
|
|
|
|
 Far Far |
| WinNavigator |
| Frigate |
| Norton
Commander |
| WinNC |
| Dos
Navigator |
| Servant
Salamander |
| Turbo
Browser |
|
|
| Winamp,
Skins, Plugins |
| Необходимые
Утилиты |
| Текстовые
редакторы |
| Юмор |
|
|

|
File managers and best utilites |
Реферат: Метаболизм. Реферат метаболизм
Реферат - Метаболизм - Медицина
Метаболизм, или обмен веществ, химические превращения, протекающие от момента поступления питательных веществ в живой организм до момента, когда конечные продукты этих превращений выделяются во внешнюю среду. К метаболизму относятся все реакции, в результате которых строятся структурные элементы клеток и тканей, и процессы, в которых из содержащихся в клетках веществ извлекается энергия. Иногда для удобства рассматривают по отдельности две стороны метаболизма – анаболизм и катаболизм, т.е. процессы созидания органических веществ и процессы их разрушения. Анаболические процессы обычно связаны с затратой энергии и приводят к образованию сложных молекул из более простых, катаболические же сопровождаются высвобождением энергии и заканчиваются образованием таких конечных продуктов (отходов) метаболизма, как мочевина, диоксид углерода, аммиак и вода.
Термин «обмен веществ» вошел в повседневную жизнь с тех пор, как врачи стали связывать избыточный или недостаточный вес, чрезмерную нервозность или, наоборот, вялость больного с повышенным или пониженным обменом. Для суждения об интенсивности метаболизма ставят тест на «основной обмен». Основной обмен – это показатель способности организма вырабатывать энергию. Тест проводят натощак в состоянии покоя; измеряют поглощение кислорода (О2) и выделение диоксида углерода (СО2). Сопоставляя эти величины, определяют, насколько полно организм использует («сжигает») питательные вещества. На интенсивность метаболизма влияют гормоны щитовидной железы, поэтому врачи при диагностике заболеваний, связанных с нарушениями обмена, в последнее время все чаще измеряют уровень этих гормонов в крови.
Методы исследования. При изучении метаболизма какого-нибудь одного из питательных веществ прослеживают все его превращения от той формы, в какой оно поступает в организм, до конечных продуктов, выводимых из организма. В таких исследованиях применяется крайне разнообразный набор биохимических методов.
Использование интактных животных или органов. Животному вводят изучаемое соединение, а затем в его моче и экскрементах определяют возможные продукты превращений (метаболиты) этого вещества. Более определенную информацию можно получить, исследуя метаболизм определенного органа, например печени или мозга. В этих случаях вещество вводят в соответствующий кровеносный сосуд, а метаболиты определяют в крови, оттекающей от данного органа.
Поскольку такого рода процедуры сопряжены с большими трудностями, часто для исследования используют тонкие срезы органов. Их инкубируют при комнатной температуре или при температуре тела в растворах с добавкой того вещества, метаболизм которого изучают. Клетки в таких препаратах не повреждены, и так как срезы очень тонкие, вещество легко проникает в клетки и легко выходит из них. Иногда затруднения возникают из-за слишком медленного прохождения вещества сквозь клеточные мембраны. В этих случаях ткани измельчают, чтобы разрушить мембраны, и с изучаемым веществом инкубируют клеточную кашицу. Именно в таких опытах было показано, что все живые клетки окисляют глюкозу до СО2 и воды и что только ткань печени способна синтезировать мочевину.
Использование клеток. Даже клетки представляют собой очень сложно организованные системы. В них имеется ядро, а в окружающей его цитоплазме находятся более мелкие тельца, т.н. органеллы, различных размеров и консистенции. С помощью соответствующей методики ткань можно «гомогенизировать», а затем подвергнуть дифференциальному центрифугированию (разделению) и получить препараты, содержащие только митохондрии, только микросомы или прозрачную жидкость – цитоплазму. Эти препараты можно по отдельности инкубировать с тем соединением, метаболизм которого изучается, и таким путем установить, какие именно субклеточные структуры участвуют в его последовательных превращениях. Известны случаи, когда начальная реакция протекает в цитоплазме, ее продукт подвергается превращению в микросомах, а продукт этого превращения вступает в новую реакцию уже в митохондриях. Инкубация изучаемого вещества с живыми клетками или с гомогенатом ткани обычно не выявляет отдельные этапы его метаболизма, и только последовательные эксперименты, в которых для инкубации используются те или иные субклеточные структуры, позволяют понять всю цепочку событий.
Использование радиоактивных изотопов. Для изучения метаболизма какого-либо вещества необходимы: 1) соответствующие аналитические методы для определения этого вещества и его метаболитов; и 2) методы, позволяющие отличать добавленное вещество от того же вещества, уже присутствующего в данном биологическом препарате. Эти требования служили главным препятствием при изучении метаболизма до тех пор, пока не были открыты радиоактивные изотопы элементов и в первую очередь радиоактивный углерод 14C. С появлением соединений, «меченных» 14C, а также приборов для измерения слабой радиоактивности эти трудности были преодолены. Если к биологическому препарату, например к суспензии митохондрий, добавляют меченную 14C жирную кислоту, то никаких специальных анализов для определения продуктов ее превращений не требуется; чтобы оценить скорость ее использования, достаточно просто измерять радиоактивность последовательно получаемых митохондриальных фракций. Эта же методика позволяет легко отличать молекулы радиоактивной жирной кислоты, введенной экспериментатором, от молекул жирной кислоты, уже присутствовавших в митохондриях к началу эксперимента.
Хроматография и электрофорез. В дополнение к вышеупомянутым требованиям биохимику необходимы и методы, позволяющие разделять смеси, состоящие из малых количеств органических веществ. Важнейший из них – хроматография, в основе которой лежит феномен адсорбции. Разделение компонентов смеси проводят при этом либо на бумаге, либо путем адсорбции на сорбенте, которым заполняют колонки (длинные стеклянные трубки), с последующей постепенной элюцией (вымыванием) каждого из компонентов.
Разделение методом электрофореза зависит от знака и числа зарядов ионизированных молекул. Электрофорез проводят на бумаге или на каком-нибудь инертном (неактивном) носителе, таком, как крахмал, целлюлоза или каучук.
Высокочувствительный и эффективный метод разделения – газовая хроматография. Им пользуются в тех случаях, когда подлежащие разделению вещества находятся в газообразном состоянии или могут быть в него переведены.
Выделение ферментов. Последнее место в описываемом ряду – животное, орган, тканевой срез, гомогенат и фракция клеточных органелл – занимает фермент, способный катализировать определенную химическую реакцию. Выделение ферментов в очищенном виде – важный раздел в изучении метаболизма.
Сочетание перечисленных методов позволило проследить главные метаболические пути у большей части организмов (в том числе у человека), установить, где именно эти различные процессы протекают, и выяснить последовательные этапы главных метаболических путей. К настоящему времени известны тысячи отдельных биохимических реакций, изучены участвующие в них ферменты.
Клеточный метаболизм. Живая клетка – это высокоорганизованная система. В ней имеются различные структуры, а также ферменты, способные их разрушить. Содержатся в ней и крупные макромолекулы, которые могут распадаться на более мелкие компоненты в результате гидролиза (расщепления под действием воды). В клетке обычно много калия и очень мало натрия, хотя клетка существует в среде, где натрия много, а калия относительно мало, и клеточная мембрана легко проницаема для обоих ионов. Следовательно, клетка – это химическая система, весьма далекая от равновесия. Равновесие наступает только в процессе посмертного автолиза (самопереваривания под действием собственных ферментов).
Потребность в энергии. Чтобы удержать систему в состоянии, далеком от химического равновесия, требуется производить работу, а для этого необходима энергия. Получение этой энергии и выполнение этой работы – непременное условие для того, чтобы клетка оставалась в своем стационарном (нормальном) состоянии, далеком от равновесия. Одновременно в ней выполняется и иная работа, связанная со взаимодействием со средой, например: в мышечных клетках – сокращение; в нервных клетках – проведение нервного импульса; в клетках почек – образование мочи, значительно отличающейся по своему составу от плазмы крови; в специализированных клетках желудочно-кишечного тракта – синтез и выделение пищеварительных ферментов; в клетках эндокринных желез – секреция гормонов; в клетках светляков – свечение; в клетках некоторых рыб – генерирование электрических разрядов и т.д.
Источники энергии. В любом из перечисленных выше примеров непосредственным источником энергии, которую клетка использует для производства работы, служит энергия, заключенная в структуре аденозинтрифосфата (АТФ). В силу особенностей своей структуры это соединение богато энергией, и разрыв связей между его фосфатными группами может происходить таким образом, что высвобождающаяся энергия используется для производства работы. Однако энергия не может стать доступной для клетки при простом гидролитическом разрыве фосфатных связей АТФ: в этом случае она расходуется впустую, выделяясь в виде тепла. Процесс должен состоять из двух последовательных этапов, в каждом из которых участвует промежуточный продукт, обозначенный здесь X–Ф (в приведенных уравнениях X и Y означают два разных органических вещества; Ф – фосфат; АДФ – аденозиндифосфат):
/>
Поскольку практически для любого проявления жизнедеятельности клеток необходим АТФ, неудивительно, что метаболическая активность живых клеток направлена в первую очередь на синтез АТФ. Этой цели служат различные сложные последовательности реакций, в которых используется потенциальная химическая энергия, заключенная в молекулах углеводов и жиров (липидов).
Метаболизм углеводов и липидов
Синтез АТФ. Анаэробный (без участия кислорода). Главная роль углеводов и липидов в клеточном метаболизме состоит в том, что их расщепление на более простые соединения обеспечивает синтез АТФ. Несомненно, что те же процессы протекали и в первых, самых примитивных клетках. Однако в атмосфере, лишенной кислорода, полное окисление углеводов и жиров до CO2 было невозможно. У этих примитивных клеток имелись все же механизмы, с помощью которых перестройка структуры молекулы глюкозы обеспечивала синтез небольших количеств АТФ. Речь идет о процессах, которые у микроорганизмов называют брожением. Лучше всего изучено сбраживание глюкозы до этилового спирта и CO2 у дрожжей.
/>
В ходе 11 последовательных реакций, необходимых для того, чтобы завершилось это превращение, образуется ряд промежуточных продуктов, представляющих собой эфиры фосфорной кислоты (фосфаты). Их фосфатная группа переносится на аденозиндифосфат (АДФ) с образованием АТФ. Чистый выход АТФ составляет 2 молекулы АТФ на каждую молекулу глюкозы, расщепленную в процессе брожения. Аналогичные процессы происходят во всех живых клетках; поскольку они поставляют необходимую для жизнедеятельности энергию, их иногда (не вполне корректно) называют анаэробным дыханием клеток.
У млекопитающих, в том числе у человека, такой процесс называется гликолизом и его конечным продуктом является молочная кислота, а не спирт и CO2. Вся последовательность реакций гликолиза, за исключением двух последних этапов, полностью идентична процессу, протекающему в дрожжевых клетках.
Аэробный (с использованием кислорода). С появлением в атмосфере кислорода, источником которого послужил, очевидно, фотосинтез растений, в ходе эволюции развился механизм, обеспечивающий полное окисление глюкозы до CO2 и воды, – аэробный процесс, в котором чистый выход АТФ составляет 38 молекул АТФ на каждую окисленную молекулу глюкозы. Этот процесс потребления клетками кислорода для образования богатых энергией соединений известен как клеточное дыхание (аэробное). В отличие от анаэробного процесса, осуществляемого ферментами цитоплазмы, окислительные процессы протекают в митохондриях. В митохондриях пировиноградная кислота – промежуточный продукт, образовавшийся в анаэробной фазе – окисляется до СО2 в шести последовательных реакциях, в каждой из которых пара электронов переносится на общий акцептор – кофермент никотинамидадениндинуклеотид (НАД). Эту последовательность реакций называют циклом трикарбоновых кислот, циклом лимонной кислоты или циклом Кребса. Из каждой молекулы глюкозы образуется 2 молекулы пировиноградной кислоты; 12 пар электронов отщепляется от молекулы глюкозы в ходе ее окисления, описываемого уравнением:
/>
Перенос электронов. В каждой митохондрии имеется механизм, посредством которого восстановленный НАД (НАДН, где Н – водород), образовавшийся в цикле трикарбоновых кислот, передает свою пару электронов кислороду. Перенос, однако, не происходит напрямую. Электроны как бы передаются «из рук в руки» и, лишь пройдя цепь переносчиков, присоединяются к кислороду. Эта «цепь переноса электронов» состоит из следующих компонентов:
НАДНН Флавинадениндинклеотид Кофермент Q
Цитохром b Цитохром c Цитохром a O2
Все компоненты этой системы, находящиеся в митохондриях, фиксированы в пространстве и сцеплены друг с другом. Такое их состояние облегчает перенос электронов.
В состав НАД входит никотиновая кислота (витамин ниацин), а в состав флавинадениндинуклеотида – рибофлавин (витамин B2). Кофермент Q представляет собой высокомолекулярный хинон, синтезируемый в печени, а цитохромы – это три разных белка, каждый из которых, подобно гемоглобину, содержит гемогруппу.
В цепи переноса электронов на каждую пару электронов, перенесенную от НАДН на O2, синтезируется 3 молекулы АТФ. Поскольку от каждой молекулы глюкозы отщепляются и передаются молекулам НАД 12 пар электронов, в общей сложности на каждую молекулу глюкозы образуется 312 = 36 молекул АТФ. Этот процесс образования АТФ в ходе окисления называется окислительным фосфорилированием.
Липиды как источник энергии. Жирные кислоты могут использоваться в качестве источника энергии приблизительно так же, как и углеводы. Окисление жирных кислот протекает путем последовательного отщепления от молекулы жирной кислоты двууглеродного фрагмента с образованием ацетилкофермента A (ацетил-КоА) и одновременной передачей двух пар электронов в цепь переноса электронов. Образовавшийся ацетил-КоА – нормальный компонент цикла трикарбоновых кислот, и в дальнейшем его судьба не отличается от судьбы ацетил-КоА, поставляемого углеводным обменом. Таким образом, механизмы синтеза АТФ при окислении как жирных кислот, так и метаболитов глюкозы практически одинаковы.
Если организм животного получает энергию почти целиком за счет одного только окисления жирных кислот, а это бывает, например, при голодании или при сахарном диабете, то скорость образования ацетил-КоА превышает скорость его окисления в цикле трикарбоновых кислот. В этом случае лишние молекулы ацетил-КоА реагируют друг с другом, в результате чего образуются в конечном счете ацетоуксусная и -гидроксимасляная кислоты. Их накопление является причиной патологического состояния, т.н. кетоза (одного из видов ацидоза), который при тяжелом диабете может вызвать кому и смерть.
Запасание энергии. Животные питаются нерегулярно, и их организму нужно как-то запасать заключенную в пище энергию, источником которой являются поглощенные животным углеводы и жиры. Жирные кислоты могут запасаться в виде нейтральных жиров либо в печени, либо в жировой ткани. Углеводы, поступая в большом количестве, в желудочно-кишечном тракте гидролизуются до глюкозы или иных сахаров, которые затем в печени превращаются в ту же глюкозу. Здесь из глюкозы синтезируется гигантский полимер гликоген путем присоединения друг к другу остатков глюкозы с отщеплением молекул воды (число остатков глюкозы в молекулах гликогена доходит до 30 000). Когда возникает потребность в энергии, гликоген вновь распадается до глюкозы в реакции, продуктом которой является глюкозофосфат. Этот глюкозофосфат направляется на путь гликолиза – процесса, составляющего часть пути окисления глюкозы. В печени глюкозофосфат может также подвергнуться гидролизу, и образующаяся глюкоза поступает в кровоток и доставляется кровью к клеткам в разных частях тела.
Синтез липидов из углеводов. Если количество углеводов, поглощенных с пищей за один прием, больше того, какое может быть запасено в виде гликогена, то избыток углеводов превращается в жиры. Начальная последовательность реакций совпадает при этом с обычным окислительным путем, т.е. сначала из глюкозы образуется ацетил-КоА, но далее этот ацетил-КоА используется в цитоплазме клетки для синтеза длинноцепочечных жирных кислот. Процесс синтеза можно описать как обращение обычного процесса окисления жирных клеток. Затем жирные кислоты запасаются в виде нейтральных жиров (триглицеридов), отлагающихся в разных частях тела. Когда требуется энергия, нейтральные жиры подвергаются гидролизу и жирные кислоты поступают в кровь. Здесь они адсорбируются молекулами плазменных белков (альбуминов и глобулинов) и затем поглощаются клетками самых разных типов. Механизмов, способных осуществлять синтез глюкозы из жирных кислот, у животных нет, но у растений такие механизмы имеются.
--PAGE_BREAK--Метаболизм липидов. Липиды попадают в организм главным образом в форме триглицеридов жирных кислот. В кишечнике под действием ферментов поджелудочной железы они подвергаются гидролизу, продукты которого всасываются клетками стенки кишечника. Здесь из них вновь синтезируются нейтральные жиры, которые через лимфатическую систему поступают в кровь и либо транспортируются в печень, либо отлагаются в жировой ткани. Выше уже указывалось, что жирные кислоты могут также синтезироваться заново из углеводных предшественников. Следует отметить, что, хотя в клетках млекопитающих может происходить включение одной двойной связи в молекулы длинноцепочечных жирных кислот (между С–9 и С–10), включать вторую и третью двойную связь эти клетки неспособны. Поскольку жирные кислоты с двумя и тремя двойными связями играют важную роль в метаболизме млекопитающих, они в сущности являются витаминами. Поэтому линолевую (C18:2) и линоленовую (C18:3) кислоты называют незаменимыми жирными кислотами. В то же время в клетках млекопитающих в линоленовую кислоту может включаться четвертая двойная связь и путем удлинения углеродной цепи может образоваться арахидоновая кислота (C20:4), также необходимый участник метаболических процессов.
В процессе синтеза липидов остатки жирных кислот, связанные с коферментом А (ацил-КоА), переносятся на глицерофосфат – эфир фосфорной кислоты и глицерина. В результате образуется фосфатидная кислота – соединение, в котором одна гидроксильная группа глицерина этерифицирована фосфорной кислотой, а две группы – жирными кислотами. При образовании нейтральных жиров фосфорная кислота удаляется путем гидролиза, и ее место занимает третья жирная кислота в результате реакции с ацил-КоА. Кофермент А образуется из пантотеновой кислоты (одного из витаминов). В его молекуле имеется сульфгидрильная (– SH) группа, способная реагировать с кислотами с образованием тиоэфиров. При образовании фосфолипидов фосфатидная кислота реагирует непосредственно с активированным производным одного из азотистых оснований, таких, как холин, этаноламин или серин.
За исключением витамина D, все встречающиеся в организме животных стероиды (производные сложных спиртов) легко синтезируются самим организмом. Сюда относятся холестерин (холестерол), желчные кислоты, мужские и женские половые гормоны и гормоны надпочечников. В каждом случае исходным материалом для синтеза служит ацетил-КоА: из ацетильных групп путем многократно повторяющейся конденсации строится углеродный скелет синтезируемого соединения.
Метаболизм белков
Синтез аминокислот. Растения и большинство микроорганизмов могут жить и расти в среде, в которой для их питания имеются только минеральные вещества, диоксид углерода и вода. Это значит, что все обнаруживаемые в них органические вещества эти организмы синтезируют сами. Встречающиеся во всех живых клетках белки построены из 21 вида аминокислот, соединенных в различной последовательности. Аминокислоты синтезируются живыми организмами. В каждом случае ряд химических реакций приводит к образованию -кетокислоты. Одна такая -кетокислота, а именно -кетоглутаровая (обычный компонент цикла трикарбоновых кислот), участвует в связывании азота по следующему уравнению:
-Кетоглутаровая кислота + Nh4 + НАДН
Глутаминовая кислота + НАД.
Азот глутаминовой кислоты может быть затем передан любой из других -кетокислот с образованием соответствующей аминокислоты.
Организм человека и большинства других животных сохранил способность синтезировать все аминокислоты за исключением девяти т.н. незаменимых аминокислот. Поскольку кетокислоты, соответствующие этим девяти, не синтезируются, незаменимые аминокислоты должны поступать с пищей.
Синтез белков. Аминокислоты нужны для биосинтеза белка. Процесс биосинтеза протекает обычно следующим образом. В цитоплазме клетки каждая аминокислота «активируется» в реакции с АТФ, а затем присоединяется к концевой группе молекулы рибонуклеиновой кислоты, специфичной именно для данной аминокислоты. Эта сложная молекула связывается с небольшим тельцем, т.н. рибосомой, в положении, определяемом более длинной молекулой рибонуклеиновой кислоты, прикрепленной к рибосоме. После того как все эти сложные молекулы соответствующим образом выстроились, связи между исходной аминокислотой и рибонуклеиновой кислотой разрываются и возникают связи между соседними аминокислотами – синтезируется специфичный белок. Процесс биосинтеза поставляет белки не только для роста организма или для секреции в среду. Все белки живых клеток со временем претерпевают распад до составляющих их аминокислот, и для поддержания жизни клетки должны синтезироваться вновь.
Синтез других азотсодержащих соединений. В организме млекопитающих аминокислоты используются не только для биосинтеза белков, но и как исходный материал для синтеза многих азотсодержащих соединений. Аминокислота тирозин является предшественником гормонов адреналина и норадреналина. Простейшая аминокислота глицин служит исходным материалом для биосинтеза пуринов, входящих в состав нуклеиновых кислот, и порфиринов, входящих в состав цитохромов и гемоглобина. Аспарагиновая кислота – предшественник пиримидинов нуклеиновых кислот. Метильная группа метионина передается ряду других соединений в ходе биосинтеза креатина, холина и саркозина. При биосинтезе креатина от одного соединения к другому передается также и гуанидиновая группировка аргинина. Триптофан служит предшественником никотиновой кислоты, а из валина в растениях синтезируется такой витамин, как пантотеновая кислота. Все это лишь отдельные примеры использования аминокислот в процессах биосинтеза.
Азот, поглощаемый микроорганизмами и высшими растениями в виде иона аммония, расходуется почти целиком на образование аминокислот, из которых затем синтезируются многие азотсодержащие соединения живых клеток. Избыточных количеств азота ни растения, ни микроорганизмы не поглощают. В отличие от них, у животных количество поглощенного азота зависит от содержащихся в пище белков. Весь азот, поступивший в организм в виде аминокислот и не израсходованный в процессах биосинтеза, довольно быстро выводится из организма с мочой. Происходит это следующим образом. В печени неиспользованные аминокислоты передают свой азот -кетоглутаровой кислоте с образованием глутаминовой кислоты, которая дезаминируется, высвобождая аммиак. Далее азот аммиака может либо на время запасаться путем синтеза глутамина, либо сразу же использоваться для синтеза мочевины, протекающего в печени.
У глутамина есть и другая роль. Он может подвергаться гидролизу в почках с высвобождением аммиака, который поступает в мочу в обмен на ионы натрия. Этот процесс крайне важен как средство поддержания кислотно-щелочного равновесия в организме животного. Почти весь аммиак, происходящий из аминокислот и, возможно, из других источников, превращается в печени в мочевину, так что свободного аммиака в крови обычно почти нет. Однако при некоторых условиях довольно значительные количества аммиака содержит моча. Этот аммиак образуется в почках из глутамина и переходит в мочу в обмен на ионы натрия, которые таким образом реадсорбируются и задерживаются в организме. Этот процесс усиливается при развитии ацидоза – состояния, при котором организм нуждается в дополнительных количествах катионов натрия для связывания избытка ионов бикарбоната в крови.
Избыточные количества пиримидинов тоже распадаются в печени через ряд реакций, в которых высвобождается аммиак. Что касается пуринов, то их избыток подвергается окислению с образованием мочевой кислоты, выделяющейся с мочой у человека и других приматов, но не у остальных млекопитающих. У птиц отсутствует механизм синтеза мочевины, и именно мочевая кислота, а не мочевина, является у них конечным продуктом обмена всех азотсодержащих соединений.
Общие представления о метаболизме органических веществ
Можно сформулировать некоторые общие понятия, или «правила», касающиеся метаболизма. Приведенные ниже несколько главных «правил» позволяют лучше понять, как протекает и регулируется метаболизм.
1. Метаболические пути необратимы. Распад никогда не идет по пути, который являлся бы простым обращением реакций синтеза. В нем участвуют другие ферменты и другие промежуточные продукты. Нередко противоположно направленные процессы протекают в разных отсеках клетки. Так, жирные кислоты синтезируются в цитоплазме при участии одного набора ферментов, а окисляются в митохондриях при участии совсем другого набора.
2. Ферментов в живых клетках достаточно для того, чтобы все известные метаболические реакции могли протекать гораздо быстрее, чем это обычно наблюдается в организме. Следовательно, в клетках существуют какие-то регуляторные механизмы. Открыты разные типы таких механизмов.
а) Фактором, ограничивающим скорость метаболических превращений данного вещества, может быть поступление этого вещества в клетку; именно на этот процесс в таком случае и направлена регуляция. Роль инсулина, например, связана с тем, что он, по-видимому, облегчает проникновение глюкозы во все клетки, глюкоза же подвергается превращениям с той скоростью, с какой она поступает. Сходным образом проникновение железа и кальция из кишечника в кровь зависит от процессов, скорость которых регулируется.
б) Вещества далеко не всегда могут свободно переходить из одного клеточного отсека в другой; есть данные, что внутриклеточный перенос регулируется некоторыми стероидными гормонами.
в) Выявлено два типа сервомеханизмов «отрицательной обратной связи».
У бактерий были обнаружены примеры того, что присутствие продукта какой-нибудь последовательности реакций, например аминокислоты, подавляет биосинтез одного из ферментов, необходимых для образования этой аминокислоты.
/>
В каждом случае фермент, биосинтез которого оказывается затронутым, был ответствен за первый «определяющий» этап (на схеме реакция 4) метаболического пути, ведущего к синтезу данной аминокислоты.
Второй механизм хорошо изучен у млекопитающих. Это простое ингибирование конечным продуктом (в нашем случае – аминокислотой) фермента, ответственного за первый «определяющий» этап метаболического пути.
Еще один тип регулирования посредством обратной связи действует в тех случаях, когда окисление промежуточных продуктов цикла трикарбоновых кислот сопряжено с образованием АТФ из АДФ и фосфата в процессе окислительного фосфорилирования. Если весь имеющийся в клетке запас фосфата и (или) АДФ уже исчерпан, то окисление приостанавливается и может возобновиться лишь после того, как этот запас вновь станет достаточным. Таким образом, окисление, смысл которого в том, чтобы поставлять полезную энергию в форме АТФ, происходит только тогда, когда возможен синтез АТФ.
3. В биосинтетических процессах участвует сравнительно небольшое число строительных блоков, каждый из которых используется для синтеза многих соединений. Среди них можно назвать ацетилкофермент А, глицерофосфат, глицин, карбамилфосфат, поставляющий карбамильную (h3N–CO–) группу, производные фолиевой кислоты, служащие источником гидроксиметильной и формильной групп, S-аденозилметионин – источник метильных групп, глутаминовую и аспарагиновую кислоты, поставляющие аминогруппы, и наконец, глутамин – источник амидных групп. Из этого относительно небольшого числа компонентов строятся все те разнообразные соединения, которые мы находим в живых организмах.
4. Простые органические соединения редко участвуют в метаболических реакциях непосредственно. Обычно они должны быть сначала «активированы» путем присоединения к одному из ряда соединений, универсально используемых в метаболизме. Глюкоза, например, может подвергнуться окислению лишь после того, как она будет этерифицирована фосфорной кислотой, для прочих же своих превращений она должна быть этерифицирована уридиндифосфатом. Жирные кислоты не могут быть вовлечены в метаболические превращения прежде, чем они образуют эфиры с коферментом А. Каждый из этих активаторов либо родствен одному из нуклеотидов, входящих в состав рибонуклеиновой кислоты, либо образуется из какого-нибудь витамина. Легко понять в связи с этим, почему витамины требуются в таких небольших количествах. Они расходуются на образование «коферментов», а каждая молекула кофермента на протяжении жизни организма используется многократно, в отличие от основных питательных веществ (например, глюкозы), каждая молекула которых используется только один раз.
В заключение следует сказать, что термин «метаболизм», означавший ранее нечто не более сложное, чем просто использование углеводов и жиров в организме, теперь применяется для обозначения тысяч ферментативных реакций, вся совокупность которых может быть представлена как огромная сеть метаболических путей, многократно пересекающихся (из-за наличия общих промежуточных продуктов) и управляемых очень тонкими регуляторными механизмами.
Метаболизм минеральных веществ
Относительное содержание. Различные элементы, встречающиеся в живых организмах, перечислены ниже в убывающем порядке в зависимости от их относительного содержания: 1) кислород, углерод, водород и азот; 2) кальций, фосфор, калий и сера; 3) натрий, хлор, магний и железо; 4) марганец, медь, молибден, селен, йод и цинк; 5) алюминий, фтор, кремний и литий; 6) бром, мышьяк, свинец и, возможно, некоторые другие.
Кислород, углерод, водород и азот – это те элементы, из которых построены мягкие ткани тела. Они входят в состав таких соединений, как углеводы, липиды, белки, вода, диоксид углерода и аммиак. Элементы, перечисленные в пп. 2 и 3, находятся в организме обычно в виде одного или нескольких неорганических соединений, а элементы пп. 4, 5 и 6 присутствуют только в следовых количествах и потому их называют микроэлементами.
Распределение в организме. Кальций. Кальций присутствует главным образом в костной ткани и в зубах, преимущественно в виде фосфата и в небольших количествах в виде карбоната и фторида. Поступающий с пищей кальций всасывается в основном в верхних отделах кишечника, имеющих слабокислую реакцию. Этому всасыванию (у человека здесь всасывается всего 20–30% кальция пищи) способствует витамин D. Под действием витамина D клетки кишечника вырабатывают особый белок, который связывает кальций и облегчает его перенос через стенку кишечника в кровь. На всасывание влияет также присутствие некоторых других веществ, в особенности фосфата и оксалата, которые в малых количествах способствуют всасыванию, а в больших, наоборот, подавляют его.
В крови около половины кальция связано с белком, остальное составляют ионы кальция. Соотношение ионизированной и неионизированной форм зависит от общей концентрации кальция в крови, а также от содержания белка и фосфата и концентрации водородных ионов (рН крови). Доля неионизированного кальция, на которую влияет уровень белка, позволяет косвенным образом судить о качестве питания и об эффективности работы печени, в которой идет синтез плазменных белков.
На количество ионизированного кальция влияют, с одной стороны, витамин D и факторы, воздействующие на всасывание, а с другой – паратиреоидный гормон и, возможно, также витамин D, поскольку оба эти вещества регулируют как скорость отложения кальция в костной ткани, так и его мобилизацию, т.е. вымывание из костей. Избыток паратиреоидного гормона стимулирует выход кальция из костной ткани, что приводит к повышению его концентрации в плазме. Изменяя скорости всасывания и экскреции кальция и фосфата, а также скорости образования костной ткани и ее разрушения, эти механизмы строго контролируют концентрацию кальция и фосфата в сыворотке крови. Ионы кальция играют регулирующую роль во многих физиологических процессах, в том числе в нервных реакциях, мышечном сокращении, свертывании крови. Выведение кальция из организма происходит в норме в основном (на 2/3) через желчь и кишечник и в меньшей степени (1/3) – через почки.
Фосфор. Метаболизм фосфора – одного из главных компонентов костной ткани и зубов – во многом зависит от тех же факторов, что и метаболизм кальция. Фосфор в виде фосфата присутствует в организме также в сотнях различных физиологически важных органических эфиров. Паратиреоидный гормон стимулирует выведение фосфора с мочой и выход его из костной ткани; тем самым он регулирует концентрацию фосфора в плазме крови.
Натрий. Натрий – главный катион внеклеточной жидкости – вместе с белком, хлоридом и бикарбонатом играет важнейшую роль в регулировании осмотического давления и pH (концентрации водородных ионов) крови. В клетках, напротив, содержится очень мало натрия, так как они обладают механизмом для выведения ионов натрия и удержания ионов калия. Весь натрий, превышающий потребности организма, очень быстро выводится через почки.
Поскольку во всех процессах выделения натрий теряется, он должен постоянно поступать в организм с пищей. При ацидозе, когда необходимо, чтобы из организма выводились большие количества анионов (например, хлорида или ацетоацетата), почки предотвращают чрезмерную потерю натрия благодаря образованию аммиака из глутамина. Выведение натрия через почки регулируется гормоном коры надпочечников альдостероном. Под действием этого гормона в кровь возвращается достаточно натрия для поддержания нормального осмотического давления и нормального объема внеклеточной жидкости.
Суточная потребность в хлористом натрии составляет 5–10 г. Эта величина возрастает при поглощении больших количеств жидкости, когда усиливается потоотделение и выделяется больше мочи.
Калий. В отличие от натрия, калий содержится в клетках в больших количествах, но во внеклеточной жидкости его мало. Главная функция калия – регулирование внутриклеточного осмотического давления и поддержание кислотно-щелочного равновесия. Он также играет важную роль в проведении нервного импульса и во многих ферментных системах, в том числе и в тех, которые участвуют в мышечном сокращении. Калий широко распространен в природе, и его много в любой пище, так что спонтанно калиевая недостаточность возникнуть не может. В плазме концентрация калия регулируется альдостероном, стимулирующим его экскрецию с мочой.
Сера. С пищей сера поступает в организм главным образом в составе двух аминокислот – цистина и метионина. На конечных этапах метаболизма этих аминокислот сера высвобождается и в результате окисления переводится в неорганическую форму. В составе цистина и метионина сера присутствует в структурных белках. Важную роль играет также сульфгидрильная (–SH) группа цистеина, от которой зависит активность многих ферментов.
Большая часть серы выводится с мочой в виде сульфата. Небольшое количество экскретируемого сульфата обычно связано с органическими соединениями типа фенолов.
Магний. Метаболизм магния сходен с метаболизмом кальция, и в виде комплекса с фосфатом этот элемент тоже входит в состав костной ткани. Магний присутствует во всех живых клетках, где он функционирует как необходимый компонент многих ферментных систем; эта его роль была убедительно продемонстрирована на примере углеводного обмена в мышцах. Магний, как и калий, широко распространен, и вероятность возникновения его недостаточности очень мала.
Железо. Железо входит в состав гемоглобина и других гемопротеинов, а именно миоглобина (мышечного гемоглобина), цитохромов (дыхательных ферментов) и каталазы, а также в состав некоторых ферментов, не содержащих гемогруппы. Всасывается железо в верхних отделах кишечника, причем это единственный элемент, всасывающийся только тогда, когда его запас в организме полностью исчерпан. В плазме железо транспортируется в соединении с белком (трансферрином). Через почки железо не выводится; избыток его накапливается в печени в соединении с особым белком (ферритином).
Микроэлементы. У каждого микроэлемента, присутствующего в организме, своя особая функция, связанная с тем, что он стимулирует действие того или иного фермента или как-либо иначе на него влияет. Цинк необходим для кристаллизации инсулина; кроме того, он является компонентом карбоангидразы (фермента, участвующего в транспорте диоксида углерода) и некоторых других ферментов. Молибден и медь – тоже необходимые компоненты различных ферментов. Иод требуется для синтеза трииодтиронина, гормона щитовидной железы. Фтор (входящий в состав зубной эмали) способствует предотвращению кариеса.
Использование метаболитов
Углеводы. Всасывание. Моносахариды, или простые сахара, высвобождающиеся при переваривании углеводов пищи, переходят из кишечника в кровоток в результате процесса, называемого всасыванием. Механизм всасывания представляет собой сочетание простой диффузии и химической реакции (активного всасывания). Одна из гипотез, касающихся природы химической фазы процесса, предполагает, что в этой фазе моносахариды соединяются с фосфорной кислотой в реакции, катализируемой ферментом из группы киназ, после чего проникают в кровеносные сосуды и здесь высвобождаются в результате ферментативного дефосфорилирования (разрыва фосфатной связи), катализируемого одной из фосфатаз. Именно активным всасыванием объясняется то, что разные моносахариды всасываются с разной скоростью и что углеводы всасываются даже тогда, когда уровень сахара в крови выше, чем в кишечнике, т.е. в условиях, когда естественно было бы ожидать их перемещения в обратном направлении – из крови в кишечник.
Механизмы гомеостаза. Поступающие в кровоток моносахариды повышают уровень сахара в крови. При голодании концентрация глюкозы в крови колеблется обычно от 70 до 100 мг на 100 мл крови. Этот уровень поддерживается с помощью механизмов, называемых механизмами гомеостаза (самостабилизации). Как только уровень сахара в крови в результате всасывания из кишечника повышается, в действие вступают процессы, выводящие сахар из крови, так что уровень его колеблется не слишком сильно.
Подобно глюкозе, все прочие моносахариды поступают из кровотока в печень, где превращаются в глюкозу. Теперь они неотличимы как от глюкозы, которая всосалась, так и от той, что уже была в организме, и подвергаются тем же метаболическим превращениям. Один из механизмов гомеостаза углеводов, функционирующий в печени, – это гликогенез, посредством которого глюкоза переходит из крови в клетки, где превращается в гликоген. Гликоген хранится в печени до тех пор, пока не произойдет снижение уровня сахара в крови: в этой ситуации гомеостатический механизм вызовет распад накопленного гликогена до глюкозы, которая вновь поступит в кровь.
Превращения и использование. Поскольку кровь поставляет глюкозу во все ткани тела и все ткани используют ее для получения энергии, уровень глюкозы в крови снижается главным образом за счет ее использования.
В мышцах глюкоза крови превращается в гликоген. Однако мышечный гликоген не может быть использован для получения глюкозы, которая перешла бы в кровь. В нем заключен запас энергии, и скорость его использования зависит от мышечной активности. В мышечной ткани содержатся два соединения с большим запасом легко доступной энергии в форме богатых энергией фосфатных связей – креатинфосфат и аденозинтрифосфат (АТФ). При отщеплении от этих соединений их фосфатных групп высвобождается энергия для мышечного сокращения. Чтобы мышца вновь могла сокращаться, эти соединения должны быть восстановлены в своей исходной форме. Для этого требуется энергия, которую поставляет окисление продуктов распада гликогена. При мышечном сокращении гликоген превращается в глюкозофосфат, а затем – через ряд реакций – во фруктозодифосфат. Фруктозодифосфат распадается на два трехуглеродных соединения, из которых после ряда этапов образуется сначала пировиноградная кислота, а в конечном итоге – молочная кислота, как об этом уже говорилось при описании метаболизма углеводов. Это превращение гликогена в молочную кислоту, сопровождающееся высвобождением энергии, может происходить в отсутствие кислорода.
При недостатке кислорода молочная кислота накапливается в мышцах, диффундирует в кровоток и поступает в печень, где из нее вновь образуется гликоген. Если кислорода достаточно, то молочная кислота в мышцах не накапливается. Вместо этого она, как это описано выше, полностью окисляется через цикл трикарбоновых кислот до диоксида углерода и воды с образованием АТФ, который может быть использован для сокращения.
Метаболизм углеводов в нервной ткани и эритроцитах отличается от метаболизма в мышцах тем, что гликоген здесь не участвует. Однако и здесь промежуточными продуктами являются пировиноградная и молочная кислоты, образующиеся при расщеплении глюкозофосфата.
Глюкоза используется не только в клеточном дыхании, но и во многих других процессах: синтезе лактозы (молочного сахара), образовании жиров, а также особых сахаров, входящих в состав полисахаридов соединительной ткани и ряда других тканей.
Гликоген печени, синтезируемый при всасывании углеводов в кишечнике, служит самым доступным источником глюкозы, когда всасывание отсутствует. Если этот источник оказывается исчерпанным, в печени начинается процесс глюконеогенеза. Глюкоза образуется при этом из некоторых аминокислот (из 100 г белка образуется 58 г глюкозы) и нескольких других неуглеводных соединений, в том числе из глицериновых остатков нейтральных жиров.
Некоторую, хотя и не столь важную, роль в метаболизме углеводов играют почки. Они выводят из организма избыток глюкозы, когда ее концентрация в крови слишком высока; при меньших концентрациях глюкоза практически не выводится.
В регулировании метаболизма углеводов участвует несколько гормонов, в том числе гормоны поджелудочной железы, передней доли гипофиза и коры надпочечников.
Гормон поджелудочной железы инсулин снижает концентрацию глюкозы в крови и повышает ее концентрацию в клетках. По-видимому, он стимулирует также и запасание гликогена в печени. Кортикостерон, гормон коры надпочечников, и адреналин, вырабатываемый мозговым веществом надпочечников, воздействуют на метаболизм углеводов, стимулируя распад гликогена (главным образом в мышцах и печени) и синтез глюкозы (в печени).
Липиды. Всасывание. В кишечнике после переваривания жиров остаются главным образом свободные жирные кислоты с небольшой примесью холестерина и лецитина и следами жирорастворимых витаминов. Все эти вещества очень тонко диспергированы благодаря эмульгирующему и солюбилизирующему действию солей желчных кислот. Солюбилизирующее действие обычно связывают с образованием нестойких химических соединений между жирными кислотами и солями желчных кислот. Эти комплексы проникают в клетки эпителия тонкого кишечника и здесь распадаются на жирные кислоты и соли желчных кислот. Последние переносятся в печень и вновь секретируются с желчью, а жирные кислоты вступают в соединение с глицерином или холестерином. Образовавшиеся реконструированные жиры поступают в лимфатические сосуды брыжейки в форме млечного сока, т.н. «хилуса». Из сосудов брыжейки хилус по лимфатической системе через грудной проток поступает в кровеносную систему.
После переваривания пищи содержание липидов в крови возрастает приблизительно от 500 мг (уровень при голодании) до 1000 мг на 100 мл плазмы. Присутствующие в крови липиды представляют собой смесь жирных кислот, нейтральных жиров, фосфолипидов (лецитина и кефалина), холестерина и эфиров холестерина.
Распределение. Кровь доставляет липиды в разные ткани тела и прежде всего в печень. Печень обладает способностью модифицировать поступающие в нее жирные кислоты. Это особенно выражено у видов, запасающих жиры с высоким содержанием насыщенных или, наоборот, ненасыщенных жирных кислот: в печени этих животных соотношение насыщенных и ненасыщенных кислот изменяется таким образом, что отлагающийся жир по своему составу соответствует жиру, свойственному данному организму.
Жиры в печени либо используются для получения энергии, либо переходят в кровь и доставляются ею в разные ткани. Здесь они могут включаться в структурные элементы тканей, но большая их часть отлагается в жировых депо, где они хранятся до тех пор, пока не возникнет потребность в энергии; тогда они снова переносятся в печень и подвергаются здесь окислению.
Метаболизм липидов, как и углеводов, регулируется гомеостатически. Механизмы гомеостаза, воздействующие на липидный и углеводный обмен, видимо, тесно связаны, поскольку при замедлении метаболизма углеводов усиливается метаболизм липидов, и наоборот.
Превращения и использование. Четырехуглеродные кислоты – ацетоуксусная (продукт конденсации двух ацетатных единиц) и -гидроксимасляная – и трехуглеродное соединение ацетон, образующийся при отщеплении одного атома углерода от ацетоуксусной кислоты, известны под общим названием кетоновых (ацетоновых) тел. В норме кетоновые тела присутствуют в крови в небольших количествах. Избыточное их образование при тяжелом диабете ведет к повышению их содержания в крови (кетонемия) и в моче (кетонурия) – это состояние обозначают термином «кетоз».
Белки. Всасывание. При переваривании белков пищеварительными ферментами образуется смесь из аминокислот и небольших пептидов, содержащих от двух до десяти остатков аминокислот. Эти продукты всасываются слизистой кишечника, и здесь гидролиз завершается – пептиды также распадаются до аминокислот. Поступившие в кровь аминокислоты смешиваются с находящимися здесь такими же аминокислотами. В крови содержится смесь из аминокислот, поступивших из кишечника, образовавшихся при распаде тканевых белков и синтезированных организмом заново.
Синтез. В тканях непрерывно идет распад белков и их новообразование. Содержащиеся в крови аминокислоты избирательно поглощаются тканями как исходный материал для построения белков, а из тканей в кровь поступают другие аминокислоты. Синтезу и распаду подвергаются не только структурные белки, но и белки плазмы крови, а также белковые гормоны и ферменты.
Во взрослом организме аминокислоты или белки практически не запасаются, поэтому удаление аминокислот из крови происходит с такой же скоростью, как и их поступление из тканей в кровь. В растущем организме формируются новые ткани, и на этот процесс расходуется больше аминокислот, чем поступает в кровь за счет распада тканевых белков.
Печень участвует в метаболизме белков самым активным образом. Здесь синтезируются белки плазмы крови – альбумины и глобулины – а также собственные ферменты печени. Так, при потере плазменных белков содержание альбумина в плазме восстанавливается – за счет интенсивного синтеза – довольно быстро. Аминокислоты в печени используются не только для образования белков, но подвергаются также расщеплению, в ходе которого извлекается заключенная в них энергия.
Превращения и использование. Если аминокислоты используются в качестве источника энергии, то отщепляемая от них аминогруппа (–Nh3) направляется на образование мочевины, а не содержащий азота остаток молекулы окисляется приблизительно так же, как глюкоза или жирные кислоты.
Так называемый «орнитиновый цикл» описывает, как происходит превращение аммиака в мочевину. В этом цикле аминогруппа, отщепившаяся от аминокислоты в форме аммиака, присоединяется вместе с диоксидом углерода к молекуле орнитина с образованием цитруллина. Цитруллин присоединяет второй атом азота, на этот раз от аспарагиновой кислоты, и превращается в аргинин. Далее аргинин подвергается гидролизу с образованием мочевины и орнитина. Орнитин может теперь вновь вступить в цикл, а мочевина выводится из организма через почки как один из конечных продуктов метаболизма.
Список литературыЛенинджер А. Основы биохимии, тт. 1–3. М., 1985
Страйер Л. Биохимия, тт. 1–3. М., 1985
Марри Р., Греннер Д., Мейес П., Родуэлл В. Биохимия человека, тт. 1–2. М., 1993
Албертс Б., Брей Д., Льюс Д. и др. Молекулярная биология клетки, тт. 1–3. М., 1994
www.ronl.ru
Реферат Метаболизм
Реферат на тему:
План:
- Введение
- 1 Биологические молекулы
- 1.1 Аминокислоты и белки
- 1.2 Липиды
- 1.3 Углеводы
- 1.4 Нуклеотиды
- 1.5 Коферменты
- 1.6 Минералы и кофакторы
- 2 Катаболизм
- 2.1 Пищеварение
- 2.2 Получение энергии
- 3 Энергетические превращения
- 3.1 Окислительное фосфорилирование
- 3.2 Энергия из неорганических соединений
- 3.3 Энергия из солнечного света
- 4 Анаболизм
- 4.1 Связывание углерода
- 4.2 Углеводы и гликаны
- 4.3 Жирные кислоты, изопреноиды и стероиды
- 4.4 Белки
- 4.5 Нуклеотиды
- 5 Ксенобиотики и окислительный метаболизм
- 6 Термодинамика живых организмов
- 7 Регуляция и контроль
- 8 Эволюция
- 9 Методы исследования
- 10 История Примечания
Введение
Структура аденозинтрифосфата — главного посредника в энергетическом обмене веществ
Метаболи́зм (от греч. μεταβολή — «превращение, изменение»), или обмен веществ — набор химических реакций, которые возникают в живом организме для поддержания жизни. Эти процессы позволяют организмам расти и размножаться, сохранять свои структуры и отвечать на воздействия окружающей среды. Метаболизм обычно делят на две стадии: в ходе катаболизма сложные органические вещества деградируют до более простых; в процессах анаболизма с затратами энергии синтезируются такие вещества, как белки, сахара, липиды и нуклеиновые кислоты.
Серии химических реакций обмена веществ называют метаболическими путями, в них при участии ферментов одни биологически значимые молекулы последовательно превращаются в другие. Ферменты играют важную роль в метаболических процессах потому, что:
- действуют как биологические катализаторы и снижают энергию активации химической реакции;
- позволяют регулировать метаболические пути в ответ на изменения среды клетки или сигналы от других клеток.
Особенности метаболизма влияют на то, будет ли пригодна определенная молекула для использования организмом в качестве источника энергии. Так, например, некоторые прокариоты используют сероводород в качестве источника энергии, однако этот газ ядовит для животных.[1] Скорость обмена веществ также влияет на количество пищи, необходимой для организма.
Основные метаболические пути и их компоненты одинаковы для многих видов, что свидетельствует о единстве происхождения всех живых существ.[2] Например, некоторые карбоновые кислоты, являющиеся интермедиатами цикла трикарбоновых кислот присутствуют во всех организмах, начиная от бактерий и заканчивая многоклеточными организмами эукариот.[3] Сходства в обмене веществ, вероятно, связаны с высокой эффективностью метаболических путей, а также с их ранним появлением в истории эволюции.[4][5]
1. Биологические молекулы
Структура липида триглицерида
Органические вещества, входящие в состав всех живых существ (животных, растений, грибов и микроорганизмов), представлены в основном аминокислотами, углеводами, липидами (часто называемые жирами) и нуклеиновыми кислотами. Так как эти молекулы имеют важное значение для жизни, метаболические реакции сосредоточены на создании этих молекул при строительстве клеток и тканей или разрушении их с целью использования в качестве источника энергии. Многие важные биохимические реакции объединяются вместе для синтеза ДНК и белков.
| Аминокислоты | Аминокислоты | Белки (полипептиды) | Фибриллярные и глобулярные белки |
| Углеводы | Моносахариды | Полисахариды | Крахмал, гликоген, целлюлоза |
| Нуклеиновые кислоты | Нуклеотиды | Полинуклеотиды | ДНК и РНК |
1.1. Аминокислоты и белки
Белки являются линейными биополимерами и состоят из остатков аминокислот, соединённых пептидными связями. Некоторые белки являются ферментами и катализируют химические реакции. Другие белки выполняют структурную или механическую функцию (например, образуют цитоскелет).[6] Белки также играют важную роль в передаче сигнала в клетках, иммунных реакциях, агрегации клеток, активном транспорте через мембраны и регуляции клеточного цикла.[7]
1.2. Липиды
Липиды входят в состав биологических мембран, например, плазматических мембран, являются компонентами коферментов и источниками энергии.[7] Липиды являются гидрофобными или амфифильными биологическими молекулами, растворимыми в органических растворителях таких, как бензол или хлороформ.[8] Жиры — большая группа соединений, в состав которых входят жирные кислоты и глицерин. Молекула трёхатомного спирта глицерина, образующая три сложные эфирные связи с тремя молекулами жирных кислот, называется триглицеридом.[9] Наряду с остатками жирных кислот, в состав сложных липидов может входить, например, сфингозин (сфинголипиды), гидрофильные группы фосфатов (в фосфолипидах). Стероиды, например холестерол, представляют собой ещё один большой класс липидов.[10]
1.3. Углеводы
Сахара могут существовать в кольцевой или линейной форме в виде альдегидов или кетонов, имеют несколько гидроксильных групп. Углеводы являются наиболее распространёнными биологическими молекулами. Углеводы выполняют следующие функции, например, хранение и транспортировка энергии (крахмал, гликоген), структурная (целлюлоза растений, хитин у животных).[7] Наиболее распространенными мономерами сахаров являются гексозы — глюкоза, фруктоза и галактоза. Моносахариды входят в состав более сложных линейных или разветвленных полисахаридов.[11]
1.4. Нуклеотиды
Полимерные молекулы ДНК и РНК представляют собой длинные неразветвленные цепочки нуклеотидов. Нуклеиновые кислоты выполняют функцию хранения и реализации генетической информации, которые осуществляются в ходе процессов репликации,транскрипции, трансляции, и биосинтеза белка.[7] Информация, закодированная в нуклеиновых кислотах, защищается от изменений системами репарации и мультиплицируется при помощи репликации ДНК.
Некоторые вирусы имеют РНК-содержащий геном. Например, вирус иммунодефицита человека использует обратную транскрипцию для создания матрицы ДНК из собственного РНК-содержащего генома.[12] Некоторые молекулы РНК обладают каталитическими свойствами (рибозимы) и входят в состав сплайсосом и рибосом.
Нуклеозиды — продукты присоединения азотистых оснований к сахару рибозе. Примерами азотистых оснований являются гетероциклические азотсодержащие соединения — производные пуринов и пиримидинов. Некоторые нуклеотиды также выступают в качестве коферментов в реакциях переноса функциональных групп.[13]
1.5. Коферменты
Структура кофермента Ацетил-КоА. Ацетильная группа связана с атомом серы по левому краю.
Подробное рассмотрение темы: КоферментыМетаболизм включает широкий спектр химических реакций, большинство из которых относятся к нескольким основным типам реакций переноса функциональных групп.[14] Для переноса функциональных групп между ферментами, катализирующими химические реакции, используются коферменты.[13] Каждый класс химических реакций переноса функциональных групп катализируется отдельными ферментами и их кофакторами.[15]
Аденозинтрифосфат (АТР) — один из центральных коферментов, универсальный источник энергии клеток. Этот нуклеотид используется для передачи химической энергии, запасенной в макроэргических связях между различными химическими реакциями. В клетках существует небольшое количество АТР, который постоянно регенерируется из ADP и AMP. Организм человека за сутки расходует массу АТР, равную массе собственного тела.[15] АТР выступает в качестве связующего звена между катаболизмом и анаболизмом: при катаболических реакциях образуется АТР, при анаболических — энергия потребляется. АТР также выступает донором фосфатной группы в реакциях фосфорилирования.
Витамины — низкомолекулярные органические вещества, необходимые в небольших количествах, причём, например, у человека большинство витаминов не синтезируется, а получается с пищей или через микрофлору КТ. В организме человека большинство витаминов являются кофакторами ферментов. Большинство витаминов приобретают биологическую активность в измененном виде, например, все водорастворимые витамины в клетках фосфорилируются или соединяются с нуклеотидами.[16] Никотинамидадениндинуклеотид (NADH) является производным витамина B3 (ниацина), и представляет собой важный кофермент — акцептора водорода. Сотни различных ферментов дегидрогеназ отнимают электроны из молекул субстратов и переносят их на молекулы NAD+, восстанавливая его до NADH. Окисленная форма кофермента выступает является субстратом для различных редуктаз в клетке.[17] NAD в клетке существует в двух связанных формах NADH и NADPH. NAD+/NADH больше важен для протекания катаболических реакций, а NADP+/NADPH чаще используется в анаболических реакциях.
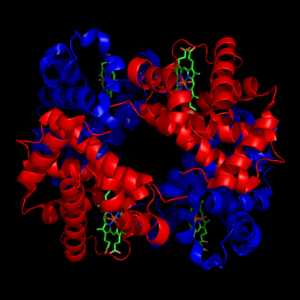
Структура гемоглобина. Белковые субъединицы окрашены красным и синим, а железосодержащий гем — зелёным. Из PDB 1GZX.
1.6. Минералы и кофакторы
Неорганические элементы играют важнейшую роль в обмене веществ. Около 99 % массы млекопитающего состоит из углерода, азота, кальция, натрия, магния, хлора, калия, водорода, фосфора, кислорода и серы.[18] Биологически значимые органические соединения (белки, жиры, углеводы и нуклеиновые кислоты) содержат большое количество углерода, водорода, кислорода, азота и фосфора.[18]
Многие неорганические соединения являются ионными электролитами. Наиболее важны для организма ионы натрия, калия, кальция, магния, хлоридов, фосфатов гидрокарбонатов. Баланс этих ионов внутри клетки во внеклеточной среде определяет осмотическое давление и рН.[19] Концентрации ионов также играют важную роль для функционирования нервных и мышечных клеток. Потенциал действия в возбудимых тканях возникает при обмене ионами между внеклеточной жидкостью и цитоплазмой.[20] Электролиты входят и выходят из клетки через ионные каналы в плазматической мембране. Например, в ходе мышечного сокращения в плазматической мембране, цитоплазме и Т-трубочках перемещаются ионы кальция, натрия и калия.[21]
Переходные металлы в организме являются микроэлементами, наиболее распространены цинк и железо.[22][23] Эти металлы используются некоторыми белками (например, ферментами в качестве кофакторов) и имеют важное значение для регуляции активности ферментов и транспортных белков.[24] Кофакторы ферментов обычно прочно связаны со специфическим белком, однако могут модифицироваться процессе катализа, при этом после окончания катализа всегда возвращаются к своему первоначальному состоянию (не расходуются). Металлы-микроэлементы усваиваются организмом при помощи специальных транспортных белков и не встречаются в организме в свободном состоянии, так как связаны со специфическими белками-переносчиками (например, ферритином или металлотионеинами).[25][26]
2. Катаболизм
Катаболизмом называют метаболические процессы, при которых расщепляются относительно крупные органические молекулы сахаров, жиров, аминокислот. В ходе катаболизма образуются более простые органические молекулы, необходимые для реакций анаболизма (биосинтеза). Часто, именно в ходе реакций катаболизма организм мобилизует энергию, переводя энергию химических связей органических молекул, полученных в процессе переваривания пищи, в доступные формы: в виде АТФ, восстановленных коферментов и трансмембранного электрохимического потенциала. Термин катаболизм не является синонимом «энергетического обмена»: у многих организмов (например, у фототрофов) основные процессы запасания энергии не связаны напрямую с расщеплением органических молекул. Классификация организмов по типу метаболизма может быть основана на источнике получения энергии и углерода, что отражено в таблице ниже. Органические молекулы используются в качестве источника энергии в органотрофами, литотрофы используют неорганические субстраты, а фототрофы потребляют энергию солнечного света. Однако, все эти различные формы обмена веществ зависят от окислительно-восстановительных реакций, которые связаны с передачей электронов от восстановленных доноров молекул, таких как органические молекулы, вода, аммиак, сероводород, на акцепторные молекулы, такие как кислород, нитраты или сульфат.[27] У животных эти реакции сопряжены с расщеплением сложных органических молекул до более простых, таких как двуокись углерода и воду. В фотосинтезирующих организмах — растениях и цианобактериях — реакции переноса электрона не высвобождают энергию, но они используются как способ запасания энергии, поглощаемой из солнечного света.[28]
| Источник энергии | Солнечный свет | фототрофы |
| Первичные молекулы | хемотрофы | |
| Донор электронов | Органическое соединение | органотрофы |
| Неорганическое соединение | литотрофы | |
| Источник углерода | Органическое соединение | гетеротрофы |
| Неорганическое соединение | автотрофы |
Катаболизм у животных может быть разделён на три основных этапа. Во-первых, крупные органические молекулы, такие как белки, полисахариды и липиды расщепляются до более мелких компонентов вне клеток. Далее эти небольшие молекулы попадают в клетки и превращается в ещё более мелкие молекулы, например, ацетил-КоА. В свою очередь, ацетильная группа кофермента А окисляется до воды и углекислого газа в цикле Кребса и дыхательной цепи, высвобождая при этом энергию, которая запасается в форме АТР.
2.1. Пищеварение
Подробное рассмотрение темы: Пищеварение и Желудочно-кишечный трактТакие макромолекулы, как крахмал, целлюлоза или белки, должны расщепляться до более мелких единиц прежде, чем они могут быть использованы клетками. Несколько классов ферментов принимают участие в деградации: протеазы, которые расщепляют белки до пептидов и аминокислот, гликозидазы, которые расщепляют полисахариды до олиго- и моносахаридов.
Микроорганизмы выделяют гидролитические ферменты в пространство вокруг себя,[29][30] чем отличаются от животных, которые выделяют такие ферменты только из специализированных железистых клеток.[31] Аминокислоты и моносахариды, образующиеся в результате активности внеклеточных ферментов, затем поступают в клетки с помощью активного транспорта.[32][33]

Упрощённая схема катаболизма белков, сахаров и липидов
2.2. Получение энергии
Подробное рассмотрение темы: Клеточное дыхание, Брожение, ЛиполизВ ходе катаболизма углеводов сложные сахара расщепляются до моносахаридов, которые усваиваются клетками.[34] Попав внутрь, сахара (например, глюкоза и фруктоза) в процессе гликолиза превращаются в пируват, при этом вырабатывается некоторое количество АТР.[35] Пировиноградная кислота (пируват) является промежуточным продуктом в нескольких метаболических путях. Основной путь метаболизма пирувата — превращаение в ацетил-КоА и далее поступление в цикл трикарбоновых кислот. При этом в цикле Кребса в форме АТР запасается часть энергии, а также восстанавливаются молекулы NADH и FAD. В процессе гликолиза и цикла трикарбоновых кислот образуется диоксид углерода, который является побочным продуктом жизнедеятельности. В анаэробных условиях в результате гликолиза из пирувата при участии фермента лактатдегидрогеназы образуется лактат, и происходит окисление NADH до NAD+, который повторно используется в реакциях гликолиза. Существует также альтернативный путь метаболизма моносахаридов — пентозофосфатный путь, в ходе реакций которого энергия запасается в форме восстановленного кофермента NADPH и образуются пентозы, например, рибоза, необходимая для синтеза нуклеиновых кислот.
Жиры на первом этапе катаболизма гидролизуются в свободные жирные кислоты и глицерин. Жирные кислоты расщепляются в процессе бета-окисления с образованием ацетил-КоА, который в свою очередь далее катаболизируется в цикле Кребса, либо идет на синтез новых жирных кислот. Жирные кислоты выделяют больше энергии, чем углеводы, так как жиры содержат удельно больше атомов водорода в своей структуре.
Аминокислоты либо используются для синтеза белков и других биомолекул, либо окисляются до мочевины, диоксида углерода и служат источником энергии.[36] Окислительный путь катаболизма аминокислот начинается с удаления аминогруппы ферментами трансаминазами. Аминогруппы утилизируются в цикле мочевины; аминокислоты, лишённые аминогрупп называют кетокислотами. Некоторые кетокислоты — промежуточные продукты цикла Кребса. Например, при дезаминировании глутамата образуется альфа-кетоглутаровая кислота.[37] Гликогенные аминокислоты также могут быть преобразованы в глюкозу в реакциях глюконеогенеза.[38]
3. Энергетические превращения
3.1. Окислительное фосфорилирование
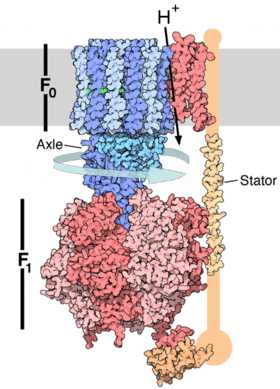
Структура АТФ-синтазы. Канал, по которому проходят протоны и вращающаяся часть фермента показаны синим, субъединицы синтазы — красным, внутренняя мембрана митохондрии — серым цветом.
Подробное рассмотрение темы: Окислительное фосфорилирование, Хемиосмос и МитохондрияПри окислительном фосфорилировании электроны, удалённые из пищевых молекул в метаболических путях (например, в цикле Кребса), переносятся на кислород, а выделяющаяся энергия используется для синтеза АТР. У эукариот данный процесс осуществляется при участии ряда белков, закрепленных в мембранах митохондрий, называемые дыхательной цепью переноса электронов. У прокариот эти белки присутствуют во внутренней мембране клеточной стенки.[39] Белки цепи переноса электронов используют энергию, полученную при передаче электронов от восстановленных молекул (например NADH) на кислород, для перекачки протонов через мембрану.[40]
При перекачке протонов создаётся разница концентраций ионов водорода и возникает электрохимический градиент.[41] Эта сила возвращает протоны обратно в митохондрии через основание АТР-синтазы. Поток протонов заставляет вращаться кольцо из c-субъединиц фермента, в результате чего активный центр синтазы изменяет форму и фосфорилирует аденозиндифосфат, превращая его в АТР.[15]
3.2. Энергия из неорганических соединений
Хемолитотрофами называют прокариот, имеющих особый тип обмена веществ, при котором энергия образуется в результате окисления неорганических соединений. Хемолитотрофы могут окислять молекулярный водород водород,[42] соединения серы (например, сульфиды, сероводород и тиосульфат),[1] оксид железа(II)[43] или аммиак.[44] При этом энергия от окисления этих соединений образуется с помощью акцепторов электронов, таких как кислород или нитриты.[45] Процессы получения энергии из неорганических веществ играют важную роль в таких биогеохимических циклах, как ацетогенез, нитрификация и денитрификация.[46][47]
3.3. Энергия из солнечного света
Энергия солнечного света поглощается растениями, цианобактериями, пурпурными бактериями, зелёными серными бактериями и некоторыми простейшими. Этот процесс часто сочетается с превращением диоксида углерода в органические соединения, как часть процесса фотосинтеза (см. ниже). Системы захвата энергии и фиксации углерода у некоторых прокариот могут работать раздельно (например, у пурпурных и зелёных серных бактерий).[48][49]
У многих организмов поглощение солнечной энергии в принципе аналогично окислительному фосфорилированию, так как при этом энергия запасается в форме градиента концентрации протонов и движущая сила протонов приводит к синтезу АТР.[15] Электроны, необходимые для этой цепи переноса, поступают от светособирающих белков, называемых центрами фотосинтетических реакций (примером являются родопсины). В зависимости от вида фотосинтетических пигментов классифицируют два типа центров реакций; в настоящее время большинство фотосинтезирующих бактерий имеют только один тип, в то время как растения и цианобактерии два.[50]
У растений, водорослей и цианобактерий, фотосистема II использует энергию света для удаления электронов из воды, при этом молекулярный кислород выделяется как побочный продукт реакции. Электроны затем поступают в комплекс цитохрома b6f, который использует энергию для перекачки протонов через тилакоидную мембрану в хлоропластах.[7] Под действием электрохимического градиента протоны движутся обратно через мембрану и запускают АТР-синтазу. Электроны затем проходят через фотосистему I и могут быть использованы для окисления кофермента NADP+, для использования в цикле Кальвина или рециркуляции для образования дополнительных молекул АТР.[51]
4. Анаболизм
Подробное рассмотрение темы: АнаболизмАнаболизм — совокупность метаболических процессов биосинтеза сложных молекул с затратой энергии. Сложные молекулы, входящие в состав клеточных структур, синтезируются последовательно из более простых предшественников. Анаболизм включает три основных этапа, каждый из которых катализируется специализированным ферментом. На первом этапе синтезируются молекулы-предшественники, например, аминокислоты, моносахариды, терпеноиды и нуклеотиды. На втором этапе предшественники с затратой энергии АТР преобразуются в активированные формы. На третьем этапе активированные мономеры объединяются в более сложные молекулы, например, белки, полисахариды, липиды и нуклеиновые кислоты.
Не все живые организмы могут синтезировать все биологически активные молекулы. Автотрофы (например, растения) могут синтезировать сложные органические молекулы из таких простых неорганических низкомолекулярных веществ, как углекислый газ и вода. Гетеротрофам необходим источник более сложных веществ, таких как моносахариды и аминокислоты, для создания более сложных молекул. Организмы классифицируют по их основным источникам энергии: фотоавтотрофы и фотогетеротрофы получают энергию из солнечного света, в то время как хемоавтотрофы и хемогетеротрофы получают энергию из неорганических реакций окисления.
4.1. Связывание углерода
Подробное рассмотрение темы: Фотосинтез и Хемосинтез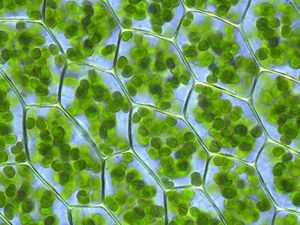
Растительные клетки содержат хлоропласты (зелёного цвета), в тилакоидах которых происходят процессы фотосинтеза. Plagiomnium affine из семейства Mniaceae отдела Настоящие мхи (Bryophyta)
Фотосинтезом называют процесс биосинтеза сахаров из углекислого газа, при котором необходимая энергия поглощается из солнечного света. У растений, цианобактерий и водорослей, при кислородном фотосинтезе происходит фотолиз воды, при этом, как побочный продукт, выделяется кислород. Для преобразования CO2 в 3-фосфоглицерат используется энергия АТР и NADPH, запасенная в фотосистемах. Реакция связывания углерода осуществляется с помощью фермента рибулозобисфосфаткарбоксилазы и является частью цикла Кальвина.[52] У растений классифицируют три типа фотосинтеза — по пути трехуглеродых молекул, по пути четырехуглеродых молекул (С4), и CAM фотосинтез. Три типа фотосинтеза отличаются по пути связывания углекислого газа и его вхождения в цикл Кальвина; у C3 растений связывание CO2 происходит непосредственно в цикле Кальвина, а при С4 и CAM CO2 предварительно включается в состав других соединений. Разные формы фотосинтеза являются приспособлениями к к интенсивному потоку солнечных лучей и к сухим условиям.[53]
У фотосинтезирующих прокариот механизмы связывания углерода более разнообразны. Углекислый газ может быть фиксирован в цикле Кальвина, в обратном цикле Кребса,[54] или в реакциях карбоксилирования ацетил-КоА.[55][56] Прокариоты — хемоавтотрофы также связывают CO2 через цикл Кальвина, но для протекания реакции используют энергию из неорганических соединений.[57]
4.2. Углеводы и гликаны
Подробное рассмотрение темы: Глюконеогенез и ГликозилированиеВ процессе анаболизма сахаров простые органические кислоты могут быть преобразованы в моносахариды, например, в глюкозу, и затем использованы для синтеза полисахаридов, таких как крахмал. Образование глюкозы из соединений, как пируват, лактат, глицерин, 3-фосфоглицерат и аминокислот называют глюконеогенезом. В процессе глюконеогенеза пируват превращается глюкозо-6-фосфат через ряд промежуточных соединений, многие из которых образуются и при гликолизе.[35] Однако, глюконеогенез не просто является гликолизом в обратном направлении, так как несколько химических реакций катализируют специальные ферменты, что дает возможность независимо регулировать процессы образования и распада глюкозы.[58][59]
Многие организмы запасают питательные вещества в форме липидов и жиров, однако, позвоночные не имеют ферментов, катализирующих превращение ацетил-КоА (продукта метаболизма жирных кислот) в пируват (субстрат глюконеогенеза).[60] После длительного голодания позвоночные начинают синтезировать кетоновые тела из жирных кислот, которые могут заменять глюкозу в таких тканях, таких как головной мозг.[61] У растений и бактерии, данная метаболическая проблема решается использованием глиоксилатного цикла, который обходит этап декарбоксилирования в цикле лимонной кислоты и позволяет превращать ацетил-КоА в оксалоацетат, и далее использовать для синтеза глюкозы.[60][62]
Полисахариды выполняют структурные и метаболические функции, а также могут быть соединены с липидами (гликолипиды) и белками (гликопротеиды) при помощи ферментов олигосахаридтрансфераз.[63][64]
4.3. Жирные кислоты, изопреноиды и стероиды
Подробное рассмотрение темы: Стероиды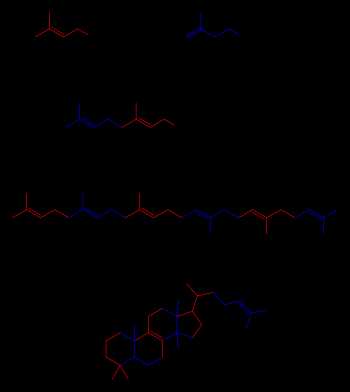
Синтез стероидов из изопентилпирофосфата, диметилаллилпирофосфата, геранилпирофосфата и сквалена. Некоторые промежуточные продукты не показаны
Жирные кислоты образуются синтазами жирных кислот из ацетил-КоА. Углеродный скелет жирных кислот удлиняется в цикле реакций, в которых сначала присоединяется ацетильная группа, далее карбонильная группа восстанавливается до гидроксильной, затем происходит дегидратация и последующее восстановление. Ферменты биосинтеза жирных кислот классифицируют на две группы: у животных и грибов все реакции синтеза жирных кислот осуществляются одним многофункциональным белком I типа,[65] в пластидах растений и у бактерий каждый этап катализируют отдельные ферменты II типа.[66][67]
Терпены и терпеноиды являются представителями самого многочисленного класса растительных натуральных продуктов.[68] Представители данной группы веществ являются производными изопрена и образуются из активированных предшественников изопентилпирофосфата и диметилаллилпирофосфата, которые, в свою очередь, образуются в разных реакциях обмена веществ.[69] У животных и архей изопентилпирофосфат и диметилаллилпирофосфат синтезируются из ацетил-КоА в мевалонатном пути,[70] в то время как у растений и бактерий субстратами не-мевалонатного пути являются пируват и глицеральдегид-3-фосфат.[69][71] В реакциях биосинтеза стероидов молекулы изопрена объединяются и образуют сквалены, которые далее формируют циклические структуры с образованием ланостерола.[72] Ланостерол может быть преобразован в другие стероиды, например холестерин и эргостерин.[72][73]
4.4. Белки
Подробное рассмотрение темы: Биосинтез белкаОрганизмы различаются по способности к синтезу 20 общих аминокислот. Большинство бактерий и растений могут синтезировать все 20, но млекопитающие способны синтезировать лишь 11 заменимых аминокислот.[7] Таким образом, в случае млекопитающих 9 незаменимых аминокислот должны быть получены из пищи. Все аминокислоты синтезируются из промежуточных продуктов гликолиза, цикла лимонной кислоты или пентозомонофосфатного пути. Перенос аминогрупп с аминокислот на альфа-кетокислоты называется трансаминированием. Донорами аминогрупп являются глутамат и глутамин. [74]
Аминокислоты, соединенными пептидными связями, образуют белки. Каждый белок имеет уникальную последовательность аминокислотных остатков (первичная структура белка). Подобно тому, как буквы алфавита могут комбинироваться с образованием почти бесконечных вариаций слов, аминокислоты могут связываться в той или иной последовательности и формировать разнообразные белки. Фермент Аминоацил-тРНК-синтетаза катализирует АТР-зависимое присоединение аминокислот к тРНК сложноэфирными связями, при этом образуются аминоацил-тРНК.[75] Аминоацил-тРНК являются субстратами для рибосом, которая объединяют аминокислоты в длинные полипептидные цепочки, используя матрицу мРНК.[76]
4.5. Нуклеотиды
Подробное рассмотрение темы: Пурин, пиримидинНуклеотиды образуются из аминокислот, углекислого газа и муравьиной кислоты в цепи реакций, для протекания которых требуется большое количество энергии.[77][78] Именно поэтому большинство организмов имеют эффективные системы сохранения ранее синтезированных нуклеотидов и азотистых оснований.[77][79] Пурины синтезируются как нуклеозиды (в основном связанные с рибозой). Аденин и гуанин образуются из инозин-монофосфата, который синтезируется из глицина, глутамина и аспартата при участии метенил-тетрагидрофолата. Пиримидины синтезируются из оротата, который образуется из глутамина и аспартата.[80]
5. Ксенобиотики и окислительный метаболизм
Подробное рассмотрение темы: АнтиоксидантыВсе организмы постоянно подвергаются воздействию соединений, накопление которых может быть вредно для клеток. Такие потенциально опасные чужеродные соединения называются ксенобиотиками.[81] Ксенобиотики, например синтетические лекарства и яды природного происхождения, детоксифицируются специализированными ферментами. У человека такие ферменты представлены, например, цитохром-оксидазами,[82] глюкуронилтрансферазой,[83] и глутатион S-трансферазой.[84] Эта система ферментов действует в три этапа: на первой стадии ксенобиотики окисляются, затем происходит конъюгирование водорастворимых групп в молекулы, далее модифицированные водорастворимые ксенобиотики могут быть удалены из клеток и метаболизированы перед их экскрецией. Описанные реакции играют важную роль в разложении микробами загрязняющих веществ и биоремедиации загрязнённых земель и разливов нефти.[85] Многие подобные реакции протекают при участии многоклеточных организмов, однако, ввиду невероятного разнообразия, микроорганизмы справляются с гораздо более широким спектром ксенобиотиков, чем многоклеточные организмы, и способны даже разрушать стойкие органические загрязнители, например хлорорганические соединения.[86]
Связанной с этим проблемой для аэробных организмов является оксидативный стресс.[87] В процессе окислительного фосфорилирования и образования дисульфидных связей при укладке белка образуются активные формы кислорода, например пероксид водорода.[88] Эти повреждающие оксиданты удаляются антиоксидантами, например глутатионом и ферментами каталазой и пероксидазами.[89][90]
6. Термодинамика живых организмов
Живые организмы подчиняются началам термодинамики, которые описывают превращения тепла и работы. Второе начало термодинамики гласит, что в любой изолированной системе энтропия не уменьшается. Хотя невероятная сложность живых организмов очевидно противоречит этому закону, жизнь возможна, так как все организмы открытые системы, которые обмениваются веществом и энергией с окружающей средой. Таким образом живые системы не находятся в термодинамическом равновесии, но вместо этого выступает диссипативной системой, которая поддерживают своё состояние сложно организованности, вызывая большее увеличение энтропии окружающей средой.[91] В метаболизм клеток это достигается путём сочетания спонтанных процессов катаболизма с не спонтанных процессов анаболизма. В термодинамических условиях, метаболизм поддерживает порядок за счёт создания беспорядка.[92]
7. Регуляция и контроль
Подробное рассмотрение темы: Гормоны, Передача сигнала в клеткеГомеостазом называют постоянство внутренней среды организма. Так как внешняя среда, окружающая большинство организмов, постоянно меняется, для поддержания постоянных условий внутри клеток, реакции обмена веществ должны точно регулироваться.[93][94] Регуляция метаболизма позволяет организмам отвечать на сигналы и активно взаимодействовать с окружающей средой.[95] В случае фермента, регуляция заключается в повышении и снижении его активности в ответ на сигналы. С другой стороны, фермент оказывает некоторый контроль над метаболическим путем, который определяется как эффект от изменения активности фермента на данный метаболический путь.[96]
Влияние инсулина на поглощение глюкозы и обмен веществ. Инсулин связывается со своим рецептором (1), который в свою очередь запускает касакад реакций активации множества белков (2). К ним относятся: транслокация переносчика GLUT-4 к плазматической мембране и поступление глюкозы в клетку (3), синтез гликогена (4), гликолиз (5) и синтез жирных кислот (6).
Выделяют несколько уровней регуляции метаболизма. В метаболическом пути происходит саморегуляция на уровне субстрата или продукта; например, уменьшение количества продукта может компенсировано увеличить поток субстрата реакции по данному пути пути.[97] Этот тип регулирования часто включает аллостерическое регулирование активности некоторых ферментов в метаболических путях.[98] Внешний контроль включает клетку многоклеточного организма, изменяющую свой метаболизм в ответ на сигналы от других клеток. Эти сигналы, как правило, в виде растворимых мессенджеров, например гормоны и факторы роста, определяются специфическими рецепторами на поверхности клеток.[99] Затем эти сигналы передаются внутрь клетки системой вторичных мессенджеров, которые зачастую связаны с фосфорилированием белков.[100]
Хорошо изученный пример внешнего контроля — регуляция метаболизма глюкозы инсулином.[101] Инсулин вырабатывается в ответ на повышение уровня глюкозы в крови. Гормон связывается с инсулиновым рецептором на поверхности клетки, затем активируется каскад протеинкиназ, которые обеспечивают поглощение молекул глюкозы клетками и преобразовать их в молекулы жирных кислот и гликогена.[102] Метаболизм гликогена контролируется активностью фосфорилазы (фермента, который расщепляет гликоген) и гликогенсинтазы (фермента, который образует его). Эти ферменты взаимосвязаны; фосфорилирование ингибируется гликогенсинтазой, но активируется фосфорилазой. Инсулин вызывает синтез гликогена путём активации белковых фосфатаз и уменьшает фосфорилирование этих ферментов.[103]
8. Эволюция
Подробное рассмотрение темы: Филогенетика
Филогенетическое дерево показывает общее происхождение организмов из всех трёх доменов. Бактерии окрашены голубым, эукариоты — красным, и археи — зелёным. Взаимосвязанные позиции некоторых типов показаны вокруг дерева.
Главные пути метаболизма, описанные выше, например, гликолиза и цикла Кребса, присутствуют у всех трёх доменах живых существ и обнаруживаются у последнего универсального общего предка.[3][104] Этот универсальный предок был прокариотом и, вероятно, метаногеном с аминокислотным, нуклеотидным, углеводным и липидным метаболизмом.[105][106] Сохранение этих древних метаболических путей в эволюции может быть результатом того, что эти реакции оптимальны для решения конкретных проблем с метаболизмом. Так, конечные продукты гликолиза и цикла Кребса образуются с высокой эффективностью и с минимальным количеством стадий.[4][5] Первые метаболические пути на основе ферментов могли быть частями пуринового метаболизма нуклеотидов с предыдущим метаболических путей были частью древнего мира РНК.[107]
Многие модели были предложены для описания механизмов, посредством которых новые метаболические пути эволюционировали. К ним относятся последовательное добавление новых ферментов на короткий предковый путь, дупликация, а затем дивергенция всех путей, а также набор уже существующих ферментов и их сборка в новый путь реакций.[108] Относительную важность этих механизмов неясна, однако геномные исследования показали, что ферменты в метаболическом пути, скорее всего, имеют общее происхождение, предполагая, что многие пути эволюционировали шаг за шагом с новыми функциями, созданными из уже существующих этапов пути.[109] Альтернативная модель основана на исследованиях, в которых прослеживается эволюция структуры белков в метаболических связях; предполагают, что ферменты собирались для выполнения схожих функций в различных метаболических путях[110] Эти процессы сборки привели к эволюционированию ферментативной мозаики.[111] Некоторые части обмена веществ возможно существовали в качестве «модулей», которые могли быть повторно использованы в различных путях для выполнения схожих функций.[112]
Эволюция также может приводить к потере метаболических функций. Например, у некоторых паразитов метаболические процессы, которые не важны для выживания, утрачены и готовые аминокислоты, нуклеотиды и углеводы получаются от хозяина.[113] Подобные упрощения метаболических возможностей наблюдают у эндосимбиотических организмов.[114]
9. Методы исследования
Подробное рассмотрение темы: Протеомика, Метабономика
Метаболическая сеть цикла Кребса Arabidopsis thaliana. Ферменты и метаболиты обозначены красными квадратами, а взаимодействия между ними — как чёрные линии.
Классически, метаболизм изучается упрощённым подходом, который фокусируется на одном метаболическом пути. Особенно ценно использование меченых атомов на организменном, тканевом и клеточном уровнях, которые определяют пути от предшественников до конечных продуктов путём выявления радиоактивно меченых промежуточных продуктов.[115] Ферменты, которые катализируют эти химические реакции, могут затем быть выделены для исследования их кинетики и ответа на ингибиторы. Параллельный подход заключается в выявлении небольших молекул в клетки или ткани; полный набор этих молекул называется метаболом. В целом, эти исследования дают хорошее представление о структуре и функциях простых путей метаболизма, но недостаточны в применении к более сложных системам, например полной метаболизм клетки.[116]
Идея сложности метаболических сетей в клетках, которые содержат тысячи различных ферментов, отражена на изображении справа, показывающее взаимодействия только между 43 белками и 40 метаболитами, которые регулируются 45000 генов.[117] Тем не менее, сейчас можно использовать такие данные о геномах для воссоздания полной сети биохимических реакций и образовывать более целостные математические модели, которые могут объяснить и предсказать их поведение.[118] Эти модели особенно сильны, когда используются для интеграции данных о путях и метаболитах, полученных на основе классических методов, с данными по экспрессии генов из протеомных и ДНК-микрочиповых исследований.[119] С помощью этих методов, модель человеческого метаболизма в настоящее время создаётся, которая будет служить ориентиром для будущих исследований лекарств и биохимических исследований.[120] Эти модели в настоящее время используются в анализах сети, для классификации болезней человека по группам, которые различаются по общим белкам или метаболитам.[121][122]
Яркий пример бактериальных метаболических сетей — устройство галстук-бабочки[123][124][125], структура которой позволяет вводить широкий спектр питательных веществ и производить большое разнообразие продуктов и сложных макромолекул, используя сравнительно немного общих промежуточных веществ.
Основная технологическая основа этой информации — метаболическая инженерия. Здесь организмы, например дрожжи, растения или бактерии, генетически модифицируются, чтобы сделать их более эффективными в биотехнологии и помочь в производстве лекарств, например антибиотиков или промышленных химических веществ, таких как 1,3-пропандиола и шикимовой кислоты.[126] Эти генетические модификации обычно направлены на уменьшение количества энергии, используемой для производства продукции, повышения урожайности и снижения производственных отходов.[127]
10. История
Санторио взвешивает сам себя до и после принятия пищи, из Ars de statica medicina, впервые опубликованной в 1614 году
История изучения метаболизма охватывает несколько столетий. Исследования начинались с изучения организмов животных, в современной биохимии изучают отдельные метаболические реакции. Понятие обмена веществ впервые встречается в работах Ибн аль-Нафиса (1213—1288), который писал, что «тело и его части находятся в постоянном состоянии распада и питания, так что оно неизбежно претерпевает постоянные изменения».[128] Первые контролируемые эксперименты по метаболизму у человека были опубликованы Санторио Санторио в 1614 году в книге итал. Ars de statica medicina.[129] Он рассказал, как он сам взвесил себя до и после приёма пищи, сна, работы, секса, натощак, после питья и выделения мочи. Он обнаружил, что большая часть пищи, которую он принял, была утрачена в результате процесса, названного «незаметным испарением».
В ранних исследованиях механизмы метаболических реакций не были обнаружены и считалось, что живой тканью управляет живая сила.[130] В XIX веке при исследовании ферментации сахара спирта дрожжами Луи Пастер сделал вывод, что брожение катализируется веществами из дрожжевых клеток, которые он назвал ферментами. Пастер писал, что «алкогольное брожение — действие, связанное с жизнью и организуется дрожжевыми клетками, не связано со смертью или разложением клеток».[131] Это открытие, вместе с публикацией Фридриха Вёлера в 1828 году о химическом синтезе мочевины,[132] доказали, что органические соединения и химические реакции, обнаруженные в клетках, не имеют различий в принципе, как и любые другие разделы химии.
Открытие ферментов в начале XX века Эдуардом Бухнером разделило изучение метаболических реакций от изучения клеток и дало начало развитию биохимии как науки.[133] Одним из успешных биохимиков начала двадцатого века был Ханс Адольф Кребс, который внёс огромный вклад в изучение метаболизма.[134] Кребс описал цикл мочевины и позднее, работая вместе с Хансом Корнбергом, цикл лимонной кислоты и глиоксилатный цикл.[135][62] В современных биохимических исследованиях широко используют новые методы, такие как хроматография, рентгеноструктурный анализ, ЯМР-спектроскопия, электронная микроскопия и метод классической молекулярной динамики. Эти методы позволяют открывать и подробно изучать множество молекул и метаболических путей в клетках.
Примечания
- ↑ 12Friedrich C (1998). «Physiology and genetics of sulfur-oxidizing bacteria». Adv Microb Physiol 39: 235–89. DOI:10.1016/S0065-2911(08)60018-1 - dx.doi.org/10.1016/S0065-2911(08)60018-1. PMID 9328649 - www.ncbi.nlm.nih.gov/pubmed/9328649?dopt=Abstract.
- Pace NR (January 2001). «The universal nature of biochemistry». Proc. Natl. Acad. Sci. U.S.A. 98 (3): 805–8. DOI:10.1073/pnas.98.3.805 - dx.doi.org/10.1073/pnas.98.3.805. PMID 11158550 - www.ncbi.nlm.nih.gov/pubmed/11158550?dopt=Abstract.
- ↑ 12Smith E, Morowitz H (2004). «Universality in intermediary metabolism - www.pnas.org/cgi/pmidlookup?view=long&pmid=15340153». Proc Natl Acad Sci USA 101 (36): 13168–73. DOI:10.1073/pnas.0404922101 - dx.doi.org/10.1073/pnas.0404922101. PMID 15340153 - www.ncbi.nlm.nih.gov/pubmed/15340153?dopt=Abstract.
- ↑ 12Ebenhöh O, Heinrich R (2001). «Evolutionary optimization of metabolic pathways. Theoretical reconstruction of the stoichiometry of ATP and NADH producing systems». Bull Math Biol 63 (1): 21–55. DOI:10.1006/bulm.2000.0197 - dx.doi.org/10.1006/bulm.2000.0197. PMID 11146883 - www.ncbi.nlm.nih.gov/pubmed/11146883?dopt=Abstract.
- ↑ 12Meléndez-Hevia E, Waddell T, Cascante M (1996). «The puzzle of the Krebs citric acid cycle: assembling the pieces of chemically feasible reactions, and opportunism in the design of metabolic pathways during evolution». J Mol Evol 43 (3): 293–303. DOI:10.1007/BF02338838 - dx.doi.org/10.1007/BF02338838. PMID 8703096 - www.ncbi.nlm.nih.gov/pubmed/8703096?dopt=Abstract.
- Michie K, Löwe J (2006). «Dynamic filaments of the bacterial cytoskeleton». Annu Rev Biochem 75: 467–92. DOI:10.1146/annurev.biochem.75.103004.142452 - dx.doi.org/10.1146/annurev.biochem.75.103004.142452. PMID 16756499 - www.ncbi.nlm.nih.gov/pubmed/16756499?dopt=Abstract.
- ↑ 123456Nelson David L. Lehninger Principles of Biochemistry. — New York: W. H. Freeman and company, 2005. — P. 841. — ISBN 0-7167-4339-6
- Fahy E, Subramaniam S, Brown H, Glass C, Merrill A, Murphy R, Raetz C, Russell D, Seyama Y, Shaw W, Shimizu T, Spener F, van Meer G, VanNieuwenhze M, White S, Witztum J, Dennis E (2005). «A comprehensive classification system for lipids - www.jlr.org/cgi/content/full/46/5/839». J Lipid Res 46 (5): 839–61. DOI:10.1194/jlr.E400004-JLR200 - dx.doi.org/10.1194/jlr.E400004-JLR200. PMID 15722563 - www.ncbi.nlm.nih.gov/pubmed/15722563?dopt=Abstract.
- Nomenclature of Lipids - www.chem.qmul.ac.uk/iupac/lipid/. IUPAC-IUB Commission on Biochemical Nomenclature (CBN).
- Hegardt F (1999). «Mitochondrial 3-hydroxy-3-methylglutaryl-CoA synthase: a control enzyme in ketogenesis». Biochem J 338 (Pt 3): 569–82. DOI:10.1042/0264-6021:3380569 - dx.doi.org/10.1042/0264-6021:3380569. PMID 10051425 - www.ncbi.nlm.nih.gov/pubmed/10051425?dopt=Abstract.
- Raman R, Raguram S, Venkataraman G, Paulson J, Sasisekharan R (2005). «Glycomics: an integrated systems approach to structure-function relationships of glycans». Nat Methods 2 (11): 817–24. DOI:10.1038/nmeth807 - dx.doi.org/10.1038/nmeth807. PMID 16278650 - www.ncbi.nlm.nih.gov/pubmed/16278650?dopt=Abstract.
- Sierra S, Kupfer B, Kaiser R (2005). «Basics of the virology of HIV-1 and its replication». J Clin Virol 34 (4): 233–44. DOI:10.1016/j.jcv.2005.09.004 - dx.doi.org/10.1016/j.jcv.2005.09.004. PMID 16198625 - www.ncbi.nlm.nih.gov/pubmed/16198625?dopt=Abstract.
- ↑ 12Wimmer M, Rose I (1978). «Mechanisms of enzyme-catalyzed group transfer reactions». Annu Rev Biochem 47: 1031–78. DOI:10.1146/annurev.bi.47.070178.005123 - dx.doi.org/10.1146/annurev.bi.47.070178.005123. PMID 354490 - www.ncbi.nlm.nih.gov/pubmed/354490?dopt=Abstract.
- Mitchell P (1979). «The Ninth Sir Hans Krebs Lecture. Compartmentation and communication in living systems. Ligand conduction: a general catalytic principle in chemical, osmotic and chemiosmotic reaction systems». Eur J Biochem 95 (1): 1–20. DOI:10.1111/j.1432-1033.1979.tb12934.x - dx.doi.org/10.1111/j.1432-1033.1979.tb12934.x. PMID 378655 - www.ncbi.nlm.nih.gov/pubmed/378655?dopt=Abstract.
- ↑ 1234Dimroth P, von Ballmoos C, Meier T (March 2006). «Catalytic and mechanical cycles in F-ATP synthases. Fourth in the Cycles Review Series». EMBO Rep 7 (3): 276–82. DOI:10.1038/sj.embor.7400646 - dx.doi.org/10.1038/sj.embor.7400646. PMID 16607397 - www.ncbi.nlm.nih.gov/pubmed/16607397?dopt=Abstract.
- Stanford School of Medicine Nutrition Courses. — SUMMIT, 2006.
- Pollak N, Dölle C, Ziegler M (2007). «The power to reduce: pyridine nucleotides—small molecules with a multitude of functions». Biochem J 402 (2): 205–18. DOI:10.1042/BJ20061638 - dx.doi.org/10.1042/BJ20061638. PMID 17295611 - www.ncbi.nlm.nih.gov/pubmed/17295611?dopt=Abstract.
- ↑ 12Heymsfield S, Waki M, Kehayias J, Lichtman S, Dilmanian F, Kamen Y, Wang J, Pierson R (1991). «Chemical and elemental analysis of humans in vivo using improved body composition models». Am J Physiol 261 (2 Pt 1): E190–8. PMID 1872381 - www.ncbi.nlm.nih.gov/pubmed/1872381?dopt=Abstract.
- Sychrová H (2004). «Yeast as a model organism to study transport and homeostasis of alkali metal cations - www.biomed.cas.cz/physiolres/pdf/53 Suppl 1/53_S91.pdf» (PDF). Physiol Res 53 Suppl 1: S91–8. PMID 15119939 - www.ncbi.nlm.nih.gov/pubmed/15119939?dopt=Abstract.
- Levitan I (1988). «Modulation of ion channels in neurons and other cells». Annu Rev Neurosci 11: 119–36. DOI:10.1146/annurev.ne.11.030188.001003 - dx.doi.org/10.1146/annurev.ne.11.030188.001003. PMID 2452594 - www.ncbi.nlm.nih.gov/pubmed/2452594?dopt=Abstract.
- Dulhunty A (2006). «Excitation-contraction coupling from the 1950s into the new millennium». Clin Exp Pharmacol Physiol 33 (9): 763–72. DOI:10.1111/j.1440-1681.2006.04441.x - dx.doi.org/10.1111/j.1440-1681.2006.04441.x. PMID 16922804 - www.ncbi.nlm.nih.gov/pubmed/16922804?dopt=Abstract.
- Mahan D, Shields R (1998). «Macro- and micromineral composition of pigs from birth to 145 kilograms of body weight - jas.fass.org/cgi/reprint/76/2/506». J Anim Sci 76 (2): 506–12. PMID 9498359 - www.ncbi.nlm.nih.gov/pubmed/9498359?dopt=Abstract.
- Husted S, Mikkelsen B, Jensen J, Nielsen N (2004). «Elemental fingerprint analysis of barley (Hordeum vulgare) using inductively coupled plasma mass spectrometry, isotope-ratio mass spectrometry, and multivariate statistics». Anal Bioanal Chem 378 (1): 171–82. DOI:10.1007/s00216-003-2219-0 - dx.doi.org/10.1007/s00216-003-2219-0. PMID 14551660 - www.ncbi.nlm.nih.gov/pubmed/14551660?dopt=Abstract.
- Finney L, O'Halloran T (2003). «Transition metal speciation in the cell: insights from the chemistry of metal ion receptors». Science 300 (5621): 931–6. DOI:10.1126/science.1085049 - dx.doi.org/10.1126/science.1085049. PMID 12738850 - www.ncbi.nlm.nih.gov/pubmed/12738850?dopt=Abstract.
- Cousins R, Liuzzi J, Lichten L (2006). «Mammalian zinc transport, trafficking, and signals - www.jbc.org/cgi/content/full/281/34/24085». J Biol Chem 281 (34): 24085–9. DOI:10.1074/jbc.R600011200 - dx.doi.org/10.1074/jbc.R600011200. PMID 16793761 - www.ncbi.nlm.nih.gov/pubmed/16793761?dopt=Abstract.
- Dunn L, Rahmanto Y, Richardson D (2007). «Iron uptake and metabolism in the new millennium». Trends Cell Biol 17 (2): 93–100. DOI:10.1016/j.tcb.2006.12.003 - dx.doi.org/10.1016/j.tcb.2006.12.003. PMID 17194590 - www.ncbi.nlm.nih.gov/pubmed/17194590?dopt=Abstract.
- Nealson K, Conrad P (1999). «Life: past, present and future - rstb.royalsocietypublishing.org/cgi/pmidlookup?view=long&pmid=10670014». Philos Trans R Soc Lond B Biol Sci 354 (1392): 1923–39. DOI:10.1098/rstb.1999.0532 - dx.doi.org/10.1098/rstb.1999.0532. PMID 10670014 - www.ncbi.nlm.nih.gov/pubmed/10670014?dopt=Abstract.
- Nelson N, Ben-Shem A (2004). «The complex architecture of oxygenic photosynthesis». Nat Rev Mol Cell Biol 5 (12): 971–82. DOI:10.1038/nrm1525 - dx.doi.org/10.1038/nrm1525. PMID 15573135 - www.ncbi.nlm.nih.gov/pubmed/15573135?dopt=Abstract.
- Häse C, Finkelstein R (December 1993). «Bacterial extracellular zinc-containing metalloproteases - mmbr.asm.org/cgi/pmidlookup?view=long&pmid=8302217». Microbiol Rev 57 (4): 823–37. PMID 8302217 - www.ncbi.nlm.nih.gov/pubmed/8302217?dopt=Abstract.
- Gupta R, Gupta N, Rathi P (2004). «Bacterial lipases: an overview of production, purification and biochemical properties». Appl Microbiol Biotechnol 64 (6): 763–81. DOI:10.1007/s00253-004-1568-8 - dx.doi.org/10.1007/s00253-004-1568-8. PMID 14966663 - www.ncbi.nlm.nih.gov/pubmed/14966663?dopt=Abstract.
- Hoyle T (1997). «The digestive system: linking theory and practice». Br J Nurs 6 (22): 1285–91. PMID 9470654 - www.ncbi.nlm.nih.gov/pubmed/9470654?dopt=Abstract.
- Souba W, Pacitti A (1992). «How amino acids get into cells: mechanisms, models, menus, and mediators». JPEN J Parenter Enteral Nutr 16 (6): 569–78. DOI:10.1177/0148607192016006569 - dx.doi.org/10.1177/0148607192016006569. PMID 1494216 - www.ncbi.nlm.nih.gov/pubmed/1494216?dopt=Abstract.
- Barrett M, Walmsley A, Gould G (1999). «Structure and function of facilitative sugar transporters». Curr Opin Cell Biol 11 (4): 496–502. DOI:10.1016/S0955-0674(99)80072-6 - dx.doi.org/10.1016/S0955-0674(99)80072-6. PMID 10449337 - www.ncbi.nlm.nih.gov/pubmed/10449337?dopt=Abstract.
- Bell G, Burant C, Takeda J, Gould G (1993). «Structure and function of mammalian facilitative sugar transporters». J Biol Chem 268 (26): 19161–4. PMID 8366068 - www.ncbi.nlm.nih.gov/pubmed/8366068?dopt=Abstract.
- ↑ 12Bouché C, Serdy S, Kahn C, Goldfine A (2004). «The cellular fate of glucose and its relevance in type 2 diabetes - edrv.endojournals.org/cgi/content/full/25/5/807». Endocr Rev 25 (5): 807–30. DOI:10.1210/er.2003-0026 - dx.doi.org/10.1210/er.2003-0026. PMID 15466941 - www.ncbi.nlm.nih.gov/pubmed/15466941?dopt=Abstract.
- Sakami W, Harrington H (1963). «Amino acid metabolism». Annu Rev Biochem 32: 355–98. DOI:10.1146/annurev.bi.32.070163.002035 - dx.doi.org/10.1146/annurev.bi.32.070163.002035. PMID 14144484 - www.ncbi.nlm.nih.gov/pubmed/14144484?dopt=Abstract.
- Brosnan J (2000). «Glutamate, at the interface between amino acid and carbohydrate metabolism - jn.nutrition.org/cgi/content/full/130/4/988S». J Nutr 130 (4S Suppl): 988S–90S. PMID 10736367 - www.ncbi.nlm.nih.gov/pubmed/10736367?dopt=Abstract.
- Young V, Ajami A (2001). «Glutamine: the emperor or his clothes? - jn.nutrition.org/cgi/content/full/131/9/2449S». J Nutr 131 (9 Suppl): 2449S–59S; discussion 2486S–7S. PMID 11533293 - www.ncbi.nlm.nih.gov/pubmed/11533293?dopt=Abstract.
- Hosler J, Ferguson-Miller S, Mills D (2006). «Energy transduction: proton transfer through the respiratory complexes». Annu Rev Biochem 75: 165–87. DOI:10.1146/annurev.biochem.75.062003.101730 - dx.doi.org/10.1146/annurev.biochem.75.062003.101730. PMID 16756489 - www.ncbi.nlm.nih.gov/pubmed/16756489?dopt=Abstract.
- Schultz B, Chan S (2001). «Structures and proton-pumping strategies of mitochondrial respiratory enzymes». Annu Rev Biophys Biomol Struct 30: 23–65. DOI:10.1146/annurev.biophys.30.1.23 - dx.doi.org/10.1146/annurev.biophys.30.1.23. PMID 11340051 - www.ncbi.nlm.nih.gov/pubmed/11340051?dopt=Abstract.
- Capaldi R, Aggeler R (2002). «Mechanism of the F(1)F(0)-type ATP synthase, a biological rotary motor». Trends Biochem Sci 27 (3): 154–60. DOI:10.1016/S0968-0004(01)02051-5 - dx.doi.org/10.1016/S0968-0004(01)02051-5. PMID 11893513 - www.ncbi.nlm.nih.gov/pubmed/11893513?dopt=Abstract.
- Friedrich B, Schwartz E (1993). «Molecular biology of hydrogen utilization in aerobic chemolithotrophs». Annu Rev Microbiol 47: 351–83. DOI:10.1146/annurev.mi.47.100193.002031 - dx.doi.org/10.1146/annurev.mi.47.100193.002031. PMID 8257102 - www.ncbi.nlm.nih.gov/pubmed/8257102?dopt=Abstract.
- Weber K, Achenbach L, Coates J (2006). «Microorganisms pumping iron: anaerobic microbial iron oxidation and reduction». Nat Rev Microbiol 4 (10): 752–64. DOI:10.1038/nrmicro1490 - dx.doi.org/10.1038/nrmicro1490. PMID 16980937 - www.ncbi.nlm.nih.gov/pubmed/16980937?dopt=Abstract.
- Jetten M, Strous M, van de Pas-Schoonen K, Schalk J, van Dongen U, van de Graaf A, Logemann S, Muyzer G, van Loosdrecht M, Kuenen J (1998). «The anaerobic oxidation of ammonium». FEMS Microbiol Rev 22 (5): 421–37. DOI:10.1111/j.1574-6976.1998.tb00379.x - dx.doi.org/10.1111/j.1574-6976.1998.tb00379.x. PMID 9990725 - www.ncbi.nlm.nih.gov/pubmed/9990725?dopt=Abstract.
- Simon J (2002). «Enzymology and bioenergetics of respiratory nitrite ammonification». FEMS Microbiol Rev 26 (3): 285–309. DOI:10.1111/j.1574-6976.2002.tb00616.x - dx.doi.org/10.1111/j.1574-6976.2002.tb00616.x. PMID 12165429 - www.ncbi.nlm.nih.gov/pubmed/12165429?dopt=Abstract.
- Conrad R (1996). «Soil microorganisms as controllers of atmospheric trace gases (h3, CO, Ch5, OCS, N2O, and NO) - mmbr.asm.org/cgi/pmidlookup?view=long&pmid=8987358». Microbiol Rev 60 (4): 609–40. PMID 8987358 - www.ncbi.nlm.nih.gov/pubmed/8987358?dopt=Abstract.
- Barea J, Pozo M, Azcón R, Azcón-Aguilar C (2005). «Microbial co-operation in the rhizosphere - jxb.oxfordjournals.org/cgi/content/full/56/417/1761». J Exp Bot 56 (417): 1761–78. DOI:10.1093/jxb/eri197 - dx.doi.org/10.1093/jxb/eri197. PMID 15911555 - www.ncbi.nlm.nih.gov/pubmed/15911555?dopt=Abstract.
- van der Meer M, Schouten S, Bateson M, Nübel U, Wieland A, Kühl M, de Leeuw J, Sinninghe Damsté J, Ward D (July 2005). «Diel variations in carbon metabolism by green nonsulfur-like bacteria in alkaline siliceous hot spring microbial mats from Yellowstone National Park - aem.asm.org/cgi/pmidlookup?view=long&pmid=16000812». Appl Environ Microbiol 71 (7): 3978–86. DOI:10.1128/AEM.71.7.3978-3986.2005 - dx.doi.org/10.1128/AEM.71.7.3978-3986.2005. PMID 16000812 - www.ncbi.nlm.nih.gov/pubmed/16000812?dopt=Abstract.
- Tichi M, Tabita F (2001). «Interactive control of Rhodobacter capsulatus redox-balancing systems during phototrophic metabolism - jb.asm.org/cgi/pmidlookup?view=long&pmid=11591679». J Bacteriol 183 (21): 6344–54. DOI:10.1128/JB.183.21.6344-6354.2001 - dx.doi.org/10.1128/JB.183.21.6344-6354.2001. PMID 11591679 - www.ncbi.nlm.nih.gov/pubmed/11591679?dopt=Abstract.
- Allen J, Williams J (1998). «Photosynthetic reaction centers». FEBS Lett 438 (1–2): 5–9. DOI:10.1016/S0014-5793(98)01245-9 - dx.doi.org/10.1016/S0014-5793(98)01245-9. PMID 9821949 - www.ncbi.nlm.nih.gov/pubmed/9821949?dopt=Abstract.
- Munekage Y, Hashimoto M, Miyake C, Tomizawa K, Endo T, Tasaka M, Shikanai T (2004). «Cyclic electron flow around photosystem I is essential for photosynthesis». Nature 429 (6991): 579–82. DOI:10.1038/nature02598 - dx.doi.org/10.1038/nature02598. PMID 15175756 - www.ncbi.nlm.nih.gov/pubmed/15175756?dopt=Abstract.
- Miziorko H, Lorimer G (1983). «Ribulose-1,5-bisphosphate carboxylase-oxygenase». Annu Rev Biochem 52: 507–35. DOI:10.1146/annurev.bi.52.070183.002451 - dx.doi.org/10.1146/annurev.bi.52.070183.002451. PMID 6351728 - www.ncbi.nlm.nih.gov/pubmed/6351728?dopt=Abstract.
- Dodd A, Borland A, Haslam R, Griffiths H, Maxwell K (2002). «Crassulacean acid metabolism: plastic, fantastic - jxb.oxfordjournals.org/cgi/content/full/53/369/569». J Exp Bot 53 (369): 569–80. DOI:10.1093/jexbot/53.369.569 - dx.doi.org/10.1093/jexbot/53.369.569. PMID 11886877 - www.ncbi.nlm.nih.gov/pubmed/11886877?dopt=Abstract.
- Hügler M, Wirsen C, Fuchs G, Taylor C, Sievert S (May 2005). «Evidence for autotrophic CO2 fixation via the reductive tricarboxylic acid cycle by members of the epsilon subdivision of proteobacteria - jb.asm.org/cgi/pmidlookup?view=long&pmid=15838028». J Bacteriol 187 (9): 3020–7. DOI:10.1128/JB.187.9.3020-3027.2005 - dx.doi.org/10.1128/JB.187.9.3020-3027.2005. PMID 15838028 - www.ncbi.nlm.nih.gov/pubmed/15838028?dopt=Abstract.
- Strauss G, Fuchs G (1993). «Enzymes of a novel autotrophic CO2 fixation pathway in the phototrophic bacterium Chloroflexus aurantiacus, the 3-hydroxypropionate cycle». Eur J Biochem 215 (3): 633–43. DOI:10.1111/j.1432-1033.1993.tb18074.x - dx.doi.org/10.1111/j.1432-1033.1993.tb18074.x. PMID 8354269 - www.ncbi.nlm.nih.gov/pubmed/8354269?dopt=Abstract.
- Wood H (1991). «Life with CO or CO2 and h3 as a source of carbon and energy - www.fasebj.org/cgi/reprint/5/2/156». FASEB J 5 (2): 156–63. PMID 1900793 - www.ncbi.nlm.nih.gov/pubmed/1900793?dopt=Abstract.
- Shively J, van Keulen G, Meijer W (1998). «Something from almost nothing: carbon dioxide fixation in chemoautotrophs». Annu Rev Microbiol 52: 191–230. DOI:10.1146/annurev.micro.52.1.191 - dx.doi.org/10.1146/annurev.micro.52.1.191. PMID 9891798 - www.ncbi.nlm.nih.gov/pubmed/9891798?dopt=Abstract.
- Boiteux A, Hess B (1981). «Design of glycolysis». Philos Trans R Soc Lond B Biol Sci 293 (1063): 5–22. DOI:10.1098/rstb.1981.0056 - dx.doi.org/10.1098/rstb.1981.0056. PMID 6115423 - www.ncbi.nlm.nih.gov/pubmed/6115423?dopt=Abstract.
- Pilkis S, el-Maghrabi M, Claus T (1990). «Fructose-2,6-bisphosphate in control of hepatic gluconeogenesis. From metabolites to molecular genetics». Diabetes Care 13 (6): 582–99. DOI:10.2337/diacare.13.6.582 - dx.doi.org/10.2337/diacare.13.6.582. PMID 2162755 - www.ncbi.nlm.nih.gov/pubmed/2162755?dopt=Abstract.
- ↑ 12Ensign S (2006). «Revisiting the glyoxylate cycle: alternate pathways for microbial acetate assimilation». Mol Microbiol 61 (2): 274–6. DOI:10.1111/j.1365-2958.2006.05247.x - dx.doi.org/10.1111/j.1365-2958.2006.05247.x. PMID 16856935 - www.ncbi.nlm.nih.gov/pubmed/16856935?dopt=Abstract.
- Finn P, Dice J (2006). «Proteolytic and lipolytic responses to starvation». Nutrition 22 (7–8): 830–44. DOI:10.1016/j.nut.2006.04.008 - dx.doi.org/10.1016/j.nut.2006.04.008. PMID 16815497 - www.ncbi.nlm.nih.gov/pubmed/16815497?dopt=Abstract.
- ↑ 12Kornberg H, Krebs H (1957). «Synthesis of cell constituents from C2-units by a modified tricarboxylic acid cycle». Nature 179 (4568): 988–91. DOI:10.1038/179988a0 - dx.doi.org/10.1038/179988a0. PMID 13430766 - www.ncbi.nlm.nih.gov/pubmed/13430766?dopt=Abstract.
- Opdenakker G, Rudd P, Ponting C, Dwek R (1993). «Concepts and principles of glycobiology - www.fasebj.org/cgi/reprint/7/14/1330». FASEB J 7 (14): 1330–7. PMID 8224606 - www.ncbi.nlm.nih.gov/pubmed/8224606?dopt=Abstract.
- McConville M, Menon A (2000). «Recent developments in the cell biology and biochemistry of glycosylphosphatidylinositol lipids (review)». Mol Membr Biol 17 (1): 1–16. DOI:10.1080/096876800294443 - dx.doi.org/10.1080/096876800294443. PMID 10824734 - www.ncbi.nlm.nih.gov/pubmed/10824734?dopt=Abstract.
- Chirala S, Wakil S (2004). «Structure and function of animal fatty acid synthase». Lipids 39 (11): 1045–53. DOI:10.1007/s11745-004-1329-9 - dx.doi.org/10.1007/s11745-004-1329-9. PMID 15726818 - www.ncbi.nlm.nih.gov/pubmed/15726818?dopt=Abstract.
- White S, Zheng J, Zhang Y (2005). «The structural biology of type II fatty acid biosynthesis». Annu Rev Biochem 74: 791–831. DOI:10.1146/annurev.biochem.74.082803.133524 - dx.doi.org/10.1146/annurev.biochem.74.082803.133524. PMID 15952903 - www.ncbi.nlm.nih.gov/pubmed/15952903?dopt=Abstract.
- Ohlrogge J, Jaworski J (1997). «Regulation of fatty acid synthesis». Annu Rev Plant Physiol Plant Mol Biol 48: 109–136. DOI:10.1146/annurev.arplant.48.1.109 - dx.doi.org/10.1146/annurev.arplant.48.1.109. PMID 15012259 - www.ncbi.nlm.nih.gov/pubmed/15012259?dopt=Abstract.
- Dubey V, Bhalla R, Luthra R (2003). «An overview of the non-mevalonate pathway for terpenoid biosynthesis in plants - www.ias.ac.in/jbiosci/sep2003/637.pdf» (PDF). J Biosci 28 (5): 637–46. DOI:10.1007/BF02703339 - dx.doi.org/10.1007/BF02703339. PMID 14517367 - www.ncbi.nlm.nih.gov/pubmed/14517367?dopt=Abstract.
- ↑ 12Kuzuyama T, Seto H (2003). «Diversity of the biosynthesis of the isoprene units». Nat Prod Rep 20 (2): 171–83. DOI:10.1039/b109860h - dx.doi.org/10.1039/b109860h. PMID 12735695 - www.ncbi.nlm.nih.gov/pubmed/12735695?dopt=Abstract.
- Grochowski L, Xu H, White R (May 2006). «Methanocaldococcus jannaschii uses a modified mevalonate pathway for biosynthesis of isopentenyl diphosphate - jb.asm.org/cgi/pmidlookup?view=long&pmid=16621811». J Bacteriol 188 (9): 3192–8. DOI:10.1128/JB.188.9.3192-3198.2006 - dx.doi.org/10.1128/JB.188.9.3192-3198.2006. PMID 16621811 - www.ncbi.nlm.nih.gov/pubmed/16621811?dopt=Abstract.
- Lichtenthaler H (1999). «The 1-Ddeoxy-D-xylulose-5-phosphate pathway of isoprenoid biosynthesis in plants». Annu Rev Plant Physiol Plant Mol Biol 50: 47–65. DOI:10.1146/annurev.arplant.50.1.47 - dx.doi.org/10.1146/annurev.arplant.50.1.47. PMID 15012203 - www.ncbi.nlm.nih.gov/pubmed/15012203?dopt=Abstract.
- ↑ 12Schroepfer G (1981). «Sterol biosynthesis». Annu Rev Biochem 50: 585–621. DOI:10.1146/annurev.bi.50.070181.003101 - dx.doi.org/10.1146/annurev.bi.50.070181.003101. PMID 7023367 - www.ncbi.nlm.nih.gov/pubmed/7023367?dopt=Abstract.
- Lees N, Skaggs B, Kirsch D, Bard M (1995). «Cloning of the late genes in the ergosterol biosynthetic pathway of Saccharomyces cerevisiae—a review». Lipids 30 (3): 221–6. DOI:10.1007/BF02537824 - dx.doi.org/10.1007/BF02537824. PMID 7791529 - www.ncbi.nlm.nih.gov/pubmed/7791529?dopt=Abstract.
- Guyton Arthur C. Textbook of Medical Physiology. — Philadelphia: Elsevier, 2006. — P. 855–6. — ISBN 0-7216-0240-1
- Ibba M, Söll D (2001). «The renaissance of aminoacyl-tRNA synthesis - www.molcells.org/home/journal/include/downloadPdf.asp?articleuid={A158E3B4-2423-4806-9A30-4B93CDA76DA0}». EMBO Rep 2 (5): 382–7. PMID 11375928 - www.ncbi.nlm.nih.gov/pubmed/11375928?dopt=Abstract.
- Lengyel P, Söll D (1969). «Mechanism of protein biosynthesis». Bacteriol Rev 33 (2): 264–301. PMID 4896351 - www.ncbi.nlm.nih.gov/pubmed/4896351?dopt=Abstract.
- ↑ 12Rudolph F (1994). «The biochemistry and physiology of nucleotides». J Nutr 124 (1 Suppl): 124S–127S. PMID 8283301 - www.ncbi.nlm.nih.gov/pubmed/8283301?dopt=Abstract.
- Zrenner R, Stitt M, Sonnewald U, Boldt R (2006). «Pyrimidine and purine biosynthesis and degradation in plants». Annu Rev Plant Biol 57: 805–36. DOI:10.1146/annurev.arplant.57.032905.105421 - dx.doi.org/10.1146/annurev.arplant.57.032905.105421. PMID 16669783 - www.ncbi.nlm.nih.gov/pubmed/16669783?dopt=Abstract.
- Stasolla C, Katahira R, Thorpe T, Ashihara H (2003). «Purine and pyrimidine nucleotide metabolism in higher plants». J Plant Physiol 160 (11): 1271–95. DOI:10.1078/0176-1617-01169 - dx.doi.org/10.1078/0176-1617-01169. PMID 14658380 - www.ncbi.nlm.nih.gov/pubmed/14658380?dopt=Abstract.
- Smith J (1995). «Enzymes of nucleotide synthesis». Curr Opin Struct Biol 5 (6): 752–7. DOI:10.1016/0959-440X(95)80007-7 - dx.doi.org/10.1016/0959-440X(95)80007-7. PMID 8749362 - www.ncbi.nlm.nih.gov/pubmed/8749362?dopt=Abstract.
- Testa B, Krämer S (2006). «The biochemistry of drug metabolism—an introduction: part 1. Principles and overview». Chem Biodivers 3 (10): 1053–101. DOI:10.1002/cbdv.200690111 - dx.doi.org/10.1002/cbdv.200690111. PMID 17193224 - www.ncbi.nlm.nih.gov/pubmed/17193224?dopt=Abstract.
- Danielson P (2002). «The cytochrome P450 superfamily: biochemistry, evolution and drug metabolism in humans». Curr Drug Metab 3 (6): 561–97. DOI:10.2174/1389200023337054 - dx.doi.org/10.2174/1389200023337054. PMID 12369887 - www.ncbi.nlm.nih.gov/pubmed/12369887?dopt=Abstract.
- King C, Rios G, Green M, Tephly T (2000). «UDP-glucuronosyltransferases». Curr Drug Metab 1 (2): 143–61. DOI:10.2174/1389200003339171 - dx.doi.org/10.2174/1389200003339171. PMID 11465080 - www.ncbi.nlm.nih.gov/pubmed/11465080?dopt=Abstract.
- Sheehan D, Meade G, Foley V, Dowd C (November 2001). «Structure, function and evolution of glutathione transferases: implications for classification of non-mammalian members of an ancient enzyme superfamily - www.biochemj.org/bj/360/0001/bj3600001.htm». Biochem J 360 (Pt 1): 1–16. DOI:10.1042/0264-6021:3600001 - dx.doi.org/10.1042/0264-6021:3600001. PMID 11695986 - www.ncbi.nlm.nih.gov/pubmed/11695986?dopt=Abstract.
- Galvão T, Mohn W, de Lorenzo V (2005). «Exploring the microbial biodegradation and biotransformation gene pool». Trends Biotechnol 23 (10): 497–506. DOI:10.1016/j.tibtech.2005.08.002 - dx.doi.org/10.1016/j.tibtech.2005.08.002. PMID 16125262 - www.ncbi.nlm.nih.gov/pubmed/16125262?dopt=Abstract.
- Janssen D, Dinkla I, Poelarends G, Terpstra P (2005). «Bacterial degradation of xenobiotic compounds: evolution and distribution of novel enzyme activities». Environ Microbiol 7 (12): 1868–82. DOI:10.1111/j.1462-2920.2005.00966.x - dx.doi.org/10.1111/j.1462-2920.2005.00966.x. PMID 16309386 - www.ncbi.nlm.nih.gov/pubmed/16309386?dopt=Abstract.
- Davies K (1995). «Oxidative stress: the paradox of aerobic life». Biochem Soc Symp 61: 1–31. PMID 8660387 - www.ncbi.nlm.nih.gov/pubmed/8660387?dopt=Abstract.
- Tu B, Weissman J (2004). «Oxidative protein folding in eukaryotes: mechanisms and consequences - www.jcb.org/cgi/content/full/164/3/341». J Cell Biol 164 (3): 341–6. DOI:10.1083/jcb.200311055 - dx.doi.org/10.1083/jcb.200311055. PMID 14757749 - www.ncbi.nlm.nih.gov/pubmed/14757749?dopt=Abstract.
- Sies H (1997). «Oxidative stress: oxidants and antioxidants - ep.physoc.org/cgi/reprint/82/2/291.pdf» (PDF). Exp Physiol 82 (2): 291–5. PMID 9129943 - www.ncbi.nlm.nih.gov/pubmed/9129943?dopt=Abstract.
- Vertuani S, Angusti A, Manfredini S (2004). «The antioxidants and pro-antioxidants network: an overview». Curr Pharm Des 10 (14): 1677–94. DOI:10.2174/1381612043384655 - dx.doi.org/10.2174/1381612043384655. PMID 15134565 - www.ncbi.nlm.nih.gov/pubmed/15134565?dopt=Abstract.
- von Stockar U, Liu J (1999). «Does microbial life always feed on negative entropy? Thermodynamic analysis of microbial growth». Biochim Biophys Acta 1412 (3): 191–211. DOI:10.1016/S0005-2728(99)00065-1 - dx.doi.org/10.1016/S0005-2728(99)00065-1. PMID 10482783 - www.ncbi.nlm.nih.gov/pubmed/10482783?dopt=Abstract.
- Demirel Y, Sandler S (2002). «Thermodynamics and bioenergetics». Biophys Chem 97 (2–3): 87–111. DOI:10.1016/S0301-4622(02)00069-8 - dx.doi.org/10.1016/S0301-4622(02)00069-8. PMID 12050002 - www.ncbi.nlm.nih.gov/pubmed/12050002?dopt=Abstract.
- Albert R (2005). «Scale-free networks in cell biology - jcs.biologists.org/cgi/content/full/118/21/4947». J Cell Sci 118 (Pt 21): 4947–57. DOI:10.1242/jcs.02714 - dx.doi.org/10.1242/jcs.02714. PMID 16254242 - www.ncbi.nlm.nih.gov/pubmed/16254242?dopt=Abstract.
- Brand M (1997). «Regulation analysis of energy metabolism - jeb.biologists.org/cgi/reprint/200/2/193». J Exp Biol 200 (Pt 2): 193–202. PMID 9050227 - www.ncbi.nlm.nih.gov/pubmed/9050227?dopt=Abstract.
- Soyer O, Salathé M, Bonhoeffer S (2006). «Signal transduction networks: topology, response and biochemical processes». J Theor Biol 238 (2): 416–25. DOI:10.1016/j.jtbi.2005.05.030 - dx.doi.org/10.1016/j.jtbi.2005.05.030. PMID 16045939 - www.ncbi.nlm.nih.gov/pubmed/16045939?dopt=Abstract.
- Westerhoff H, Groen A, Wanders R (1984). «Modern theories of metabolic control and their applications (review)». Biosci Rep 4 (1): 1–22. DOI:10.1007/BF01120819 - dx.doi.org/10.1007/BF01120819. PMID 6365197 - www.ncbi.nlm.nih.gov/pubmed/6365197?dopt=Abstract.
- Salter M, Knowles R, Pogson C (1994). «Metabolic control». Essays Biochem 28: 1–12. PMID 7925313 - www.ncbi.nlm.nih.gov/pubmed/7925313?dopt=Abstract.
- Fell D, Thomas S (1995). «Physiological control of metabolic flux: the requirement for multisite modulation». Biochem J 311 (Pt 1): 35–9. PMID 7575476 - www.ncbi.nlm.nih.gov/pubmed/7575476?dopt=Abstract.
- Hendrickson W (2005). «Transduction of biochemical signals across cell membranes». Q Rev Biophys 38 (4): 321–30. DOI:10.1017/S0033583506004136 - dx.doi.org/10.1017/S0033583506004136. PMID 16600054 - www.ncbi.nlm.nih.gov/pubmed/16600054?dopt=Abstract.
- Cohen P (2000). «The regulation of protein function by multisite phosphorylation—a 25 year update». Trends Biochem Sci 25 (12): 596–601. DOI:10.1016/S0968-0004(00)01712-6 - dx.doi.org/10.1016/S0968-0004(00)01712-6. PMID 11116185 - www.ncbi.nlm.nih.gov/pubmed/11116185?dopt=Abstract.
- Lienhard G, Slot J, James D, Mueckler M (1992). «How cells absorb glucose». Sci Am 266 (1): 86–91. DOI:10.1038/scientificamerican0192-86 - dx.doi.org/10.1038/scientificamerican0192-86. PMID 1734513 - www.ncbi.nlm.nih.gov/pubmed/1734513?dopt=Abstract.
- Roach P (2002). «Glycogen and its metabolism». Curr Mol Med 2 (2): 101–20. DOI:10.2174/1566524024605761 - dx.doi.org/10.2174/1566524024605761. PMID 11949930 - www.ncbi.nlm.nih.gov/pubmed/11949930?dopt=Abstract.
- Newgard C, Brady M, O'Doherty R, Saltiel A (2000). «Organizing glucose disposal: emerging roles of the glycogen targeting subunits of protein phosphatase-1 - diabetes.diabetesjournals.org/cgi/reprint/49/12/1967.pdf» (PDF). Diabetes 49 (12): 1967–77. DOI:10.2337/diabetes.49.12.1967 - dx.doi.org/10.2337/diabetes.49.12.1967. PMID 11117996 - www.ncbi.nlm.nih.gov/pubmed/11117996?dopt=Abstract.
- Romano A, Conway T (1996). «Evolution of carbohydrate metabolic pathways». Res Microbiol 147 (6–7): 448–55. DOI:10.1016/0923-2508(96)83998-2 - dx.doi.org/10.1016/0923-2508(96)83998-2. PMID 9084754 - www.ncbi.nlm.nih.gov/pubmed/9084754?dopt=Abstract.
- Koch A (1998). «How did bacteria come to be?». Adv Microb Physiol 40: 353–99. DOI:10.1016/S0065-2911(08)60135-6 - dx.doi.org/10.1016/S0065-2911(08)60135-6. PMID 9889982 - www.ncbi.nlm.nih.gov/pubmed/9889982?dopt=Abstract.
- Ouzounis C, Kyrpides N (1996). «The emergence of major cellular processes in evolution». FEBS Lett 390 (2): 119–23. DOI:10.1016/0014-5793(96)00631-X - dx.doi.org/10.1016/0014-5793(96)00631-X. PMID 8706840 - www.ncbi.nlm.nih.gov/pubmed/8706840?dopt=Abstract.
- Caetano-Anolles G, Kim HS, Mittenthal JE (2007). «The origin of modern metabolic networks inferred from phylogenomic analysis of protein architecture». Proc Natl Acad Sci USA 104 (22): 9358–63. DOI:10.1073/pnas.0701214104 - dx.doi.org/10.1073/pnas.0701214104. PMID 17517598 - www.ncbi.nlm.nih.gov/pubmed/17517598?dopt=Abstract.
- Schmidt S, Sunyaev S, Bork P, Dandekar T (2003). «Metabolites: a helping hand for pathway evolution?». Trends Biochem Sci 28 (6): 336–41. DOI:10.1016/S0968-0004(03)00114-2 - dx.doi.org/10.1016/S0968-0004(03)00114-2. PMID 12826406 - www.ncbi.nlm.nih.gov/pubmed/12826406?dopt=Abstract.
- Light S, Kraulis P (2004). «Network analysis of metabolic enzyme evolution in Escherichia coli». BMC Bioinformatics 5: 15. DOI:10.1186/1471-2105-5-15 - dx.doi.org/10.1186/1471-2105-5-15. PMID 15113413 - www.ncbi.nlm.nih.gov/pubmed/15113413?dopt=Abstract. Alves R, Chaleil R, Sternberg M (2002). «Evolution of enzymes in metabolism: a network perspective». J Mol Biol 320 (4): 751–70. DOI:10.1016/S0022-2836(02)00546-6 - dx.doi.org/10.1016/S0022-2836(02)00546-6. PMID 12095253 - www.ncbi.nlm.nih.gov/pubmed/12095253?dopt=Abstract.
- Kim HS, Mittenthal JE, Caetano-Anolles G (2006). «MANET: tracing evolution of protein architecture in metabolic networks». BMC Bioinformatics 19 (7): 351. DOI:10.1186/1471-2105-7-351 - dx.doi.org/10.1186/1471-2105-7-351. PMID 16854231 - www.ncbi.nlm.nih.gov/pubmed/16854231?dopt=Abstract.
- Teichmann SA, Rison SC, Thornton JM, Riley M, Gough J, Chothia C (2001). «Small-molecule metabolsim: an enzyme mosaic». Trends Biotechnol 19 (12): 482–6. DOI:10.1016/S0167-7799(01)01813-3 - dx.doi.org/10.1016/S0167-7799(01)01813-3. PMID 11711174 - www.ncbi.nlm.nih.gov/pubmed/11711174?dopt=Abstract.
- Spirin V, Gelfand M, Mironov A, Mirny L (June 2006). «A metabolic network in the evolutionary context: multiscale structure and modularity - www.pnas.org/cgi/pmidlookup?view=long&pmid=16731630». Proc Natl Acad Sci USA 103 (23): 8774–9. DOI:10.1073/pnas.0510258103 - dx.doi.org/10.1073/pnas.0510258103. PMID 16731630 - www.ncbi.nlm.nih.gov/pubmed/16731630?dopt=Abstract.
- Lawrence J (2005). «Common themes in the genome strategies of pathogens». Curr Opin Genet Dev 15 (6): 584–8. DOI:10.1016/j.gde.2005.09.007 - dx.doi.org/10.1016/j.gde.2005.09.007. PMID 16188434 - www.ncbi.nlm.nih.gov/pubmed/16188434?dopt=Abstract. Wernegreen J (2005). «For better or worse: genomic consequences of intracellular mutualism and parasitism». Curr Opin Genet Dev 15 (6): 572–83. DOI:10.1016/j.gde.2005.09.013 - dx.doi.org/10.1016/j.gde.2005.09.013. PMID 16230003 - www.ncbi.nlm.nih.gov/pubmed/16230003?dopt=Abstract.
- Pál C, Papp B, Lercher M, Csermely P, Oliver S, Hurst L (2006). «Chance and necessity in the evolution of minimal metabolic networks». Nature 440 (7084): 667–70. DOI:10.1038/nature04568 - dx.doi.org/10.1038/nature04568. PMID 16572170 - www.ncbi.nlm.nih.gov/pubmed/16572170?dopt=Abstract.
- Rennie M (1999). «An introduction to the use of tracers in nutrition and metabolism». Proc Nutr Soc 58 (4): 935–44. DOI:10.1017/S002966519900124X - dx.doi.org/10.1017/S002966519900124X. PMID 10817161 - www.ncbi.nlm.nih.gov/pubmed/10817161?dopt=Abstract.
- Phair R (1997). «Development of kinetic models in the nonlinear world of molecular cell biology». Metabolism 46 (12): 1489–95. DOI:10.1016/S0026-0495(97)90154-2 - dx.doi.org/10.1016/S0026-0495(97)90154-2. PMID 9439549 - www.ncbi.nlm.nih.gov/pubmed/9439549?dopt=Abstract.
- Sterck L, Rombauts S, Vandepoele K, Rouzé P, Van de Peer Y (2007). «How many genes are there in plants (... and why are they there)?». Curr Opin Plant Biol 10 (2): 199–203. DOI:10.1016/j.pbi.2007.01.004 - dx.doi.org/10.1016/j.pbi.2007.01.004. PMID 17289424 - www.ncbi.nlm.nih.gov/pubmed/17289424?dopt=Abstract.
- Borodina I, Nielsen J (2005). «From genomes to in silico cells via metabolic networks». Curr Opin Biotechnol 16 (3): 350–5. DOI:10.1016/j.copbio.2005.04.008 - dx.doi.org/10.1016/j.copbio.2005.04.008. PMID 15961036 - www.ncbi.nlm.nih.gov/pubmed/15961036?dopt=Abstract.
- Gianchandani E, Brautigan D, Papin J (2006). «Systems analyses characterize integrated functions of biochemical networks». Trends Biochem Sci 31 (5): 284–91. DOI:10.1016/j.tibs.2006.03.007 - dx.doi.org/10.1016/j.tibs.2006.03.007. PMID 16616498 - www.ncbi.nlm.nih.gov/pubmed/16616498?dopt=Abstract.
- Duarte NC, Becker SA, Jamshidi N, et al. (February 2007). «Global reconstruction of the human metabolic network based on genomic and bibliomic data - www.pnas.org/cgi/pmidlookup?view=long&pmid=17267599». Proc. Natl. Acad. Sci. U.S.A. 104 (6): 1777–82. DOI:10.1073/pnas.0610772104 - dx.doi.org/10.1073/pnas.0610772104. PMID 17267599 - www.ncbi.nlm.nih.gov/pubmed/17267599?dopt=Abstract.
- Goh KI, Cusick ME, Valle D, Childs B, Vidal M, Barabási AL (May 2007). «The human disease network». Proc. Natl. Acad. Sci. U.S.A. 104 (21): 8685–90. DOI:10.1073/pnas.0701361104 - dx.doi.org/10.1073/pnas.0701361104. PMID 17502601 - www.ncbi.nlm.nih.gov/pubmed/17502601?dopt=Abstract.
- Lee DS, Park J, Kay KA, Christakis NA, Oltvai ZN, Barabási AL (July 2008). «The implications of human metabolic network topology for disease comorbidity - www.pnas.org/lookup/pmid?view=long&pmid=18599447». Proc. Natl. Acad. Sci. U.S.A. 105 (29): 9880–9885. DOI:10.1073/pnas.0802208105 - dx.doi.org/10.1073/pnas.0802208105. PMID 18599447 - www.ncbi.nlm.nih.gov/pubmed/18599447?dopt=Abstract.
- Csete M, Doyle J (2004). «Bow ties, metabolism and disease». Trends Biotechnol. 22 (9): 446–50. DOI:10.1016/j.tibtech.2004.07.007 - dx.doi.org/10.1016/j.tibtech.2004.07.007. PMID 5249808 - www.ncbi.nlm.nih.gov/pubmed/5249808?dopt=Abstract.
- Ma HW, Zeng AP (2003). «The connectivity structure, giant strong component and centrality of metabolic networks». Bioinformatics 19 (11): 1423–30. DOI:10.1093/bioinformatics/btg177 - dx.doi.org/10.1093/bioinformatics/btg177. PMID 12874056 - www.ncbi.nlm.nih.gov/pubmed/12874056?dopt=Abstract.
- Zhao J, Yu H, Luo JH, Cao ZW, Li YX (2006). «Hierarchical modularity of nested bow-ties in metabolic networks». BMC Bioinformatics 7: 386. DOI:10.1186/1471-2105-7-386 - dx.doi.org/10.1186/1471-2105-7-386. PMID 16916470 - www.ncbi.nlm.nih.gov/pubmed/16916470?dopt=Abstract.
- Thykaer J, Nielsen J (2003). «Metabolic engineering of beta-lactam production». Metab Eng 5 (1): 56–69. DOI:10.1016/S1096-7176(03)00003-X - dx.doi.org/10.1016/S1096-7176(03)00003-X. PMID 12749845 - www.ncbi.nlm.nih.gov/pubmed/12749845?dopt=Abstract. González-Pajuelo M, Meynial-Salles I, Mendes F, Andrade J, Vasconcelos I, Soucaille P (2005). «Metabolic engineering of Clostridium acetobutylicum for the industrial production of 1,3-propanediol from glycerol». Metab Eng 7 (5–6): 329–36. DOI:10.1016/j.ymben.2005.06.001 - dx.doi.org/10.1016/j.ymben.2005.06.001. PMID 16095939 - www.ncbi.nlm.nih.gov/pubmed/16095939?dopt=Abstract. Krämer M, Bongaerts J, Bovenberg R, Kremer S, Müller U, Orf S, Wubbolts M, Raeven L (2003). «Metabolic engineering for microbial production of shikimic acid». Metab Eng 5 (4): 277–83. DOI:10.1016/j.ymben.2003.09.001 - dx.doi.org/10.1016/j.ymben.2003.09.001. PMID 14642355 - www.ncbi.nlm.nih.gov/pubmed/14642355?dopt=Abstract.
- Koffas M, Roberge C, Lee K, Stephanopoulos G (1999). «Metabolic engineering». Annu Rev Biomed Eng 1: 535–57. DOI:10.1146/annurev.bioeng.1.1.535 - dx.doi.org/10.1146/annurev.bioeng.1.1.535. PMID 11701499 - www.ncbi.nlm.nih.gov/pubmed/11701499?dopt=Abstract.
- Dr. Abu Shadi Al-Roubi (1982), «Ibn Al-Nafis as a philosopher», Symposium on Ibn al Nafis, Second International Conference on Islamic Medicine: Islamic Medical Organization, Kuwait (cf. Ibnul-Nafees As a Philosopher - www.islamset.com/isc/nafis/drroubi.html, Encyclopedia of Islamic World [1] - web.archive.org/web/20080206072116/http://www.islamset.com/isc/nafis/drroubi.html).
- Eknoyan G (1999). «Santorio Sanctorius (1561–1636) - founding father of metabolic balance studies». Am J Nephrol 19 (2): 226–33. DOI:10.1159/000013455 - dx.doi.org/10.1159/000013455. PMID 10213823 - www.ncbi.nlm.nih.gov/pubmed/10213823?dopt=Abstract.
- Williams, H. S. (1904) A History of Science: in Five Volumes. Volume IV: Modern Development of the Chemical and Biological Sciences - etext.lib.virginia.edu/toc/modeng/public/Wil4Sci.html Harper and Brothers (New York) Retrieved on 2007-03-26
- Dubos J. (1951). «Louis Pasteur: Free Lance of Science, Gollancz. Quoted in Manchester K. L. (1995) Louis Pasteur (1822–1895)—chance and the prepared mind». Trends Biotechnol 13 (12): 511–515. DOI:10.1016/S0167-7799(00)89014-9 - dx.doi.org/10.1016/S0167-7799(00)89014-9. PMID 8595136 - www.ncbi.nlm.nih.gov/pubmed/8595136?dopt=Abstract.
- Kinne-Saffran E, Kinne R (1999). «Vitalism and synthesis of urea. From Friedrich Wöhler to Hans A. Krebs». Am J Nephrol 19 (2): 290–4. DOI:10.1159/000013463 - dx.doi.org/10.1159/000013463. PMID 10213830 - www.ncbi.nlm.nih.gov/pubmed/10213830?dopt=Abstract.
- Eduard Buchner’s 1907 Nobel lecture - nobelprize.org/nobel_prizes/chemistry/laureates/1907/buchner-lecture.html at http://nobelprize.org - nobelprize.org Accessed 2007-03-20
- Kornberg H (2000). «Krebs and his trinity of cycles». Nat Rev Mol Cell Biol 1 (3): 225–8. DOI:10.1038/35043073 - dx.doi.org/10.1038/35043073. PMID 11252898 - www.ncbi.nlm.nih.gov/pubmed/11252898?dopt=Abstract.
- Krebs HA, Henseleit K (1932). «Untersuchungen über die Harnstoffbildung im tierkorper». Z. Physiol. Chem. 210: 33–66.Krebs H, Johnson W (April 1937). «Metabolism of ketonic acids in animal tissues». Biochem J 31 (4): 645–60. PMID 16746382 - www.ncbi.nlm.nih.gov/pubmed/16746382?dopt=Abstract.
Категории: Физиология, Метаболизм.
Текст доступен по лицензии Creative Commons Attribution-ShareAlike.wreferat.baza-referat.ru
Реферат - Метаболизм - Биология
Кировская гимназия имени героя Советского Союза Султана Баймагамбетова
Экзаменционный реферат по биологиитема: ”Метаболизм”
Выполнил: Сысоев М.Ю.
Преподаватель: Савченко И.В.
Кировск 2010
Содержание
ВведениеОсновная часть
Определение метаболизма Анаболизм и катаболизм Энергетический обмен в клетке
Протеины Определения протеинов Синтез протеинов в живых организмах Классификация, структура и функции
Синтез липидов в живых организмах Компоненты липидов Образование липидов Некоторые виды липидов
Синтез углеводов в живых организмах Классификация углеводов Синтез Использование в живых организмах
Заключительная часть
Приложения
Список литературы
Введение
Мы довольно много знаем о процессах, протекающих в клетке на молекулярном уровне. Достаточно упомянуть о том, что была расшифрована структура генетического материала -ДНК, были получены интересные и принципиально новые данные об активности генов в бактериальной клетке и клетках высших организмов.
На современном этапе развития науки следует больше внимания уделять организму как целому, связывать протекающие процессы с его функционированием, помнить о том, что наша конечная цель состоит в понимании структуры и функция организма в целом на основе знаний об отдельных его системах. Однако достичь этой цели невозможно без знания реакциях метаболизма, ведь, не зная что и как получается в организме, нельзя предположить что может получится в будущем.
В процессе написания сего реферата, я хочу узнать подробней о том что такое метаболизм, как проиходят реакции метаболизма в организме и какие ученые занимались изучением сих реакций.
Метаболизм
Анаболизм
Любая живая клетка, осуществляя многообразные процессы синтеза и распада веществ, подобна сложнейшему химическому комбинату. Для нормального протекания этиъ химических процессов необходим постоянный обмен веществами между клеткой и окружающей средой, а также постоянное превращение энергии в клетке. Получаемые извне белки, жиры, углеводы, витамины, микроэлементы расходуются клетками на синтез необходимых им соединений, построение клеточных структур.
Из поступающих в клетку веществ, под воздействием ферментов образуются новые вещества, необходимые для замены израсходованных веществ и построения органоидов. Вест набор реакций биосинтеза веществ в клетке называется ассимиляцией или пластического обмена. Также эти реакции называют реакциями анаболизма. Эти реакции происходят с поглощением энергии и потому называются эндотермическими.
КатаболизмОчевидно, что синтез каких-либо веществ невозможен без затрат энергии. Особенно интенсивно реакции анаболизма происходят в растущей, развивающейся клетке. Важнейшими из таких реакций являются синтез белка и фотосинтез. Как же клетка получает энергию для реакций биосинтеза? Наряду с процессами синтеза новых веществ, в клетках происходит постоянный распад запасенных при ассимиляции сложных органических веществ. При участии ферментов эти вещества распадаются до более простых соединений: при этом высвобождается энергия. Чаще всего эта энергия запасается в виде аденозинтри-фосфата (АТФ). Далее эта энергия используется для различных нужд клетки, в том числе и для реакций анаболизма. Совокупность этих реакций называется диссимиляцией или энергетический обмен. Также они называются реакциями катаболизма. Эти реакции происходят с выделением энергии и потому называются экзотермическими.
Разложение углеводовРазложение углеводов начинается уже в ротовой полости. Когда углеводы попадают туда, начинает выделяться слюна, в которой помимо воды и солей различных кислот, содержит ферменты, которые позволяют начать расщепление простых углеводов, таких как крахмал и мальтозу. Расщепление происходит до появления, как правило, глюкозы. Далее, появившаяся глюкоза всасывается, а остальные не поддающиеся расщеплению в ротовой полости углеводы идут по ЖКТ дальше и попадают в желудок, а затем и в кишечный тракт.
Разложение белковПопадая в наш организм, белки сначала оказываются в ротовой полости, где ферменты слюны ничего с ними поделать не могут, и затем попадают в желудок. В желудке, выделяется желудочный сок, который содержит самые различные ферменты, соляную кислоту и некоторые другие вещества. Благодаря богатому набору ферментов, большинство белков перевариваются там. Некоторые аминокислоты, полученные в результате пищеварения, используются для синтеза белков организма, а остальные превращаются в глюкозу.
Ферменты которые расщепляют белки называются протеазами.
Разложение липидовКак и белки, липиды начинают разлагаться только в желудке, да и то
-далеко не все липиды, а только простейшие. Дальшейшее разложение липидов происходит в кишечнике. Попадая в кишечник, пища, смоченная слюной и желудочным соком, подвергается действию кишечного сока, желчи, сока поджелудочной железы, здесь же происходит всасывание продуктов переваривания в кровеносные и лимфатические капилляры.
МетаболизмАнаболизм и катаболизм -противоположные процессы: в одном случае вещества образуются, в другом -разрушаются. Но они тесно связаны и друг без друга невозможны. Ведь если в клетке не будет синтеза веществ, то нечему будет распадаться, т.е. не будет материала для распада и получения энергии. Таким образом метаболизм -совокупность реакций анаболизма и катаболизма, т.е. обмен веществ.
Энергетический обмен в клетке
АТФ обеспечивает энергией все функции клетки: механическую работу, биосинтез, деление и т.д. В среднем, содержание АТФ в клетке составляет около 0,05% ее массы, но в тех клетках, где затраты АТФ велики (например, в клетках печени, поперечно-полосатых мышц), ее содержание может доходить до 0,5%. Синтез АТФ в клетках происходит главным образом в митохондриях. Этот процесс можно условно разделить на три этапа: 1) Подготовительный: при этом этапе крупные пищевые полимерные молекулы распадаются на более мелкие фрагменты: жиры распадаются на жирные кислоты и глицерин, белки до аминокислот и далее. В ходе этих превращений энергии выделяется мало и она рассеивается в виде тепла, т.е. АТФ не образуется. 2) Неполное бескислородное расщепление: на этом этапе вещества, образовавшиеся во время первого этапа, разлагаются под действием ферментов и в отсутствие кислорода. Ниже я постараюсь привести пример на основе гликолиза. Гликолиз происходит в животных клетках и у некоторых микроорганизмов. Суммарно, процесс можно представить так:
C₆H₁₂O₆+2H₃PO₄+2АДФ = 2C₃H₆O₃+2H₂O+2АТФ Где можно наглядно увидеть, что при гликолизе из одной молекулы глюкозы образуется 2 молекулы пировиноградной кислоты, которая затем во многих клетках превращается в молочную кислоту. Причем, образовавшейся энергии хватает для образования двух молекул АТФ. Несмотря на кажущуюся простоту, гликолиз -очень сложный и многоступенчатый процесс, насчитывающий более десяти ступеней, катализируемых различными ферментами. В результате, только 40% энергии запасается в виде АТФ, остальная часть энергии рассеивается в виде тепла. Гликолиз происходит в цитоплазме клеток. 3) Полное кислородное расщепление или клеточное дыхание: на этом этапе вещества, образовавшиеся во время второго этапа, распадаются до конечных CO₂и H₂O. Этот этап упрощенно можно представить в виде следующего уравнения:
2C₃H₆O₃+ 6O₂+ 36H₃PO₄+ 36АДФ→CO₂+ 42H₂O +36АТФ Таким образом, окисление двух молекул молочной кислоты, приводит к выделению большого количества энергии, достаточной для образования 36 молекул АТФ. Клеточное дыхание происходит на кристах митохондрий. КПД этого процесса выше чем у гликолиза и составляет примерно 55%. В результате полного расщепления одной молекулы глюкозы образуется 38 молекул АТФ. Для получения энергии в клетках используются не только сахариды, но также липиды и, в некоторых случаях, белки. Однако, чаще всего используются сахара.
Протеины
Определение протеиновПротеины (белки) -это сложные органические соединения, состоящие из углерода, кислорода, водорода и азота. В некоторых белках также может содержатся сера и фосфор.Часть белков образует комплексы с другими молекулами, содержащими фосфор, железо, цинк и медь.
Молекулы белков — (цепи построенные из аминокислот) -очень велики; это макромолекулы, молекулярная масса которых колеблется от нескольких тысяч, до нескольких миллионов. В природных белках встречается двадцать различных аминокислот. Однако, потенциально разнообразие белков безгранично, поскольку каждому белку свойственна своя особая аминокислотная последовательность, генетически контролируемая, то есть -закодированная в ДНК клетки, вырабатывающей данный белок. Белков в данных клетках больше, чем каких-бы то ни было других органических соединений: на их долю приходится больше 50% общей сухой массы клеток.
Они -важный элемент пищи животных и могут превращаться в жиры, углеводы и другие белки. А их большое разнообразие позволяет им выполнять в живом организме множество различных функций, такие как:
1) Пластическая функция 2) Метаболическая функция 3) Энергетическая 4) Транспортная 5) Регуляторная 6) Сигнальная 7) Защитная
Классификация белковСложность строения белковых молекул и чрезвычайное разнообразие разнообразие их функций крайне затрудняют создание единой четкой классификации белков на какой-либо основе. Я постараюсь изобразить некоторые классификации белков в виде таблиц.
Белки
/> Простые СложныеСостоят только изСостоят из глобулярныхаминокислотбелков и небелкового материалаПростые белки
Название Свойства Пример АльбуминыНейтральные, растворимы в воде и разбавленных соленых растворахЯичный альбумин Сывороточный альбумин в кровиГлобулиныНейтральные. Нерастворимы в воде. Растворимы в разбавленных соленых растворахСодержащиеся в крови антитела ФибринГистоныОсновные Растворимы в воде Нерастворимы в разбавленном водном растворе аммиакаСвязаны с нуклеиновыми кислотами в нуклеопротеидах клеткиСклеропротеиныНерастворимы в воде и в большей части других растворителейКератин волос, кожи, коллаген сухожилий, эластин связокСложные белки
Название Простетическая группа Пример ФосфопротеиныФосфорная кислотаКазеин молока Вителлин яичного желткаГликопротеиныУглеводПлазма крови Муцин (компонент слюны)НуклеопротеиныНуклеиновая кислотаКомпоненты вирусов Хромосомы РибосомыХромопротеиныПигментГемоглобин-гем (железосодержащий пигмент)Фитохром (пигмент растительного происхождения)Цитохром (дыхательный пигмент)ЛипопротеиныЛипидКомпонент мембран Липопротеины крови -траспортная форма липидовФлавопротеиныФАД (флавинадениндинуклеот ид)Компонент цепи переноса электронов при дыханииМеталлопротеиныМеталлНитратредуктаза -фермент, катализирующий в растениях превращение нитрата в нитритТакже белки могут классицироваться не только по сложности и составу, но и их функциям и структуре. Ниже представлены таблицы классификации белков по их структуре и по их функциям.
Классификация белков по их структуре
Класс белков Характеристика Функция ФибриллярныеНаиболее важна вторичная структура (третичная почти или совсем не выражена)Нерастворимы в воде. Отличаются большой механической прочностью. Длинные параллельные полипептидные цепи, скрепленные друг с другом поперечными сшивками, образуют длинные волокна или слоистые структуры.Выполняют в клетках и в организме структурные функции, например в составе соединительной ткани; к этой группе относятся среди других коллаген (сухожилия, межклеточное вещество костной ткани), миозин в мышцах, фиброин (шелк, паутина), кератин (волосы, рога, ногти, перья)ГлобулярныеНаиболее важна третичная структура. Полипептидные цепи свернуты в компактные глобулы. Растворимы -легко образуют коллоидные суспензии.Выполняют функции ферментов, антител (глобулины сыворотки крови определяют имуннологическую активность) и в некоторых случаях гормонов (например -инсулин)ПромежуточныеФибриллярной природы, но растворимые.Примером может служить фибриноген, превращающийся в нерастворимый фибрин при свертывании крови.Также, при описании структуры белков рассматривают обычно три разных уровня организации. Первичная структура: под ней обычно подразумевают число и последовательность аминокислот, соединенных друг с другом пептидными связями в полипептидной цепи. Вторичная структура: α-спираль, возможная благодаря множеству водородных связей, возникаемых между находящимися поблизости друг от друга СО-и NH-группами. Третичная структура: у большинства белков полипептидные цепи свернуты особым образом в компактную глобулу. Этот способ свертывания полипептидных цепей глобулярных белков называется третичной структурой
Классификация белков по их функциям
Класс белков Характеристика Локализация/Функция Структурные белкиКоллаген Склеротин α-Кератин Эластин Мукопротеины Белки оболочки вирусовКомпонент соединительной ткани Наружный скелет насекомых Кожа, перья, рога Связки Слизистые секреты КапсидФерментыТрипсин Липаза Амилаза ГлутаминсинтетазаКатализ белков, жиров, углеводов. Катализ образования глутамина из глутаминовой кислоты и аммикаГормоныИнсулин ГлюкагонРегуляция обмена глюкозыТранспортные белкиГемоглобин Гемоцианин Миоглобин Сывороточный альбуминТранспорт О₂ в крови позвоночных и некоторых беспозвоночных. Транспорт жирных кислот и липидов и т.п.Защитные белкиАнтитела Фибриноген ТромбинОбразуют защитные комплексы с инородными телами. Участие в свертывании кровиСократительные белкиМиозин АктинПодвижные нити миофибрилл саркомера Неподвижные нити миофибрилл саркомераЗапасные белкиЯичный альбумин КазеинБелок яйца Белок молокаТоксиныЗмеиный яд Дифтерийный токсинФерменты Токсин, вырабатываемый дифтерийной палочкойСинтез белковСинтез белков в клетках -это очень сложный и энергозатратный процесс, он требует очень большого количества ферментов и других специфических макромолекул, общее количество которых, близко к трёхстам. Часть из них, к тому же объединены в сложную трёхмерную структуру рибосом. Но несмотря на большую сложность, синтез протекает с чрезвычайно высокой скоростью (десятки аминокислотных остатков в секунду). Процесс может замедляться и даже останавливаться ингибиторами-антибиотиками… Он протекает как в клетках прокариотах, так и в клетках эукариотах.
Синтез белков проводится рибосомами. Рибосома состоит из двух субчастиц: большой и малой. (см. приложение1)
Каждая субчастица состоит из нескольких десятков белков, каждый из которых уже изучен, известно, каким образом каждый белок уложен в субчастицу. При исследовании белков используют метод электрофореза, то есть в электрическом поле в специальном геле или специальном носителе молекулы белков разъединяются в зависимости от их заряда и молекулярного веса, то есть под действием поля они начинают двигаться и могут отодвигаться друг от друга на разное расстояние. Другим методом разделения белков является хроматография, в результате этого метода на носителе получают пятнышки, каждый из которых соответствует отдельному белку.
Белки в рибосоме держатся на каркасе, состоящем из рибосомной РНК. Формирование рибосомы начинается с того, что рибосомная РНК сворачивается и на нее в определенном порядке начинают налипать белки. На рисунке представлена рибосомная РНК. В ней самокомплементарные участки нити РНК спариваются, образуя шпильки (вторичная структура), и затем РНК сворачивается (третичная структура РНК), образуя каркас субчастиц. (см. приложение1)
Еще один вид РНК, участвующей в синтезе белка, это транспортная РНК (тРНК). Молекулы тРНК относительно небольшие (по сравнению с рибосомной или матричной РНК). За счет спаривания комплементарных участков молекулы тРНК образуется три «стебля» с петлями на концах и один «стебель», образованный 5'-и 3'-концами молекулы тРНК. Изображение этой структуры похоже на крест или клеверный лист. «Голова» на этом листе представлена антикодонной петлей, здесь находится антикодон – те три нуклеотида, которые комплементарно взаимодействуют с кодоном в мРНК. Противоположный антикодонной петле стебель, образованный концами молекулы, называется акцепторным стеблем – сюда присоединяется соответствующая аминокислота.
Распознают подходящие друг другу тРНК и аминокислоты специальные ферменты, называемые аминоацил-тРНК синтетазами. Для каждой аминокислоты есть своя аминоацил-тРНК синтетаза.(см.
приложение2)
В рибосоме находится матричная РНК (мРНК). С кодоном (тремя нуклеотидами) мРНК комплементарно связан антикодон транспортной РНК, на которой висит остаток аминокислоты. На рисунке видна такая структура (тРНК вместе с аминокислотой, которая называется аминоцил-тРНК).(см. приложение3)
Процесс трансляции, также как и процесс транскрипции, связан с перемещением вдоль молекулы нуклеиновой кислоты, разница в том, что рибосома шагает на три нуклеотида, в то время как РНК-полимераза -на один.
Аминоцил т-РНК входит в рибосому, комплементарно связываясь с кодоном мРНК, затем происходит реакция при которой аминокислотные остатки связываются друг с другом, а т-РНК удаляется. (см. приложение4)
«Словарь» для перевода с языка нуклеотидов на язык аминокислот называется генетическим кодом. Аминокислот — 20, нуклеотидов – 4, число комбинаций из 4 по 2 = 16, а аминокислот 20, поэтому кодировка не двух, а трехбуквенная, каждая тройка называется кодоном. Каждая аминокислота кодируется тремя нуклеотидами в мРНК (которая, в свою очередь, кодируется ДНК).
В таблице на рисунке боковые столбцы кодируют левую и правую букву кодона, верхняя строка – среднюю. Например кодон AUG кодирует аминокислоту метионин. Число комбинаций из 4 по 3 = 64, то есть некоторые аминокислоты кодируются несколькими кодонами. Три кодона не кодируют никакую аминокислоту, они называются терминирующими. Когда они попадаются в мРНК, рибосома прекращает свою работу и готовая полипептидная цепь выбрасывается наружу.(см. приложение5)
Таблица генетического кода была составлена в 60-х годах. Начало положили Ниренберг и Маттеию. Они пытались производить в пробирке эксперименты на клеточных экстрактах, к которым были добавлены искусственные матрицы РНК. В то время считалось, что кодоны, состоящие из одного нуклеотида (UUU или ААА) не кодируют аминокислоты. Ниренберг и Маттеи использовали полиU-РНК (то есть состоящую только из урацилов) в качестве контроля в своих опытах, но именно в этой пробирке прошла реакция. Стало ясно, что кодон UUU кодирует аминокислоту фенилаланин. Затем была составлена таблица генетического кода.
Генетический код универсален. Он один и тот же у всех микроорганизмов. Есть небольшие отличия в генетическом коде митохондрий.
Липиды
Определение и классификация
Липиды -обширная группа жироподобных веществ, нерастворимых в воде. Большинство липидов состоит из высокомолекулярных жирных кислот и трехатомного спирта глицерина.
Содержание липидов в разных клетках сильно варьируется: от 2-3% до 50-90% в клетках семян некоторых растений и жировой ткани животных соответсвенно.
Липиды присутствуют во всех, без исключения клетках, выполняя самые различные функции. 1) Энергетическая функция. Наиболее простые и широкораспростра-
ненные липиды играют роли источников энергии.
2) Защитная функция. Благодаря низкой теплопроводности, липиды используются как теплоизоляционный материал, например в подкожном жировом слое.
3) Строительная функция. Так как липиды водонерастворимы, они используются при построении клеточных мембран. 4) Регуляторная функция. Многие гормоны (например гормоны коры надпочечников) являются производными липидов.
Классификация липидов.
В соответствии с химическим строением различают три основные группы липидов: 1) Жирные кислоты и продукты их ферментативного окисления, 2) Глицеролипиды (содержат в молекуле остаток глицерина), 3) Липиды, не содержащие в молекуле остаток глицерина (за
исключением соеденений, входящих в первую группу). По другой классификации (она приведена на схеме), липиды подразделяют на нейтральные липиды, фосфолипиды и гликолипиды.
Компоненты липидов
Жирные кислоты содержат в своей молекуле кислотную группу -COOH( карбоксильная группа). Жирными их называют потому, что некоторые высокомолекулярные члены этой группы входят в состав жиров. Общая формула жирных кислот имеет вид R⋄COOH, где R -атом водорода, или алкильный радикал, типа -CH₃, -C₂H₅ и далее.В липидах, радикал R обычно представлен длинной цепью углеродных атомов. Большая часть жирных кислот содержит четное количество атомов углерода, от 14 до 22 (чаще всего 16 или 18). В приложении по липидам даны рисунки, на которых можно увидеть строение двух наиболее распространенных жирных кислот. При просмотре рисунков, советую обратить внимание на длинные “хвосты” из атомов углерода и водорода. Углеводородные хвосты молекул определяют многие свойства липидов, в том чиле и нерастворимость в воде.
Иногда в жирных кислотах имеются одна или несколько двойных связей (C=C). В этом случае жирные кислоты, и содержащие их липиды называются ненасыщенными. Соответсвенно, те кислоты где двойных связей нет, называются насыщенными. Ненасыщенные жирные кислоты плавятся при более низких кислотах, например олеиновая кислота -основной компонент оливкого масла -в обычных условиях бывает жидкой, тогда как стеариновая кислота при обычных условиях -твердая.
Большая часть липидов -это сложные эфиры спирта глицерола. Поэтому их называют глицеридами.
Синтез липидов.
У глицерола имеется три гидроксильные группы (-ОН), каждая из которых способна вступать в реакцию с жирной кислотой, то есть образовывать сложный эфир. Обычно, в реакцию вступают все три гидроксильные группы глицерола (схему образования сложного эфира можно найти в приложениях по липидам), поэтому продукт реакции называют триглицеридом. Биосинтез жирных кислот протекает в цитоплазме, в основном в печени, жировой ткани, почках, легких и молочных железах. Главным источником атомов углерода является глюкоза.
Свойства и функции триглицеридов
Триглицериды -самые распространенные из липидов, встречающихся в природе. Их принято разделять на жиры и масла, в зависимости от того, остаются ли они твердыми при 20℃ (жиры) или имеют жидкую консистенцию при этой температуре (масла). Температура плавления липида тем ниже, чем выше в нем доля ненасыщенных жирных кислот.
Триглицериды неполярны и поэтому нерастворимы в воде. Их плотность ниже, чем у воды, поэтому в воде они всплывают. Калорийность липидов выше чем у углеводов, т.е. данная масса липида выделяет при окислении больше энергии, чем равная ей масса углевода. Это можно объяснить тем, что в липидах, по сравнению с углеводами больше водорода и совсем мало кислорода. Одним из продуктов окисления жиров является вода. Эта метаболическая вода очень важна для обитателей пустыни, так как жир запасаемый в их организмах, используется именно для этой цели.
УглеводыУглеводами называют вещества с общей формулой Cx(h3O)y, где y и x могут иметь различные значения. Название “углеводы” отражает тот факт, что в молекулах этих веществ присутствуют атомы водорода и кислорода в том же соотношении, что и в молекулах воды. Углеводы подразделяют на три главных класса: моносахариды, дисахариды и полисахариды.
/>Синтез углеводов
В животных организмах, углеводы напрямую не синтезируются. Биосинтез -начинаеться в фотосинтезирующих и хемосинтезирующих организмах. Подробнее остановлюсь на фотосинтезирующих -это растения (высшие и низшие) и синезелёные водоросли. В них происходит фотосинтез, образование высшими растениями, водорослями, фотосинтезирующими бактериями сложных органических веществ, необходимых для жизнедеятельности как самих растений, так и всех других организмов, из простых соединений за счёт энергии света, поглощаемой хлорофиллом и другими фотосинтетическими пигментами. Один из важнейших биологических процессов, постоянно и в огромных масштабах совершающийся на нашей планете.
Суммарное (и очень сильно упрощенное) уравнение фотосинтеза выражается следующим образом: 6СО₂ + 6Н₂О = С₆Н₁₂О₆ + 6О₂. Там образуеться много промежуточных органических кислот. При использовании в Фотосинтезе только CO₂и H₂О образуются углеводы.
Из простых углеводов образуются более сложные (сахароза, фруктоза, крохмал и прочие), которые выполняют функции «долгосрочных» запасных веществ. Молекулы углеводов -более-менее универсальны для всех организмов.
Попадая в гетеротрофные организмы -углеводы и белки разбираются на составные части. И из них либо собираются свои молекулы, либо добывается энергия для жизненноважных процессов. Углеводы могут переситезироваться в жиры и кислоты.
Приложения
Синтез белков
Приложение1
/>Приложение2
/>Приложение3
/>Приложение4
/>В таблице на рисунке боковые столбцы кодируют левую и правую букву кодона, верхняя строка – среднюю. Например кодон AUG кодирует аминокислоту метионин. Число комбинаций из 4 по 3 = 64, то есть некоторые аминокислоты кодируются несколькими кодонами. Три кодона не кодируют никакую аминокислоту, они называются терминирующими. Когда они попадаются в мРНК, рибосома прекращает свою работу и готовая полипептидная цепь выбрасывается наружу.
Липиды
Приложение6 Приложение7
/>Стеариновая кислота
/>O
❘❘
CH₃-CH₂-CH₂-CH₂-CH₂-CH₂-CH₂-CH₂-CH₂-CH₂-CH₂-CH₂-CH₂-CH₂-CH₂-CH₂-CH₂-C-OH
Олеиновая кислота
/>О
❘❘
CH₃-CH₂-CH₂-CH₂-CH₂-CH₂-CH₂-CH₂-CH=CH-CH₂-CH₂-CH₂-CH₂-CH₂-CH₂-CH₂-С-ОН
Приложение8 Образование липида(триацилглицерола)
O O
❘❘ ❘❘
CH₂OH HO-C-R¹CH₂OC-R¹
O O ❘❘ ❘❘CHOH HO-C-R²3H₂O + CHOC-R²O O ❘❘ ❘❘CH₂OH HO-C-R³CH₂OC-R³ глицерол три жирные кислоты(три сложноэфирные связи)Углеводы Приложение 9 Схема фотосинтеза
/>ЗаключениеВ процессе написания реферата, я узнал подробнее о том что такое метаболизм, как проиходят реакции метаболизма в организме и какие ученые занимались изучением сих реакций.
Я узнал много нового о процессах, протекающих в клетке на молекулярном уровне. Например, я изучил подробней синтез белка, липидов и углеводов.
На современном этапе развития науки следует больше внимания уделять организму как целому, связывать протекающие процессы с его функционированием, помнить о том, что наша конечная цель состоит в понимании структуры и функция организма в целом на основе знаний об отдельных его системах.
Список литературы1) Биология. Под редакцией Р.Сопера.
Авторы: Н.Грин, У.Стаут, Д.Тейлор.
1996 Москва “Мир”
2) Биология. Человек: Учеб. для 9 кл. общеобразоват. убеч. заведений/ А.С. Батуев, А.Д. Ноздрачев и др.; под ред. А.С. Батуева
3) Animal growth and development by David R. Newth
Edward Arnold (Publishers) Ltd 1970
4) Общая химия: учебное пособие / Н.Л. Глинка — М.: КНОРУС, 2010. -752 с.
5) Гусев М.В., Минеева Л.А. Микробиология. М.: Изд-во МГУ, 1992, 448 с.
www.ronl.ru
Реферат: Метаболизм
Метаболизм
Метаболизм, или обмен веществ, химические превращения, протекающие от момента поступления питательных веществ в живой организм до момента, когда конечные продукты этих превращений выделяются во внешнюю среду. К метаболизму относятся все реакции, в результате которых строятся структурные элементы клеток и тканей, и процессы, в которых из содержащихся в клетках веществ извлекается энергия. Иногда для удобства рассматривают по отдельности две стороны метаболизма – анаболизм и катаболизм, т.е. процессы созидания органических веществ и процессы их разрушения. Анаболические процессы обычно связаны с затратой энергии и приводят к образованию сложных молекул из более простых, катаболические же сопровождаются высвобождением энергии и заканчиваются образованием таких конечных продуктов (отходов) метаболизма, как мочевина, диоксид углерода, аммиак и вода.
Термин «обмен веществ» вошел в повседневную жизнь с тех пор, как врачи стали связывать избыточный или недостаточный вес, чрезмерную нервозность или, наоборот, вялость больного с повышенным или пониженным обменом. Для суждения об интенсивности метаболизма ставят тест на «основной обмен». Основной обмен – это показатель способности организма вырабатывать энергию. Тест проводят натощак в состоянии покоя; измеряют поглощение кислорода (О2) и выделение диоксида углерода (СО2). Сопоставляя эти величины, определяют, насколько полно организм использует («сжигает») питательные вещества. На интенсивность метаболизма влияют гормоны щитовидной железы, поэтому врачи при диагностике заболеваний, связанных с нарушениями обмена, в последнее время все чаще измеряют уровень этих гормонов в крови.
Методы исследования. При изучении метаболизма какого-нибудь одного из питательных веществ прослеживают все его превращения от той формы, в какой оно поступает в организм, до конечных продуктов, выводимых из организма. В таких исследованиях применяется крайне разнообразный набор биохимических методов.
Использование интактных животных или органов. Животному вводят изучаемое соединение, а затем в его моче и экскрементах определяют возможные продукты превращений (метаболиты) этого вещества. Более определенную информацию можно получить, исследуя метаболизм определенного органа, например печени или мозга. В этих случаях вещество вводят в соответствующий кровеносный сосуд, а метаболиты определяют в крови, оттекающей от данного органа.
Поскольку такого рода процедуры сопряжены с большими трудностями, часто для исследования используют тонкие срезы органов. Их инкубируют при комнатной температуре или при температуре тела в растворах с добавкой того вещества, метаболизм которого изучают. Клетки в таких препаратах не повреждены, и так как срезы очень тонкие, вещество легко проникает в клетки и легко выходит из них. Иногда затруднения возникают из-за слишком медленного прохождения вещества сквозь клеточные мембраны. В этих случаях ткани измельчают, чтобы разрушить мембраны, и с изучаемым веществом инкубируют клеточную кашицу. Именно в таких опытах было показано, что все живые клетки окисляют глюкозу до СО2 и воды и что только ткань печени способна синтезировать мочевину.
Использование клеток. Даже клетки представляют собой очень сложно организованные системы. В них имеется ядро, а в окружающей его цитоплазме находятся более мелкие тельца, т.н. органеллы, различных размеров и консистенции. С помощью соответствующей методики ткань можно «гомогенизировать», а затем подвергнуть дифференциальному центрифугированию (разделению) и получить препараты, содержащие только митохондрии, только микросомы или прозрачную жидкость – цитоплазму. Эти препараты можно по отдельности инкубировать с тем соединением, метаболизм которого изучается, и таким путем установить, какие именно субклеточные структуры участвуют в его последовательных превращениях. Известны случаи, когда начальная реакция протекает в цитоплазме, ее продукт подвергается превращению в микросомах, а продукт этого превращения вступает в новую реакцию уже в митохондриях. Инкубация изучаемого вещества с живыми клетками или с гомогенатом ткани обычно не выявляет отдельные этапы его метаболизма, и только последовательные эксперименты, в которых для инкубации используются те или иные субклеточные структуры, позволяют понять всю цепочку событий.
Использование радиоактивных изотопов. Для изучения метаболизма какого-либо вещества необходимы: 1) соответствующие аналитические методы для определения этого вещества и его метаболитов; и 2) методы, позволяющие отличать добавленное вещество от того же вещества, уже присутствующего в данном биологическом препарате. Эти требования служили главным препятствием при изучении метаболизма до тех пор, пока не были открыты радиоактивные изотопы элементов и в первую очередь радиоактивный углерод 14C. С появлением соединений, «меченных» 14C, а также приборов для измерения слабой радиоактивности эти трудности были преодолены. Если к биологическому препарату, например к суспензии митохондрий, добавляют меченную 14C жирную кислоту, то никаких специальных анализов для определения продуктов ее превращений не требуется; чтобы оценить скорость ее использования, достаточно просто измерять радиоактивность последовательно получаемых митохондриальных фракций. Эта же методика позволяет легко отличать молекулы радиоактивной жирной кислоты, введенной экспериментатором, от молекул жирной кислоты, уже присутствовавших в митохондриях к началу эксперимента.
Хроматография и электрофорез. В дополнение к вышеупомянутым требованиям биохимику необходимы и методы, позволяющие разделять смеси, состоящие из малых количеств органических веществ. Важнейший из них – хроматография, в основе которой лежит феномен адсорбции. Разделение компонентов смеси проводят при этом либо на бумаге, либо путем адсорбции на сорбенте, которым заполняют колонки (длинные стеклянные трубки), с последующей постепенной элюцией (вымыванием) каждого из компонентов.
Разделение методом электрофореза зависит от знака и числа зарядов ионизированных молекул. Электрофорез проводят на бумаге или на каком-нибудь инертном (неактивном) носителе, таком, как крахмал, целлюлоза или каучук.
Высокочувствительный и эффективный метод разделения – газовая хроматография. Им пользуются в тех случаях, когда подлежащие разделению вещества находятся в газообразном состоянии или могут быть в него переведены.
Выделение ферментов. Последнее место в описываемом ряду – животное, орган, тканевой срез, гомогенат и фракция клеточных органелл – занимает фермент, способный катализировать определенную химическую реакцию. Выделение ферментов в очищенном виде – важный раздел в изучении метаболизма.
Сочетание перечисленных методов позволило проследить главные метаболические пути у большей части организмов (в том числе у человека), установить, где именно эти различные процессы протекают, и выяснить последовательные этапы главных метаболических путей. К настоящему времени известны тысячи отдельных биохимических реакций, изучены участвующие в них ферменты.
Клеточный метаболизм. Живая клетка – это высокоорганизованная система. В ней имеются различные структуры, а также ферменты, способные их разрушить. Содержатся в ней и крупные макромолекулы, которые могут распадаться на более мелкие компоненты в результате гидролиза (расщепления под действием воды). В клетке обычно много калия и очень мало натрия, хотя клетка существует в среде, где натрия много, а калия относительно мало, и клеточная мембрана легко проницаема для обоих ионов. Следовательно, клетка – это химическая система, весьма далекая от равновесия. Равновесие наступает только в процессе посмертного автолиза (самопереваривания под действием собственных ферментов).
Потребность в энергии. Чтобы удержать систему в состоянии, далеком от химического равновесия, требуется производить работу, а для этого необходима энергия. Получение этой энергии и выполнение этой работы – непременное условие для того, чтобы клетка оставалась в своем стационарном (нормальном) состоянии, далеком от равновесия. Одновременно в ней выполняется и иная работа, связанная со взаимодействием со средой, например: в мышечных клетках – сокращение; в нервных клетках – проведение нервного импульса; в клетках почек – образование мочи, значительно отличающейся по своему составу от плазмы крови; в специализированных клетках желудочно-кишечного тракта – синтез и выделение пищеварительных ферментов; в клетках эндокринных желез – секреция гормонов; в клетках светляков – свечение; в клетках некоторых рыб – генерирование электрических разрядов и т.д.
Источники энергии. В любом из перечисленных выше примеров непосредственным источником энергии, которую клетка использует для производства работы, служит энергия, заключенная в структуре аденозинтрифосфата (АТФ). В силу особенностей своей структуры это соединение богато энергией, и разрыв связей между его фосфатными группами может происходить таким образом, что высвобождающаяся энергия используется для производства работы. Однако энергия не может стать доступной для клетки при простом гидролитическом разрыве фосфатных связей АТФ: в этом случае она расходуется впустую, выделяясь в виде тепла. Процесс должен состоять из двух последовательных этапов, в каждом из которых участвует промежуточный продукт, обозначенный здесь X–Ф (в приведенных уравнениях X и Y означают два разных органических вещества; Ф – фосфат; АДФ – аденозиндифосфат):
Поскольку практически для любого проявления жизнедеятельности клеток необходим АТФ, неудивительно, что метаболическая активность живых клеток направлена в первую очередь на синтез АТФ. Этой цели служат различные сложные последовательности реакций, в которых используется потенциальная химическая энергия, заключенная в молекулах углеводов и жиров (липидов).
Метаболизм углеводов и липидов
Синтез АТФ. Анаэробный (без участия кислорода). Главная роль углеводов и липидов в клеточном метаболизме состоит в том, что их расщепление на более простые соединения обеспечивает синтез АТФ. Несомненно, что те же процессы протекали и в первых, самых примитивных клетках. Однако в атмосфере, лишенной кислорода, полное окисление углеводов и жиров до CO2 было невозможно. У этих примитивных клеток имелись все же механизмы, с помощью которых перестройка структуры молекулы глюкозы обеспечивала синтез небольших количеств АТФ. Речь идет о процессах, которые у микроорганизмов называют брожением. Лучше всего изучено сбраживание глюкозы до этилового спирта и CO2 у дрожжей.
В ходе 11 последовательных реакций, необходимых для того, чтобы завершилось это превращение, образуется ряд промежуточных продуктов, представляющих собой эфиры фосфорной кислоты (фосфаты). Их фосфатная группа переносится на аденозиндифосфат (АДФ) с образованием АТФ. Чистый выход АТФ составляет 2 молекулы АТФ на каждую молекулу глюкозы, расщепленную в процессе брожения. Аналогичные процессы происходят во всех живых клетках; поскольку они поставляют необходимую для жизнедеятельности энергию, их иногда (не вполне корректно) называют анаэробным дыханием клеток.
У млекопитающих, в том числе у человека, такой процесс называется гликолизом и его конечным продуктом является молочная кислота, а не спирт и CO2. Вся последовательность реакций гликолиза, за исключением двух последних этапов, полностью идентична процессу, протекающему в дрожжевых клетках.
Аэробный (с использованием кислорода). С появлением в атмосфере кислорода, источником которого послужил, очевидно, фотосинтез растений, в ходе эволюции развился механизм, обеспечивающий полное окисление глюкозы до CO2 и воды, – аэробный процесс, в котором чистый выход АТФ составляет 38 молекул АТФ на каждую окисленную молекулу глюкозы. Этот процесс потребления клетками кислорода для образования богатых энергией соединений известен как клеточное дыхание (аэробное). В отличие от анаэробного процесса, осуществляемого ферментами цитоплазмы, окислительные процессы протекают в митохондриях. В митохондриях пировиноградная кислота – промежуточный продукт, образовавшийся в анаэробной фазе – окисляется до СО2 в шести последовательных реакциях, в каждой из которых пара электронов переносится на общий акцептор – кофермент никотинамидадениндинуклеотид (НАД). Эту последовательность реакций называют циклом трикарбоновых кислот, циклом лимонной кислоты или циклом Кребса. Из каждой молекулы глюкозы образуется 2 молекулы пировиноградной кислоты; 12 пар электронов отщепляется от молекулы глюкозы в ходе ее окисления, описываемого уравнением:
Перенос электронов. В каждой митохондрии имеется механизм, посредством которого восстановленный НАД (НАДН, где Н – водород), образовавшийся в цикле трикарбоновых кислот, передает свою пару электронов кислороду. Перенос, однако, не происходит напрямую. Электроны как бы передаются «из рук в руки» и, лишь пройдя цепь переносчиков, присоединяются к кислороду. Эта «цепь переноса электронов» состоит из следующих компонентов:
НАДНН Флавинадениндинклеотид Кофермент Q
Цитохром b Цитохром c Цитохром a O2
Все компоненты этой системы, находящиеся в митохондриях, фиксированы в пространстве и сцеплены друг с другом. Такое их состояние облегчает перенос электронов.
В состав НАД входит никотиновая кислота (витамин ниацин), а в состав флавинадениндинуклеотида – рибофлавин (витамин B2). Кофермент Q представляет собой высокомолекулярный хинон, синтезируемый в печени, а цитохромы – это три разных белка, каждый из которых, подобно гемоглобину, содержит гемогруппу.
В цепи переноса электронов на каждую пару электронов, перенесенную от НАДН на O2, синтезируется 3 молекулы АТФ. Поскольку от каждой молекулы глюкозы отщепляются и передаются молекулам НАД 12 пар электронов, в общей сложности на каждую молекулу глюкозы образуется 312 = 36 молекул АТФ. Этот процесс образования АТФ в ходе окисления называется окислительным фосфорилированием.
Липиды как источник энергии. Жирные кислоты могут использоваться в качестве источника энергии приблизительно так же, как и углеводы. Окисление жирных кислот протекает путем последовательного отщепления от молекулы жирной кислоты двууглеродного фрагмента с образованием ацетилкофермента A (ацетил-КоА) и одновременной передачей двух пар электронов в цепь переноса электронов. Образовавшийся ацетил-КоА – нормальный компонент цикла трикарбоновых кислот, и в дальнейшем его судьба не отличается от судьбы ацетил-КоА, поставляемого углеводным обменом. Таким образом, механизмы синтеза АТФ при окислении как жирных кислот, так и метаболитов глюкозы практически одинаковы.
Если организм животного получает энергию почти целиком за счет одного только окисления жирных кислот, а это бывает, например, при голодании или при сахарном диабете, то скорость образования ацетил-КоА превышает скорость его окисления в цикле трикарбоновых кислот. В этом случае лишние молекулы ацетил-КоА реагируют друг с другом, в результате чего образуются в конечном счете ацетоуксусная и -гидроксимасляная кислоты. Их накопление является причиной патологического состояния, т.н. кетоза (одного из видов ацидоза), который при тяжелом диабете может вызвать кому и смерть.
Запасание энергии. Животные питаются нерегулярно, и их организму нужно как-то запасать заключенную в пище энергию, источником которой являются поглощенные животным углеводы и жиры. Жирные кислоты могут запасаться в виде нейтральных жиров либо в печени, либо в жировой ткани. Углеводы, поступая в большом количестве, в желудочно-кишечном тракте гидролизуются до глюкозы или иных сахаров, которые затем в печени превращаются в ту же глюкозу. Здесь из глюкозы синтезируется гигантский полимер гликоген путем присоединения друг к другу остатков глюкозы с отщеплением молекул воды (число остатков глюкозы в молекулах гликогена доходит до 30 000). Когда возникает потребность в энергии, гликоген вновь распадается до глюкозы в реакции, продуктом которой является глюкозофосфат. Этот глюкозофосфат направляется на путь гликолиза – процесса, составляющего часть пути окисления глюкозы. В печени глюкозофосфат может также подвергнуться гидролизу, и образующаяся глюкоза поступает в кровоток и доставляется кровью к клеткам в разных частях тела.
Синтез липидов из углеводов. Если количество углеводов, поглощенных с пищей за один прием, больше того, какое может быть запасено в виде гликогена, то избыток углеводов превращается в жиры. Начальная последовательность реакций совпадает при этом с обычным окислительным путем, т.е. сначала из глюкозы образуется ацетил-КоА, но далее этот ацетил-КоА используется в цитоплазме клетки для синтеза длинноцепочечных жирных кислот. Процесс синтеза можно описать как обращение обычного процесса окисления жирных клеток. Затем жирные кислоты запасаются в виде нейтральных жиров (триглицеридов), отлагающихся в разных частях тела. Когда требуется энергия, нейтральные жиры подвергаются гидролизу и жирные кислоты поступают в кровь. Здесь они адсорбируются молекулами плазменных белков (альбуминов и глобулинов) и затем поглощаются клетками самых разных типов. Механизмов, способных осуществлять синтез глюкозы из жирных кислот, у животных нет, но у растений такие механизмы имеются.
Метаболизм липидов. Липиды попадают в организм главным образом в форме триглицеридов жирных кислот. В кишечнике под действием ферментов поджелудочной железы они подвергаются гидролизу, продукты которого всасываются клетками стенки кишечника. Здесь из них вновь синтезируются нейтральные жиры, которые через лимфатическую систему поступают в кровь и либо транспортируются в печень, либо отлагаются в жировой ткани. Выше уже указывалось, что жирные кислоты могут также синтезироваться заново из углеводных предшественников. Следует отметить, что, хотя в клетках млекопитающих может происходить включение одной двойной связи в молекулы длинноцепочечных жирных кислот (между С–9 и С–10), включать вторую и третью двойную связь эти клетки неспособны. Поскольку жирные кислоты с двумя и тремя двойными связями играют важную роль в метаболизме млекопитающих, они в сущности являются витаминами. Поэтому линолевую (C18:2) и линоленовую (C18:3) кислоты называют незаменимыми жирными кислотами. В то же время в клетках млекопитающих в линоленовую кислоту может включаться четвертая двойная связь и путем удлинения углеродной цепи может образоваться арахидоновая кислота (C20:4), также необходимый участник метаболических процессов.
В процессе синтеза липидов остатки жирных кислот, связанные с коферментом А (ацил-КоА), переносятся на глицерофосфат – эфир фосфорной кислоты и глицерина. В результате образуется фосфатидная кислота – соединение, в котором одна гидроксильная группа глицерина этерифицирована фосфорной кислотой, а две группы – жирными кислотами. При образовании нейтральных жиров фосфорная кислота удаляется путем гидролиза, и ее место занимает третья жирная кислота в результате реакции с ацил-КоА. Кофермент А образуется из пантотеновой кислоты (одного из витаминов). В его молекуле имеется сульфгидрильная (– SH) группа, способная реагировать с кислотами с образованием тиоэфиров. При образовании фосфолипидов фосфатидная кислота реагирует непосредственно с активированным производным одного из азотистых оснований, таких, как холин, этаноламин или серин.
За исключением витамина D, все встречающиеся в организме животных стероиды (производные сложных спиртов) легко синтезируются самим организмом. Сюда относятся холестерин (холестерол), желчные кислоты, мужские и женские половые гормоны и гормоны надпочечников. В каждом случае исходным материалом для синтеза служит ацетил-КоА: из ацетильных групп путем многократно повторяющейся конденсации строится углеродный скелет синтезируемого соединения.
Метаболизм белков
Синтез аминокислот. Растения и большинство микроорганизмов могут жить и расти в среде, в которой для их питания имеются только минеральные вещества, диоксид углерода и вода. Это значит, что все обнаруживаемые в них органические вещества эти организмы синтезируют сами. Встречающиеся во всех живых клетках белки построены из 21 вида аминокислот, соединенных в различной последовательности. Аминокислоты синтезируются живыми организмами. В каждом случае ряд химических реакций приводит к образованию -кетокислоты. Одна такая -кетокислота, а именно -кетоглутаровая (обычный компонент цикла трикарбоновых кислот), участвует в связывании азота по следующему уравнению:
-Кетоглутаровая кислота + Nh4 + НАДН
Глутаминовая кислота + НАД.
Азот глутаминовой кислоты может быть затем передан любой из других -кетокислот с образованием соответствующей аминокислоты.
Организм человека и большинства других животных сохранил способность синтезировать все аминокислоты за исключением девяти т.н. незаменимых аминокислот. Поскольку кетокислоты, соответствующие этим девяти, не синтезируются, незаменимые аминокислоты должны поступать с пищей.
Синтез белков. Аминокислоты нужны для биосинтеза белка. Процесс биосинтеза протекает обычно следующим образом. В цитоплазме клетки каждая аминокислота «активируется» в реакции с АТФ, а затем присоединяется к концевой группе молекулы рибонуклеиновой кислоты, специфичной именно для данной аминокислоты. Эта сложная молекула связывается с небольшим тельцем, т.н. рибосомой, в положении, определяемом более длинной молекулой рибонуклеиновой кислоты, прикрепленной к рибосоме. После того как все эти сложные молекулы соответствующим образом выстроились, связи между исходной аминокислотой и рибонуклеиновой кислотой разрываются и возникают связи между соседними аминокислотами – синтезируется специфичный белок. Процесс биосинтеза поставляет белки не только для роста организма или для секреции в среду. Все белки живых клеток со временем претерпевают распад до составляющих их аминокислот, и для поддержания жизни клетки должны синтезироваться вновь.
Синтез других азотсодержащих соединений. В организме млекопитающих аминокислоты используются не только для биосинтеза белков, но и как исходный материал для синтеза многих азотсодержащих соединений. Аминокислота тирозин является предшественником гормонов адреналина и норадреналина. Простейшая аминокислота глицин служит исходным материалом для биосинтеза пуринов, входящих в состав нуклеиновых кислот, и порфиринов, входящих в состав цитохромов и гемоглобина. Аспарагиновая кислота – предшественник пиримидинов нуклеиновых кислот. Метильная группа метионина передается ряду других соединений в ходе биосинтеза креатина, холина и саркозина. При биосинтезе креатина от одного соединения к другому передается также и гуанидиновая группировка аргинина. Триптофан служит предшественником никотиновой кислоты, а из валина в растениях синтезируется такой витамин, как пантотеновая кислота. Все это лишь отдельные примеры использования аминокислот в процессах биосинтеза.
Азот, поглощаемый микроорганизмами и высшими растениями в виде иона аммония, расходуется почти целиком на образование аминокислот, из которых затем синтезируются многие азотсодержащие соединения живых клеток. Избыточных количеств азота ни растения, ни микроорганизмы не поглощают. В отличие от них, у животных количество поглощенного азота зависит от содержащихся в пище белков. Весь азот, поступивший в организм в виде аминокислот и не израсходованный в процессах биосинтеза, довольно быстро выводится из организма с мочой. Происходит это следующим образом. В печени неиспользованные аминокислоты передают свой азот -кетоглутаровой кислоте с образованием глутаминовой кислоты, которая дезаминируется, высвобождая аммиак. Далее азот аммиака может либо на время запасаться путем синтеза глутамина, либо сразу же использоваться для синтеза мочевины, протекающего в печени.
У глутамина есть и другая роль. Он может подвергаться гидролизу в почках с высвобождением аммиака, который поступает в мочу в обмен на ионы натрия. Этот процесс крайне важен как средство поддержания кислотно-щелочного равновесия в организме животного. Почти весь аммиак, происходящий из аминокислот и, возможно, из других источников, превращается в печени в мочевину, так что свободного аммиака в крови обычно почти нет. Однако при некоторых условиях довольно значительные количества аммиака содержит моча. Этот аммиак образуется в почках из глутамина и переходит в мочу в обмен на ионы натрия, которые таким образом реадсорбируются и задерживаются в организме. Этот процесс усиливается при развитии ацидоза – состояния, при котором организм нуждается в дополнительных количествах катионов натрия для связывания избытка ионов бикарбоната в крови.
Избыточные количества пиримидинов тоже распадаются в печени через ряд реакций, в которых высвобождается аммиак. Что касается пуринов, то их избыток подвергается окислению с образованием мочевой кислоты, выделяющейся с мочой у человека и других приматов, но не у остальных млекопитающих. У птиц отсутствует механизм синтеза мочевины, и именно мочевая кислота, а не мочевина, является у них конечным продуктом обмена всех азотсодержащих соединений.
Общие представления о метаболизме органических веществ
Можно сформулировать некоторые общие понятия, или «правила», касающиеся метаболизма. Приведенные ниже несколько главных «правил» позволяют лучше понять, как протекает и регулируется метаболизм.
1. Метаболические пути необратимы. Распад никогда не идет по пути, который являлся бы простым обращением реакций синтеза. В нем участвуют другие ферменты и другие промежуточные продукты. Нередко противоположно направленные процессы протекают в разных отсеках клетки. Так, жирные кислоты синтезируются в цитоплазме при участии одного набора ферментов, а окисляются в митохондриях при участии совсем другого набора.
2. Ферментов в живых клетках достаточно для того, чтобы все известные метаболические реакции могли протекать гораздо быстрее, чем это обычно наблюдается в организме. Следовательно, в клетках существуют какие-то регуляторные механизмы. Открыты разные типы таких механизмов.
а) Фактором, ограничивающим скорость метаболических превращений данного вещества, может быть поступление этого вещества в клетку; именно на этот процесс в таком случае и направлена регуляция. Роль инсулина, например, связана с тем, что он, по-видимому, облегчает проникновение глюкозы во все клетки, глюкоза же подвергается превращениям с той скоростью, с какой она поступает. Сходным образом проникновение железа и кальция из кишечника в кровь зависит от процессов, скорость которых регулируется.
б) Вещества далеко не всегда могут свободно переходить из одного клеточного отсека в другой; есть данные, что внутриклеточный перенос регулируется некоторыми стероидными гормонами.
в) Выявлено два типа сервомеханизмов «отрицательной обратной связи».
У бактерий были обнаружены примеры того, что присутствие продукта какой-нибудь последовательности реакций, например аминокислоты, подавляет биосинтез одного из ферментов, необходимых для образования этой аминокислоты.
В каждом случае фермент, биосинтез которого оказывается затронутым, был ответствен за первый «определяющий» этап (на схеме реакция 4) метаболического пути, ведущего к синтезу данной аминокислоты.
Второй механизм хорошо изучен у млекопитающих. Это простое ингибирование конечным продуктом (в нашем случае – аминокислотой) фермента, ответственного за первый «определяющий» этап метаболического пути.
Еще один тип регулирования посредством обратной связи действует в тех случаях, когда окисление промежуточных продуктов цикла трикарбоновых кислот сопряжено с образованием АТФ из АДФ и фосфата в процессе окислительного фосфорилирования. Если весь имеющийся в клетке запас фосфата и (или) АДФ уже исчерпан, то окисление приостанавливается и может возобновиться лишь после того, как этот запас вновь станет достаточным. Таким образом, окисление, смысл которого в том, чтобы поставлять полезную энергию в форме АТФ, происходит только тогда, когда возможен синтез АТФ.
3. В биосинтетических процессах участвует сравнительно небольшое число строительных блоков, каждый из которых используется для синтеза многих соединений. Среди них можно назвать ацетилкофермент А, глицерофосфат, глицин, карбамилфосфат, поставляющий карбамильную (h3N–CO–) группу, производные фолиевой кислоты, служащие источником гидроксиметильной и формильной групп, S-аденозилметионин – источник метильных групп, глутаминовую и аспарагиновую кислоты, поставляющие аминогруппы, и наконец, глутамин – источник амидных групп. Из этого относительно небольшого числа компонентов строятся все те разнообразные соединения, которые мы находим в живых организмах.
4. Простые органические соединения редко участвуют в метаболических реакциях непосредственно. Обычно они должны быть сначала «активированы» путем присоединения к одному из ряда соединений, универсально используемых в метаболизме. Глюкоза, например, может подвергнуться окислению лишь после того, как она будет этерифицирована фосфорной кислотой, для прочих же своих превращений она должна быть этерифицирована уридиндифосфатом. Жирные кислоты не могут быть вовлечены в метаболические превращения прежде, чем они образуют эфиры с коферментом А. Каждый из этих активаторов либо родствен одному из нуклеотидов, входящих в состав рибонуклеиновой кислоты, либо образуется из какого-нибудь витамина. Легко понять в связи с этим, почему витамины требуются в таких небольших количествах. Они расходуются на образование «коферментов», а каждая молекула кофермента на протяжении жизни организма используется многократно, в отличие от основных питательных веществ (например, глюкозы), каждая молекула которых используется только один раз.
В заключение следует сказать, что термин «метаболизм», означавший ранее нечто не более сложное, чем просто использование углеводов и жиров в организме, теперь применяется для обозначения тысяч ферментативных реакций, вся совокупность которых может быть представлена как огромная сеть метаболических путей, многократно пересекающихся (из-за наличия общих промежуточных продуктов) и управляемых очень тонкими регуляторными механизмами.
Метаболизм минеральных веществ
Относительное содержание. Различные элементы, встречающиеся в живых организмах, перечислены ниже в убывающем порядке в зависимости от их относительного содержания: 1) кислород, углерод, водород и азот; 2) кальций, фосфор, калий и сера; 3) натрий, хлор, магний и железо; 4) марганец, медь, молибден, селен, йод и цинк; 5) алюминий, фтор, кремний и литий; 6) бром, мышьяк, свинец и, возможно, некоторые другие.
Кислород, углерод, водород и азот – это те элементы, из которых построены мягкие ткани тела. Они входят в состав таких соединений, как углеводы, липиды, белки, вода, диоксид углерода и аммиак. Элементы, перечисленные в пп. 2 и 3, находятся в организме обычно в виде одного или нескольких неорганических соединений, а элементы пп. 4, 5 и 6 присутствуют только в следовых количествах и потому их называют микроэлементами.
Распределение в организме. Кальций. Кальций присутствует главным образом в костной ткани и в зубах, преимущественно в виде фосфата и в небольших количествах в виде карбоната и фторида. Поступающий с пищей кальций всасывается в основном в верхних отделах кишечника, имеющих слабокислую реакцию. Этому всасыванию (у человека здесь всасывается всего 20–30% кальция пищи) способствует витамин D. Под действием витамина D клетки кишечника вырабатывают особый белок, который связывает кальций и облегчает его перенос через стенку кишечника в кровь. На всасывание влияет также присутствие некоторых других веществ, в особенности фосфата и оксалата, которые в малых количествах способствуют всасыванию, а в больших, наоборот, подавляют его.
В крови около половины кальция связано с белком, остальное составляют ионы кальция. Соотношение ионизированной и неионизированной форм зависит от общей концентрации кальция в крови, а также от содержания белка и фосфата и концентрации водородных ионов (рН крови). Доля неионизированного кальция, на которую влияет уровень белка, позволяет косвенным образом судить о качестве питания и об эффективности работы печени, в которой идет синтез плазменных белков.
На количество ионизированного кальция влияют, с одной стороны, витамин D и факторы, воздействующие на всасывание, а с другой – паратиреоидный гормон и, возможно, также витамин D, поскольку оба эти вещества регулируют как скорость отложения кальция в костной ткани, так и его мобилизацию, т.е. вымывание из костей. Избыток паратиреоидного гормона стимулирует выход кальция из костной ткани, что приводит к повышению его концентрации в плазме. Изменяя скорости всасывания и экскреции кальция и фосфата, а также скорости образования костной ткани и ее разрушения, эти механизмы строго контролируют концентрацию кальция и фосфата в сыворотке крови. Ионы кальция играют регулирующую роль во многих физиологических процессах, в том числе в нервных реакциях, мышечном сокращении, свертывании крови. Выведение кальция из организма происходит в норме в основном (на 2/3) через желчь и кишечник и в меньшей степени (1/3) – через почки.
Фосфор. Метаболизм фосфора – одного из главных компонентов костной ткани и зубов – во многом зависит от тех же факторов, что и метаболизм кальция. Фосфор в виде фосфата присутствует в организме также в сотнях различных физиологически важных органических эфиров. Паратиреоидный гормон стимулирует выведение фосфора с мочой и выход его из костной ткани; тем самым он регулирует концентрацию фосфора в плазме крови.
Натрий. Натрий – главный катион внеклеточной жидкости – вместе с белком, хлоридом и бикарбонатом играет важнейшую роль в регулировании осмотического давления и pH (концентрации водородных ионов) крови. В клетках, напротив, содержится очень мало натрия, так как они обладают механизмом для выведения ионов натрия и удержания ионов калия. Весь натрий, превышающий потребности организма, очень быстро выводится через почки.
Поскольку во всех процессах выделения натрий теряется, он должен постоянно поступать в организм с пищей. При ацидозе, когда необходимо, чтобы из организма выводились большие количества анионов (например, хлорида или ацетоацетата), почки предотвращают чрезмерную потерю натрия благодаря образованию аммиака из глутамина. Выведение натрия через почки регулируется гормоном коры надпочечников альдостероном. Под действием этого гормона в кровь возвращается достаточно натрия для поддержания нормального осмотического давления и нормального объема внеклеточной жидкости.
Суточная потребность в хлористом натрии составляет 5–10 г. Эта величина возрастает при поглощении больших количеств жидкости, когда усиливается потоотделение и выделяется больше мочи.
Калий. В отличие от натрия, калий содержится в клетках в больших количествах, но во внеклеточной жидкости его мало. Главная функция калия – регулирование внутриклеточного осмотического давления и поддержание кислотно-щелочного равновесия. Он также играет важную роль в проведении нервного импульса и во многих ферментных системах, в том числе и в тех, которые участвуют в мышечном сокращении. Калий широко распространен в природе, и его много в любой пище, так что спонтанно калиевая недостаточность возникнуть не может. В плазме концентрация калия регулируется альдостероном, стимулирующим его экскрецию с мочой.
Сера. С пищей сера поступает в организм главным образом в составе двух аминокислот – цистина и метионина. На конечных этапах метаболизма этих аминокислот сера высвобождается и в результате окисления переводится в неорганическую форму. В составе цистина и метионина сера присутствует в структурных белках. Важную роль играет также сульфгидрильная (–SH) группа цистеина, от которой зависит активность многих ферментов.
Большая часть серы выводится с мочой в виде сульфата. Небольшое количество экскретируемого сульфата обычно связано с органическими соединениями типа фенолов.
Магний. Метаболизм магния сходен с метаболизмом кальция, и в виде комплекса с фосфатом этот элемент тоже входит в состав костной ткани. Магний присутствует во всех живых клетках, где он функционирует как необходимый компонент многих ферментных систем; эта его роль была убедительно продемонстрирована на примере углеводного обмена в мышцах. Магний, как и калий, широко распространен, и вероятность возникновения его недостаточности очень мала.
Железо. Железо входит в состав гемоглобина и других гемопротеинов, а именно миоглобина (мышечного гемоглобина), цитохромов (дыхательных ферментов) и каталазы, а также в состав некоторых ферментов, не содержащих гемогруппы. Всасывается железо в верхних отделах кишечника, причем это единственный элемент, всасывающийся только тогда, когда его запас в организме полностью исчерпан. В плазме железо транспортируется в соединении с белком (трансферрином). Через почки железо не выводится; избыток его накапливается в печени в соединении с особым белком (ферритином).
Микроэлементы. У каждого микроэлемента, присутствующего в организме, своя особая функция, связанная с тем, что он стимулирует действие того или иного фермента или как-либо иначе на него влияет. Цинк необходим для кристаллизации инсулина; кроме того, он является компонентом карбоангидразы (фермента, участвующего в транспорте диоксида углерода) и некоторых других ферментов. Молибден и медь – тоже необходимые компоненты различных ферментов. Иод требуется для синтеза трииодтиронина, гормона щитовидной железы. Фтор (входящий в состав зубной эмали) способствует предотвращению кариеса.
Использование метаболитов
Углеводы. Всасывание. Моносахариды, или простые сахара, высвобождающиеся при переваривании углеводов пищи, переходят из кишечника в кровоток в результате процесса, называемого всасыванием. Механизм всасывания представляет собой сочетание простой диффузии и химической реакции (активного всасывания). Одна из гипотез, касающихся природы химической фазы процесса, предполагает, что в этой фазе моносахариды соединяются с фосфорной кислотой в реакции, катализируемой ферментом из группы киназ, после чего проникают в кровеносные сосуды и здесь высвобождаются в результате ферментативного дефосфорилирования (разрыва фосфатной связи), катализируемого одной из фосфатаз. Именно активным всасыванием объясняется то, что разные моносахариды всасываются с разной скоростью и что углеводы всасываются даже тогда, когда уровень сахара в крови выше, чем в кишечнике, т.е. в условиях, когда естественно было бы ожидать их перемещения в обратном направлении – из крови в кишечник.
Механизмы гомеостаза. Поступающие в кровоток моносахариды повышают уровень сахара в крови. При голодании концентрация глюкозы в крови колеблется обычно от 70 до 100 мг на 100 мл крови. Этот уровень поддерживается с помощью механизмов, называемых механизмами гомеостаза (самостабилизации). Как только уровень сахара в крови в результате всасывания из кишечника повышается, в действие вступают процессы, выводящие сахар из крови, так что уровень его колеблется не слишком сильно.
Подобно глюкозе, все прочие моносахариды поступают из кровотока в печень, где превращаются в глюкозу. Теперь они неотличимы как от глюкозы, которая всосалась, так и от той, что уже была в организме, и подвергаются тем же метаболическим превращениям. Один из механизмов гомеостаза углеводов, функционирующий в печени, – это гликогенез, посредством которого глюкоза переходит из крови в клетки, где превращается в гликоген. Гликоген хранится в печени до тех пор, пока не произойдет снижение уровня сахара в крови: в этой ситуации гомеостатический механизм вызовет распад накопленного гликогена до глюкозы, которая вновь поступит в кровь.
Превращения и использование. Поскольку кровь поставляет глюкозу во все ткани тела и все ткани используют ее для получения энергии, уровень глюкозы в крови снижается главным образом за счет ее использования.
В мышцах глюкоза крови превращается в гликоген. Однако мышечный гликоген не может быть использован для получения глюкозы, которая перешла бы в кровь. В нем заключен запас энергии, и скорость его использования зависит от мышечной активности. В мышечной ткани содержатся два соединения с большим запасом легко доступной энергии в форме богатых энергией фосфатных связей – креатинфосфат и аденозинтрифосфат (АТФ). При отщеплении от этих соединений их фосфатных групп высвобождается энергия для мышечного сокращения. Чтобы мышца вновь могла сокращаться, эти соединения должны быть восстановлены в своей исходной форме. Для этого требуется энергия, которую поставляет окисление продуктов распада гликогена. При мышечном сокращении гликоген превращается в глюкозофосфат, а затем – через ряд реакций – во фруктозодифосфат. Фруктозодифосфат распадается на два трехуглеродных соединения, из которых после ряда этапов образуется сначала пировиноградная кислота, а в конечном итоге – молочная кислота, как об этом уже говорилось при описании метаболизма углеводов. Это превращение гликогена в молочную кислоту, сопровождающееся высвобождением энергии, может происходить в отсутствие кислорода.
При недостатке кислорода молочная кислота накапливается в мышцах, диффундирует в кровоток и поступает в печень, где из нее вновь образуется гликоген. Если кислорода достаточно, то молочная кислота в мышцах не накапливается. Вместо этого она, как это описано выше, полностью окисляется через цикл трикарбоновых кислот до диоксида углерода и воды с образованием АТФ, который может быть использован для сокращения.
Метаболизм углеводов в нервной ткани и эритроцитах отличается от метаболизма в мышцах тем, что гликоген здесь не участвует. Однако и здесь промежуточными продуктами являются пировиноградная и молочная кислоты, образующиеся при расщеплении глюкозофосфата.
Глюкоза используется не только в клеточном дыхании, но и во многих других процессах: синтезе лактозы (молочного сахара), образовании жиров, а также особых сахаров, входящих в состав полисахаридов соединительной ткани и ряда других тканей.
Гликоген печени, синтезируемый при всасывании углеводов в кишечнике, служит самым доступным источником глюкозы, когда всасывание отсутствует. Если этот источник оказывается исчерпанным, в печени начинается процесс глюконеогенеза. Глюкоза образуется при этом из некоторых аминокислот (из 100 г белка образуется 58 г глюкозы) и нескольких других неуглеводных соединений, в том числе из глицериновых остатков нейтральных жиров.
Некоторую, хотя и не столь важную, роль в метаболизме углеводов играют почки. Они выводят из организма избыток глюкозы, когда ее концентрация в крови слишком высока; при меньших концентрациях глюкоза практически не выводится.
В регулировании метаболизма углеводов участвует несколько гормонов, в том числе гормоны поджелудочной железы, передней доли гипофиза и коры надпочечников.
Гормон поджелудочной железы инсулин снижает концентрацию глюкозы в крови и повышает ее концентрацию в клетках. По-видимому, он стимулирует также и запасание гликогена в печени. Кортикостерон, гормон коры надпочечников, и адреналин, вырабатываемый мозговым веществом надпочечников, воздействуют на метаболизм углеводов, стимулируя распад гликогена (главным образом в мышцах и печени) и синтез глюкозы (в печени).
Липиды. Всасывание. В кишечнике после переваривания жиров остаются главным образом свободные жирные кислоты с небольшой примесью холестерина и лецитина и следами жирорастворимых витаминов. Все эти вещества очень тонко диспергированы благодаря эмульгирующему и солюбилизирующему действию солей желчных кислот. Солюбилизирующее действие обычно связывают с образованием нестойких химических соединений между жирными кислотами и солями желчных кислот. Эти комплексы проникают в клетки эпителия тонкого кишечника и здесь распадаются на жирные кислоты и соли желчных кислот. Последние переносятся в печень и вновь секретируются с желчью, а жирные кислоты вступают в соединение с глицерином или холестерином. Образовавшиеся реконструированные жиры поступают в лимфатические сосуды брыжейки в форме млечного сока, т.н. «хилуса». Из сосудов брыжейки хилус по лимфатической системе через грудной проток поступает в кровеносную систему.
После переваривания пищи содержание липидов в крови возрастает приблизительно от 500 мг (уровень при голодании) до 1000 мг на 100 мл плазмы. Присутствующие в крови липиды представляют собой смесь жирных кислот, нейтральных жиров, фосфолипидов (лецитина и кефалина), холестерина и эфиров холестерина.
Распределение. Кровь доставляет липиды в разные ткани тела и прежде всего в печень. Печень обладает способностью модифицировать поступающие в нее жирные кислоты. Это особенно выражено у видов, запасающих жиры с высоким содержанием насыщенных или, наоборот, ненасыщенных жирных кислот: в печени этих животных соотношение насыщенных и ненасыщенных кислот изменяется таким образом, что отлагающийся жир по своему составу соответствует жиру, свойственному данному организму.
Жиры в печени либо используются для получения энергии, либо переходят в кровь и доставляются ею в разные ткани. Здесь они могут включаться в структурные элементы тканей, но большая их часть отлагается в жировых депо, где они хранятся до тех пор, пока не возникнет потребность в энергии; тогда они снова переносятся в печень и подвергаются здесь окислению.
Метаболизм липидов, как и углеводов, регулируется гомеостатически. Механизмы гомеостаза, воздействующие на липидный и углеводный обмен, видимо, тесно связаны, поскольку при замедлении метаболизма углеводов усиливается метаболизм липидов, и наоборот.
Превращения и использование. Четырехуглеродные кислоты – ацетоуксусная (продукт конденсации двух ацетатных единиц) и -гидроксимасляная – и трехуглеродное соединение ацетон, образующийся при отщеплении одного атома углерода от ацетоуксусной кислоты, известны под общим названием кетоновых (ацетоновых) тел. В норме кетоновые тела присутствуют в крови в небольших количествах. Избыточное их образование при тяжелом диабете ведет к повышению их содержания в крови (кетонемия) и в моче (кетонурия) – это состояние обозначают термином «кетоз».
Белки. Всасывание. При переваривании белков пищеварительными ферментами образуется смесь из аминокислот и небольших пептидов, содержащих от двух до десяти остатков аминокислот. Эти продукты всасываются слизистой кишечника, и здесь гидролиз завершается – пептиды также распадаются до аминокислот. Поступившие в кровь аминокислоты смешиваются с находящимися здесь такими же аминокислотами. В крови содержится смесь из аминокислот, поступивших из кишечника, образовавшихся при распаде тканевых белков и синтезированных организмом заново.
Синтез. В тканях непрерывно идет распад белков и их новообразование. Содержащиеся в крови аминокислоты избирательно поглощаются тканями как исходный материал для построения белков, а из тканей в кровь поступают другие аминокислоты. Синтезу и распаду подвергаются не только структурные белки, но и белки плазмы крови, а также белковые гормоны и ферменты.
Во взрослом организме аминокислоты или белки практически не запасаются, поэтому удаление аминокислот из крови происходит с такой же скоростью, как и их поступление из тканей в кровь. В растущем организме формируются новые ткани, и на этот процесс расходуется больше аминокислот, чем поступает в кровь за счет распада тканевых белков.
Печень участвует в метаболизме белков самым активным образом. Здесь синтезируются белки плазмы крови – альбумины и глобулины – а также собственные ферменты печени. Так, при потере плазменных белков содержание альбумина в плазме восстанавливается – за счет интенсивного синтеза – довольно быстро. Аминокислоты в печени используются не только для образования белков, но подвергаются также расщеплению, в ходе которого извлекается заключенная в них энергия.
Превращения и использование. Если аминокислоты используются в качестве источника энергии, то отщепляемая от них аминогруппа (–Nh3) направляется на образование мочевины, а не содержащий азота остаток молекулы окисляется приблизительно так же, как глюкоза или жирные кислоты.
Так называемый «орнитиновый цикл» описывает, как происходит превращение аммиака в мочевину. В этом цикле аминогруппа, отщепившаяся от аминокислоты в форме аммиака, присоединяется вместе с диоксидом углерода к молекуле орнитина с образованием цитруллина. Цитруллин присоединяет второй атом азота, на этот раз от аспарагиновой кислоты, и превращается в аргинин. Далее аргинин подвергается гидролизу с образованием мочевины и орнитина. Орнитин может теперь вновь вступить в цикл, а мочевина выводится из организма через почки как один из конечных продуктов метаболизма.
Список литературы
Ленинджер А. Основы биохимии, тт. 1–3. М., 1985
Страйер Л. Биохимия, тт. 1–3. М., 1985
Марри Р., Греннер Д., Мейес П., Родуэлл В. Биохимия человека, тт. 1–2. М., 1993
Албертс Б., Брей Д., Льюс Д. и др. Молекулярная биология клетки, тт. 1–3. М., 1994
www.referatmix.ru
Реферат: Метаболизм
Метаболизм, или обмен веществ, химические превращения, протекающие от момента поступления питательных веществ в живой организм до момента, когда конечные продукты этих превращений выделяются во внешнюю среду. К метаболизму относятся все реакции, в результате которых строятся структурные элементы клеток и тканей, и процессы, в которых из содержащихся в клетках веществ извлекается энергия. Иногда для удобства рассматривают по отдельности две стороны метаболизма – анаболизм и катаболизм, т.е. процессы созидания органических веществ и процессы их разрушения. Анаболические процессы обычно связаны с затратой энергии и приводят к образованию сложных молекул из более простых, катаболические же сопровождаются высвобождением энергии и заканчиваются образованием таких конечных продуктов (отходов) метаболизма, как мочевина, диоксид углерода, аммиак и вода.
Термин «обмен веществ» вошел в повседневную жизнь с тех пор, как врачи стали связывать избыточный или недостаточный вес, чрезмерную нервозность или, наоборот, вялость больного с повышенным или пониженным обменом. Для суждения об интенсивности метаболизма ставят тест на «основной обмен». Основной обмен – это показатель способности организма вырабатывать энергию. Тест проводят натощак в состоянии покоя; измеряют поглощение кислорода (О2) и выделение диоксида углерода (СО2). Сопоставляя эти величины, определяют, насколько полно организм использует («сжигает») питательные вещества. На интенсивность метаболизма влияют гормоны щитовидной железы, поэтому врачи при диагностике заболеваний, связанных с нарушениями обмена, в последнее время все чаще измеряют уровень этих гормонов в крови.
Методы исследования. При изучении метаболизма какого-нибудь одного из питательных веществ прослеживают все его превращения от той формы, в какой оно поступает в организм, до конечных продуктов, выводимых из организма. В таких исследованиях применяется крайне разнообразный набор биохимических методов.
Использование интактных животных или органов. Животному вводят изучаемое соединение, а затем в его моче и экскрементах определяют возможные продукты превращений (метаболиты) этого вещества. Более определенную информацию можно получить, исследуя метаболизм определенного органа, например печени или мозга. В этих случаях вещество вводят в соответствующий кровеносный сосуд, а метаболиты определяют в крови, оттекающей от данного органа.
Поскольку такого рода процедуры сопряжены с большими трудностями, часто для исследования используют тонкие срезы органов. Их инкубируют при комнатной температуре или при температуре тела в растворах с добавкой того вещества, метаболизм которого изучают. Клетки в таких препаратах не повреждены, и так как срезы очень тонкие, вещество легко проникает в клетки и легко выходит из них. Иногда затруднения возникают из-за слишком медленного прохождения вещества сквозь клеточные мембраны. В этих случаях ткани измельчают, чтобы разрушить мембраны, и с изучаемым веществом инкубируют клеточную кашицу. Именно в таких опытах было показано, что все живые клетки окисляют глюкозу до СО2 и воды и что только ткань печени способна синтезировать мочевину.
Использование клеток. Даже клетки представляют собой очень сложно организованные системы. В них имеется ядро, а в окружающей его цитоплазме находятся более мелкие тельца, т.н. органеллы, различных размеров и консистенции. С помощью соответствующей методики ткань можно «гомогенизировать», а затем подвергнуть дифференциальному центрифугированию (разделению) и получить препараты, содержащие только митохондрии, только микросомы или прозрачную жидкость – цитоплазму. Эти препараты можно по отдельности инкубировать с тем соединением, метаболизм которого изучается, и таким путем установить, какие именно субклеточные структуры участвуют в его последовательных превращениях. Известны случаи, когда начальная реакция протекает в цитоплазме, ее продукт подвергается превращению в микросомах, а продукт этого превращения вступает в новую реакцию уже в митохондриях. Инкубация изучаемого вещества с живыми клетками или с гомогенатом ткани обычно не выявляет отдельные этапы его метаболизма, и только последовательные эксперименты, в которых для инкубации используются те или иные субклеточные структуры, позволяют понять всю цепочку событий.
Использование радиоактивных изотопов. Для изучения метаболизма какого-либо вещества необходимы: 1) соответствующие аналитические методы для определения этого вещества и его метаболитов; и 2) методы, позволяющие отличать добавленное вещество от того же вещества, уже присутствующего в данном биологическом препарате. Эти требования служили главным препятствием при изучении метаболизма до тех пор, пока не были открыты радиоактивные изотопы элементов и в первую очередь радиоактивный углерод 14C. С появлением соединений, «меченных» 14C, а также приборов для измерения слабой радиоактивности эти трудности были преодолены. Если к биологическому препарату, например к суспензии митохондрий, добавляют меченную 14C жирную кислоту, то никаких специальных анализов для определения продуктов ее превращений не требуется; чтобы оценить скорость ее использования, достаточно просто измерять радиоактивность последовательно получаемых митохондриальных фракций. Эта же методика позволяет легко отличать молекулы радиоактивной жирной кислоты, введенной экспериментатором, от молекул жирной кислоты, уже присутствовавших в митохондриях к началу эксперимента.
Хроматография и электрофорез. В дополнение к вышеупомянутым требованиям биохимику необходимы и методы, позволяющие разделять смеси, состоящие из малых количеств органических веществ. Важнейший из них – хроматография, в основе которой лежит феномен адсорбции. Разделение компонентов смеси проводят при этом либо на бумаге, либо путем адсорбции на сорбенте, которым заполняют колонки (длинные стеклянные трубки), с последующей постепенной элюцией (вымыванием) каждого из компонентов.
Разделение методом электрофореза зависит от знака и числа зарядов ионизированных молекул. Электрофорез проводят на бумаге или на каком-нибудь инертном (неактивном) носителе, таком, как крахмал, целлюлоза или каучук.
Высокочувствительный и эффективный метод разделения – газовая хроматография. Им пользуются в тех случаях, когда подлежащие разделению вещества находятся в газообразном состоянии или могут быть в него переведены.
Выделение ферментов. Последнее место в описываемом ряду – животное, орган, тканевой срез, гомогенат и фракция клеточных органелл – занимает фермент, способный катализировать определенную химическую реакцию. Выделение ферментов в очищенном виде – важный раздел в изучении метаболизма.
Сочетание перечисленных методов позволило проследить главные метаболические пути у большей части организмов (в том числе у человека), установить, где именно эти различные процессы протекают, и выяснить последовательные этапы главных метаболических путей. К настоящему времени известны тысячи отдельных биохимических реакций, изучены участвующие в них ферменты.
Клеточный метаболизм. Живая клетка – это высокоорганизованная система. В ней имеются различные структуры, а также ферменты, способные их разрушить. Содержатся в ней и крупные макромолекулы, которые могут распадаться на более мелкие компоненты в результате гидролиза (расщепления под действием воды). В клетке обычно много калия и очень мало натрия, хотя клетка существует в среде, где натрия много, а калия относительно мало, и клеточная мембрана легко проницаема для обоих ионов. Следовательно, клетка – это химическая система, весьма далекая от равновесия. Равновесие наступает только в процессе посмертного автолиза (самопереваривания под действием собственных ферментов).
Потребность в энергии. Чтобы удержать систему в состоянии, далеком от химического равновесия, требуется производить работу, а для этого необходима энергия. Получение этой энергии и выполнение этой работы – непременное условие для того, чтобы клетка оставалась в своем стационарном (нормальном) состоянии, далеком от равновесия. Одновременно в ней выполняется и иная работа, связанная со взаимодействием со средой, например: в мышечных клетках – сокращение; в нервных клетках – проведение нервного импульса; в клетках почек – образование мочи, значительно отличающейся по своему составу от плазмы крови; в специализированных клетках желудочно-кишечного тракта – синтез и выделение пищеварительных ферментов; в клетках эндокринных желез – секреция гормонов; в клетках светляков – свечение; в клетках некоторых рыб – генерирование электрических разрядов и т.д.
Источники энергии. В любом из перечисленных выше примеров непосредственным источником энергии, которую клетка использует для производства работы, служит энергия, заключенная в структуре аденозинтрифосфата (АТФ). В силу особенностей своей структуры это соединение богато энергией, и разрыв связей между его фосфатными группами может происходить таким образом, что высвобождающаяся энергия используется для производства работы. Однако энергия не может стать доступной для клетки при простом гидролитическом разрыве фосфатных связей АТФ: в этом случае она расходуется впустую, выделяясь в виде тепла. Процесс должен состоять из двух последовательных этапов, в каждом из которых участвует промежуточный продукт, обозначенный здесь X–Ф (в приведенных уравнениях X и Y означают два разных органических вещества; Ф – фосфат; АДФ – аденозиндифосфат):
Поскольку практически для любого проявления жизнедеятельности клеток необходим АТФ, неудивительно, что метаболическая активность живых клеток направлена в первую очередь на синтез АТФ. Этой цели служат различные сложные последовательности реакций, в которых используется потенциальная химическая энергия, заключенная в молекулах углеводов и жиров (липидов).
Метаболизм углеводов и липидов
Синтез АТФ. Анаэробный (без участия кислорода). Главная роль углеводов и липидов в клеточном метаболизме состоит в том, что их расщепление на более простые соединения обеспечивает синтез АТФ. Несомненно, что те же процессы протекали и в первых, самых примитивных клетках. Однако в атмосфере, лишенной кислорода, полное окисление углеводов и жиров до CO2 было невозможно. У этих примитивных клеток имелись все же механизмы, с помощью которых перестройка структуры молекулы глюкозы обеспечивала синтез небольших количеств АТФ. Речь идет о процессах, которые у микроорганизмов называют брожением. Лучше всего изучено сбраживание глюкозы до этилового спирта и CO2 у дрожжей.
В ходе 11 последовательных реакций, необходимых для того, чтобы завершилось это превращение, образуется ряд промежуточных продуктов, представляющих собой эфиры фосфорной кислоты (фосфаты). Их фосфатная группа переносится на аденозиндифосфат (АДФ) с образованием АТФ. Чистый выход АТФ составляет 2 молекулы АТФ на каждую молекулу глюкозы, расщепленную в процессе брожения. Аналогичные процессы происходят во всех живых клетках; поскольку они поставляют необходимую для жизнедеятельности энергию, их иногда (не вполне корректно) называют анаэробным дыханием клеток.
У млекопитающих, в том числе у человека, такой процесс называется гликолизом и его конечным продуктом является молочная кислота, а не спирт и CO2. Вся последовательность реакций гликолиза, за исключением двух последних этапов, полностью идентична процессу, протекающему в дрожжевых клетках.
Аэробный (с использованием кислорода). С появлением в атмосфере кислорода, источником которого послужил, очевидно, фотосинтез растений, в ходе эволюции развился механизм, обеспечивающий полное окисление глюкозы до CO2 и воды, – аэробный процесс, в котором чистый выход АТФ составляет 38 молекул АТФ на каждую окисленную молекулу глюкозы. Этот процесс потребления клетками кислорода для образования богатых энергией соединений известен как клеточное дыхание (аэробное). В отличие от анаэробного процесса, осуществляемого ферментами цитоплазмы, окислительные процессы протекают в митохондриях. В митохондриях пировиноградная кислота – промежуточный продукт, образовавшийся в анаэробной фазе – окисляется до СО2 в шести последовательных реакциях, в каждой из которых пара электронов переносится на общий акцептор – кофермент никотинамидадениндинуклеотид (НАД). Эту последовательность реакций называют циклом трикарбоновых кислот, циклом лимонной кислоты или циклом Кребса. Из каждой молекулы глюкозы образуется 2 молекулы пировиноградной кислоты; 12 пар электронов отщепляется от молекулы глюкозы в ходе ее окисления, описываемого уравнением:
Перенос электронов. В каждой митохондрии имеется механизм, посредством которого восстановленный НАД (НАДН, где Н – водород), образовавшийся в цикле трикарбоновых кислот, передает свою пару электронов кислороду. Перенос, однако, не происходит напрямую. Электроны как бы передаются «из рук в руки» и, лишь пройдя цепь переносчиков, присоединяются к кислороду. Эта «цепь переноса электронов» состоит из следующих компонентов:
НАДНН Флавинадениндинклеотид Кофермент Q
Цитохром b Цитохром c Цитохром a O2
Все компоненты этой системы, находящиеся в митохондриях, фиксированы в пространстве и сцеплены друг с другом. Такое их состояние облегчает перенос электронов.
В состав НАД входит никотиновая кислота (витамин ниацин), а в состав флавинадениндинуклеотида – рибофлавин (витамин B2). Кофермент Q представляет собой высокомолекулярный хинон, синтезируемый в печени, а цитохромы – это три разных белка, каждый из которых, подобно гемоглобину, содержит гемогруппу.
В цепи переноса электронов на каждую пару электронов, перенесенную от НАДН на O2, синтезируется 3 молекулы АТФ. Поскольку от каждой молекулы глюкозы отщепляются и передаются молекулам НАД 12 пар электронов, в общей сложности на каждую молекулу глюкозы образуется 312 = 36 молекул АТФ. Этот процесс образования АТФ в ходе окисления называется окислительным фосфорилированием.
Липиды как источник энергии. Жирные кислоты могут использоваться в качестве источника энергии приблизительно так же, как и углеводы. Окисление жирных кислот протекает путем последовательного отщепления от молекулы жирной кислоты двууглеродного фрагмента с образованием ацетилкофермента A (ацетил-КоА) и одновременной передачей двух пар электронов в цепь переноса электронов. Образовавшийся ацетил-КоА – нормальный компонент цикла трикарбоновых кислот, и в дальнейшем его судьба не отличается от судьбы ацетил-КоА, поставляемого углеводным обменом. Таким образом, механизмы синтеза АТФ при окислении как жирных кислот, так и метаболитов глюкозы практически одинаковы.
Если организм животного получает энергию почти целиком за счет одного только окисления жирных кислот, а это бывает, например, при голодании или при сахарном диабете, то скорость образования ацетил-КоА превышает скорость его окисления в цикле трикарбоновых кислот. В этом случае лишние молекулы ацетил-КоА реагируют друг с другом, в результате чего образуются в конечном счете ацетоуксусная и -гидроксимасляная кислоты. Их накопление является причиной патологического состояния, т.н. кетоза (одного из видов ацидоза), который при тяжелом диабете может вызвать кому и смерть.
Запасание энергии. Животные питаются нерегулярно, и их организму нужно как-то запасать заключенную в пище энергию, источником которой являются поглощенные животным углеводы и жиры. Жирные кислоты могут запасаться в виде нейтральных жиров либо в печени, либо в жировой ткани. Углеводы, поступая в большом количестве, в желудочно-кишечном тракте гидролизуются до глюкозы или иных сахаров, которые затем в печени превращаются в ту же глюкозу. Здесь из глюкозы синтезируется гигантский полимер гликоген путем присоединения друг к другу остатков глюкозы с отщеплением молекул воды (число остатков глюкозы в молекулах гликогена доходит до 30 000). Когда возникает потребность в энергии, гликоген вновь распадается до глюкозы в реакции, продуктом которой является глюкозофосфат. Этот глюкозофосфат направляется на путь гликолиза – процесса, составляющего часть пути окисления глюкозы. В печени глюкозофосфат может также подвергнуться гидролизу, и образующаяся глюкоза поступает в кровоток и доставляется кровью к клеткам в разных частях тела.
Синтез липидов из углеводов. Если количество углеводов, поглощенных с пищей за один прием, больше того, какое может быть запасено в виде гликогена, то избыток углеводов превращается в жиры. Начальная последовательность реакций совпадает при этом с обычным окислительным путем, т.е. сначала из глюкозы образуется ацетил-КоА, но далее этот ацетил-КоА используется в цитоплазме клетки для синтеза длинноцепочечных жирных кислот. Процесс синтеза можно описать как обращение обычного процесса окисления жирных клеток. Затем жирные кислоты запасаются в виде нейтральных жиров (триглицеридов), отлагающихся в разных частях тела. Когда требуется энергия, нейтральные жиры подвергаются гидролизу и жирные кислоты поступают в кровь. Здесь они адсорбируются молекулами плазменных белков (альбуминов и глобулинов) и затем поглощаются клетками самых разных типов. Механизмов, способных осуществлять синтез глюкозы из жирных кислот, у животных нет, но у растений такие механизмы имеются.
Метаболизм липидов. Липиды попадают в организм главным образом в форме триглицеридов жирных кислот. В кишечнике под действием ферментов поджелудочной железы они подвергаются гидролизу, продукты которого всасываются клетками стенки кишечника. Здесь из них вновь синтезируются нейтральные жиры, которые через лимфатическую систему поступают в кровь и либо транспортируются в печень, либо отлагаются в жировой ткани. Выше уже указывалось, что жирные кислоты могут также синтезироваться заново из углеводных предшественников. Следует отметить, что, хотя в клетках млекопитающих может происходить включение одной двойной связи в молекулы длинноцепочечных жирных кислот (между С–9 и С–10), включать вторую и третью двойную связь эти клетки неспособны. Поскольку жирные кислоты с двумя и тремя двойными связями играют важную роль в метаболизме млекопитающих, они в сущности являются витаминами. Поэтому линолевую (C18:2) и линоленовую (C18:3) кислоты называют незаменимыми жирными кислотами. В то же время в клетках млекопитающих в линоленовую кислоту может включаться четвертая двойная связь и путем удлинения углеродной цепи может образоваться арахидоновая кислота (C20:4), также необходимый участник метаболических процессов.
В процессе синтеза липидов остатки жирных кислот, связанные с коферментом А (ацил-КоА), переносятся на глицерофосфат – эфир фосфорной кислоты и глицерина. В результате образуется фосфатидная кислота – соединение, в котором одна гидроксильная группа глицерина этерифицирована фосфорной кислотой, а две группы – жирными кислотами. При образовании нейтральных жиров фосфорная кислота удаляется путем гидролиза, и ее место занимает третья жирная кислота в результате реакции с ацил-КоА. Кофермент А образуется из пантотеновой кислоты (одного из витаминов). В его молекуле имеется сульфгидрильная (– SH) группа, способная реагировать с кислотами с образованием тиоэфиров. При образовании фосфолипидов фосфатидная кислота реагирует непосредственно с активированным производным одного из азотистых оснований, таких, как холин, этаноламин или серин.
За исключением витамина D, все встречающиеся в организме животных стероиды (производные сложных спиртов) легко синтезируются самим организмом. Сюда относятся холестерин (холестерол), желчные кислоты, мужские и женские половые гормоны и гормоны надпочечников. В каждом случае исходным материалом для синтеза служит ацетил-КоА: из ацетильных групп путем многократно повторяющейся конденсации строится углеродный скелет синтезируемого соединения.
Метаболизм белков
Синтез аминокислот. Растения и большинство микроорганизмов могут жить и расти в среде, в которой для их питания имеются только минеральные вещества, диоксид углерода и вода. Это значит, что все обнаруживаемые в них органические вещества эти организмы синтезируют сами. Встречающиеся во всех живых клетках белки построены из 21 вида аминокислот, соединенных в различной последовательности. Аминокислоты синтезируются живыми организмами. В каждом случае ряд химических реакций приводит к образованию -кетокислоты. Одна такая -кетокислота, а именно -кетоглутаровая (обычный компонент цикла трикарбоновых кислот), участвует в связывании азота по следующему уравнению:
-Кетоглутаровая кислота + Nh4 + НАДН
Глутаминовая кислота + НАД.
Азот глутаминовой кислоты может быть затем передан любой из других -кетокислот с образованием соответствующей аминокислоты.
Организм человека и большинства других животных сохранил способность синтезировать все аминокислоты за исключением девяти т.н. незаменимых аминокислот. Поскольку кетокислоты, соответствующие этим девяти, не синтезируются, незаменимые аминокислоты должны поступать с пищей.
Синтез белков. Аминокислоты нужны для биосинтеза белка. Процесс биосинтеза протекает обычно следующим образом. В цитоплазме клетки каждая аминокислота «активируется» в реакции с АТФ, а затем присоединяется к концевой группе молекулы рибонуклеиновой кислоты, специфичной именно для данной аминокислоты. Эта сложная молекула связывается с небольшим тельцем, т.н. рибосомой, в положении, определяемом более длинной молекулой рибонуклеиновой кислоты, прикрепленной к рибосоме. После того как все эти сложные молекулы соответствующим образом выстроились, связи между исходной аминокислотой и рибонуклеиновой кислотой разрываются и возникают связи между соседними аминокислотами – синтезируется специфичный белок. Процесс биосинтеза поставляет белки не только для роста организма или для секреции в среду. Все белки живых клеток со временем претерпевают распад до составляющих их аминокислот, и для поддержания жизни клетки должны синтезироваться вновь.
Синтез других азотсодержащих соединений. В организме млекопитающих аминокислоты используются не только для биосинтеза белков, но и как исходный материал для синтеза многих азотсодержащих соединений. Аминокислота тирозин является предшественником гормонов адреналина и норадреналина. Простейшая аминокислота глицин служит исходным материалом для биосинтеза пуринов, входящих в состав нуклеиновых кислот, и порфиринов, входящих в состав цитохромов и гемоглобина. Аспарагиновая кислота – предшественник пиримидинов нуклеиновых кислот. Метильная группа метионина передается ряду других соединений в ходе биосинтеза креатина, холина и саркозина. При биосинтезе креатина от одного соединения к другому передается также и гуанидиновая группировка аргинина. Триптофан служит предшественником никотиновой кислоты, а из валина в растениях синтезируется такой витамин, как пантотеновая кислота. Все это лишь отдельные примеры использования аминокислот в процессах биосинтеза.
Азот, поглощаемый микроорганизмами и высшими растениями в виде иона аммония, расходуется почти целиком на образование аминокислот, из которых затем синтезируются многие азотсодержащие соединения живых клеток. Избыточных количеств азота ни растения, ни микроорганизмы не поглощают. В отличие от них, у животных количество поглощенного азота зависит от содержащихся в пище белков. Весь азот, поступивший в организм в виде аминокислот и не израсходованный в процессах биосинтеза, довольно быстро выводится из организма с мочой. Происходит это следующим образом. В печени неиспользованные аминокислоты передают свой азот -кетоглутаровой кислоте с образованием глутаминовой кислоты, которая дезаминируется, высвобождая аммиак. Далее азот аммиака может либо на время запасаться путем синтеза глутамина, либо сразу же использоваться для синтеза мочевины, протекающего в печени.
У глутамина есть и другая роль. Он может подвергаться гидролизу в почках с высвобождением аммиака, который поступает в мочу в обмен на ионы натрия. Этот процесс крайне важен как средство поддержания кислотно-щелочного равновесия в организме животного. Почти весь аммиак, происходящий из аминокислот и, возможно, из других источников, превращается в печени в мочевину, так что свободного аммиака в крови обычно почти нет. Однако при некоторых условиях довольно значительные количества аммиака содержит моча. Этот аммиак образуется в почках из глутамина и переходит в мочу в обмен на ионы натрия, которые таким образом реадсорбируются и задерживаются в организме. Этот процесс усиливается при развитии ацидоза – состояния, при котором организм нуждается в дополнительных количествах катионов натрия для связывания избытка ионов бикарбоната в крови.
Избыточные количества пиримидинов тоже распадаются в печени через ряд реакций, в которых высвобождается аммиак. Что касается пуринов, то их избыток подвергается окислению с образованием мочевой кислоты, выделяющейся с мочой у человека и других приматов, но не у остальных млекопитающих. У птиц отсутствует механизм синтеза мочевины, и именно мочевая кислота, а не мочевина, является у них конечным продуктом обмена всех азотсодержащих соединений.
Общие представления о метаболизме органических веществ
Можно сформулировать некоторые общие понятия, или «правила», касающиеся метаболизма. Приведенные ниже несколько главных «правил» позволяют лучше понять, как протекает и регулируется метаболизм.
1. Метаболические пути необратимы. Распад никогда не идет по пути, который являлся бы простым обращением реакций синтеза. В нем участвуют другие ферменты и другие промежуточные продукты. Нередко противоположно направленные процессы протекают в разных отсеках клетки. Так, жирные кислоты синтезируются в цитоплазме при участии одного набора ферментов, а окисляются в митохондриях при участии совсем другого набора.
2. Ферментов в живых клетках достаточно для того, чтобы все известные метаболические реакции могли протекать гораздо быстрее, чем это обычно наблюдается в организме. Следовательно, в клетках существуют какие-то регуляторные механизмы. Открыты разные типы таких механизмов.
а) Фактором, ограничивающим скорость метаболических превращений данного вещества, может быть поступление этого вещества в клетку; именно на этот процесс в таком случае и направлена регуляция. Роль инсулина, например, связана с тем, что он, по-видимому, облегчает проникновение глюкозы во все клетки, глюкоза же подвергается превращениям с той скоростью, с какой она поступает. Сходным образом проникновение железа и кальция из кишечника в кровь зависит от процессов, скорость которых регулируется.
б) Вещества далеко не всегда могут свободно переходить из одного клеточного отсека в другой; есть данные, что внутриклеточный перенос регулируется некоторыми стероидными гормонами.
в) Выявлено два типа сервомеханизмов «отрицательной обратной связи».
У бактерий были обнаружены примеры того, что присутствие продукта какой-нибудь последовательности реакций, например аминокислоты, подавляет биосинтез одного из ферментов, необходимых для образования этой аминокислоты.
В каждом случае фермент, биосинтез которого оказывается затронутым, был ответствен за первый «определяющий» этап (на схеме реакция 4) метаболического пути, ведущего к синтезу данной аминокислоты.
Второй механизм хорошо изучен у млекопитающих. Это простое ингибирование конечным продуктом (в нашем случае – аминокислотой) фермента, ответственного за первый «определяющий» этап метаболического пути.
Еще один тип регулирования посредством обратной связи действует в тех случаях, когда окисление промежуточных продуктов цикла трикарбоновых кислот сопряжено с образованием АТФ из АДФ и фосфата в процессе окислительного фосфорилирования. Если весь имеющийся в клетке запас фосфата и (или) АДФ уже исчерпан, то окисление приостанавливается и может возобновиться лишь после того, как этот запас вновь станет достаточным. Таким образом, окисление, смысл которого в том, чтобы поставлять полезную энергию в форме АТФ, происходит только тогда, когда возможен синтез АТФ.
3. В биосинтетических процессах участвует сравнительно небольшое число строительных блоков, каждый из которых используется для синтеза многих соединений. Среди них можно назвать ацетилкофермент А, глицерофосфат, глицин, карбамилфосфат, поставляющий карбамильную (h3N–CO–) группу, производные фолиевой кислоты, служащие источником гидроксиметильной и формильной групп, S-аденозилметионин – источник метильных групп, глутаминовую и аспарагиновую кислоты, поставляющие аминогруппы, и наконец, глутамин – источник амидных групп. Из этого относительно небольшого числа компонентов строятся все те разнообразные соединения, которые мы находим в живых организмах.
4. Простые органические соединения редко участвуют в метаболических реакциях непосредственно. Обычно они должны быть сначала «активированы» путем присоединения к одному из ряда соединений, универсально используемых в метаболизме. Глюкоза, например, может подвергнуться окислению лишь после того, как она будет этерифицирована фосфорной кислотой, для прочих же своих превращений она должна быть этерифицирована уридиндифосфатом. Жирные кислоты не могут быть вовлечены в метаболические превращения прежде, чем они образуют эфиры с коферментом А. Каждый из этих активаторов либо родствен одному из нуклеотидов, входящих в состав рибонуклеиновой кислоты, либо образуется из какого-нибудь витамина. Легко понять в связи с этим, почему витамины требуются в таких небольших количествах. Они расходуются на образование «коферментов», а каждая молекула кофермента на протяжении жизни организма используется многократно, в отличие от основных питательных веществ (например, глюкозы), каждая молекула которых используется только один раз.
В заключение следует сказать, что термин «метаболизм», означавший ранее нечто не более сложное, чем просто использование углеводов и жиров в организме, теперь применяется для обозначения тысяч ферментативных реакций, вся совокупность которых может быть представлена как огромная сеть метаболических путей, многократно пересекающихся (из-за наличия общих промежуточных продуктов) и управляемых очень тонкими регуляторными механизмами.
Метаболизм минеральных веществ
Относительное содержание. Различные элементы, встречающиеся в живых организмах, перечислены ниже в убывающем порядке в зависимости от их относительного содержания: 1) кислород, углерод, водород и азот; 2) кальций, фосфор, калий и сера; 3) натрий, хлор, магний и железо; 4) марганец, медь, молибден, селен, йод и цинк; 5) алюминий, фтор, кремний и литий; 6) бром, мышьяк, свинец и, возможно, некоторые другие.
Кислород, углерод, водород и азот – это те элементы, из которых построены мягкие ткани тела. Они входят в состав таких соединений, как углеводы, липиды, белки, вода, диоксид углерода и аммиак. Элементы, перечисленные в пп. 2 и 3, находятся в организме обычно в виде одного или нескольких неорганических соединений, а элементы пп. 4, 5 и 6 присутствуют только в следовых количествах и потому их называют микроэлементами.
Распределение в организме. Кальций. Кальций присутствует главным образом в костной ткани и в зубах, преимущественно в виде фосфата и в небольших количествах в виде карбоната и фторида. Поступающий с пищей кальций всасывается в основном в верхних отделах кишечника, имеющих слабокислую реакцию. Этому всасыванию (у человека здесь всасывается всего 20–30% кальция пищи) способствует витамин D. Под действием витамина D клетки кишечника вырабатывают особый белок, который связывает кальций и облегчает его перенос через стенку кишечника в кровь. На всасывание влияет также присутствие некоторых других веществ, в особенности фосфата и оксалата, которые в малых количествах способствуют всасыванию, а в больших, наоборот, подавляют его.
В крови около половины кальция связано с белком, остальное составляют ионы кальция. Соотношение ионизированной и неионизированной форм зависит от общей концентрации кальция в крови, а также от содержания белка и фосфата и концентрации водородных ионов (рН крови). Доля неионизированного кальция, на которую влияет уровень белка, позволяет косвенным образом судить о качестве питания и об эффективности работы печени, в которой идет синтез плазменных белков.
На количество ионизированного кальция влияют, с одной стороны, витамин D и факторы, воздействующие на всасывание, а с другой – паратиреоидный гормон и, возможно, также витамин D, поскольку оба эти вещества регулируют как скорость отложения кальция в костной ткани, так и его мобилизацию, т.е. вымывание из костей. Избыток паратиреоидного гормона стимулирует выход кальция из костной ткани, что приводит к повышению его концентрации в плазме. Изменяя скорости всасывания и экскреции кальция и фосфата, а также скорости образования костной ткани и ее разрушения, эти механизмы строго контролируют концентрацию кальция и фосфата в сыворотке крови. Ионы кальция играют регулирующую роль во многих физиологических процессах, в том числе в нервных реакциях, мышечном сокращении, свертывании крови. Выведение кальция из организма происходит в норме в основном (на 2/3) через желчь и кишечник и в меньшей степени (1/3) – через почки.
Фосфор. Метаболизм фосфора – одного из главных компонентов костной ткани и зубов – во многом зависит от тех же факторов, что и метаболизм кальция. Фосфор в виде фосфата присутствует в организме также в сотнях различных физиологически важных органических эфиров. Паратиреоидный гормон стимулирует выведение фосфора с мочой и выход его из костной ткани; тем самым он регулирует концентрацию фосфора в плазме крови.
Натрий. Натрий – главный катион внеклеточной жидкости – вместе с белком, хлоридом и бикарбонатом играет важнейшую роль в регулировании осмотического давления и pH (концентрации водородных ионов) крови. В клетках, напротив, содержится очень мало натрия, так как они обладают механизмом для выведения ионов натрия и удержания ионов калия. Весь натрий, превышающий потребности организма, очень быстро выводится через почки.
Поскольку во всех процессах выделения натрий теряется, он должен постоянно поступать в организм с пищей. При ацидозе, когда необходимо, чтобы из организма выводились большие количества анионов (например, хлорида или ацетоацетата), почки предотвращают чрезмерную потерю натрия благодаря образованию аммиака из глутамина. Выведение натрия через почки регулируется гормоном коры надпочечников альдостероном. Под действием этого гормона в кровь возвращается достаточно натрия для поддержания нормального осмотического давления и нормального объема внеклеточной жидкости.
Суточная потребность в хлористом натрии составляет 5–10 г. Эта величина возрастает при поглощении больших количеств жидкости, когда усиливается потоотделение и выделяется больше мочи.
Калий. В отличие от натрия, калий содержится в клетках в больших количествах, но во внеклеточной жидкости его мало. Главная функция калия – регулирование внутриклеточного осмотического давления и поддержание кислотно-щелочного равновесия. Он также играет важную роль в проведении нервного импульса и во многих ферментных системах, в том числе и в тех, которые участвуют в мышечном сокращении. Калий широко распространен в природе, и его много в любой пище, так что спонтанно калиевая недостаточность возникнуть не может. В плазме концентрация калия регулируется альдостероном, стимулирующим его экскрецию с мочой.
Сера. С пищей сера поступает в организм главным образом в составе двух аминокислот – цистина и метионина. На конечных этапах метаболизма этих аминокислот сера высвобождается и в результате окисления переводится в неорганическую форму. В составе цистина и метионина сера присутствует в структурных белках. Важную роль играет также сульфгидрильная (–SH) группа цистеина, от которой зависит активность многих ферментов.
Большая часть серы выводится с мочой в виде сульфата. Небольшое количество экскретируемого сульфата обычно связано с органическими соединениями типа фенолов.
Магний. Метаболизм магния сходен с метаболизмом кальция, и в виде комплекса с фосфатом этот элемент тоже входит в состав костной ткани. Магний присутствует во всех живых клетках, где он функционирует как необходимый компонент многих ферментных систем; эта его роль была убедительно продемонстрирована на примере углеводного обмена в мышцах. Магний, как и калий, широко распространен, и вероятность возникновения его недостаточности очень мала.
Железо. Железо входит в состав гемоглобина и других гемопротеинов, а именно миоглобина (мышечного гемоглобина), цитохромов (дыхательных ферментов) и каталазы, а также в состав некоторых ферментов, не содержащих гемогруппы. Всасывается железо в верхних отделах кишечника, причем это единственный элемент, всасывающийся только тогда, когда его запас в организме полностью исчерпан. В плазме железо транспортируется в соединении с белком (трансферрином). Через почки железо не выводится; избыток его накапливается в печени в соединении с особым белком (ферритином).
Микроэлементы. У каждого микроэлемента, присутствующего в организме, своя особая функция, связанная с тем, что он стимулирует действие того или иного фермента или как-либо иначе на него влияет. Цинк необходим для кристаллизации инсулина; кроме того, он является компонентом карбоангидразы (фермента, участвующего в транспорте диоксида углерода) и некоторых других ферментов. Молибден и медь – тоже необходимые компоненты различных ферментов. Иод требуется для синтеза трииодтиронина, гормона щитовидной железы. Фтор (входящий в состав зубной эмали) способствует предотвращению кариеса.
Использование метаболитов
Углеводы. Всасывание. Моносахариды, или простые сахара, высвобождающиеся при переваривании углеводов пищи, переходят из кишечника в кровоток в результате процесса, называемого всасыванием. Механизм всасывания представляет собой сочетание простой диффузии и химической реакции (активного всасывания). Одна из гипотез, касающихся природы химической фазы процесса, предполагает, что в этой фазе моносахариды соединяются с фосфорной кислотой в реакции, катализируемой ферментом из группы киназ, после чего проникают в кровеносные сосуды и здесь высвобождаются в результате ферментативного дефосфорилирования (разрыва фосфатной связи), катализируемого одной из фосфатаз. Именно активным всасыванием объясняется то, что разные моносахариды всасываются с разной скоростью и что углеводы всасываются даже тогда, когда уровень сахара в крови выше, чем в кишечнике, т.е. в условиях, когда естественно было бы ожидать их перемещения в обратном направлении – из крови в кишечник.
Механизмы гомеостаза. Поступающие в кровоток моносахариды повышают уровень сахара в крови. При голодании концентрация глюкозы в крови колеблется обычно от 70 до 100 мг на 100 мл крови. Этот уровень поддерживается с помощью механизмов, называемых механизмами гомеостаза (самостабилизации). Как только уровень сахара в крови в результате всасывания из кишечника повышается, в действие вступают процессы, выводящие сахар из крови, так что уровень его колеблется не слишком сильно.
Подобно глюкозе, все прочие моносахариды поступают из кровотока в печень, где превращаются в глюкозу. Теперь они неотличимы как от глюкозы, которая всосалась, так и от той, что уже была в организме, и подвергаются тем же метаболическим превращениям. Один из механизмов гомеостаза углеводов, функционирующий в печени, – это гликогенез, посредством которого глюкоза переходит из крови в клетки, где превращается в гликоген. Гликоген хранится в печени до тех пор, пока не произойдет снижение уровня сахара в крови: в этой ситуации гомеостатический механизм вызовет распад накопленного гликогена до глюкозы, которая вновь поступит в кровь.
Превращения и использование. Поскольку кровь поставляет глюкозу во все ткани тела и все ткани используют ее для получения энергии, уровень глюкозы в крови снижается главным образом за счет ее использования.
В мышцах глюкоза крови превращается в гликоген. Однако мышечный гликоген не может быть использован для получения глюкозы, которая перешла бы в кровь. В нем заключен запас энергии, и скорость его использования зависит от мышечной активности. В мышечной ткани содержатся два соединения с большим запасом легко доступной энергии в форме богатых энергией фосфатных связей – креатинфосфат и аденозинтрифосфат (АТФ). При отщеплении от этих соединений их фосфатных групп высвобождается энергия для мышечного сокращения. Чтобы мышца вновь могла сокращаться, эти соединения должны быть восстановлены в своей исходной форме. Для этого требуется энергия, которую поставляет окисление продуктов распада гликогена. При мышечном сокращении гликоген превращается в глюкозофосфат, а затем – через ряд реакций – во фруктозодифосфат. Фруктозодифосфат распадается на два трехуглеродных соединения, из которых после ряда этапов образуется сначала пировиноградная кислота, а в конечном итоге – молочная кислота, как об этом уже говорилось при описании метаболизма углеводов. Это превращение гликогена в молочную кислоту, сопровождающееся высвобождением энергии, может происходить в отсутствие кислорода.
При недостатке кислорода молочная кислота накапливается в мышцах, диффундирует в кровоток и поступает в печень, где из нее вновь образуется гликоген. Если кислорода достаточно, то молочная кислота в мышцах не накапливается. Вместо этого она, как это описано выше, полностью окисляется через цикл трикарбоновых кислот до диоксида углерода и воды с образованием АТФ, который может быть использован для сокращения.
Метаболизм углеводов в нервной ткани и эритроцитах отличается от метаболизма в мышцах тем, что гликоген здесь не участвует. Однако и здесь промежуточными продуктами являются пировиноградная и молочная кислоты, образующиеся при расщеплении глюкозофосфата.
Глюкоза используется не только в клеточном дыхании, но и во многих других процессах: синтезе лактозы (молочного сахара), образовании жиров, а также особых сахаров, входящих в состав полисахаридов соединительной ткани и ряда других тканей.
Гликоген печени, синтезируемый при всасывании углеводов в кишечнике, служит самым доступным источником глюкозы, когда всасывание отсутствует. Если этот источник оказывается исчерпанным, в печени начинается процесс глюконеогенеза. Глюкоза образуется при этом из некоторых аминокислот (из 100 г белка образуется 58 г глюкозы) и нескольких других неуглеводных соединений, в том числе из глицериновых остатков нейтральных жиров.
Некоторую, хотя и не столь важную, роль в метаболизме углеводов играют почки. Они выводят из организма избыток глюкозы, когда ее концентрация в крови слишком высока; при меньших концентрациях глюкоза практически не выводится.
В регулировании метаболизма углеводов участвует несколько гормонов, в том числе гормоны поджелудочной железы, передней доли гипофиза и коры надпочечников.
Гормон поджелудочной железы инсулин снижает концентрацию глюкозы в крови и повышает ее концентрацию в клетках. По-видимому, он стимулирует также и запасание гликогена в печени. Кортикостерон, гормон коры надпочечников, и адреналин, вырабатываемый мозговым веществом надпочечников, воздействуют на метаболизм углеводов, стимулируя распад гликогена (главным образом в мышцах и печени) и синтез глюкозы (в печени).
Липиды. Всасывание. В кишечнике после переваривания жиров остаются главным образом свободные жирные кислоты с небольшой примесью холестерина и лецитина и следами жирорастворимых витаминов. Все эти вещества очень тонко диспергированы благодаря эмульгирующему и солюбилизирующему действию солей желчных кислот. Солюбилизирующее действие обычно связывают с образованием нестойких химических соединений между жирными кислотами и солями желчных кислот. Эти комплексы проникают в клетки эпителия тонкого кишечника и здесь распадаются на жирные кислоты и соли желчных кислот. Последние переносятся в печень и вновь секретируются с желчью, а жирные кислоты вступают в соединение с глицерином или холестерином. Образовавшиеся реконструированные жиры поступают в лимфатические сосуды брыжейки в форме млечного сока, т.н. «хилуса». Из сосудов брыжейки хилус по лимфатической системе через грудной проток поступает в кровеносную систему.
После переваривания пищи содержание липидов в крови возрастает приблизительно от 500 мг (уровень при голодании) до 1000 мг на 100 мл плазмы. Присутствующие в крови липиды представляют собой смесь жирных кислот, нейтральных жиров, фосфолипидов (лецитина и кефалина), холестерина и эфиров холестерина.
Распределение. Кровь доставляет липиды в разные ткани тела и прежде всего в печень. Печень обладает способностью модифицировать поступающие в нее жирные кислоты. Это особенно выражено у видов, запасающих жиры с высоким содержанием насыщенных или, наоборот, ненасыщенных жирных кислот: в печени этих животных соотношение насыщенных и ненасыщенных кислот изменяется таким образом, что отлагающийся жир по своему составу соответствует жиру, свойственному данному организму.
Жиры в печени либо используются для получения энергии, либо переходят в кровь и доставляются ею в разные ткани. Здесь они могут включаться в структурные элементы тканей, но большая их часть отлагается в жировых депо, где они хранятся до тех пор, пока не возникнет потребность в энергии; тогда они снова переносятся в печень и подвергаются здесь окислению.
Метаболизм липидов, как и углеводов, регулируется гомеостатически. Механизмы гомеостаза, воздействующие на липидный и углеводный обмен, видимо, тесно связаны, поскольку при замедлении метаболизма углеводов усиливается метаболизм липидов, и наоборот.
Превращения и использование. Четырехуглеродные кислоты – ацетоуксусная (продукт конденсации двух ацетатных единиц) и -гидроксимасляная – и трехуглеродное соединение ацетон, образующийся при отщеплении одного атома углерода от ацетоуксусной кислоты, известны под общим названием кетоновых (ацетоновых) тел. В норме кетоновые тела присутствуют в крови в небольших количествах. Избыточное их образование при тяжелом диабете ведет к повышению их содержания в крови (кетонемия) и в моче (кетонурия) – это состояние обозначают термином «кетоз».
Белки. Всасывание. При переваривании белков пищеварительными ферментами образуется смесь из аминокислот и небольших пептидов, содержащих от двух до десяти остатков аминокислот. Эти продукты всасываются слизистой кишечника, и здесь гидролиз завершается – пептиды также распадаются до аминокислот. Поступившие в кровь аминокислоты смешиваются с находящимися здесь такими же аминокислотами. В крови содержится смесь из аминокислот, поступивших из кишечника, образовавшихся при распаде тканевых белков и синтезированных организмом заново.
Синтез. В тканях непрерывно идет распад белков и их новообразование. Содержащиеся в крови аминокислоты избирательно поглощаются тканями как исходный материал для построения белков, а из тканей в кровь поступают другие аминокислоты. Синтезу и распаду подвергаются не только структурные белки, но и белки плазмы крови, а также белковые гормоны и ферменты.
Во взрослом организме аминокислоты или белки практически не запасаются, поэтому удаление аминокислот из крови происходит с такой же скоростью, как и их поступление из тканей в кровь. В растущем организме формируются новые ткани, и на этот процесс расходуется больше аминокислот, чем поступает в кровь за счет распада тканевых белков.
Печень участвует в метаболизме белков самым активным образом. Здесь синтезируются белки плазмы крови – альбумины и глобулины – а также собственные ферменты печени. Так, при потере плазменных белков содержание альбумина в плазме восстанавливается – за счет интенсивного синтеза – довольно быстро. Аминокислоты в печени используются не только для образования белков, но подвергаются также расщеплению, в ходе которого извлекается заключенная в них энергия.
Превращения и использование. Если аминокислоты используются в качестве источника энергии, то отщепляемая от них аминогруппа (–Nh3) направляется на образование мочевины, а не содержащий азота остаток молекулы окисляется приблизительно так же, как глюкоза или жирные кислоты.
Так называемый «орнитиновый цикл» описывает, как происходит превращение аммиака в мочевину. В этом цикле аминогруппа, отщепившаяся от аминокислоты в форме аммиака, присоединяется вместе с диоксидом углерода к молекуле орнитина с образованием цитруллина. Цитруллин присоединяет второй атом азота, на этот раз от аспарагиновой кислоты, и превращается в аргинин. Далее аргинин подвергается гидролизу с образованием мочевины и орнитина. Орнитин может теперь вновь вступить в цикл, а мочевина выводится из организма через почки как один из конечных продуктов метаболизма.
Список литературы
Ленинджер А. Основы биохимии, тт. 1–3. М., 1985
Страйер Л. Биохимия, тт. 1–3. М., 1985
Марри Р., Греннер Д., Мейес П., Родуэлл В. Биохимия человека, тт. 1–2. М., 1993
Албертс Б., Брей Д., Льюс Д. и др. Молекулярная биология клетки, тт. 1–3. М., 1994
www.neuch.ru
Метаболизм
Метаболизм, или обмен веществ, химические превращения, протекающие от момента поступления питательных веществ в живой организм до момента, когда конечные продукты этих превращений выделяются во внешнюю среду. К метаболизму относятся все реакции, в результате которых строятся структурные элементы клеток и тканей, и процессы, в которых из содержащихся в клетках веществ извлекается энергия. Иногда для удобства рассматривают по отдельности две стороны метаболизма – анаболизм и катаболизм, т.е. процессы созидания органических веществ и процессы их разрушения. Анаболические процессы обычно связаны с затратой энергии и приводят к образованию сложных молекул из более простых, катаболические же сопровождаются высвобождением энергии и заканчиваются образованием таких конечных продуктов (отходов) метаболизма, как мочевина, диоксид углерода, аммиак и вода.
Термин «обмен веществ» вошел в повседневную жизнь с тех пор, как врачи стали связывать избыточный или недостаточный вес, чрезмерную нервозность или, наоборот, вялость больного с повышенным или пониженным обменом. Для суждения об интенсивности метаболизма ставят тест на «основной обмен». Основной обмен – это показатель способности организма вырабатывать энергию. Тест проводят натощак в состоянии покоя; измеряют поглощение кислорода (О2) и выделение диоксида углерода (СО2). Сопоставляя эти величины, определяют, насколько полно организм использует («сжигает») питательные вещества. На интенсивность метаболизма влияют гормоны щитовидной железы, поэтому врачи при диагностике заболеваний, связанных с нарушениями обмена, в последнее время все чаще измеряют уровень этих гормонов в крови.
Методы исследования. При изучении метаболизма какого-нибудь одного из питательных веществ прослеживают все его превращения от той формы, в какой оно поступает в организм, до конечных продуктов, выводимых из организма. В таких исследованиях применяется крайне разнообразный набор биохимических методов.
Использование интактных животных или органов. Животному вводят изучаемое соединение, а затем в его моче и экскрементах определяют возможные продукты превращений (метаболиты) этого вещества. Более определенную информацию можно получить, исследуя метаболизм определенного органа, например печени или мозга. В этих случаях вещество вводят в соответствующий кровеносный сосуд, а метаболиты определяют в крови, оттекающей от данного органа.
Поскольку такого рода процедуры сопряжены с большими трудностями, часто для исследования используют тонкие срезы органов. Их инкубируют при комнатной температуре или при температуре тела в растворах с добавкой того вещества, метаболизм которого изучают. Клетки в таких препаратах не повреждены, и так как срезы очень тонкие, вещество легко проникает в клетки и легко выходит из них. Иногда затруднения возникают из-за слишком медленного прохождения вещества сквозь клеточные мембраны. В этих случаях ткани измельчают, чтобы разрушить мембраны, и с изучаемым веществом инкубируют клеточную кашицу. Именно в таких опытах было показано, что все живые клетки окисляют глюкозу до СО2 и воды и что только ткань печени способна синтезировать мочевину.
Использование клеток. Даже клетки представляют собой очень сложно организованные системы. В них имеется ядро, а в окружающей его цитоплазме находятся более мелкие тельца, т.н. органеллы, различных размеров и консистенции. С помощью соответствующей методики ткань можно «гомогенизировать», а затем подвергнуть дифференциальному центрифугированию (разделению) и получить препараты, содержащие только митохондрии, только микросомы или прозрачную жидкость – цитоплазму. Эти препараты можно по отдельности инкубировать с тем соединением, метаболизм которого изучается, и таким путем установить, какие именно субклеточные структуры участвуют в его последовательных превращениях. Известны случаи, когда начальная реакция протекает в цитоплазме, ее продукт подвергается превращению в микросомах, а продукт этого превращения вступает в новую реакцию уже в митохондриях. Инкубация изучаемого вещества с живыми клетками или с гомогенатом ткани обычно не выявляет отдельные этапы его метаболизма, и только последовательные эксперименты, в которых для инкубации используются те или иные субклеточные структуры, позволяют понять всю цепочку событий.
Использование радиоактивных изотопов. Для изучения метаболизма какого-либо вещества необходимы: 1) соответствующие аналитические методы для определения этого вещества и его метаболитов; и 2) методы, позволяющие отличать добавленное вещество от того же вещества, уже присутствующего в данном биологическом препарате. Эти требования служили главным препятствием при изучении метаболизма до тех пор, пока не были открыты радиоактивные изотопы элементов и в первую очередь радиоактивный углерод 14C. С появлением соединений, «меченных» 14C, а также приборов для измерения слабой радиоактивности эти трудности были преодолены. Если к биологическому препарату, например к суспензии митохондрий, добавляют меченную 14C жирную кислоту, то никаких специальных анализов для определения продуктов ее превращений не требуется; чтобы оценить скорость ее использования, достаточно просто измерять радиоактивность последовательно получаемых митохондриальных фракций. Эта же методика позволяет легко отличать молекулы радиоактивной жирной кислоты, введенной экспериментатором, от молекул жирной кислоты, уже присутствовавших в митохондриях к началу эксперимента.
Хроматография и электрофорез. В дополнение к вышеупомянутым требованиям биохимику необходимы и методы, позволяющие разделять смеси, состоящие из малых количеств органических веществ. Важнейший из них – хроматография, в основе которой лежит феномен адсорбции. Разделение компонентов смеси проводят при этом либо на бумаге, либо путем адсорбции на сорбенте, которым заполняют колонки (длинные стеклянные трубки), с последующей постепенной элюцией (вымыванием) каждого из компонентов.
Разделение методом электрофореза зависит от знака и числа зарядов ионизированных молекул. Электрофорез проводят на бумаге или на каком-нибудь инертном (неактивном) носителе, таком, как крахмал, целлюлоза или каучук.
Высокочувствительный и эффективный метод разделения – газовая хроматография. Им пользуются в тех случаях, когда подлежащие разделению вещества находятся в газообразном состоянии или могут быть в него переведены.
Выделение ферментов. Последнее место в описываемом ряду – животное, орган, тканевой срез, гомогенат и фракция клеточных органелл – занимает фермент, способный катализировать определенную химическую реакцию. Выделение ферментов в очищенном виде – важный раздел в изучении метаболизма.
Сочетание перечисленных методов позволило проследить главные метаболические пути у большей части организмов (в том числе у человека), установить, где именно эти различные процессы протекают, и выяснить последовательные этапы главных метаболических путей. К настоящему времени известны тысячи отдельных биохимических реакций, изучены участвующие в них ферменты.
Клеточный метаболизм. Живая клетка – это высокоорганизованная система. В ней имеются различные структуры, а также ферменты, способные их разрушить. Содержатся в ней и крупные макромолекулы, которые могут распадаться на более мелкие компоненты в результате гидролиза (расщепления под действием воды). В клетке обычно много калия и очень мало натрия, хотя клетка существует в среде, где натрия много, а калия относительно мало, и клеточная мембрана легко проницаема для обоих ионов. Следовательно, клетка – это химическая система, весьма далекая от равновесия. Равновесие наступает только в процессе посмертного автолиза (самопереваривания под действием собственных ферментов).
Потребность в энергии. Чтобы удержать систему в состоянии, далеком от химического равновесия, требуется производить работу, а для этого необходима энергия. Получение этой энергии и выполнение этой работы – непременное условие для того, чтобы клетка оставалась в своем стационарном (нормальном) состоянии, далеком от равновесия. Одновременно в ней выполняется и иная работа, связанная со взаимодействием со средой, например: в мышечных клетках – сокращение; в нервных клетках – проведение нервного импульса; в клетках почек – образование мочи, значительно отличающейся по своему составу от плазмы крови; в специализированных клетках желудочно-кишечного тракта – синтез и выделение пищеварительных ферментов; в клетках эндокринных желез – секреция гормонов; в клетках светляков – свечение; в клетках некоторых рыб – генерирование электрических разрядов и т.д.
Источники энергии. В любом из перечисленных выше примеров непосредственным источником энергии, которую клетка использует для производства работы, служит энергия, заключенная в структуре аденозинтрифосфата (АТФ). В силу особенностей своей структуры это соединение богато энергией, и разрыв связей между его фосфатными группами может происходить таким образом, что высвобождающаяся энергия используется для производства работы. Однако энергия не может стать доступной для клетки при простом гидролитическом разрыве фосфатных связей АТФ: в этом случае она расходуется впустую, выделяясь в виде тепла. Процесс должен состоять из двух последовательных этапов, в каждом из которых участвует промежуточный продукт, обозначенный здесь X–Ф (в приведенных уравнениях X и Y означают два разных органических вещества; Ф – фосфат; АДФ – аденозиндифосфат):
Поскольку практически для любого проявления жизнедеятельности клеток необходим АТФ, неудивительно, что метаболическая активность живых клеток направлена в первую очередь на синтез АТФ. Этой цели служат различные сложные последовательности реакций, в которых используется потенциальная химическая энергия, заключенная в молекулах углеводов и жиров (липидов).
Метаболизм углеводов и липидов
Синтез АТФ. Анаэробный (без участия кислорода). Главная роль углеводов и липидов в клеточном метаболизме состоит в том, что их расщепление на более простые соединения обеспечивает синтез АТФ. Несомненно, что те же процессы протекали и в первых, самых примитивных клетках. Однако в атмосфере, лишенной кислорода, полное окисление углеводов и жиров до CO2 было невозможно. У этих примитивных клеток имелись все же механизмы, с помощью которых перестройка структуры молекулы глюкозы обеспечивала синтез небольших количеств АТФ. Речь идет о процессах, которые у микроорганизмов называют брожением. Лучше всего изучено сбраживание глюкозы до этилового спирта и CO2 у дрожжей.
В ходе 11 последовательных реакций, необходимых для того, чтобы завершилось это превращение, образуется ряд промежуточных продуктов, представляющих собой эфиры фосфорной кислоты (фосфаты). Их фосфатная группа переносится на аденозиндифосфат (АДФ) с образованием АТФ. Чистый выход АТФ составляет 2 молекулы АТФ на каждую молекулу глюкозы, расщепленную в процессе брожения. Аналогичные процессы происходят во всех живых клетках; поскольку они поставляют необходимую для жизнедеятельности энергию, их иногда (не вполне корректно) называют анаэробным дыханием клеток.
У млекопитающих, в том числе у человека, такой процесс называется гликолизом и его конечным продуктом является молочная кислота, а не спирт и CO2. Вся последовательность реакций гликолиза, за исключением двух последних этапов, полностью идентична процессу, протекающему в дрожжевых клетках.
Аэробный (с использованием кислорода). С появлением в атмосфере кислорода, источником которого послужил, очевидно, фотосинтез растений, в ходе эволюции развился механизм, обеспечивающий полное окисление глюкозы до CO2 и воды, – аэробный процесс, в котором чистый выход АТФ составляет 38 молекул АТФ на каждую окисленную молекулу глюкозы. Этот процесс потребления клетками кислорода для образования богатых энергией соединений известен как клеточное дыхание (аэробное). В отличие от анаэробного процесса, осуществляемого ферментами цитоплазмы, окислительные процессы протекают в митохондриях. В митохондриях пировиноградная кислота – промежуточный продукт, образовавшийся в анаэробной фазе – окисляется до СО2 в шести последовательных реакциях, в каждой из которых пара электронов переносится на общий акцептор – кофермент никотинамидадениндинуклеотид (НАД). Эту последовательность реакций называют циклом трикарбоновых кислот, циклом лимонной кислоты или циклом Кребса. Из каждой молекулы глюкозы образуется 2 молекулы пировиноградной кислоты; 12 пар электронов отщепляется от молекулы глюкозы в ходе ее окисления, описываемого уравнением:
Перенос электронов. В каждой митохондрии имеется механизм, посредством которого восстановленный НАД (НАДН, где Н – водород), образовавшийся в цикле трикарбоновых кислот, передает свою пару электронов кислороду. Перенос, однако, не происходит напрямую. Электроны как бы передаются «из рук в руки» и, лишь пройдя цепь переносчиков, присоединяются к кислороду. Эта «цепь переноса электронов» состоит из следующих компонентов:
НАДНН Флавинадениндинклеотид Кофермент Q
Цитохром b Цитохром c Цитохром a O2
Все компоненты этой системы, находящиеся в митохондриях, фиксированы в пространстве и сцеплены друг с другом. Такое их состояние облегчает перенос электронов.
В состав НАД входит никотиновая кислота (витамин ниацин), а в состав флавинадениндинуклеотида – рибофлавин (витамин B2). Кофермент Q представляет собой высокомолекулярный хинон, синтезируемый в печени, а цитохромы – это три разных белка, каждый из которых, подобно гемоглобину, содержит гемогруппу.
В цепи переноса электронов на каждую пару электронов, перенесенную от НАДН на O2, синтезируется 3 молекулы АТФ. Поскольку от каждой молекулы глюкозы отщепляются и передаются молекулам НАД 12 пар электронов, в общей сложности на каждую молекулу глюкозы образуется 312 = 36 молекул АТФ. Этот процесс образования АТФ в ходе окисления называется окислительным фосфорилированием.
Липиды как источник энергии. Жирные кислоты могут использоваться в качестве источника энергии приблизительно так же, как и углеводы. Окисление жирных кислот протекает путем последовательного отщепления от молекулы жирной кислоты двууглеродного фрагмента с образованием ацетилкофермента A (ацетил-КоА) и одновременной передачей двух пар электронов в цепь переноса электронов. Образовавшийся ацетил-КоА – нормальный компонент цикла трикарбоновых кислот, и в дальнейшем его судьба не отличается от судьбы ацетил-КоА, поставляемого углеводным обменом. Таким образом, механизмы синтеза АТФ при окислении как жирных кислот, так и метаболитов глюкозы практически одинаковы.
Если организм животного получает энергию почти целиком за счет одного только окисления жирных кислот, а это бывает, например, при голодании или при сахарном диабете, то скорость образования ацетил-КоА превышает скорость его окисления в цикле трикарбоновых кислот. В этом случае лишние молекулы ацетил-КоА реагируют друг с другом, в результате чего образуются в конечном счете ацетоуксусная и -гидроксимасляная кислоты. Их накопление является причиной патологического состояния, т.н. кетоза (одного из видов ацидоза), который при тяжелом диабете может вызвать кому и смерть.
Запасание энергии. Животные питаются нерегулярно, и их организму нужно как-то запасать заключенную в пище энергию, источником которой являются поглощенные животным углеводы и жиры. Жирные кислоты могут запасаться в виде нейтральных жиров либо в печени, либо в жировой ткани. Углеводы, поступая в большом количестве, в желудочно-кишечном тракте гидролизуются до глюкозы или иных сахаров, которые затем в печени превращаются в ту же глюкозу. Здесь из глюкозы синтезируется гигантский полимер гликоген путем присоединения друг к другу остатков глюкозы с отщеплением молекул воды (число остатков глюкозы в молекулах гликогена доходит до 30 000). Когда возникает потребность в энергии, гликоген вновь распадается до глюкозы в реакции, продуктом которой является глюкозофосфат. Этот глюкозофосфат направляется на путь гликолиза – процесса, составляющего часть пути окисления глюкозы. В печени глюкозофосфат может также подвергнуться гидролизу, и образующаяся глюкоза поступает в кровоток и доставляется кровью к клеткам в разных частях тела.
Синтез липидов из углеводов. Если количество углеводов, поглощенных с пищей за один прием, больше того, какое может быть запасено в виде гликогена, то избыток углеводов превращается в жиры. Начальная последовательность реакций совпадает при этом с обычным окислительным путем, т.е. сначала из глюкозы образуется ацетил-КоА, но далее этот ацетил-КоА используется в цитоплазме клетки для синтеза длинноцепочечных жирных кислот. Процесс синтеза можно описать как обращение обычного процесса окисления жирных клеток. Затем жирные кислоты запасаются в виде нейтральных жиров (триглицеридов), отлагающихся в разных частях тела. Когда требуется энергия, нейтральные жиры подвергаются гидролизу и жирные кислоты поступают в кровь. Здесь они адсорбируются молекулами плазменных белков (альбуминов и глобулинов) и затем поглощаются клетками самых разных типов. Механизмов, способных осуществлять синтез глюкозы из жирных кислот, у животных нет, но у растений такие механизмы имеются.
www.coolreferat.com
Метаболизм | Рефераты KM.RU
Метаболизм, или обмен веществ, химические превращения, протекающие от момента поступления питательных веществ в живой организм до момента, когда конечные продукты этих превращений выделяются во внешнюю среду. К метаболизму относятся все реакции, в результате которых строятся структурные элементы клеток и тканей, и процессы, в которых из содержащихся в клетках веществ извлекается энергия. Иногда для удобства рассматривают по отдельности две стороны метаболизма – анаболизм и катаболизм, т.е. процессы созидания органических веществ и процессы их разрушения. Анаболические процессы обычно связаны с затратой энергии и приводят к образованию сложных молекул из более простых, катаболические же сопровождаются высвобождением энергии и заканчиваются образованием таких конечных продуктов (отходов) метаболизма, как мочевина, диоксид углерода, аммиак и вода.
Термин «обмен веществ» вошел в повседневную жизнь с тех пор, как врачи стали связывать избыточный или недостаточный вес, чрезмерную нервозность или, наоборот, вялость больного с повышенным или пониженным обменом. Для суждения об интенсивности метаболизма ставят тест на «основной обмен». Основной обмен – это показатель способности организма вырабатывать энергию. Тест проводят натощак в состоянии покоя; измеряют поглощение кислорода (О2) и выделение диоксида углерода (СО2). Сопоставляя эти величины, определяют, насколько полно организм использует («сжигает») питательные вещества. На интенсивность метаболизма влияют гормоны щитовидной железы, поэтому врачи при диагностике заболеваний, связанных с нарушениями обмена, в последнее время все чаще измеряют уровень этих гормонов в крови.
Методы исследования. При изучении метаболизма какого-нибудь одного из питательных веществ прослеживают все его превращения от той формы, в какой оно поступает в организм, до конечных продуктов, выводимых из организма. В таких исследованиях применяется крайне разнообразный набор биохимических методов.
Использование интактных животных или органов. Животному вводят изучаемое соединение, а затем в его моче и экскрементах определяют возможные продукты превращений (метаболиты) этого вещества. Более определенную информацию можно получить, исследуя метаболизм определенного органа, например печени или мозга. В этих случаях вещество вводят в соответствующий кровеносный сосуд, а метаболиты определяют в крови, оттекающей от данного органа.
Поскольку такого рода процедуры сопряжены с большими трудностями, часто для исследования используют тонкие срезы органов. Их инкубируют при комнатной температуре или при температуре тела в растворах с добавкой того вещества, метаболизм которого изучают. Клетки в таких препаратах не повреждены, и так как срезы очень тонкие, вещество легко проникает в клетки и легко выходит из них. Иногда затруднения возникают из-за слишком медленного прохождения вещества сквозь клеточные мембраны. В этих случаях ткани измельчают, чтобы разрушить мембраны, и с изучаемым веществом инкубируют клеточную кашицу. Именно в таких опытах было показано, что все живые клетки окисляют глюкозу до СО2 и воды и что только ткань печени способна синтезировать мочевину.
Использование клеток. Даже клетки представляют собой очень сложно организованные системы. В них имеется ядро, а в окружающей его цитоплазме находятся более мелкие тельца, т.н. органеллы, различных размеров и консистенции. С помощью соответствующей методики ткань можно «гомогенизировать», а затем подвергнуть дифференциальному центрифугированию (разделению) и получить препараты, содержащие только митохондрии, только микросомы или прозрачную жидкость – цитоплазму. Эти препараты можно по отдельности инкубировать с тем соединением, метаболизм которого изучается, и таким путем установить, какие именно субклеточные структуры участвуют в его последовательных превращениях. Известны случаи, когда начальная реакция протекает в цитоплазме, ее продукт подвергается превращению в микросомах, а продукт этого превращения вступает в новую реакцию уже в митохондриях. Инкубация изучаемого вещества с живыми клетками или с гомогенатом ткани обычно не выявляет отдельные этапы его метаболизма, и только последовательные эксперименты, в которых для инкубации используются те или иные субклеточные структуры, позволяют понять всю цепочку событий.
Использование радиоактивных изотопов. Для изучения метаболизма какого-либо вещества необходимы: 1) соответствующие аналитические методы для определения этого вещества и его метаболитов; и 2) методы, позволяющие отличать добавленное вещество от того же вещества, уже присутствующего в данном биологическом препарате. Эти требования служили главным препятствием при изучении метаболизма до тех пор, пока не были открыты радиоактивные изотопы элементов и в первую очередь радиоактивный углерод 14C. С появлением соединений, «меченных» 14C, а также приборов для измерения слабой радиоактивности эти трудности были преодолены. Если к биологическому препарату, например к суспензии митохондрий, добавляют меченную 14C жирную кислоту, то никаких специальных анализов для определения продуктов ее превращений не требуется; чтобы оценить скорость ее использования, достаточно просто измерять радиоактивность последовательно получаемых митохондриальных фракций. Эта же методика позволяет легко отличать молекулы радиоактивной жирной кислоты, введенной экспериментатором, от молекул жирной кислоты, уже присутствовавших в митохондриях к началу эксперимента.
Хроматография и электрофорез. В дополнение к вышеупомянутым требованиям биохимику необходимы и методы, позволяющие разделять смеси, состоящие из малых количеств органических веществ. Важнейший из них – хроматография, в основе которой лежит феномен адсорбции. Разделение компонентов смеси проводят при этом либо на бумаге, либо путем адсорбции на сорбенте, которым заполняют колонки (длинные стеклянные трубки), с последующей постепенной элюцией (вымыванием) каждого из компонентов.
Разделение методом электрофореза зависит от знака и числа зарядов ионизированных молекул. Электрофорез проводят на бумаге или на каком-нибудь инертном (неактивном) носителе, таком, как крахмал, целлюлоза или каучук.
Высокочувствительный и эффективный метод разделения – газовая хроматография. Им пользуются в тех случаях, когда подлежащие разделению вещества находятся в газообразном состоянии или могут быть в него переведены.
Выделение ферментов. Последнее место в описываемом ряду – животное, орган, тканевой срез, гомогенат и фракция клеточных органелл – занимает фермент, способный катализировать определенную химическую реакцию. Выделение ферментов в очищенном виде – важный раздел в изучении метаболизма.
Сочетание перечисленных методов позволило проследить главные метаболические пути у большей части организмов (в том числе у человека), установить, где именно эти различные процессы протекают, и выяснить последовательные этапы главных метаболических путей. К настоящему времени известны тысячи отдельных биохимических реакций, изучены участвующие в них ферменты.
Клеточный метаболизм. Живая клетка – это высокоорганизованная система. В ней имеются различные структуры, а также ферменты, способные их разрушить. Содержатся в ней и крупные макромолекулы, которые могут распадаться на более мелкие компоненты в результате гидролиза (расщепления под действием воды). В клетке обычно много калия и очень мало натрия, хотя клетка существует в среде, где натрия много, а калия относительно мало, и клеточная мембрана легко проницаема для обоих ионов. Следовательно, клетка – это химическая система, весьма далекая от равновесия. Равновесие наступает только в процессе посмертного автолиза (самопереваривания под действием собственных ферментов).
Потребность в энергии. Чтобы удержать систему в состоянии, далеком от химического равновесия, требуется производить работу, а для этого необходима энергия. Получение этой энергии и выполнение этой работы – непременное условие для того, чтобы клетка оставалась в своем стационарном (нормальном) состоянии, далеком от равновесия. Одновременно в ней выполняется и иная работа, связанная со взаимодействием со средой, например: в мышечных клетках – сокращение; в нервных клетках – проведение нервного импульса; в клетках почек – образование мочи, значительно отличающейся по своему составу от плазмы крови; в специализированных клетках желудочно-кишечного тракта – синтез и выделение пищеварительных ферментов; в клетках эндокринных желез – секреция гормонов; в клетках светляков – свечение; в клетках некоторых рыб – генерирование электрических разрядов и т.д.
Источники энергии. В любом из перечисленных выше примеров непосредственным источником энергии, которую клетка использует для производства работы, служит энергия, заключенная в структуре аденозинтрифосфата (АТФ). В силу особенностей своей структуры это соединение богато энергией, и разрыв связей между его фосфатными группами может происходить таким образом, что высвобождающаяся энергия используется для производства работы. Однако энергия не может стать доступной для клетки при простом гидролитическом разрыве фосфатных связей АТФ: в этом случае она расходуется впустую, выделяясь в виде тепла. Процесс должен состоять из двух последовательных этапов, в каждом из которых участвует промежуточный продукт, обозначенный здесь X–Ф (в приведенных уравнениях X и Y означают два разных органических вещества; Ф – фосфат; АДФ – аденозиндифосфат):
Поскольку практически для любого проявления жизнедеятельности клеток необходим АТФ, неудивительно, что метаболическая активность живых клеток направлена в первую очередь на синтез АТФ. Этой цели служат различные сложные последовательности реакций, в которых используется потенциальная химическая энергия, заключенная в молекулах углеводов и жиров (липидов).
Метаболизм углеводов и липидов
Синтез АТФ. Анаэробный (без участия кислорода). Главная роль углеводов и липидов в клеточном метаболизме состоит в том, что их расщепление на более простые соединения обеспечивает синтез АТФ. Несомненно, что те же процессы протекали и в первых, самых примитивных клетках. Однако в атмосфере, лишенной кислорода, полное окисление углеводов и жиров до CO2 было невозможно. У этих примитивных клеток имелись все же механизмы, с помощью которых перестройка структуры молекулы глюкозы обеспечивала синтез небольших количеств АТФ. Речь идет о процессах, которые у микроорганизмов называют брожением. Лучше всего изучено сбраживание глюкозы до этилового спирта и CO2 у дрожжей.
В ходе 11 последовательных реакций, необходимых для того, чтобы завершилось это превращение, образуется ряд промежуточных продуктов, представляющих собой эфиры фосфорной кислоты (фосфаты). Их фосфатная группа переносится на аденозиндифосфат (АДФ) с образованием АТФ. Чистый выход АТФ составляет 2 молекулы АТФ на каждую молекулу глюкозы, расщепленную в процессе брожения. Аналогичные процессы происходят во всех живых клетках; поскольку они поставляют необходимую для жизнедеятельности энергию, их иногда (не вполне корректно) называют анаэробным дыханием клеток.
У млекопитающих, в том числе у человека, такой процесс называется гликолизом и его конечным продуктом является молочная кислота, а не спирт и CO2. Вся последовательность реакций гликолиза, за исключением двух последних этапов, полностью идентична процессу, протекающему в дрожжевых клетках.
Аэробный (с использованием кислорода). С появлением в атмосфере кислорода, источником которого послужил, очевидно, фотосинтез растений, в ходе эволюции развился механизм, обеспечивающий полное окисление глюкозы до CO2 и воды, – аэробный процесс, в котором чистый выход АТФ составляет 38 молекул АТФ на каждую окисленную молекулу глюкозы. Этот процесс потребления клетками кислорода для образования богатых энергией соединений известен как клеточное дыхание (аэробное). В отличие от анаэробного процесса, осуществляемого ферментами цитоплазмы, окислительные процессы протекают в митохондриях. В митохондриях пировиноградная кислота – промежуточный продукт, образовавшийся в анаэробной фазе – окисляется до СО2 в шести последовательных реакциях, в каждой из которых пара электронов переносится на общий акцептор – кофермент никотинамидадениндинуклеотид (НАД). Эту последовательность реакций называют циклом трикарбоновых кислот, циклом лимонной кислоты или циклом Кребса. Из каждой молекулы глюкозы образуется 2 молекулы пировиноградной кислоты; 12 пар электронов отщепляется от молекулы глюкозы в ходе ее окисления, описываемого уравнением:
Перенос электронов. В каждой митохондрии имеется механизм, посредством которого восстановленный НАД (НАДН, где Н – водород), образовавшийся в цикле трикарбоновых кислот, передает свою пару электронов кислороду. Перенос, однако, не происходит напрямую. Электроны как бы передаются «из рук в руки» и, лишь пройдя цепь переносчиков, присоединяются к кислороду. Эта «цепь переноса электронов» состоит из следующих компонентов:
НАДНН Флавинадениндинклеотид Кофермент Q
Цитохром b Цитохром c Цитохром a O2
Все компоненты этой системы, находящиеся в митохондриях, фиксированы в пространстве и сцеплены друг с другом. Такое их состояние облегчает перенос электронов.
В состав НАД входит никотиновая кислота (витамин ниацин), а в состав флавинадениндинуклеотида – рибофлавин (витамин B2). Кофермент Q представляет собой высокомолекулярный хинон, синтезируемый в печени, а цитохромы – это три разных белка, каждый из которых, подобно гемоглобину, содержит гемогруппу.
В цепи переноса электронов на каждую пару электронов, перенесенную от НАДН на O2, синтезируется 3 молекулы АТФ. Поскольку от каждой молекулы глюкозы отщепляются и передаются молекулам НАД 12 пар электронов, в общей сложности на каждую молекулу глюкозы образуется 312 = 36 молекул АТФ. Этот процесс образования АТФ в ходе окисления называется окислительным фосфорилированием.
Липиды как источник энергии. Жирные кислоты могут использоваться в качестве источника энергии приблизительно так же, как и углеводы. Окисление жирных кислот протекает путем последовательного отщепления от молекулы жирной кислоты двууглеродного фрагмента с образованием ацетилкофермента A (ацетил-КоА) и одновременной передачей двух пар электронов в цепь переноса электронов. Образовавшийся ацетил-КоА – нормальный компонент цикла трикарбоновых кислот, и в дальнейшем его судьба не отличается от судьбы ацетил-КоА, поставляемого углеводным обменом. Таким образом, механизмы синтеза АТФ при окислении как жирных кислот, так и метаболитов глюкозы практически одинаковы.
Если организм животного получает энергию почти целиком за счет одного только окисления жирных кислот, а это бывает, например, при голодании или при сахарном диабете, то скорость образования ацетил-КоА превышает скорость его окисления в цикле трикарбоновых кислот. В этом случае лишние молекулы ацетил-КоА реагируют друг с другом, в результате чего образуются в конечном счете ацетоуксусная и -гидроксимасляная кислоты. Их накопление является причиной патологического состояния, т.н. кетоза (одного из видов ацидоза), который при тяжелом диабете может вызвать кому и смерть.
Запасание энергии. Животные питаются нерегулярно, и их организму нужно как-то запасать заключенную в пище энергию, источником которой являются поглощенные животным углеводы и жиры. Жирные кислоты могут запасаться в виде нейтральных жиров либо в печени, либо в жировой ткани. Углеводы, поступая в большом количестве, в желудочно-кишечном тракте гидролизуются до глюкозы или иных сахаров, которые затем в печени превращаются в ту же глюкозу. Здесь из глюкозы синтезируется гигантский полимер гликоген путем присоединения друг к другу остатков глюкозы с отщеплением молекул воды (число остатков глюкозы в молекулах гликогена доходит до 30 000). Когда возникает потребность в энергии, гликоген вновь распадается до глюкозы в реакции, продуктом которой является глюкозофосфат. Этот глюкозофосфат направляется на путь гликолиза – процесса, составляющего часть пути окисления глюкозы. В печени глюкозофосфат может также подвергнуться гидролизу, и образующаяся глюкоза поступает в кровоток и доставляется кровью к клеткам в разных частях тела.
Синтез липидов из углеводов. Если количество углеводов, поглощенных с пищей за один прием, больше того, какое может быть запасено в виде гликогена, то избыток углеводов превращается в жиры. Начальная последовательность реакций совпадает при этом с обычным окислительным путем, т.е. сначала из глюкозы образуется ацетил-КоА, но далее этот ацетил-КоА используется в цитоплазме клетки для синтеза длинноцепочечных жирных кислот. Процесс синтеза можно описать как обращение обычного процесса окисления жирных клеток. Затем жирные кислоты запасаются в виде нейтральных жиров (триглицеридов), отлагающихся в разных частях тела. Когда требуется энергия, нейтральные жиры подвергаются гидролизу и жирные кислоты поступают в кровь. Здесь они адсорбируются молекулами плазменных белков (альбуминов и глобулинов) и затем поглощаются клетками самых разных типов. Механизмов, способных осуществлять синтез глюкозы из жирных кислот, у животных нет, но у растений такие механизмы имеются.
Метаболизм липидов. Липиды попадают в организм главным образом в форме триглицеридов жирных кислот. В кишечнике под действием ферментов поджелудочной железы они подвергаются гидролизу, продукты которого всасываются клетками стенки кишечника. Здесь из них вновь синтезируются нейтральные жиры, которые через лимфатическую систему поступают в кровь и либо транспортируются в печень, либо отлагаются в жировой ткани. Выше уже указывалось, что жирные кислоты могут также синтезироваться заново из углеводных предшественников. Следует отметить, что, хотя в клетках млекопитающих может происходить включение одной двойной связи в молекулы длинноцепочечных жирных кислот (между С–9 и С–10), включать вторую и третью двойную связь эти клетки неспособны. Поскольку жирные кислоты с двумя и тремя двойными связями играют важную роль в метаболизме млекопитающих, они в сущности являются витаминами. Поэтому линолевую (C18:2) и линоленовую (C18:3) кислоты называют незаменимыми жирными кислотами. В то же время в клетках млекопитающих в линоленовую кислоту может включаться четвертая двойная связь и путем удлинения углеродной цепи может образоваться арахидоновая кислота (C20:4), также необходимый участник метаболических процессов.
В процессе синтеза липидов остатки жирных кислот, связанные с коферментом А (ацил-КоА), переносятся на глицерофосфат – эфир фосфорной кислоты и глицерина. В результате образуется фосфатидная кислота – соединение, в котором одна гидроксильная группа глицерина этерифицирована фосфорной кислотой, а две группы – жирными кислотами. При образовании нейтральных жиров фосфорная кислота удаляется путем гидролиза, и ее место занимает третья жирная кислота в результате реакции с ацил-КоА. Кофермент А образуется из пантотеновой кислоты (одного из витаминов). В его молекуле имеется сульфгидрильная (– SH) группа, способная реагировать с кислотами с образованием тиоэфиров. При образовании фосфолипидов фосфатидная кислота реагирует непосредственно с активированным производным одного из азотистых оснований, таких, как холин, этаноламин или серин.
За исключением витамина D, все встречающиеся в организме животных стероиды (производные сложных спиртов) легко синтезируются самим организмом. Сюда относятся холестерин (холестерол), желчные кислоты, мужские и женские половые гормоны и гормоны надпочечников. В каждом случае исходным материалом для синтеза служит ацетил-КоА: из ацетильных групп путем многократно повторяющейся конденсации строится углеродный скелет синтезируемого соединения.
Метаболизм белков
Синтез аминокислот. Растения и большинство микроорганизмов могут жить и расти в среде, в которой для их питания имеются только минеральные вещества, диоксид углерода и вода. Это значит, что все обнаруживаемые в них органические вещества эти организмы синтезируют сами. Встречающиеся во всех живых клетках белки построены из 21 вида аминокислот, соединенных в различной последовательности. Аминокислоты синтезируются живыми организмами. В каждом случае ряд химических реакций приводит к образованию -кетокислоты. Одна такая -кетокислота, а именно -кетоглутаровая (обычный компонент цикла трикарбоновых кислот), участвует в связывании азота по следующему уравнению:
-Кетоглутаровая кислота + Nh4 + НАДН
Глутаминовая кислота + НАД.
Азот глутаминовой кислоты может быть затем передан любой из других -кетокислот с образованием соответствующей аминокислоты.
Организм человека и большинства других животных сохранил способность синтезировать все аминокислоты за исключением девяти т.н. незаменимых аминокислот. Поскольку кетокислоты, соответствующие этим девяти, не синтезируются, незаменимые аминокислоты должны поступать с пищей.
Синтез белков. Аминокислоты нужны для биосинтеза белка. Процесс биосинтеза протекает обычно следующим образом. В цитоплазме клетки каждая аминокислота «активируется» в реакции с АТФ, а затем присоединяется к концевой группе молекулы рибонуклеиновой кислоты, специфичной именно для данной аминокислоты. Эта сложная молекула связывается с небольшим тельцем, т.н. рибосомой, в положении, определяемом более длинной молекулой рибонуклеиновой кислоты, прикрепленной к рибосоме. После того как все эти сложные молекулы соответствующим образом выстроились, связи между исходной аминокислотой и рибонуклеиновой кислотой разрываются и возникают связи между соседними аминокислотами – синтезируется специфичный белок. Процесс биосинтеза поставляет белки не только для роста организма или для секреции в среду. Все белки живых клеток со временем претерпевают распад до составляющих их аминокислот, и для поддержания жизни клетки должны синтезироваться вновь.
Синтез других азотсодержащих соединений. В организме млекопитающих аминокислоты используются не только для биосинтеза белков, но и как исходный материал для синтеза многих азотсодержащих соединений. Аминокислота тирозин является предшественником гормонов адреналина и норадреналина. Простейшая аминокислота глицин служит исходным материалом для биосинтеза пуринов, входящих в состав нуклеиновых кислот, и порфиринов, входящих в состав цитохромов и гемоглобина. Аспарагиновая кислота – предшественник пиримидинов нуклеиновых кислот. Метильная группа метионина передается ряду других соединений в ходе биосинтеза креатина, холина и саркозина. При биосинтезе креатина от одного соединения к другому передается также и гуанидиновая группировка аргинина. Триптофан служит предшественником никотиновой кислоты, а из валина в растениях синтезируется такой витамин, как пантотеновая кислота. Все это лишь отдельные примеры использования аминокислот в процессах биосинтеза.
Азот, поглощаемый микроорганизмами и высшими растениями в виде иона аммония, расходуется почти целиком на образование аминокислот, из которых затем синтезируются многие азотсодержащие соединения живых клеток. Избыточных количеств азота ни растения, ни микроорганизмы не поглощают. В отличие от них, у животных количество поглощенного азота зависит от содержащихся в пище белков. Весь азот, поступивший в организм в виде аминокислот и не израсходованный в процессах биосинтеза, довольно быстро выводится из организма с мочой. Происходит это следующим образом. В печени неиспользованные аминокислоты передают свой азот -кетоглутаровой кислоте с образованием глутаминовой кислоты, которая дезаминируется, высвобождая аммиак. Далее азот аммиака может либо на время запасаться путем синтеза глутамина, либо сразу же использоваться для синтеза мочевины, протекающего в печени.
У глутамина есть и другая роль. Он может подвергаться гидролизу в почках с высвобождением аммиака, который поступает в мочу в обмен на ионы натрия. Этот процесс крайне важен как средство поддержания кислотно-щелочного равновесия в организме животного. Почти весь аммиак, происходящий из аминокислот и, возможно, из других источников, превращается в печени в мочевину, так что свободного аммиака в крови обычно почти нет. Однако при некоторых условиях довольно значительные количества аммиака содержит моча. Этот аммиак образуется в почках из глутамина и переходит в мочу в обмен на ионы натрия, которые таким образом реадсорбируются и задерживаются в организме. Этот процесс усиливается при развитии ацидоза – состояния, при котором организм нуждается в дополнительных количествах катионов натрия для связывания избытка ионов бикарбоната в крови.
Избыточные количества пиримидинов тоже распадаются в печени через ряд реакций, в которых высвобождается аммиак. Что касается пуринов, то их избыток подвергается окислению с образованием мочевой кислоты, выделяющейся с мочой у человека и других приматов, но не у остальных млекопитающих. У птиц отсутствует механизм синтеза мочевины, и именно мочевая кислота, а не мочевина, является у них конечным продуктом обмена всех азотсодержащих соединений.
Общие представления о метаболизме органических веществ
Можно сформулировать некоторые общие понятия, или «правила», касающиеся метаболизма. Приведенные ниже несколько главных «правил» позволяют лучше понять, как протекает и регулируется метаболизм.
1. Метаболические пути необратимы. Распад никогда не идет по пути, который являлся бы простым обращением реакций синтеза. В нем участвуют другие ферменты и другие промежуточные продукты. Нередко противоположно направленные процессы протекают в разных отсеках клетки. Так, жирные кислоты синтезируются в цитоплазме при участии одного набора ферментов, а окисляются в митохондриях при участии совсем другого набора.
2. Ферментов в живых клетках достаточно для того, чтобы все известные метаболические реакции могли протекать гораздо быстрее, чем это обычно наблюдается в организме. Следовательно, в клетках существуют какие-то регуляторные механизмы. Открыты разные типы таких механизмов.
а) Фактором, ограничивающим скорость метаболических превращений данного вещества, может быть поступление этого вещества в клетку; именно на этот процесс в таком случае и направлена регуляция. Роль инсулина, например, связана с тем, что он, по-видимому, облегчает проникновение глюкозы во все клетки, глюкоза же подвергается превращениям с той скоростью, с какой она поступает. Сходным образом проникновение железа и кальция из кишечника в кровь зависит от процессов, скорость которых регулируется.
б) Вещества далеко не всегда могут свободно переходить из одного клеточного отсека в другой; есть данные, что внутриклеточный перенос регулируется некоторыми стероидными гормонами.
в) Выявлено два типа сервомеханизмов «отрицательной обратной связи».
У бактерий были обнаружены примеры того, что присутствие продукта какой-нибудь последовательности реакций, например аминокислоты, подавляет биосинтез одного из ферментов, необходимых для образования этой аминокислоты.
В каждом случае фермент, биосинтез которого оказывается затронутым, был ответствен за первый «определяющий» этап (на схеме реакция 4) метаболического пути, ведущего к синтезу данной аминокислоты.
Второй механизм хорошо изучен у млекопитающих. Это простое ингибирование конечным продуктом (в нашем случае – аминокислотой) фермента, ответственного за первый «определяющий» этап метаболического пути.
Еще один тип регулирования посредством обратной связи действует в тех случаях, когда окисление промежуточных продуктов цикла трикарбоновых кислот сопряжено с образованием АТФ из АДФ и фосфата в процессе окислительного фосфорилирования. Если весь имеющийся в клетке запас фосфата и (или) АДФ уже исчерпан, то окисление приостанавливается и может возобновиться лишь после того, как этот запас вновь станет достаточным. Таким образом, окисление, смысл которого в том, чтобы поставлять полезную энергию в форме АТФ, происходит только тогда, когда возможен синтез АТФ.
3. В биосинтетических процессах участвует сравнительно небольшое число строительных блоков, каждый из которых используется для синтеза многих соединений. Среди них можно назвать ацетилкофермент А, глицерофосфат, глицин, карбамилфосфат, поставляющий карбамильную (h3N–CO–) группу, производные фолиевой кислоты, служащие источником гидроксиметильной и формильной групп, S-аденозилметионин – источник метильных групп, глутаминовую и аспарагиновую кислоты, поставляющие аминогруппы, и наконец, глутамин – источник амидных групп. Из этого относительно небольшого числа компонентов строятся все те разнообразные соединения, которые мы находим в живых организмах.
4. Простые органические соединения редко участвуют в метаболических реакциях непосредственно. Обычно они должны быть сначала «активированы» путем присоединения к одному из ряда соединений, универсально используемых в метаболизме. Глюкоза, например, может подвергнуться окислению лишь после того, как она будет этерифицирована фосфорной кислотой, для прочих же своих превращений она должна быть этерифицирована уридиндифосфатом. Жирные кислоты не могут быть вовлечены в метаболические превращения прежде, чем они образуют эфиры с коферментом А. Каждый из этих активаторов либо родствен одному из нуклеотидов, входящих в состав рибонуклеиновой кислоты, либо образуется из какого-нибудь витамина. Легко понять в связи с этим, почему витамины требуются в таких небольших количествах. Они расходуются на образование «коферментов», а каждая молекула кофермента на протяжении жизни организма используется многократно, в отличие от основных питательных веществ (например, глюкозы), каждая молекула которых используется только один раз.
В заключение следует сказать, что термин «метаболизм», означавший ранее нечто не более сложное, чем просто использование углеводов и жиров в организме, теперь применяется для обозначения тысяч ферментативных реакций, вся совокупность которых может быть представлена как огромная сеть метаболических путей, многократно пересекающихся (из-за наличия общих промежуточных продуктов) и управляемых очень тонкими регуляторными механизмами.
Метаболизм минеральных веществ
Относительное содержание. Различные элементы, встречающиеся в живых организмах, перечислены ниже в убывающем порядке в зависимости от их относительного содержания: 1) кислород, углерод, водород и азот; 2) кальций, фосфор, калий и сера; 3) натрий, хлор, магний и железо; 4) марганец, медь, молибден, селен, йод и цинк; 5) алюминий, фтор, кремний и литий; 6) бром, мышьяк, свинец и, возможно, некоторые другие.
Кислород, углерод, водород и азот – это те элементы, из которых построены мягкие ткани тела. Они входят в состав таких соединений, как углеводы, липиды, белки, вода, диоксид углерода и аммиак. Элементы, перечисленные в пп. 2 и 3, находятся в организме обычно в виде одного или нескольких неорганических соединений, а элементы пп. 4, 5 и 6 присутствуют только в следовых количествах и потому их называют микроэлементами.
Распределение в организме. Кальций. Кальций присутствует главным образом в костной ткани и в зубах, преимущественно в виде фосфата и в небольших количествах в виде карбоната и фторида. Поступающий с пищей кальций всасывается в основном в верхних отделах кишечника, имеющих слабокислую реакцию. Этому всасыванию (у человека здесь всасывается всего 20–30% кальция пищи) способствует витамин D. Под действием витамина D клетки кишечника вырабатывают особый белок, который связывает кальций и облегчает его перенос через стенку кишечника в кровь. На всасывание влияет также присутствие некоторых других веществ, в особенности фосфата и оксалата, которые в малых количествах способствуют всасыванию, а в больших, наоборот, подавляют его.
В крови около половины кальция связано с белком, остальное составляют ионы кальция. Соотношение ионизированной и неионизированной форм зависит от общей концентрации кальция в крови, а также от содержания белка и фосфата и концентрации водородных ионов (рН крови). Доля неионизированного кальция, на которую влияет уровень белка, позволяет косвенным образом судить о качестве питания и об эффективности работы печени, в которой идет синтез плазменных белков.
На количество ионизированного кальция влияют, с одной стороны, витамин D и факторы, воздействующие на всасывание, а с другой – паратиреоидный гормон и, возможно, также витамин D, поскольку оба эти вещества регулируют как скорость отложения кальция в костной ткани, так и его мобилизацию, т.е. вымывание из костей. Избыток паратиреоидного гормона стимулирует выход кальция из костной ткани, что приводит к повышению его концентрации в плазме. Изменяя скорости всасывания и экскреции кальция и фосфата, а также скорости образования костной ткани и ее разрушения, эти механизмы строго контролируют концентрацию кальция и фосфата в сыворотке крови. Ионы кальция играют регулирующую роль во многих физиологических процессах, в том числе в нервных реакциях, мышечном сокращении, свертывании крови. Выведение кальция из организма происходит в норме в основном (на 2/3) через желчь и кишечник и в меньшей степени (1/3) – через почки.
Фосфор. Метаболизм фосфора – одного из главных компонентов костной ткани и зубов – во многом зависит от тех же факторов, что и метаболизм кальция. Фосфор в виде фосфата присутствует в организме также в сотнях различных физиологически важных органических эфиров. Паратиреоидный гормон стимулирует выведение фосфора с мочой и выход его из костной ткани; тем самым он регулирует концентрацию фосфора в плазме крови.
Натрий. Натрий – главный катион внеклеточной жидкости – вместе с белком, хлоридом и бикарбонатом играет важнейшую роль в регулировании осмотического давления и pH (концентрации водородных ионов) крови. В клетках, напротив, содержится очень мало натрия, так как они обладают механизмом для выведения ионов натрия и удержания ионов калия. Весь натрий, превышающий потребности организма, очень быстро выводится через почки.
Поскольку во всех процессах выделения натрий теряется, он должен постоянно поступать в организм с пищей. При ацидозе, когда необходимо, чтобы из организма выводились большие количества анионов (например, хлорида или ацетоацетата), почки предотвращают чрезмерную потерю натрия благодаря образованию аммиака из глутамина. Выведение натрия через почки регулируется гормоном коры надпочечников альдостероном. Под действием этого гормона в кровь возвращается достаточно натрия для поддержания нормального осмотического давления и нормального объема внеклеточной жидкости.
Суточная потребность в хлористом натрии составляет 5–10 г. Эта величина возрастает при поглощении больших количеств жидкости, когда усиливается потоотделение и выделяется больше мочи.
Калий. В отличие от натрия, калий содержится в клетках в больших количествах, но во внеклеточной жидкости его мало. Главная функция калия – регулирование внутриклеточного осмотического давления и поддержание кислотно-щелочного равновесия. Он также играет важную роль в проведении нервного импульса и во многих ферментных системах, в том числе и в тех, которые участвуют в мышечном сокращении. Калий широко распространен в природе, и его много в любой пище, так что спонтанно калиевая недостаточность возникнуть не может. В плазме концентрация калия регулируется альдостероном, стимулирующим его экскрецию с мочой.
Сера. С пищей сера поступает в организм главным образом в составе двух аминокислот – цистина и метионина. На конечных этапах метаболизма этих аминокислот сера высвобождается и в результате окисления переводится в неорганическую форму. В составе цистина и метионина сера присутствует в структурных белках. Важную роль играет также сульфгидрильная (–SH) группа цистеина, от которой зависит активность многих ферментов.
Большая часть серы выводится с мочой в виде сульфата. Небольшое количество экскретируемого сульфата обычно связано с органическими соединениями типа фенолов.
Магний. Метаболизм магния сходен с метаболизмом кальция, и в виде комплекса с фосфатом этот элемент тоже входит в состав костной ткани. Магний присутствует во всех живых клетках, где он функционирует как необходимый компонент многих ферментных систем; эта его роль была убедительно продемонстрирована на примере углеводного обмена в мышцах. Магний, как и калий, широко распространен, и вероятность возникновения его недостаточности очень мала.
Железо. Железо входит в состав гемоглобина и других гемопротеинов, а именно миоглобина (мышечного гемоглобина), цитохромов (дыхательных ферментов) и каталазы, а также в состав некоторых ферментов, не содержащих гемогруппы. Всасывается железо в верхних отделах кишечника, причем это единственный элемент, всасывающийся только тогда, когда его запас в организме полностью исчерпан. В плазме железо транспортируется в соединении с белком (трансферрином). Через почки железо не выводится; избыток его накапливается в печени в соединении с особым белком (ферритином).
Микроэлементы. У каждого микроэлемента, присутствующего в организме, своя особая функция, связанная с тем, что он стимулирует действие того или иного фермента или как-либо иначе на него влияет. Цинк необходим для кристаллизации инсулина; кроме того, он является компонентом карбоангидразы (фермента, участвующего в транспорте диоксида углерода) и некоторых других ферментов. Молибден и медь – тоже необходимые компоненты различных ферментов. Иод требуется для синтеза трииодтиронина, гормона щитовидной железы. Фтор (входящий в состав зубной эмали) способствует предотвращению кариеса.
Использование метаболитов
Углеводы. Всасывание. Моносахариды, или простые сахара, высвобождающиеся при переваривании углеводов пищи, переходят из кишечника в кровоток в результате процесса, называемого всасыванием. Механизм всасывания представляет собой сочетание простой диффузии и химической реакции (активного всасывания). Одна из гипотез, касающихся природы химической фазы процесса, предполагает, что в этой фазе моносахариды соединяются с фосфорной кислотой в реакции, катализируемой ферментом из группы киназ, после чего проникают в кровеносные сосуды и здесь высвобождаются в результате ферментативного дефосфорилирования (разрыва фосфатной связи), катализируемого одной из фосфатаз. Именно активным всасыванием объясняется то, что разные моносахариды всасываются с разной скоростью и что углеводы всасываются даже тогда, когда уровень сахара в крови выше, чем в кишечнике, т.е. в условиях, когда естественно было бы ожидать их перемещения в обратном направлении – из крови в кишечник.
Механизмы гомеостаза. Поступающие в кровоток моносахариды повышают уровень сахара в крови. При голодании концентрация глюкозы в крови колеблется обычно от 70 до 100 мг на 100 мл крови. Этот уровень поддерживается с помощью механизмов, называемых механизмами гомеостаза (самостабилизации). Как только уровень сахара в крови в результате всасывания из кишечника повышается, в действие вступают процессы, выводящие сахар из крови, так что уровень его колеблется не слишком сильно.
Подобно глюкозе, все прочие моносахариды поступают из кровотока в печень, где превращаются в глюкозу. Теперь они неотличимы как от глюкозы, которая всосалась, так и от той, что уже была в организме, и подвергаются тем же метаболическим превращениям. Один из механизмов гомеостаза углеводов, функционирующий в печени, – это гликогенез, посредством которого глюкоза переходит из крови в клетки, где превращается в гликоген. Гликоген хранится в печени до тех пор, пока не произойдет снижение уровня сахара в крови: в этой ситуации гомеостатический механизм вызовет распад накопленного гликогена до глюкозы, которая вновь поступит в кровь.
Превращения и использование. Поскольку кровь поставляет глюкозу во все ткани тела и все ткани используют ее для получения энергии, уровень глюкозы в крови снижается главным образом за счет ее использования.
В мышцах глюкоза крови превращается в гликоген. Однако мышечный гликоген не может быть использован для получения глюкозы, которая перешла бы в кровь. В нем заключен запас энергии, и скорость его использования зависит от мышечной активности. В мышечной ткани содержатся два соединения с большим запасом легко доступной энергии в форме богатых энергией фосфатных связей – креатинфосфат и аденозинтрифосфат (АТФ). При отщеплении от этих соединений их фосфатных групп высвобождается энергия для мышечного сокращения. Чтобы мышца вновь могла сокращаться, эти соединения должны быть восстановлены в своей исходной форме. Для этого требуется энергия, которую поставляет окисление продуктов распада гликогена. При мышечном сокращении гликоген превращается в глюкозофосфат, а затем – через ряд реакций – во фруктозодифосфат. Фруктозодифосфат распадается на два трехуглеродных соединения, из которых после ряда этапов образуется сначала пировиноградная кислота, а в конечном итоге – молочная кислота, как об этом уже говорилось при описании метаболизма углеводов. Это превращение гликогена в молочную кислоту, сопровождающееся высвобождением энергии, может происходить в отсутствие кислорода.
При недостатке кислорода молочная кислота накапливается в мышцах, диффундирует в кровоток и поступает в печень, где из нее вновь образуется гликоген. Если кислорода достаточно, то молочная кислота в мышцах не накапливается. Вместо этого она, как это описано выше, полностью окисляется через цикл трикарбоновых кислот до диоксида углерода и воды с образованием АТФ, который может быть использован для сокращения.
Метаболизм углеводов в нервной ткани и эритроцитах отличается от метаболизма в мышцах тем, что гликоген здесь не участвует. Однако и здесь промежуточными продуктами являются пировиноградная и молочная кислоты, образующиеся при расщеплении глюкозофосфата.
Глюкоза используется не только в клеточном дыхании, но и во многих других процессах: синтезе лактозы (молочного сахара), образовании жиров, а также особых сахаров, входящих в состав полисахаридов соединительной ткани и ряда других тканей.
Гликоген печени, синтезируемый при всасывании углеводов в кишечнике, служит самым доступным источником глюкозы, когда всасывание отсутствует. Если этот источник оказывается исчерпанным, в печени начинается процесс глюконеогенеза. Глюкоза образуется при этом из некоторых аминокислот (из 100 г белка образуется 58 г глюкозы) и нескольких других неуглеводных соединений, в том числе из глицериновых остатков нейтральных жиров.
Некоторую, хотя и не столь важную, роль в метаболизме углеводов играют почки. Они выводят из организма избыток глюкозы, когда ее концентрация в крови слишком высока; при меньших концентрациях глюкоза практически не выводится.
В регулировании метаболизма углеводов участвует несколько гормонов, в том числе гормоны поджелудочной железы, передней доли гипофиза и коры надпочечников.
Гормон поджелудочной железы инсулин снижает концентрацию глюкозы в крови и повышает ее концентрацию в клетках. По-видимому, он стимулирует также и запасание гликогена в печени. Кортикостерон, гормон коры надпочечников, и адреналин, вырабатываемый мозговым веществом надпочечников, воздействуют на метаболизм углеводов, стимулируя распад гликогена (главным образом в мышцах и печени) и синтез глюкозы (в печени).
Липиды. Всасывание. В кишечнике после переваривания жиров остаются главным образом свободные жирные кислоты с небольшой примесью холестерина и лецитина и следами жирорастворимых витаминов. Все эти вещества очень тонко диспергированы благодаря эмульгирующему и солюбилизирующему действию солей желчных кислот. Солюбилизирующее действие обычно связывают с образованием нестойких химических соединений между жирными кислотами и солями желчных кислот. Эти комплексы проникают в клетки эпителия тонкого кишечника и здесь распадаются на жирные кислоты и соли желчных кислот. Последние переносятся в печень и вновь секретируются с желчью, а жирные кислоты вступают в соединение с глицерином или холестерином. Образовавшиеся реконструированные жиры поступают в лимфатические сосуды брыжейки в форме млечного сока, т.н. «хилуса». Из сосудов брыжейки хилус по лимфатической системе через грудной проток поступает в кровеносную систему.
После переваривания пищи содержание липидов в крови возрастает приблизительно от 500 мг (уровень при голодании) до 1000 мг на 100 мл плазмы. Присутствующие в крови липиды представляют собой смесь жирных кислот, нейтральных жиров, фосфолипидов (лецитина и кефалина), холестерина и эфиров холестерина.
Распределение. Кровь доставляет липиды в разные ткани тела и прежде всего в печень. Печень обладает способностью модифицировать поступающие в нее жирные кислоты. Это особенно выражено у видов, запасающих жиры с высоким содержанием насыщенных или, наоборот, ненасыщенных жирных кислот: в печени этих животных соотношение насыщенных и ненасыщенных кислот изменяется таким образом, что отлагающийся жир по своему составу соответствует жиру, свойственному данному организму.
Жиры в печени либо используются для получения энергии, либо переходят в кровь и доставляются ею в разные ткани. Здесь они могут включаться в структурные элементы тканей, но большая их часть отлагается в жировых депо, где они хранятся до тех пор, пока не возникнет потребность в энергии; тогда они снова переносятся в печень и подвергаются здесь окислению.
Метаболизм липидов, как и углеводов, регулируется гомеостатически. Механизмы гомеостаза, воздействующие на липидный и углеводный обмен, видимо, тесно связаны, поскольку при замедлении метаболизма углеводов усиливается метаболизм липидов, и наоборот.
Превращения и использование. Четырехуглеродные кислоты – ацетоуксусная (продукт конденсации двух ацетатных единиц) и -гидроксимасляная – и трехуглеродное соединение ацетон, образующийся при отщеплении одного атома углерода от ацетоуксусной кислоты, известны под общим названием кетоновых (ацетоновых) тел. В норме кетоновые тела присутствуют в крови в небольших количествах. Избыточное их образование при тяжелом диабете ведет к повышению их содержания в крови (кетонемия) и в моче (кетонурия) – это состояние обозначают термином «кетоз».
Белки. Всасывание. При переваривании белков пищеварительными ферментами образуется смесь из аминокислот и небольших пептидов, содержащих от двух до десяти остатков аминокислот. Эти продукты всасываются слизистой кишечника, и здесь гидролиз завершается – пептиды также распадаются до аминокислот. Поступившие в кровь аминокислоты смешиваются с находящимися здесь такими же аминокислотами. В крови содержится смесь из аминокислот, поступивших из кишечника, образовавшихся при распаде тканевых белков и синтезированных организмом заново.
Синтез. В тканях непрерывно идет распад белков и их новообразование. Содержащиеся в крови аминокислоты избирательно поглощаются тканями как исходный материал для построения белков, а из тканей в кровь поступают другие аминокислоты. Синтезу и распаду подвергаются не только структурные белки, но и белки плазмы крови, а также белковые гормоны и ферменты.
Во взрослом организме аминокислоты или белки практически не запасаются, поэтому удаление аминокислот из крови происходит с такой же скоростью, как и их поступление из тканей в кровь. В растущем организме формируются новые ткани, и на этот процесс расходуется больше аминокислот, чем поступает в кровь за счет распада тканевых белков.
Печень участвует в метаболизме белков самым активным образом. Здесь синтезируются белки плазмы крови – альбумины и глобулины – а также собственные ферменты печени. Так, при потере плазменных белков содержание альбумина в плазме восстанавливается – за счет интенсивного синтеза – довольно быстро. Аминокислоты в печени используются не только для образования белков, но подвергаются также расщеплению, в ходе которого извлекается заключенная в них энергия.
Превращения и использование. Если аминокислоты используются в качестве источника энергии, то отщепляемая от них аминогруппа (–Nh3) направляется на образование мочевины, а не содержащий азота остаток молекулы окисляется приблизительно так же, как глюкоза или жирные кислоты.
Так называемый «орнитиновый цикл» описывает, как происходит превращение аммиака в мочевину. В этом цикле аминогруппа, отщепившаяся от аминокислоты в форме аммиака, присоединяется вместе с диоксидом углерода к молекуле орнитина с образованием цитруллина. Цитруллин присоединяет второй атом азота, на этот раз от аспарагиновой кислоты, и превращается в аргинин. Далее аргинин подвергается гидролизу с образованием мочевины и орнитина. Орнитин может теперь вновь вступить в цикл, а мочевина выводится из организма через почки как один из конечных продуктов метаболизма.
Список литературы
Ленинджер А. Основы биохимии, тт. 1–3. М., 1985
Страйер Л. Биохимия, тт. 1–3. М., 1985
Марри Р., Греннер Д., Мейес П., Родуэлл В. Биохимия человека, тт. 1–2. М., 1993
Албертс Б., Брей Д., Льюс Д. и др. Молекулярная биология клетки, тт. 1–3. М., 1994
Для подготовки данной работы были использованы материалы с сайта http://bio.freehostia.com
Дата добавления: 01.11.2006
www.km.ru





|
|
..:::Счетчики:::.. |
|
|
|
|
|
|
|
|