Так, глицилаланин — это дипептид, состоящий из глицина (N-концевая аминокислота) и аланина(С-концевая аминокислота). В сокращенном виде название пишут gly-ala. Аланилглицин (ala-gly) — это изомерный дипептид, в котором N-концевой аминокислотой является аланин, а С-концевой — глицин.
АНАЛИЗБЕЛКОВ
Белки, как и прочие амидыгидролизуются под действием кислот и щелочей. После полного гидролизаможно установить качественный и количественный аминокислотный состав, но неточную последовательность аминокислот. Если перед гидролизом обработатьполипептид реактивом Сэнгера, то можно будет затем идентифицировать N-концевую аминокислоту, так как онадаст устойчивое окрашенное производное анилина, которое не разрушается пригидролизе.
/>
В результате неполногогидролиза образуются небольшие пептидные фрагменты. Меняя условиягидролиза, можно разбивать полипептид на различные фрагменты, которыеперекрываются по составляющим их аминокислотным остаткам. Для воссоздания первичнойструктуры необходимо идентифицировать аминокислоты, которые входят в составкаждого из фрагментов, и определить последовательность, в которой этиаминокислоты соединяются друг с другом в исходном полипептиде. Для этого можнопровести полный гидролиз фрагментов, идентифицировать аминокислоты, а затемосуществить химический синтез фрагментов. Существует и другой путь — избирательныйгидролиз. В этом случае от фрагмента отщепляют по одной аминокислоте накаждом этапе, чаще всего при помощи ферментов поджелудочной железы (карбоксипептидаз).Эти ферменты способны гидролизовать только С-концевые аминокислоты и,следовательно, разрушать пептидный фрагмент с С-конца.
N-концевую кислоту можно удалить припомощи фенилизотиоцианата. В результате следующая аминокислота становится N-концевой, ее также можно отщепитьдействием фенилизотиоцианата и т.д. (деградация по Эдману).
/>
В результате реакциифенилизотиоцианата с N-концевойаминокислотой образуется аддукт (тиомочевина), который циклизуется в уксуснойкислоте. При кислотном гидролизе аддукт расщепляется, давая N-концевую аминокислоту и неустойчивоепромежуточное соединение, которое изомеризуется в тиогидантоин. Специфическийтиогидантоин служит для идентификации первой N-концевой аминокислоты. Проводя во второй раз всюпоследовательность реакций, можно определить следующую аминокислоту, так какона стала N-концевой в результате деградации поЭдману.
СИНТЕЗБЕЛКОВ
Реакции, протекающие при синтезе белка- см. Реакции аминокислот. Вышеприведенная схема допускает некоторые изменения- например, для активации карбоксильной группы вместо этилхлорформиатаиспользуют дициклогексилкарбодиимид. В любом случае, синтез белка по этой схеметребует нескольких стадий, что и служит ограничением использования этогометода.
Для синтеза белков стали применятьметод, впервые предложенный Р. Меррифилдом и получивший название твердофазногосинтеза пептидов (ТФСП). На первой стадии синтеза аминокислоту,которая будет стоять на С-конце будущего белка, присоединяют через еекарбоксильную группу к полимеру. При этом нередко используют образованиеэфирной связи между этой группой и хлорметильной группой полимера:
/>
Затем проводят ацилирование свободнойаминогруппы этой аминокислоты путем реакции с дициклогексилкарбодиимидом идругой аминокислотой, которая содержит защищенную аминогруппу. В результатеобразуется первая пептидная связь:
/>
После удаления трет-бутоксикарбонильнойгруппы и нейтрализации полимер, несущий пептид, готов для следующей стадииацилирования:
/>
Вслед за присоединением последнегоаминокислотного остатка аддукт полипептида и полимера обрабатывают смесьюбромистого водорода и трифторуксусной кислоты. В результате полипептидосвобождается от полимера, а с N-концевойаминокислоты снимается защитная группа.
Преимущество этого метода посравнению с классическими методами синтеза состоит в том, что ни на одной изстадий не требуется выделения полипептидной цепи. В силу чрезвычайно низкойрастворимости аддукта пептида и полимера легко отмывается после каждой реакцииот побочных продуктов, растворителей и избытка реагентов без потери пептида,после чего аддукт готов к следующей реакции. В настоящее время методавтоматизирован, и запрограммированные аминокислотные синтезаторы могутбез труда присоединить шесть аминокислот к растущей полипептидной цепи за 24 ч.Эти приборы добавляют реактивы в надлежащей последовательности, меняют условияреакций, обеспечивают необходимое время реакций, отмывают побочные продукты,после чего начинают операцию снова. С помощью метода ТФСП былисинтезированы инсулин и фермент рибонуклеаза, состоящий из124 аминокислот.
www.ronl.ru
Доклад - Белки и аминокислоты
АЗБУКА ЖИВОЙ МАТЕРИИ. БЕЛКИ
Более 4 млрд лет назад на Земле из маленьких неорганических молекул непостижимым образом возникли белки, ставшие строительными блоками живых организмов. Своим бесконечным разнообразием всё живое обязано именно уникальным молекулам белка, и иные формы жизни во Вселенной науке пока неизвестны.
Белки, или протеины (от греч. «протос» — «первый»), — это природные органические соединения, которые обеспечивают все жизненные процессы любого организма. Из белков построены хрусталик глаза и паутина, панцирь черепахи и ядовитые вещества грибов… С помощью белков мы перевариваем пищу и боремся с болезнями. Благодаря особым белкам по ночам светятся светлячки, а в глубинах океана мерцают таинственным светом медузы.
Белковых молекул в живой клетке во много раз больше, чем всех других (кроме воды, разумеется!). Учёные выяснили, что у большинства организмов белки составляют более половины их сухой массы. И разнообразие видов белков очень велико — в одной клетке такого маленького организма, как бактерия Escherichia сой' (см. дополнительный очерк «Объект исследования — прокариоты»), насчитывается около 3 тыс. различных белков.
Впервые белок был выделен (в виде клейковины) в 1728 г. итальянцем Якопо Бартоломео Беккари (1682— 1766) из пшеничной муки. Это событие принято считать рождением химии белка. С тех пор почти за три столетия из природных источников получены тысячи различных белков и исследованы их свойства.

БИОЛОГИЧЕСКИЕ «БУСЫ»

 Молекула белка очень длинная. Химики называют такие молекулы полимерными (от греч. «поли» — «много» и «мерос» — «часть», «доля»). Действительно, длинная молекула полимера состоит из множества маленьких молекул, связанных друг с другом. Так нанизываются на нить бусинки в ожерелье. В полимерах роль нити играют химические связи между бусинками-молекулами.
Молекула белка очень длинная. Химики называют такие молекулы полимерными (от греч. «поли» — «много» и «мерос» — «часть», «доля»). Действительно, длинная молекула полимера состоит из множества маленьких молекул, связанных друг с другом. Так нанизываются на нить бусинки в ожерелье. В полимерах роль нити играют химические связи между бусинками-молекулами.
Секрет белков спрятан в особенностях этих самых бусинок. Большинство полимеров не принимает устойчивой формы в пространстве, уподобляясь тем же бусам, у которых и не может быть пространственной структуры: повесишь их на шею — они примут форму кольца или овала, положишь в коробку — свернутся в клубок неопределённой формы. А теперь представим себе, что некоторые бусинки могут «слипаться» друг с другом. Например, красные притягиваются к жёлтым. Тогда вся цепочка примет определённую форму, обязанную своим существованием «слипа-нию» жёлтых и красных бусинок
Нечто подобное происходит и в белках. Отдельные маленькие молекулы, входящие в состав белка, обладают способностью «слипаться», так как между ними действуют силы притяжения. В результате у любой белковой цепи есть характерная только для неё пространственная структура. Именно она определяет чудесные свойства белков. Без такой структуры они не могли бы выполнять те функции, которые осуществляют в живой клетке.
При длительном кипячении белков в присутствии сильных кислот или щелочей белковые цепи распадаются на составляющие их молекулы,
называемые аминокислотами. Аминокислоты — это и есть те «бусинки», из которых состоит белок, и устроены они сравнительно просто.
КАК УСТРОЕНА АМИНОКИСЛОТА
В каждой молекуле аминокислоты есть атом углерода, связанный с четырьмя заместителями. Один из них — атом водорода, второй — карбоксильная группа —СООН. Она легко «отпускает на волю» ион водорода Н+, благодаря чему в названии аминокислот и присутствует слово «кислота». Третий заместитель — аминогруппа —Nh3 и, наконец, четвёртый заместитель — группа атомов, которую в общем случае обозначают R. У всех аминокислот R-группы разные, и каждая из них играет свою, очень важную роль.
Свойства «бусинок», отличающие одну аминокислоту от другой, скрыты в R-группах (их ещё называют боковыми цепями). Что же касается группы —СООН, то химики-органики относятся к ней с большим почтением: всем другим атомам углерода в молекуле даются обозначения в зависимости от степени их удалённости от карбоксильной группы. Ближайший к ней атом именуют а-атомом, второй — в-атомом, следующий — у-атомом и т. д. Атом углерода в аминокислотах, который находится ближе всех к карбоксильной группе, т. е. а-атом, связан также с аминогруппой, поэтому природные аминокислоты, входящие в состав белка, называют а-аминокислотами.
В природе встречаются также аминокислоты, в которых NH^-группа связана с более отдалёнными от карбоксильной группы атомами углерода. Однако для построения белков природа выбрала именно а-аминокислоты. Это обусловлено прежде всего тем, что только а-аминокислоты, соединённые в длинные цепи, способны обеспечить достаточную прочность и устойчивость структуры больших белковых молекул.
Число а-аминокислот, различающихся R-группой, велико. Но чаще других в белках встречается всего 20 разных аминокислот. Их можно рассматривать как алфавит «языка» белковой молекулы. Химики называют эти главные аминокислоты стандартными, основными или нормальными. Условно основные аминокислоты делят на четыре класса.
В первый входят аминокислоты с неполярными боковыми цепями. Во второй — аминокислоты, содержащие полярную группу. Следующие два составляют аминокислоты с боковыми цепями, которые могут заряжаться положительно (они объединяются в третий класс) или отрицательно (четвёртый). Например, диссоциация карбоксильной группы даёт анион — СОО-, а протонирование атома азота — катион, например —Nh4+. Боковые цепи аспарагиновой и глута-миновой кислот имеют ещё по одной карбоксильной группе —СООН, которая при значениях рН, характерных для живой клетки (рН = 7), расстаётся с ионом водорода (Н+) и приобретает отрицательный заряд. Боковые цепи аминокислот лизина, аргинина и гистидина заряжены положительно, поскольку у них есть атомы азота, которые, наоборот, могут ион водорода присоединять.
Каждая а-аминокислота (кроме глицина) в зависимости от взаимного расположения четырёх заместителей может существовать в двух формах. Они отличаются друг от друга, как предмет от своего зеркального отражения или как правая рука от левой. Такие соединения получили название хоральных (от грен. «хир» — «рука»). Хиральные молекулы открыл в 1848 г. великий французский учёный Луи Пастер. Два типа оптических изомеров органических молекул получили названия Д-форма (от лат. dexter — «правый») и Z-форма (от лат. laevus — «левый»). Кстати, одно из названий других хиральных молекул — глюкозы и фруктозы — декстроза и левулоза. Примечательно, что в состав белков входят только Z-аминокислоты, и вся белковая жизнь на Земле — «левая».
Для нормальной жизнедеятельности организм нуждается в полном наборе из 20 основных a-Z-аминокислот. Но одни из них могут быть синтезированы в клетках самого организма, а другие — должны поступать в готовом виде из пищевых продуктов. В первом случае аминокислоты называют заменимыми, а во втором — незаменимыми. Набор последних для разных организмов различен. Например, для белой крысы незаменимыми являются 10 аминокислот, а для молочнокислых бактерий — 16. Растения могут самостоятельно синтезировать самые разнообразные аминокислоты, создавать такие, которые не встречаются в белках.
Для удобства 20 главных аминокислот обозначают символами, используя одну или первые три буквы русского или английского названия аминокислоты, например аланин — Ала или А, глицин — Гли или G.
ЧТО ТАКОЕ ПЕПТИД
Полимерная молекула белка образуется при соединении в длинную цепочку бусинок-аминокислот. Они нанизываются на нить химических связей благодаря имеющимся у всех аминокислот амино- и карбоксильной группам, присоединённым к а-атому углерода.
Образующиеся в результате такой реакции соединения называются пеп-тидами; (—СО—NH—группировка в них — это пептидная группа, а связь между атомами углерода и азота — пептидная связь (её ещё называют амидной). Соединяя аминокислоты посредством пептидных связей, можно получить пептиды, состоящие из остатков очень многих аминокислот. Такие соединения получили название полипептиды. Полипептидное строение белковой молекулы доказал в 1902 г. немецкий химик Эмиль Герман Фишер.
На концах аминокислотной цепочки находятся свободные амино-и карбоксильная группы; эти концы цепочки называют N- и С-концами. Аминокислотные остатки в полипеп-тидной цепочке принято нумеровать с N-конца.
Общее число аминокислотных остатков в белковой молекуле изменяется в очень широких пределах. Так, человеческий инсулин состоит из 51 аминокислотного остатка, а лизо-цим молока кормящей матери — из 130. В гемоглобине человека 4 аминокислотные цепочки, каждая из которых построена из примерно 140 аминокислот. Существуют белки, имеющие почти 3 тыс. аминокислотных остатков в единой цепи.
Молекулярные массы белков лежат в диапазоне примерно от 11 тыс. для малых белков, состоящих из 100 аминокислотных остатков, до 1 млн и более для белков с очень длинными полипептидными цепями или для белков, состоящих из нескольких по-липептидных цепей.
Возникает вопрос: как же всё огромное многообразие белков с различными функциями и свойствами может быть создано всего из 20 молекул? А разгадка этого секрета природы проста — каждый белок имеет свой неповторимый аминокислотный состав и уникальный порядок соединения аминокислот, называемый первичной структурой белка.
СПИРАЛИ И СЛОИ
В начале 50-х гг. XX в. американские химики Лайнус Карл Полинг (1901— 1994), награждённый Нобелевской премией за исследования природы химической связи, и Роберт Кори (1897—1971) предположили, что некоторые участки аминокислотной цепочки в белках закручены в спираль. Благодаря совершенствованию экспериментальных методов (структуру белков изучают с помощью рентгеновских лучей) через несколько лет эта гениальная догадка подтвердилась.
Действительно, полипептидные цепи очень часто образуют спираль, закрученную в правую сторону. Это первый, самый низкий уровень пространственной организации белковых цепочек Здесь-то и начинают играть роль слабые взаимодействия «бусинок»-аминокислот: группа С=0 и группа N—H из разных пептидных связей могут образовывать между собой водородную связь. Оказалось, что в открытой Полингом и Кори спирали такая связь образована между группой С=0 каждой г-й аминокислоты и группой N—H (i + 4)-й аминокислоты, т. е. между собой связаны аминокислотные остатки, отстоящие друг от друга на четыре «бусинки». Эти водородные связи и стабилизируют такую спираль в целом. Она получила название a.-спирали.
Позднее выснилось, что а-спираль — не единственный способ укладки аминокислотных цепочек. Помимо спиралей они образуют ещё и слои. Благодаря всё тем же водородным связям между группами С=0 и N—H друг с другом могут «слипаться» сразу несколько разных фрагментов одной полипептидной цепи. В результате получается целый слой — его назвали ^-слоем.
В большинстве белков а-спирали и р-слои перемежаются всевозможными изгибами и фрагментами цепи без какой-либо определённой структуры. Когда имеют дело с пространственной структурой отдельных участков белка, говорят о вторичной структуре белковой молекулы.
БЕЛОК В ПРОСТРАНСТВЕ
Для того чтобы получить полный «портрет» молекулы белка, знания первичной и вторичной структуры недостаточно. Эти сведения ещё не дают представления ни об объёме, ни о форме молекулы, ни тем более о расположении участков цепи по отношению друг к другу. А ведь все спирали и слои каким-то образом размещены в пространстве. Общая пространственная структура поли-пептидной цепи называется третичной структурой белка.
Первые пространственные модели молекул белка — миоглобина и гемоглобина — построили в конце 50-х гг. XX в. английские биохимики Джон Ко-удери Кендрю (родился в 1917 г.) и Макс Фердинанд Перуц (родился в 1914 г.). При этом они использовали данные экспериментов с рентгеновскими лучами. За исследования в области строения белков Кендрю и Перуц в 1962 г. были удостоены Нобелевской премии. А в конце столетия была определена третичная структура уже нескольких тысяч белков.
При образовании третичной структуры белка наконец-то проявляют активность R-группы — боковые цепи аминокислот. Именно благодаря им «слипаются» между собой большинство «бусинок»-аминокислот, придавая цепи определённую форму в пространстве.
В живом организме белки всегда находятся в водной среде. А самое большое число основных аминокислот — восемь — содержат неполярные R-группы. Разумеется, белок стремится надёжно спрятать внутрь своей молекулы неполярные боковые цепи, чтобы ограничить их контакт с водой. Учёные называют это возникновением гидрофобных взаимодействий (см. статью «Мельчайшая единица живого»).
Благодаря гидрофобным взаимодействиям вся полипептидная цепочка принимает определённую форму в пространстве, т. е. образует третичную структуру.
В молекуле белка действуют и другие силы. Часть боковых цепей основных аминокислот заряжена отрицательно, а часть — положительно. Так как отрицательные заряды притягиваются к положительным, соответствующие «бусинки» «слипаются». Электростатические взаимодействия, или, как их называют иначе, солевые мостики, — ещё одна важная сила, стабилизирующая третичную структуру.
У семи основных аминокислот есть полярные боковые цепи. Между ними могут возникать водородные связи, тоже играющие немалую роль в поддержании пространственной структуры белка.
Между двумя аминокислотными остатками цистеина иногда образуются ковалентные связи (—S—S—), которые очень прочно фиксируют расположение разных участков белковой цепи по отношению друг к другу. Такие связи называют дисуль-фидными мостиками. Это самые немногочисленные взаимодействия в белках (в некоторых случаях они вообще отсутствуют), зато по прочности они не имеют равных.
ВЫСШИЙ УРОВЕНЬ ПРОСТРАНСТВЕННОЙ ОРГАНИЗАЦИИ БЕЛКОВ
Молекула белка может состоять не из одной, а из нескольких полипептидных цепей. Каждая такая цепь представляет собой самостоятельную пространственную структуру — субь-единицу. Например, белок гемоглобин состоит из четырёх субъединиц, которые образуют единую молекулу, располагаясь в вершинах почти правильного тетраэдра. Субъединицы «прилипают» друг к другу благодаря тем же самым силам, что стабилизируют третичную структуру. Это гидрофобные взаимодействия, солевые мостики и водородные связи.
Если белок состоит из нескольких субъединиц, говорят, что он обладает четвертичной структурой. Такая структура представляет собой высший уровень организации белковой молекулы. В отличие от первых трёх уровней четвертичная структура есть далеко не у всех белков. Приблизительно половина из известных на сегодняшний день белков её не имеют.
ПОЧЕМУ БЕЛКИ БОЯТСЯ ТЕПЛА
Связи, поддерживающие пространственную структуру белка, довольно легко разрушаются. Мы с детства знаем, что при варке яиц прозрачный яичный белок превращается в упругую белую массу, а молоко при скисании загустевает. Происходит это из-за разрушения пространственной структуры белков альбумина в яичном белке и казеина (огглат. caseus — «сыр») в молоке. Такой процесс называется денатурацией. В первом случае её вызывает нагревание, а во втором — значительное увеличение кислотности (в результате жизнедеятельности обитающих в молоке бактерий). При денатурации белок теряет способность выполнять присущие ему в организме функции (отсюда и название процесса: от лат. denaturare — «лишать природных свойств»). Денатурированные белки легче усваиваются организмом, поэтому одной из целей термической обработки пищевых продуктов является денатурация белков.
ЗАЧЕМ НУЖНА ПРОСТРАНСТВЕННАЯ СТРУКТУРА
В природе почти ничего не происходит случайно. Если белок принял определённую форму в пространстве, это должно служить достижению какой-то цели. Действительно, только белок с «правильной» пространственной структурой может обладать определёнными свойствами, т. е. выполнять те функции в организме, которые ему предписаны. А делает он это с помощью всё тех же R-групп аминокислот. Оказывается, боковые цепи не только поддерживают «правильную» форму молекулы белка в пространстве. R-группы могут связывать другие органические и неорганические молекулы, принимать участие в химических реакциях, выступая, например, в роли катализатора.
Часто сама пространственная организация полипептидной цепи как раз' и нужна для того, чтобы сосредоточить в определённых точках пространства необходимый для выполнения той или иной функции набор боковых цепей. Пожалуй, ни один процесс в живом организме не проходит без участия белков.
В ЧЁМ СЕКРЕТ ФЕРМЕНТОВ
Все химические реакции, протекающие в клетке, происходят благодаря особому классу белков — ферментам. Это белки-катализаторы. У них есть свой секрет, который позволяет им работать гораздо эффективнее других катализаторов, ускоряя реакции в миллиарды раз.
Предположим, что несколько приятелей никак не могут встретиться. Но стоило одному из них пригласить друзей на день рождения, как результат не заставил себя ждать: все оказались в одном месте в назначенное время.
Чтобы встреча состоялась, понадобилось подтолкнуть друзей к контакту. То же самое делает и фермент. В его молекуле есть так называемые цгнтры связывания. В них расположены привлекательные для определённого типа химических соединений (и только для них!) «уютные кресла» — R-группы, связывающие какие-то участки молекул реагирующих веществ. Например, если одна из молекул имеет неполярную группу, в центре связывания находятся гидрофобные боковые цепи. Если же в молекуле есть отрицательный заряд, его будет поджидать в молекуле фермента R-группа с положительным зарядом.
В результате обе молекулы реагентов связываются с ферментом и оказываются в непосредственной близости друг от друга. Мало того, те их группы, которые должны вступить в химическую реакцию, сориентированы в пространстве нужным для реакции образом. Теперь за дело принимаются боковые цепи фермента, играющие роль катализаторов. В ферменте все «продумано» таким образом, что R-группы-катализаторы тоже расположены вблизи от места событий, которое называют активным центром. А после завершения реакции фермент «отпускает на волю» молекулы-продукты (см. статью «Ферменты — на все руки мастера»).
ОТКУДА БЕРЁТСЯ ИММУНИТЕТ
Белки выполняют в организме множество функций; они, например, защищают клетки от нежелательных вторжений, предохраняют их от повреждений. Специальные белки — антитела обладают способностью распознавать проникшие в клетки бактерии, вирусы, чужеродные полимерные молекулы и нейтрализовывать их.
У высших позвоночных от чужеродных частиц организм защищает иммунная система. Она устроена так, что организм, в который вторглись такие «агрессоры» — антигены, начинает вырабатывать антитела. Молекула антитела прочно связывается с антигеном: у антител, как и у ферментов, тоже есть центры связывания. Боковые цепи аминокислот расположены в центрах таким образом, что антиген, попавший в эту ловушку, уже не сможет вырваться из «железных лап» антитела. После связывания с антителом враг выдворяется за пределы организма.
Можно ввести в организм небольшое количество некоторых полимерных молекул, входящих в состав бактерий или вирусов-возбудителей какой-либо инфекционной болезни.
В организме немедленно появятся соответствующие антитела. Теперь попавший в кровь или лимфу «настоящий» болезнетворный микроб тотчас же подвергнется атаке этих антител, и болезнь будет побеждена. Такой способ борьбы с инфекцией есть не что иное, как нелюбимая многими прививка. Благодаря ей организм приобретает иммунитет к инфекционным болезням.
ДЛЯ ЧЕГО В ГЕМОГЛОБИНЕ ЖЕЛЕЗО
В природе существуют белки, в которых помимо аминокислот содержатся другие химические компоненты, такие, как липиды, сахара, ионы металлов. Обычно эти компоненты играют важную роль при выполнении белком его биологической функции. Так, перенос молекул и ионов из одного органа в другой осуществляют транспортные белки плазмы крови. Белок гемоглобин (от греч. «гема» — «кровь» и лат. globus — «шар», «шарик»), содержащийся в кровяных клетках — эритроцитах (от греч. «эритрос» — «красный» и «китос» — «клетка»), доставляет кислород от лёгких к тканям. В молекуле гемоглобина есть комплекс иона железа Fe24" со сложной органической молекулой, называемый гемам. Гемоглобин состоит из четырёх белковых субъединиц, и каждая из них содержит по одному гему.
В связывании кислорода в лёгких принимает участие непосредственно ион железа. Как только к нему хотя бы в одной из субъединиц присоединяется кислород, сам ион тут же чуть-чуть меняет своё расположение в молекуле белка. Движение железа «провоцирует» движение всей аминокислотной цепочки данной субъединицы, которая слегка трансформирует свою третичную структуру. Другая субъединица, ещё не присоединившая кислород, «чувствует», что произошло с соседкой. Её структура тоже начинает меняться. В итоге вторая субъединица связывает кислород легче, чем первая. Присоединение кислорода к третьей и четвёртой субъединицам происходит с ещё меньшими трудностями. Как видно, субъединицы помогают друг другу в работе. Для этого-то гемоглобину и нужна четвертичная структура. Оксид углерода СО (в просторечии угарный газ) связывается с железом в геме в сотни раз прочнее кислорода. Угарный газ смертельно опасен для человека, поскольку лишает гемоглобин возможности присоединять кислород.
А ЕЩЁ БЕЛКИ...
… Служат питательными веществами. В семенах многих растений (пшеницы, кукурузы, риса и др.) содержатся пищевые белки. К ним относятся также альбумин — основной компонент яичного белка и казеин — главный белок молока. При переваривании в организме человека белковой пищи происходит гидролиз пептидных связей. Белки «разбираются» на отдельные аминокислоты, из которых организм в дальнейшем «строит» новые пептиды или использует для получения энергии. Отсюда и название:
греческое слово «пептос» означает «переваренный». Интересно, что гидролизом пептидной связи управляют тоже белки — ферменты.
… Участвуют в регуляции клеточной и физиологической активности. К подобным белкам относятся многие гормоны (от греч. «гормао» — «побуждаю»), такие, как инсулин, регулирующий обмен глюкозы, и гормон роста.
… Наделяют организм способностью изменять форму и передвигаться. За это отвечают белки актин и миозин, из которых построены мышцы.
… Выполняют опорную и защитную функции, скрепляя биологические структуры и придавая им прочность. Кожа представляет собой почти чистый белок коллаген, а волосы, ногти и перья состоят из прочного нерастворимого белка кератина.
ЧТО ЗАПИСАНО В ГЕНАХ
Последовательность аминокислот в белках кодируется генами, которые хранятся и передаются по наследству с помощью молекул ДНК (см. статьи «Хранитель наследственной информации. ДНК» и «Экспрессия генов»). Пространственную структуру белка задаёт именно порядок расположения аминокислот. Получается, что не только первичная, но и вторичная, третичная и четвертичная структуры белков составляют содержание наследственной информации. Следовательно, и выполняемые белками функции запрограммированы генетически. Громадный перечень этих функций позволяет белкам по праву называться главными молекулами жизни. Поэтому сведения о белках и есть то бесценное сокровище, которое передаётся в природе от поколения к поколению.
Интерес человека к этим органическим соединениям с каждым годом только увеличивается. Сегодня учёные уже расшифровали структуру многих белковых молекул. Они выясняют функции самых разных белков, пытаются определить взаимосвязь функций со структурой. Установление сходства и различий у белков, выполняющих аналогичные функции у разных живых организмов, позволяет глубже проникать в тайны эволюции.
АМИНОКИСЛОТЫ — ПОКАЗАТЕЛИ ВОЗРАСТА
D- и L-формы аминокислот обладают способностью очень медленно превращаться друг в друга. За определённый (весьма длительный) период времени чистая D- или I-форма может стать смесью равных количеств обеих форм. Такая смесь называется раиемагом, а сам процесс —раие-мизаиией. Скорость рацемизации зависит от температуры и типа аминокислоты. Данное свойство можно использовать для определения возраста ископаемых остатков организмов, а при необходимости — и живых существ. Например, в белке дентина (дентин — костная ткань зубов) 1-ас-парагиновая кислота самопроизвольно раиемизуется со скоростью 0,1 % в год. У детей в период формирования зубов в дентине содержится только 1-аспарагиновая кислота. Дентин выделяют из зуба и определяют В нём содержание 0-формы. Результаты теста достаточно точны. Так, для 97-летней женщины, возраст которой был документально засвидетельствован, тест показал возраст 99 лет. Данные исследований, выполненных на ископаемых остатках доисторических животных — слонов, дельфинов, медведей, — хорошо согласуются с результатами датирования, полученными радионуклидным методом.
ЗА ЧТО СЕНГЕР ПОЛУЧИЛ НОБЕЛЕВСКИЕ ПРЕМИИ
При гидролизе белков до аминокислот (разрушении пептидной связи водой) теряется информация о последовательности их соединения. Поэтому долгое время считали, что определение первичной структуры белка представляет собой совершенно безнадежную задачу. Но в 50-х гг. XX в. английский биохимик Фредерик Сенгер (родился в 1918 г.) смог расшифровать последовательность аминокислот в полипептидных цепях гормона инсулина. За эту работу, на выполнение которой ушло несколько лет, в 1958 г. Сенгер был удостоен Нобелевской премии по химии (двадцатью годами позже он совместно с У. Гилбертом получил вторую премию за вклад в установление первичной структуры ДНК).
Принципы определения аминокислотной последовательности, впервые сформулированные Сенгером, используются и ныне, правда, со всевозможными вариациями и усовершенствованиями. Процедура установления первичной структуры белка сложна и многоступенчата: в ней около десятка различных стадий. Сначала белок расщепляют до отдельных аминокислот и устанавливают их тип и количество в данном веществе. На следующей стадии длинную белковую молекулу расщепляют уже не полностью, а на фрагменты. Затем в этих фрагментах определяют порядок соединения аминокислот, последовательно отделяя их одну за другой. Расшепление белка на фрагменты проводят несколькими способами, чтобы в разных фрагментах были перекрывающиеся участки. Выяснив порядок расположения аминокислот во всех фрагментах, получают полную информацию о том, как аминокислоты расположены в белке. К концу XX в. созданы специальные приборы, определяющие последовательность аминокислот в молекуле белка в автоматическом режиме — секвенаторы (от англ. sequence — «последовательность»).
молоко
И КИСЛОМОЛОЧНЫЕ ПРОДУКТЫ
Молоко представляет собой коллоидный раствор жира в воде. Под микроскопом хорошо видно, что оно неоднородно: в бесцветном растворе (сыворотке) плавают жировые шарики.
В коровьем молоке обычно содержится от 3 до 6 % жиров (в основном это сложные эфиры глицерина и насыщенных карбоновых кислот — пальмитиновой, стеариновой), около 3 % белков, а ешё углеводы, органические кислоты, витамины и минеральные вещества.
Белок казеин в молоке присутствует в связанном виде — ковалентно присоединённые к аминокислоте сери-ну фосфатные группы образуют соли с ионами кальция. При подкислении молока эти соли разрушаются, и казеин выделяется в виде белой творожистой массы. В желудке человека под действием особых ферментов происходит процесс, называемый “створажива-нием казеина”. Створоженный казеин выпадает в осадок и медленнее выводится из организма, а потому полнее усваивается. Казеин высоко питателен:
в нём есть почти все аминокислоты, необходимые человеку для построения собственных белков. В чистом виде он представляет собой безвкусный белый порошок, не растворимый в воде. Помимо него в молоке содержатся и другие белки, например лактальбумин. При кипячении этот белок превращается в нерастворимую форму, образуя на поверхности кипячёного молока характерную белую плёнку — пенку.
Входящий в состав молока сахар лактоза С^НддО,, изомерен сахарозе. В организме человека под действием фермента лактазы этот сахар расщепляется на моносахариды глюкозу и галактозу, которые легко усваиваются. За счёт этого, например, грудные дети пополняют запасы углеводов. Интересно, что у многих людей (в основном у представителей монголоидной расы) организм в зрелом возрасте утрачивает способность расщеплять лактозу.
Проходя через пищеварительный тракт, лактоза не усваивается, а становится питательной средой для развития различных болезнетворных микроорганизмов, что приводит к общему недомоганию. Именно поэтому народы Дальнего Востока (японцы, китайцы) практически не употребляют в пишу молочные продукты.
В промышленных условиях молоко подвергают тепловой обработке, цель которой — подавить развитие микроорганизмов и продлить срок его хранения. Для этого молоко пастеризуют — выдерживают 30 мин при 65 °С, а также используют кратковременную термообработку — нагревают в течение 10-20 с до 71 °С. По сравнению с пастеризацией термообработка лучше сохраняет питательные вещества, в первую очередь витамины. Чтобы молоко не расслаивалось на сливки и сыворотку, его гомогенизируют — пропускают под давлением через небольшие отверстия. Жировые шарики дробятся, уменьшаются в размерах, а молоко становится более вязким.
Значительная часть молока идёт на переработку — для производства сливочного масла, сыра и кисломолочных продуктов (кефира, ряженки, простокваши, сметаны).
Чтобы получить кефир, молоко сквашивают — выдерживают в течение 8-10 ч при 20-25 °С, добавляя затравку молочнокислых бактерий. Под их действием лактоза распадается до молочной кислоты:
с„н„о„ + н, о =
лактоза
== 4СНзСН(ОН)СООН. молочная (2-гидроксипропановая) кислота
Именно молочная кислота определяет специфический вкус кефира. По мере того как она накапливается в растворе, происходит коагуляция (свёртывание)казеина, который выделяется в свободном виде. Поэтому кефир имеет более густую консистенцию, чем молоко. Молочнокислое сбраживание лактозы сопровождается спиртовым брожением, из-за чего в кисломолочных продуктах, в частности в кефире, есть небольшое количество алкоголя (до 0,03 %). В кисломолочных продуктах содержатся также микроорганизмы, которые подавляют развитие болезнетворных бактерий и тем самым улучшают пишеварение.
Творог тоже получают сквашиванием молока молочнокислыми бактериями. Его главной составной частью является белок казеин.
Чтобы приготовить сливочное масло, от молочной сыворотки необходимо отделить капельки жира, входящие в состав молока. Для этого сбивают сливки — верхний, более жирный слой, образующийся при отстаивании молока.
Казеин входит также в состав сыров. Их делают, добавляя в молоко бактериальную закваску и специальные ферменты, а затем подогревая смесь до определённой температуры. В выделившийся сгусток вновь вводят ферменты и подогревают. При этом происходит частичное изменение структуры и состава казеина. Затем смесь раскладывают по формам и длительное время — до шести месяцев — выдерживают при низкой температуре (не выше 15 °С). Во время созревания казеин под действием ферментов распадается на поли-пептиды и свободные аминокислоты. Часть аминокислот окисляется кислородом воздуха, при этом образуются аммиак, альдегиды, а также кетокислоты, придающие сыру характерный аромат.
Скисание молока — привычный пример денатурации белка.
МЕДНАЯ КРОВЬ
В холодных водах Перуанского течения в Тихом океане обитает кальмар Dosidicus gigas. Его сигарообразное тело вместе со щупальцами достигает в длину 3,5 м, а масса гиганта может превышать 150 кг. Мощные мышиы выбрасывают струю воды с силой, с какой она бьёт из пожарного рукава, благодаря чему кальмар способен двигаться со скоростью до 40 км/ч. Клювом, очень крепким и острым, он может перебить стальной кабель. По свидетельству очевидцев, кальмар буквально в клочья раздирает 20-килограммовую рыбину. Этот свирепый хишник очень опасен и для человека. В книге Франка Лейна “Царство осьминога” утверждается, что “человек, упавший за борт в местах, где обитает много кальмаров, не проживёт и полминуты”.
Чтобы “зарядиться” энергией, этому обитателю океана требуется много кислорода — не менее 50 л в час. По-ступаюший из морской воды кислород разносится по телу кальмара с помошью особого белка, содержащего медь, — гемоиианина (от греч. “гема” — “кровь” и “кианос” — “лазурный”, “голубой”).
Стоит заметить, что в крови позвоночных кислород “транспортируют” атомы железа в составе гема — особой сложной молекулы, которая входит в состав белка гемоглобина. Им буквально нашпигованы красные кровяные клетки — эритроциты. Молекула гемоглобина содержит четыре гемовых фрагмента, каждый из которых способен связать молекулу кислорода. В отличие от гемоглобина, в гемоиианине атомы меди непосредственно связаны с белковыми молекулами, которые не включены ни в какие клетки, а свободно “плавают” в крови. Зато одна молекула гемоииани
на способна связать до 200 атомов меди. И ешё одна особенность гемоииани-на — его молекулы имеют огромные даже для белков размеры. У “обычных” белков, входящих в состав яиц, молока, мыши, молекулярная масса колеблется в пределах от б тыс. до 1 млн, а молекулярная масса гемоиианина может достигать 10 млн! Это один из самых крупных белков; больше по размеру и массе только белковые комплексы у вирусов.
Гемоиианин — очень древний белок. Он устроен проще, чем гемоглобин и не так эффективен. Тем не менее при малом содержании кислорода в морской воде гемоиианин довольно успешно снабжает им ткани холоднокровных животных. Так, давление кислорода в жабрах лангуста составляет всего 7 мм рт. ст. (930 Па), а в тканях — 3 мм рт. ст.; причём концентрация этого газа в крови лангуста в 20 раз выше, чем в морской воде.
Кроме кальмаров, кислород переносится “голубой кровью” также у де-сятиногих ракообразных (омары, крабы, креветки). Гемоиианин найден у всех головоногих моллюсков (осьминоги, кальмары, каракатицы), разнообразных улиток, пауков и др. А вот у морских гребешков, устриц и других двустворчатых моллюсков его нет.
Количество гемоиианина в крови может быть самым разным. Так, у шустрых осьминога и мечехвоста (морское животное типа членистоногих) концентрация этого необычного белка доходит до 10 г в 100 мл крови — почти столько же гемоглобина в крови человека. В то же время, у малоподвижного съедобного моллюска морское ушко Hatiotis tuberculataв 100 мл крови всего 0,03 г гемоиианина. Это и понятно: чем более активно животное,
чем больше кислорода необходимо ему для восполнения энергетических затрат, тем выше в крови концентрация белка, переносящего кислород.
Гемоиианин был открыт в 60-х гг. XIX в., когда биологи заметили, что кровь головоногих моллюсков при прохождении через жабры окрашивается в голубой цвет. А в 1878 г. бельгийский физиолог Леон Фредерик доказал, что голубой цвет вызван реакцией кислорода с медьсодержащим белком, который он назвал гемоиианином. Когда последний теряет кислород, он, в отличие от гемоглобина, становится бесцветным. Примечательно, что всю работу по изучению нового белка Фредерик выполнил в течение одного дня.
Из гемоиианина нетрудно полностью извлечь медь. Аля этого достаточно обработать белок в отсутствие кислорода реактивом, который прочно связывается с ионами одновалентной меди. Таким же способом можно определить содержание меди в гемоиианине. Лишённый этого металла, он теряет способность переносить кислород. Но если потом ввести в раствор белка ионы Си«1', гемоиианин восстанавливает свою физиологическую активность. Так было доказано, что в отсутствие кислорода медь гемоиианина находится в степени окисления +1. При избытке же этого газа происходит частичное окисление металла. При этом всегда на одну связанную гемоиианином молекулу кислорода приходится два атома меди. Таким образом, кислород окисляет ровно половину атомов меди. Это ещё одно отличие гемоиианина от значительно более распространённого в животном мире гемоглобина, в котором все атомы железа равноценны и имеют заряд +2 как в свободном состоянии, так и в комплексе с кислородом.
Список использованной литературы:
1) Энциклопедия для детей “Аванта+”.Том17. Химия. Москва 2000. (стр480-293)
www.ronl.ru

|
|
..:::Счетчики:::.. |
|
|
|
|
|
|
|
|