Реферат: Фауна и растительность Свердловской области. Реферат фауна
Реферат - Фауна Евразии - Теоритическая география
Рыжая лисица Лисица относится к роду волчьих. Она - очень хитрый и искусный охотник, который может найти жертву по ее следам. Чаще всего ее добычей становятся кролики или мыши. Увидев их, лиса начинает погоню и быстро догоняет свою жертву. У рыжей лисицы хорошо развит слух. Также у лисицы теплая красива шкура и красивый хвост. Живут в норах. Важный объект пушного промысла. Факты Название: Vulpes. Размеры: Тело 45-85 см в длину; хвост 30-50 см в длину. Среда обитания: леса, сухая открытая местность; все чаще города и поселки в Евразии, северной Африке, Азии, Северной Америке, Австралии. Размножение: самка рожает четыре или пять детенышей; период беременности семь-девять недель. Питание: грызуны, кролики, черви, насекомые, плоды, падаль, отбросы.Полевая мышь
Несмотря на свои маленькие размеры, эти грызуны заняты приемом пищи почти целый день. Полевая мышь живет на открытых местностях обычно среди злаковых растений, зернами которых она питается. Но так же эта мышь хорошо лазает по деревьям в поисках листьев и фруктов. Свои гнезда мыши устраивают под корнями деревьев и под землей.
Факты
Название: Clethrionomys glareolus.
Размеры: до 10 см в длину.
Среда обитания: кустарники и леса в Евразии, южной Скандинавии, западной России.
Размножение: самка рожает два или три раза от трех до пяти детенышей за раз, с весны до осени; период беременности 18-20 дней.
Питание: семена, плоды, почки.
Большой баклан
Баклан - семейство пеликанообразных. Это большие птицы, которые питаются рыбой, Они могут нырять на глубину более 23 метров и оставаться в воде целую минуту. Большой баклан - это самый многочисленный вид. Крылья относительно короткие, хвост длинный, жесткий. Стайные птицы, гнездятся колониями. Полет быстрый, но неманевренный.
Факты
Название: Phalacrocorax carbo.
Размеры: до 95 см в длину; весит до 2,3 кг.
Среда обитания: скалистые берега, а также озера и реки в Северной Америке , Евразии, Африке и Австралии.
Размножение: самка откладывает 3-4 яйца в гнездо обычно сделанное в скалах и выложенное морскими водорослями.
Питание: рыба.
Ласка
Ласка - самый маленький из собачьих млекопитающих. На охоте ласка часто нападает на жертву, которая значительно больше их самих, например кролик. Но в основном добычей ласки становятся полевые мыши. Выслеживая мышей, она приподнимается на задние лапки, чтобы дальше видеть свою добычу. У ласк когтистые лапы, небольшой хвост, вытянутое тело и ярко-белая грудь.
Факты
Название: Mustela nivalis.
Размеры: 20-25 см в длину.
Среда обитания: лиственные леса в Евразии, Северной Америке, северной африке.
Размножение: самка рожает одного или двух детенышей ежегодно; гнезда ласк находятся под землей между корнями деревьев, в отверстиях стен, между грудами камней.
Питание: мелкие грызуны, кролики.
Синица
Эта энергичная певчая птица - близкий родственник североамериканской гаички. Она летает вокруг деревьев и очень часто висит на ветках головой вниз, выискивая гусениц и других насекомых. Для птицы, гнездящейся на деревьях, синица откладывает очень много яиц - до 16 в одном гнезде. У синицы есть синие пятно на голове, а также у нее синие крылья и хвост.
Факты
Название: Parus caeruleus.
Размеры: до 12 см в длину; весом до 10 грамм.
Среда обитания: леса в Евразии, на севере Африки.
Размножение: гнездится в дуплах, самка откладывает 6-12 яиц.
Питание: мелкие насекомые, такие как гусеницы и тля; а также семена.
Ястреб-перепелятник
Ястреб-перепелятник охотится на мелких птиц. Как и многие хищные птицы, он нападает на свою жертву с неба на очень высокой скорости. Крылья ястреба перепелятника - широкие, крапчатые - позволяют ему поворачивать с очень высокой точностью. Также у него длинные, острые когти. Ястреб, поймав свою жертву в воздухе, убивает ее только на земле.
Факты
Название: Accipiter nisus.
Размеры: самец достигает 40 см в длину; самка - 30 см.
Среда обитания: леса, степи в Евразии, на северо-западе Африки.
Размножение: в гнездо из веток самка откладывает до 5 яиц и высиживает их около 5 недель.
Питание: птицы, мелкие млекопитающие.
Олень
Этого красивого лесного оленя относят к ночным животным, которые бодрствуют ночью, а отдыхают днем. Летом его шкура приобретает пеструю окраску. Как и у всех жвачных животных, рога самца-оленя намного больше рогов самок. С помощью рогов олень отпугивает врагов.
Факты
Название: Dama.
Размеры: 1,2-1,5 м в длину.
Среда обитания: лиственные леса в Евразии
Размножение: самка рожает одного детеныша в июне или в июле; уже скоро он может бегать.
Питание: трава, ягоды; желуди зимой и осенью.
www.ronl.ru
Реферат - Животный мир Донбасса
Реферат выполнил: ученик 7-Г класса ОШ I-III ступеней №20 Демидов Олег
Донецк 2003 г.
Заповедник «Хомутовская степь» как форма сохранения естественной природы Донбасса
Организация заповедных территорий и объектов — наиболее совершенная и полная форма сохранения в естественном состоянии как отдельных компонентов природы, прежде всего растительного и животного мира, так и природных комплексов в целом с присущими им естественными природными процессами. Особенно актуальна проблема сохранения уникальных и типичных ландшафтов, генофонда растительного и животного мира, а также благоприятных экологических условий на значительных пространствах, в условиях Донецкой области — наиболее экологически напряженном регионе Украины. Располагаясь в Степной зоне, регион практически не имеет целинных степных участков. Отдельные природные территории и комплексы испытывают огромное воздействие промышленности и сельского хозяйства.
В наше время уже не найдешь больших целинных степей. Человек освоил их, вспахал и засеял. Но есть на территории Донецкой области уникальный кусочек целины — Хомутовская степь, показывающей нам как выглядело Приазовье в далеком прошлом. Заповедник с таким названием был организован в 1926 году. Сейчас он является центральным отделением Украинского степного природного заповедника, в состав которого входят также и другие чудесные степные массивы — Каменные Могилы, Меловая флора и Михайловская целина. Это все, что осталось от некогда необъятного Дикого поля, занимавшего южную часть Украины. На его просторах появлялись, развивались и исчезали разнообразные народы — скифы, сарматы, печенеги, половцы и многие другие, которые волнами прокатывались бескрайней степью, оставляя на курганах часовых времени — каменных баб. К сожалению, эти идолы прошлого не смогли уберечь плодородную степь от человека, распахавшего земли. На сегодня площадь заповедника составляет 1030,4 га, причем 90 из них являются абсолютно заповедным участком, которого никогда не касался плуг и на котором 70 лет не выпасался скот и не проводилась косовица. На его территории сохранились и охраняются растения группы равнинного разнотравно — типчаково — ковыльной степи. Тут прорастает 604 вида цветочных растений и папоротников — трав, кустарников и деревьев. Из них 19 — эндемичные, т.е. распространенные на очень ограниченной территории. В заповеднике зарегистрировано также 59 видов мхов, 46 — лишайников и свыше 270 грибов. Наиболее типичными и массовыми представителями флоры являются: катран, кермек, ковыли, волошки, кашка, пырей, костер, тырса, горицвет, тюльпан, фиалки, миндаль, дрок, карагана, васильки, мятлик, шалфей, резак, скабиоза и еще сотни других растений. Каждое из них по своему и в свое время украшает степь. Только в заповеднике можно наблюдать за сезонными изменением настоящей степи — его весенним пробуждением, буйством жизни летом и осенне — зимним засыпанием. Например в начале мая, на огромных площадях неописуемую красоту создает цветущий пион. Кажется, что море огня разлилось на зеленом ковре весенней степи. Разнообразна фауна заповедника: волк, лиса, енотовидная собака, заяц, хомяк, байбак, суслик, полевка, мыши, хорек, ласка, ящерица, полоз, медянка, уж, гадюга степная, жабы, лягушки и т.д. Пернатый мир Хомутовской степи представлен более 80 видами птиц. Куропатка, перепел, фазан, утки, дрофа, стрепет, жаворонки, скворцы, луни, степной орел, сорокопут-жулан, славки, синица, иволга, кобчик, пустельга, осоед, камышевка, выпь, бекас, чибис, зимородок, цапля, кукушка — вот наиболее часто встречающиеся птицы. В настоящее время в Хомутовской степи, в рамках научного эксперимента, начаты работы по разведению коней и их выпасу. Это связано с проблемой накопления, особенно на участках с абсолютно заповедным режимом, подстилки из толстого слоя омертвевших растений. Это произошло вследствие изменений в экосистеме, а именно исчезновение или резкое сокращение представителей животного мира. Некогда здесь ходили стада тарпанов, туров, сайгаков, косуль. Позже Донские казаки выпасали тут табуны лошадей. Употребляя траву они сдерживали образование подстилки. Поэтому возрождение коневодства в Украинских степях дает уникальную возможность возрождения красочных ландшафтов. С возвышенности на левом берегу реки Грузский Еланчик открывается вид на огромный сохранившийся массив первозданной степи. Ощущение вечности, охватывающее экскурсантов, усиливается каменными бабами — изваяниями древних зодчих. Их тут более 20. Гости Хомутовской степи с интересом посещают музей, в котором квалифицированные сотрудники расскажут и покажут массу интереснейшей информации. Выйти в степь и увидеть какой она была в далекие прошедшие времена, это прекрасно!
Разнообразие животного мира Донбасса
Распределение животных подчинено закону широтной зональности, что предопределяет возникновение фаунистических комплексов в пределах каждой зоны. Животный мир территории Донецкой области относится к Голарктической зоогеографической области. Формирование современных фаунистических комплексов, так же как и растительности, началось в неогене. Возрастающая хозяйственная деятельность человека привела к уничтожению многих видов животных.
В лесах Донецкой области обитают лось, косуля, дикий кабан, лесная куница, заяц — русак, красная белка, каменная куница, европейская норка, барсук, ласка, горностай, крот (Великоанадольское лесничество). В степи водятся байбаки, землеройки, ежи, суслики, полевые мыши, степной хорек, енотовидные собаки, слепушонки, красная лисица, реже волки, ящерица прыткая, ящерица-веретенница, четырехполосный полоз, уж обыкновенный,
уж водяной, медянка, степная гадюка, желтобрюх.
Достаточно разнообразен видовой состав птиц. В донецких степях можно встретить серую куропатку, дрофу, стрепета, жаворонка, скворца, черноголовую овсянку, орлана — могильника, скопу, филина, степного орла, орлана — белохвоста, аиста, ласточку деревенскую, стрижа. В полезащитных полосах и в лесных массивах обитают иволги, коршуны, горлицы, дятлы, соловьи, синицы, грачи, галки, серые вороны, сороки, козодои, кукушки, пеночки. По берегам рек и водоемов, на косах Азовского моря много водоплавающей птицы — гусей, уток, степных куликов, чибисов, краснозобых казарок, лебедей-шипунов, кроншнепов, чаек — черноголовок, чаек — хохотуний, крячек. В степных водоемах живут болотная черепаха, озерная лягушка, прудовая лягушка, некоторые моллюски — катушка, прудовик, лужанка, раки и около 30 видов рыб. Животный мир Азовского моря насчитывает около 80 видов рыб. Наибольшее значение имеют лещ, судак, белуга, сельдь, тарань, хамса, большая камбала, бычок.
Енотовидная собака.
Приземистое животное, длина тела которого равна 65—80 см; имеет сравнительно короткий, пушистый хвост, уши маленькие, закругленные, мех густой пышный, сверху бурый, снизу более светлый с желтоватым оттенком. Этот зверек имеет «бакенбарды» — удлиненные волосы по бокам головы. Часть головы от глаз до ушей у него светлая, щеки темные. Отпечатки лап округлые с поперечником около 4,5—5 см, на них отчетливо видны следы коротких тупых когтей. Отпечатки располагаются в два ряда по ломаной линии. Длина шага 15—30 см.
Обитательница Дальнего Востока, енотовидная собака в настоящее время акклиматизировалась и в нашем регионе. Места обитания: пойменные луга с кустарником, берега озер, перелески, светлые смешанные леса. Питается разнообразной животной пищей, в том числе птицами, яйцами и птенцами. В пойменных угодьях наносит большой вред — уничтожает кладки водоплавающих птиц. С наступлением морозов впадает в дремотное состояние, забившись в стог сена, под кучу хвороста, в бурелом, в свою, барсучью или лисью нору. В оттепели пробуждается и бродит вблизи своего убежища и по проезжим дорогам. Гон в феврале-марте; щенки (обычно 6—8) появляются через 2 мес.
Байбак европейский.
Подвид с сокращающейся численностью. Длина тела 585 мм, хвоста — 145мм. Обитатель степной зоны.
В ХVIII — ХIХ вв. байбак европейский широко населял открытые ландшафты Русской равнины. Его ареал уменьшился во много раз и стал мозаичным вследствие распашки целинных степей и неумеренной охоты. Наиболее критическим положение было в 40 — 50-х гг. ХХ века, но принятые меры охраны обеспечили сохранение подвида. Нынешние места обитания байбака — это клочки нераспаханных злаково-разнотравных степей, сильно деградировавших от перевыпаса скота, сохранившиеся по оврагам и балкам, крутым склонам речных долин с характерными меловыми обнажениями. Пригодные для поселения байбака земли составляют незначительную долю общей площади, освоенной посевами. На посевах многолетних трав байбак задерживается на более длительные сроки. Площадь обособленных поселений байбаков не превышает десятков гектаров, редко достигает немногих сотен. Поселения обычно отстоят друг от друга на несколько километров. Поэтому средняя плотность, рассчитанная на площадь занимаемого видом района, составляет всего единицы особей на 1 км кв., но в пределах колоний она может достигать 200 — 300 зверьков на 1 кв.км.
Каменная куница.
Каменная куница–белодушка немного меньше, чем лесная куница; хвост относительно длиннее, шерсть у нее более грубая, пятно на горле белого цвета. Обитает в бедных лесом скалистых горах, каменоломнях, заброшенных постройках, кустарниках, в пещерах, часто вблизи селений. На деревья поднимается редко. Ловит грызунов; охотнее, чем лесная куница, поедает ягоды и фрукты.
Лебедь-шипун.
Крупная птица. Длина крыла самца 53—63 см, самки— 53—60. Масса взрослой птицы 8—13 кг. Взрослые самец и самка белые. Основание и края надклювья, уздечка и нижняя челюсть черные, остальные части клюва оранжево-красные. В верхней части клюва у основания лба характерный нарост, или «шишка», более крупный у самцов и несколько меньший у самок.
Ноги черные.
Молодые птицы серые, с буроватым оттенком. Основание клюва темное, остальные его части буровато-серые. На лбу малозаметная шишка. С возрастом окраска становится ярче, окончательный наряд лебедь приобретает лишь после 3-й или 4-й линьки.
От других видов лебедей отличается тем, что, плавая, часто изгибает шею в виде буквы S, а клюв и голову держит наклонно к воде. Шея более толстая и кажется короче, чем у лебедя-кликуна. Шипун часто сушит крылья, приподняв их над спиной и несколько распушив перья. В полете кричит редко. Голос низкий, сиплый, слегка дребезжащий. С воды поднимается с трудом, пробегая длинную дорожку и помогая себе ударами крыльев и лап. Полет ровный, стремительный. При взмахах крыльев перья издают ритмичные скрипящие звуки, слышимые на большом расстоянии.
Медянка.
Медянка, змея из семейства ужовых. Длиной до 70 см. Иногда встречаются более крупные особи. Окраска спины от серой до медно-красной, с 2-4 продольными рядами черноватых пятнышек. Зрачок у медянки круглый (у обыкновенной гадюки, которая похожа по окраске на медянку, зрачок вертикальный, узкий). Обитает в лесной зоне, чаще встречается на прогреваемых солнцем опушках, на вырубках, облесенных полянах и в зарослях подлеска.
Перечисленные животные – лишь маленькая капля в огромном океане, который называется природа.
К сожалению, деятельность человека губительно отражается на животном и растительном мире нашего края. Если люди не начнут более бережно и осмотрительно относится к природе, которая взрастила и их самих, то будущие поколения будут рассматривать ныне еще существующих животных только на картинках. Рассказы о них будут казаться чудесной сказкой. Уже сейчас создаются заповедники — образцы нетронутой, дикой природы, которые по праву называют природными лабораториями. Они особенно нужны нам сейчас, когда мы должны понять направления изменений природной Среды под влиянием деятельности человека и отыскать пути наиболее бережного и разумного использования ее богатств.
Наши потребности растут, и от природы мы берем все больше и больше. Поэтому необходимо обеспечить восполнение природных ресурсов по принципу расширенного воспроизводства. В этом главная задача охраны природы сегодня.
Список литературы
1. Человек и мир зверей./ К. А. Татаринов «Вища школа» Львов 1980.
2. География родного края. Учебное пособие. / Сост.: Н. А. Швец Н. Я. Швец. Донецк 1997.
3.Этот прекрасный загадочный мир/ Сост.: Н. В. Якименко Киев «Реклама» 1979.
Другие источники информации:
1.Сайт Экологического информационного центра г. Донецк
2.Сайт общественной организации «Донецкое аграрное развитие» г. Донецк
3.Сайт «Украинский сервер охотника»
www.ronl.ru
Фауна и растительность Свердловской области
Министерство сельского хозяйства РФ
ФГОУ ВПО «УрГСХА»
Кафедра экологии и зоогигиены
Реферат
по региональной экологии
на тему: «Фауна и растительность
Свердловской области»
Выполнил: Прилепо М.Н.
ФВМ 223
Проверил: Неверова О.П.
К.б.н., доцент каф. экологии
и зоогигиены
Екатеринбург 2008
Содержание
1. Растительность
1.1 Основные закономерности в распределении растительности
1.2 Лесная растительность
1.3 Лесостепная растительность
1.4 Растительность горной полосы Урала
1.5 Интразональная растительность
1.6 Растительные ресурсы
2. Животный мир
2.1 Главные особенности животного мира Свердловской области
2.2 Промысловые животные
2.3 Пресмыкающиеся
2.4 Обитатели рек и озер
Литература
1. Растительность
1.1 Основные
закономерности в распределении растительности
Растительность в
Свердловской области, ее характер и размещение зависят от климата, от его
широтных, долготных и высотных изменений, которые выражаются в смене
температурного режима и общего режима увлажнения, т.е. в изменении баланса
тепла и влаги. Это важнейший фактор формирования растительности как на равнинах,
так и в горах. Вместе с тем заметное влияние на растительность оказывает
рельеф, состав горных пород, грунтовые воды, почвы, деятельность человека и
другие факторы. В связи с воздействием ряда факторов в размещении
растительности на территории области можно выделить несколько основных
закономерностей:
1. Избыточно влажный климат обуславливает преобладание на
территории области лесной растительности. В условиях недостаточного увлажнения
на юго-востоке и юго-западе области формируются участки степной растительности.
2. Широтные изменения климата приводят к смене основных
типов растительности, которые образуют растительные зоны и подзоны на равнинах
области.
3. Зональные типы растительности изменяются с запада на
восток, что обусловлено секторными и барьерными различиями климата.
4. Высотные изменения климата в горной полосе приводят к
смене типов растительности с высотой и выделению нескольких высотных
растительных поясов, которые наиболее ярко выражены в среднегорьях Северного
Урала.
5. На фоне зональных типов растительности распространены
интразональные, формирование которых связано с особым режимом увлажнения. В
местах избыточного переувлажнения атмосферными или грунтовыми водами, в
пониженных элементах рельефа формируется влаголюбивая растительность пойм,
болот и водная.
6. Растительность области значительно изменена человеком в
процессе хозяйственного освоения территории.
1.2 Лесная
растительность
растительность
фауна лес зверь
Таежные хвойные леса
разделяются на темнохвойные и светлохвойные. На их распространение влияют
секторные и барьерные изменения климата, а также характер почвогрунтов,
определяемый геологическим строением, рельефом и грунтовыми водами.
В горной полосе и в
западных предгорьях Урала широко распространены темнохвойные леса, представленные
растительными сообществами, в составе которых преобладают ель, пихта и кедр.
Основная лесообразующая порода – ель. Участие пихты древостое незначительное.
Экологические условия данного района – повышенное увлажнение, более
благоприятный температурный режим – способствуют распространению здесь этих
влаголюбивых теневыносливых деревьев. Севернее широты Нижнего Тагила в составе
лесов распространен кедр сибирский, однако, чистых кедровников в области
немного. Все они объявлены памятниками природы.
Более континентальный
климат восточных предгорий Урала и равнинной западносибирской его части
вызывает смену темнохвойных лесов светлохвойными. Древостой в них образован в
основном сосной с незначительной примесью лиственницы. Широкое распространение
здесь сосны связано с ее большей приспособляемостью к различным условиям
обитания. Она растет на бедных песчаных и каменистых почвах, на крутых
каменистых склонах и даже на скалах, хорошо переносит переувлажнение и растет
на болотах, где образует низкопродуктивные насаждения из угнетенных,
низкорослых деревьев.
В области большие площади
заняты производными вторичными березовыми и осиново-березовыми лесами на месте
хвойных лесов. Береза тоже отличается исключительной неприхотливостью к
неблагоприятным условиям обитания, поэтому первой занимает вырубки, гари, луга,
конкурируя даже с сосной.
На юго-востоке области
довольно большие площади заняты березовыми лесами, представляющими собой
коренной зональный тип растительности, который, сочетаясь с луговыми степями,
образует у южных границ области лесостепную растительность.
Средняя лесистость по
области, по разным источникам, составляет около 70%. Наиболее высока лесистость
в горной полосе – от 70 до 80%, меньше всего на юго-востоке области – около 25-30%
и на юго-западе – 35% (в Артинском районе). Даже в окрестностях Екатеринбурга
лесистость довольно высока: от 66% на северо-западе до 42% на юго-востоке.
Характер лесов меняется
на территории области с севера на юг вместе с климатическими условиями, поэтому
выделяют подзональные типы растительности.
Северо-таежные леса в верховьях Пелыма и Лозьвы – это
редкостойные лишайниково-моховые и кустарничковые, на равнинах большей частью
заболоченные долгомошные и сфагновые леса. В наземном покрове таких лесов много
багульника, встречается карликовая береза. Древостой среднетаежных лесов становится
более высоким и сомкнутым. Меняется наземный покров. Распространены
зеленомошные, травяно-кустарничковые леса с брусникой и черникой, лесными
травами в наземном покрове. В южно-таежных лесах в наземном покрове
увеличивается доля лесного разнотравья, появляется богатый подлесок из
можжевельника, рябины, жимолости обыкновенной и голубой, кизильника, ракитника,
липы. Южно-таежные леса отличаются высокой полнотой и производительностью. На
юго-западе области, в западных предгорьях и на Уфимском плато выделяются широколиственно-хвойные
леса – пихтово-еловые с участием широколиственных пород в древостое и
подлеске (липа, клен, ильм). В подлеске и травостое много видов кустарников и
трав, характерных для европейских широколиственных лесов. Среди них лещина
(орешник) и бересклет, сныть и копытень европейский.
Березовые и
осиново-березовые леса на юго-востоке области отличаются большой разреженностью, осветленностью,
густым травяным покровом из злаков и разнотравья. Уникальный пример таких лесов
– 200-летняя Шипеловская роща в Белоярском районе. На песчаных грунтах, обычно
в приречных местностях, березовые леса сменяются сосновыми борами
(Припышминские боры в Талицком и Камышловском районах).
1.3 Лесостепная
растительность
На юго-востоке области,
южнее реки Пышмы, развита степная растительность, которая вместе с лесной
образует переходную зону лесостепей. В этой зоне обычны островные
осиново-березовые леса, которые называют «колками». Они занимают чаще всего
плоские понижения, западины на междуречьях, нередко заболоченные в центральной
части. В травостое этих лесов лесное разнотравье, немало степных видов, по
опушкам разрастается шиповник, ива, черемуха. Между лесными участками раньше
располагались луговые степи и остепненные луга. Для них характерно сочетание
лугового разнотравья из клеверов, чины, мышиного горошка, тысячелистника и
других видов с сухолюбивыми степными злаками (перистый ковыль, типчак, овсец,
степная тимофеевка) и степными травами (полынь, люцерна, лабазник).
Участки лесостепей есть и
на юго-западе области. Это Красноуфимская лесостепь в Юрюзано-Сылвенской
депрессии. Появление здесь лесостепных ландшафтов также связано с
климатическими причинами, но не зональными, а барьерными: Уфимское плато
перехватывает часть осадков с запада. На особенности почвенно-растительного
покрова оказывают влияние также и подстилающие карбонатные породы. В березовых
лесах Уфимской лесостепи встречаются широколиственные породы деревьев (дуб и
липа).
Лесостепные районы нашей
области значительно изменены хозяйственной деятельностью человека. Луговые
степи и часть лесов распаханы. Оставшиеся леса выполняют регулирующие и
полезащитные функции: зимой задерживают снег на полях, летом ослабляют действие
засух, которые случаются в этих районах.
1.4 Растительность
горной полосы Урала
Горная полоса Урала
характеризуется высотными изменениями растительности, которые позволяют
выделить в горах три пояса. Горные леса, поднимаясь по склонам гор до высоты
750-800 м, образуют широкий горно-таежный пояс. Горная тайга западного склона и
осевых хребтов Урала образована темнохвойными еловыми с пихтой и кедром лесами.
На менее увлажненных восточных склонах их сменяют сосновые леса.
Выше расположен переходный
от горной тайги к горным тундрам пояс редколесий из угнетенных елей, кедров,
лиственниц. Местами распространены березовые криволесья. Среди редколесий и
криволесий встречаются кусты можжевельника, ольхи, участки горных лугов с
высоким травостоем из злаков и разнотравья (вейник, герань, лютик, черемица,
аконит, ветреница пермская, живокость и др.) и горных тундр. Этот пояс называют
также подгольцовым. Он поднимается по склонам гор примерно до 800-900 м.
Редколесья сменяются
поясом горных тундр. В его составе моховые, лишайниковые, каменистые тундры, которые
прерываются каменистыми россыпями, выступами скал-останцев. Тундровые растения
представлены зарослями карликовой березки («ерники»), голубикой, водяникой,
дриадой, осокой, пушицей, цветковыми растениями.
Весь набор высотных
поясов характерен для среднегорий Северного Урала – и для осевых хребтов, и для
таких горных массивов, как Конжаковский Камень, Денежкин Камень, Чистоп и др.
Невысокие горные хребты и увалы Среднего Урала до самых вершин покрыты
горно-таежными лесами, лишь на самых высоких хребтах и вершинах появляются
разреженная растительность, скалы и каменистые россыпи.
1.5 Интразональная
растительность
Значительные площади в
области занимает болотная, луговая и пойменная растительность. Верховые болота
на плоских водоразделах междуречий на северо-востоке области покрыты сплошным
ковром сфагновых мхов, на этом фоне растут багульник, карликовая березка,
подбел, морошка, клюква, пушица. Нередко можно встретить угнетенные,
низкорослые сосенки и березки. Наиболее значительные массивы верховых болот
находятся среди северо- и среднетаежных лесов. В южной тайге их меньше (самое
крупное из них – Аятское болото).
Низинные болота
распространены по долинам рек, особенно в поймах, на пониженных участках
междуречий, где близки грунтовые воды. В растительности низинных болот
господствуют зеленые (гипновые) мхи и осоки. Часто по низинным болотам растут
ивы, ольха, мелкие березы.
Кроме верховых и низинных
болот, развиты и переходные.
Поймы крупных и мелких
рек заняты лугово-кустарниково-лесной растительностью. Это периодически
затопляемые лиственные леса из тополя, осины, ольхи, черемухи и кустарников
(смородины, малины). Такую растительность называют «уремой», а леса с
преобладанием ольхи – сероольшанниками. Среди трав в этих зарослях много
таволги, крапивы, аконита, звездчатки, сныти. На деревьях иногда встречается
хмель. Леса сменяются злаково-разнотравными лугами, иногда сырыми и
заболоченными. В травостое лугов – мятлик, костер, пырей, купальница, щавель,
подмаренник и др. На сырых и заболоченных лугах растут осоки, вейник, щучка.
В озерах и реках
разрастается водная растительность. У берегов это болотные травы, осоки,
стрелолист, сусак; на большей глубине – тростник, рогоз, камыш. Еще дальше –
пояс белых кувшинок и кубышек с плавающими листьями и красивыми белыми и
желтыми цветами. Есть и свободно плавающие растения: ряска, водокрас,
пузырчатка и др. По каменистым руслам и отмелям рек часто встречается
нардосмия, образующая значительные заросли из крупных листьев.
1.6 Растительные
ресурсы
Разнообразны и богаты
растительные ресурсы области. Самая важная их часть – лесные ресурсы. Леса
области поделены на три группы. Первая группа – неэксплуатируемые леса вокруг
городов и населенных пунктов (лесопарки и зеленые зоны), вокруг курортов;
водоохранные леса горных районов и вдоль рек и озер; полезащитные лесные
полосы. Леса второй группы – частично эксплуатируемые, в основном резервные.
Третья группа – эксплуатируемые леса.
Леса области значительно
истощены лесозаготовками. На сегодня реальные запасы лесных ресурсов, пригодных
для заготовки древесины, невелики – их хватит лет на десять. Восстановление же
лесов ведется недостаточно. Леса области дают не только древесину. Заготовка
лекарственных трав, ягод, грибов, орехов имеет не менее важное значение. Важны
рекреационные функции лесов, водоохранные, почвозащитные. Лесные и пойменные
луга – это пастбища и сенокосы.
2. Животный мир
2.1 Главные
особенности животного мира Свердловской области
Животный мир очень тесно
связан с зонально-климатическими условиями и некоторыми другими природными
особенностями территории.
1. На территории области преобладают
представители таежной фауны, а на юге появляются и лесостепные обитатели.
2. Большинство видов животных Урала
относится к европейско-западносибирской фауне, но среди представителей
животного мира есть и типично сибирские виды (колонок, бурундук, сибирская
косуля, темнозобый дрозд), и типично европейские (европейская норка и черный,
или лесной, хорь).
3. Животный мир Урала практически не
отличается от фауны соседних равнин. Лишь в горах, в горных тундрах на гольцах
Урала он имеет некоторые особенности.
4. Своеобразен животный мир болот и
речных долин, рек и озер.
5. Животный мир региона испытывал и
испытывает значительное воздействие человека. Численность некоторых видов
животных, особенно промысловых, сильно менялась за прошедшие столетия.
Интенсивная антропогенная нагрузка на окружающую природу – вырубка лесов,
распашка земель, загрязнение воздуха, вод, почвы – вызывает негативные
изменения в животном мире, обедняя его и приводя к сокращению численности
животных. Мероприятия по охране животного мира приводят к положительным
результатам: численность одних видов восстановлена, а других – выросла. Многие
виды внесены в красную книгу Среднего Урала (выхухоль, медянка, обыкновенный
еж, европейская норка, черный аист, могильник, орлан-белохвост, сапсан,
южнорусский тарантул, северный кожанок, филин и др.).
2.2 Промысловые
животные
Среди таежных животных
Свердловской области много ценных промысловых пушных зверей. Прежде всего это
соболь, колонок и куница. Ценятся также белка и лисица, ареалы которых заходят
и в лесостепные районы. Очень ценный мех имеют выдра, норка и бобр. Охотятся
также на рысь, росомаху, волка и медведя.
К началу XX в. в результате хищнической добычи
на грани исчезновения оказались соболь и бобр, бурый медведь и выдра. Лишь
запрет на отстрел некоторых видов, организация заповедников, заказников,
акклиматизация и реакклиматизация позволили восстановить ареалы и численность
таких животных, как соболь и бобр.
Соболь принадлежит к отряду хищников,
семейству куньих. Предпочитает глухие, захламленные участки ельников и
кедровников. Питается мелкими грызунами и хищниками. Красивый и прочный мех
соболя ценится очень высоко. К середине XX в. Численность соболя в области достигла критической
отметки. Поэтому в конце 40-х – начале 50-х гг. сюда было завезено и выпущено в
районе Ивделя 227 особей баргузинского соболя, из них 126 с наиболее ценным темным
мехом. В дальнейшем завезенный соболь широко расселился на севере области – в
Ивдельском, Карпинском, Серовском и Гаринском районах. Численность его, по
оценкам охотоведов, достигла уровня XVII в. С 1969 г. в области разрешен лицензионный отстрел соболя.
Близкий родственник
соболя – куница лесная живет во всех лесах нашей области, за исключением
северо-восточных районов. Серовато-коричневый мех куницы также высоко ценится.
Интересно, что на Урале, где соболь и куница живут вместе, встречается их
помесь, называемая кидасом (кидусом).
Есть в наших водоемах
хищники, приспособленные для полуводного образа жизни. Это выдра и
норка. Длина тела выдры 55-95 см, хвоста – 26-55 см. Норка помельче: длина тела до 45 см, хвост – до 20 см. Питаются эти хищники рыбой и другими водными животными. Интересно, что европейская норка постепенно вытесняется
другим видом – норкой американской, завезенной и акклиматизированной у
нас.
К хищникам относится
также хорь светлый – житель лесостепи, маленький горностай, белый
зимой, и ласка. Это обитатели лесостепи и тайги. Ласка – самый маленький
уральский хищник: длина ее тела от 11 до 26 см, хвоста – до 4 см. Ласка ловит за сезон тысячи мышей и нападает даже на крупных лесных птиц – тетеревов. Иногда
она заглядывает и в курятники.
На севере области в
таежных лесах водится зверь, похожий на лохматую собаку, и на огромную куницу.
Это росомаха. Длина ее тела 70-105 см, хвоста – 16-23 см, телосложение массивное. Шерсть густая, грубая. Окраска светло- или темно-бурая
со светлой полосой по бокам. Бегает быстро, как волк, на деревья лазает как
рысь. Все, кроме лося да медведя, боятся этого хищника. Ловит росомаха и
оленей, и косуль, и зайцев и птиц, но предпочитает падаль. Летом может
полакомиться ягодой.
Барсук – родственник росомахи и куницы, но в
отличие от них он не всеяден. Любит грибы и ягоды, корневища и орехи, мелких
позвоночных и насекомых. Живет в норах по берегам рек и по оврагам. Окраска
спины и боков серебристо-серая, низ черноватый, по бокам головы, от носа к
ушам, идут темные полосы. Сало барсука обладает лечебными свойствами.
Бобр – один из наиболее крупных
представителей отряда грызунов. Ценится своим густым, высоким мехом. Ведет
полуводный образ жизни, селится по берегам тихих лесных речек. Бобр также был
на грани исчезновения. Работы по восстановлению его численности начались в1957
г. В разных районах области было организовано 9 заказников. В результате в
начале 90-х гг. численность бобра приблизилась к 10000 особей. Лицензионный
отстрел его разрешен с 1972 г.
В 30-х гг. началась
работа по акклиматизации на Урале ондатры. В Гаринском и Таборинском
районах было выпущено 2088 особей. Ондатра хорошо прижилась во многих водоемах,
даже в озере Шарташ, которое находится в городской черте Екатеринбурга.
Постоянная численность бурых
медведей в области около 1000 особей. Их отстрел производится по особым
разрешениям в количестве 60 голов в год. Численность волков достигает
900 особей, ежегодно отстреливают 400-500 голов. Численность рыси – около 1000
особей, ее отстреливают, но немного.
Производится лицензионный
отстрел лося и косули. Лось (сохатый) – самый крупный зверь уральского
леса. Ростом лось выше крупной лошади. Высота его в холке – до 2,3 м. Длина тела – до 3 м, масса – до 570 кг. Длинные ноги позволяют лосю передвигаться по глубокому
снегу и по болотам. Зимой он питается побегами и корой ив, осины, рябины,
сосны, летом – травами, листьями, сочными корнями. Крепкие копыта, огромная
сила и страшные рога-лопаты заставляют уважать лося даже хищников. Только
больные да молодые лоси могут не выдержать нападения волчьей стаи.
Сибирская косуля – небольшой олень, средний вес ее 50 кг. Предпочитает светлые сосновые и березовые леса с травяными лугами.
Дикий северный олень по Уральским горам доходил когда-то
до самого юга области. Теперь северный олень встречается только на севере
территории. На сегодня осталось 150-200 голов диких оленей и около 500
одомашненных.
К промысловым птицам
относится боровая дичь – глухарь, тетерев и рябчик, а также водоплавающие птицы
(утки) и кулики.
2.3 Пресмыкающиеся
В наших лесах их немного.
Это гадюка обыкновенная, уж и живородящая ящерица. Самый крупный среди них – уж,
до метра длиной. Окраска у него черная, на голове по бокам желтоватые
пятна. Уж не ядовит, но, если неумело схватить его в руки, – укусит, и больно.
Похожа на ужа, но меньше его – гадюка. Это очень ядовитая змея. Укус
крупной гадюки может быть смертельным. Окрашена бывает в темно- и светло-серый
цвет. По всему хребту у нее идет зигзагом черная полоса. Гадюка водится по
лугам, вблизи болот и на каменистых склонах гор. В лесу можно встретить
небольшую и неядовитую змею – медянку, цвет которой бывает от желтого до
медно-красного.
2.4 Обитатели рек и
озер
Среди обитателей
ихтиофауны Урала наиболее интересны рыбы. В реках области водятся разнообразные
виды как ценных промысловых, так и менее значимых с экономической точки зрения
«сорных» рыб.
Практически повсеместно
распространены щука, окунь, ерш, плотва, лещ и густера, язь, налим, гольян. Для
южной части области характерны также пескарь, елец и линь. Редко встречается
таймень (семейство лососевых).
В реках Обь-Иртышского
бассейна – в Тавде, Пелыме, Сосьве, Туре – и их притоках широко распространена
пелядь (сырок) из семейства сиговых. Она имеет промысловое значение и поэтому
была разведена во многих озерах Зауралья. Есть в этих реках и ценные сибирские
виды – нельма, тугун (сосьвинская селедка). В горных реках встречается хариус
сибирский.
Для рек Волго-Камского
бассейна – Чусовой, Сылвы, Уфы – и их притоков характерны европейские виды:
красноперка, голавль, жерех, уклей, вьюн. В Чусовую раньше заходила даже
белорыбица (нельма).
В озерах Зауралья
распространен карась – золотой и серебряный. Основное местообитание золотого
карася – заболоченные, заросшие озера. В реках он встречается очень редко.
Диапазон местообитания серебряного карася шире: он живет и в озерах, и в реках.
В озерах, там, где вода более богата кислородом, обитают также плотва, окунь,
щука, лещ, линь.
В водоемы Свердловской
области из ладожского, Онежского и Псковско-Чудского озер были переселены такие
ценные промысловые рыбы, как сиг и рипус (ладожская форма ряпушки). Рипус
хорошо прижился в озерах Таватуе, Аятском, Шарташе. В 30-х гг. уловы рипуса в
Таватуе достигали 726 ц в год, а в Шарташе – 200 ц.
Во многих районах области
в прудах выращивают карпа (сазана).
Литература
1. Капустин В.Г., Корнев И.Н. Свердловская
область: природа, население, хозяйство, экология: Учебное пособие для учащихся
старших классов по курсу «География Свердловской области». – 2-е изд., испр. и
доп. – Екатеринбург: Изд-во Дома учителя, 2000. – 300 с.: ил.
2. Большая серия знаний. Страны. Народы.
Европа и Россия. / сост. В.Б. Новичков. – М.: ООО «Мир книги»; Современная
педагогика, 2003.
www.neuch.ru
Реферат: Фауна прямокрылых Республики Тува
Федеральное агентство по образованию ТОМСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ (ТГУ) Институт биологии, экологии, почвоведения, сельского и лесного хозяйства Кафедра защиты растений КУРСОВАЯ РАБОТА ФАУНА ПРЯМОКРЫЛЫХ (ORTHOPTERA) РЕСПУБЛИКИ ТУВА Научный руководитель канд. биол. наук, доцент ____________С. В. Лукьянцев Автор работы ____________Ч. К. Борарган Томск 2010. СОДЕРЖАНИЕ ВВЕДЕНИЕ.. 3 1.ФИЗИКО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА ТЕРРИТОРИИ ТУВЫ. 5 1.1. Климат. 6 1.2 Рельеф.. 8 1.3 Растительные сообщества. 10 2. ХАРАКТЕРИСТИКА ФАУНЫ ПРЯМОКРЫЛЫХ ТУВЫ... 12 ЗАКЛЮЧЕНИЕ.. 15 СПИСОК ЛИТЕРАТУРЫ... 16 ПРИЛОЖЕНИЕ.. 17 ВВЕДЕНИЕ Прямокрылые насекомые, особенно саранчовые, являются одним из важнейших компонентов многих ландшафтов, в первую очередь травянистых. Хорошо известно, что их численность и биомасса могут достигать колоссальных значений. В такие годы эти насекомые могут съедать почти весь прирост зеленой фитомассы. Особенно важно, что во многих случаях одной из причин, обусловливающих нарастание численности, может быть тот или иной тип деятельности человека, например, выпас, сенокошение, создание полей многолетних трав (Сергеев,1999). В горах Южной Сибири прямокрылые связаны в первую очередь с разнообразными степными и опустыненными ландшафтами, а также с горными лугами, тундрами и лесами (значительной меньшей степени). Полное отсутствие местных стадных видов, периодически размножающихся в массе и способных расселяться на сравнительно большие расстояния, заставляет исследователей характеризовать в первую очередь многовидовые сообщества этих насекомых (Сергеев,1999). Актуальность данной работы состоит в том, что прямокрылые ― главным образом облигатные фитофаги или зоофитофаги, составляющие значительную долю растительноядных форм в составе фауны Тувы. Известно, что продуктивность травянистых растений в изучаемом районе невысока, тогда как одна особь саранчового, в зависимости от размеров тела,потребляет от 90 до 280 мг сухой фитомассы в сутки (Пшеницына, 1997).Следовательно, нагрузка с их стороны на растительный покров здесь значительна. Недавними исследованиями для степей Внутренней Монголии показано возрастание роли саранчовых за счет уменьшения количества в таких экосистемах крупных травоядных животных (Локвуд и др., 1997). Прямокрылые участвуют в биологическом круговороте веществ и в почвообразовательных процессах, что было показано на примере луговых степей Западной Сибири (Стебаев, Гукасян, 1963). Эти насекомые служат пищей как для позвоночных (особенно это относится к птицам), так и для беспозвоночных животных (Правдин, 1974). Такие прямокрылые, как саранчовые, часто более известны в качестве вредителей сельского хозяйства. (Мищенко, 1972). В связи с сокращением сельскохозяйственных работ в последние десятилетия увеличились площади основных мест откладки кубышек (залежи и т.д.). В некоторые годы развитию прямокрылых в Туве благоприятствуют и погодные условия. Комплекс этих факторов способствует возникновению массовых вспышек размножения этих насекомых, которые при отсутствии необходимых мероприятий могут наносить экономически ощутимые потери для сельского хозяйства (Мищенко,1972). Целью данной работы являлось составление обзора фауны прямокрылых Тувы. Для достижения этой цели необходимо решить следующие задачи: 1. Дать биогеографическую характеристику региона 2. Составить предварительный список видов прямокрылых Тувы 3. Проанализировать ландшафтно-биотопическое распределение видов. При написании курсовой работы был проведен анализ литературных источников. Изучение данной проблемы отражено в трудах таких ученых, как Бей-Биенко Г.Я (1980), Мищенко Л.Л. (1951),Бережков. Р.П (1956), Сергеев М.Г. (2001), Стебаев И.В. (1963) и др. Структура и объем работы обусловлены предметом, целями и задачами исследования. Курсовая работа состоит из введения, двух глав, состоящих из трех параграфов, заключения и списка литературы. Во введении раскрывается актуальность обозначенной темы, определяются цель и задачи. В первой главе рассматривается общая характеристика Республики Тува: географическое положение, климат, рельеф, растительные сообщества. Во второй главе изучается краткий обзор фауны прямокрылых; основные очаги распространения прямокрылых в Республике Тувы. В заключение работы сделаны выводы и составлен предварительный список прямокрылых Тувы. 1.ФИЗИКО-ГЕОГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА ТЕРРИТОРИИ ТУВЫ. Республика Тува самая молодая республика в составе Российской Федерации. Республика Тува расположена на юге Восточной Сибири, в центральной части Азиатского материка между 49 45 – 53 46 Северной широты и 88 49 – 98 56 восточной долготы. На западе граничит с Республикой Алтай, на северо-западе и севере - с Красноярским краем и Республикой Хакасия, на северо-востоке – с Иркутской областью и Республикой Бурятия, на юге и востоке – с Монголией. (Рисунок 1) 
Рисунок 1. Географическое положение Республики Тува Трудно назвать другой район России, который бы на сравнительно небольшой территории имел такое разнообразие природных условий, как Тува. Расположенная в центре азиатского материка, на стыке Сибири и Центральной Азии, она сочетает в себе элементы этих двух, таких разных природных районов. На тундровых плоскогорьях с вечной мерзлотой и в песчаной пустыне; из тайги выйти в степи, с ледников и снежных полей высокогорья спуститься в межгорные котловины с обширными камышовыми плавнями. Однако тувинская горная тундра не похожа на тундру Полярного Урала, тувинская тайга выглядит не так, как тайга соседнего Восточного Саяна, а камышовые плавни Терехольской котловины совсем иные, чем плавни дельты Волги. Все это создает неповторимый облик тувинской природы, которую можно сравнить разве что с удивительной природой Уссурийского края, сочетающей в себе признаки и формы северной тайги и южных тропических лесов. Тува такой же, как Уссурийский край, уникальный район страны, обладающий рядом природных особенностей, не встречающихся больше нигде в пределах России (Сергеев, 1986). В соответствии с разнообразием природных условий и естественных ресурсов, характером экономического развития и транспортных связей Туву можно разделить на 4 части: центральную, западную, южную и восточную. С позиции природных условий географическое положение республики выгодное. Она расположена на стыке сибирских таёжных и центрально-азиатских пустынно-степных ландшафтов – в широкой полосе гор и межгорных равнин. Территория республики равна 171.3 тыс. кв. км., она простирается с востока на запад более чем на 700 км., с севера на юг в восточной части на 450 км., а в центральной части на 100 км., плотность населения составляет 1.8 человека на 1 кв. км. (Сергеев,1989). 1.1. Климат В отличие от Саян или Алтая климат Тувы характеризуется резкой континентальностью, гораздо меньшим количеством осадков, как зимой, так и летом, преобладанием солнечных дней и сильной зависимостью от рельефа (абсолютной высоты и ориентировки горных хребтов). Удаленность Тувы от морей и океанов служит причиной сильной континентальности климата, выражающейся в большом колебании сезонных и суточных температур. Днем можно изнывать от жары, а с заходом солнца замерзать от холода. Хребты Саян, закрывая Туву с запада, севера и северо-востока подковообразной горной цепью высотой свыше 2000 м, защищают ее территорию от господствующих здесь в теплое время года западных и северо-западных ветров, несущих влагу. В Туву воздушные массы приходят сильно обедненные влагой, которую они оставляют на наветренных склонах Алтая и Саян, где количество осадков доходит до 1000 мм в год. Подветренныё склоны хребтов Тувы и котловины получают всего 200-300 мм осадков в год. Но и в самой Туве малое количество осадков распределено неравномерно. Большую их часть получают наветренные склоны, меньшую - подветренные. Наветренные склоны в Туве, как правило, имеют северную экспозицию, а подветренные - южную, станет ясна причина резкого отличия климатических условий на склонах разной экспозиции. Северные наветренные склоны в основном покрыты лесами, южные, получающие меньше влаги и одновременно больше солнца, часто заняты лесостепью и степью. Контраст этот разителен (Сергеев, 1999) Для Тувы характерна продолжительная, малоснежная, очень холодная зима. Средняя температура воздуха в январе колеблется в Тувинской котловине от 30 градусов до 35 градусов ниже нуля. Зима длится с ноября по апрель. Снежный покров устанавливается в конце октября и достигает 15-20 см., в горах до 1-2 метров, сходит в середине апреля в горах–в мае. Весна (апрель – май) – поздняя, короткая, ясная, ветреная и сухая. Март здесь можно назвать зимним месяцем, особенно в горах. Даже в мае горы и высокие горные распадки покрыты снегом. Лето – сухое, теплое, в межгорных котловинах даже жаркое, в горах – короткое и прохладное. Средняя температура в равнинной части – 20-30 С, (в горах – 13-15 С), в отдельные дни может подниматься выше 35 С. Самый теплый месяц – июль. Осень - сухая, солнечная, самое лучшее время года. В сентябре бывают возвраты тепла, которые сопровождаются ясной солнечной погодой и вторичным цветением травянистой растительности в межгорных котловинах (Сергеев,1989) Сезонная разница температур достигает 54 градусов Цельсия. В горных районах суточный перепад температур достигает 30 градусов. Очень своеобразен климатический режим котловин. Закрытые со всех сторон от ветров, они сильно прогреваются днем солнцем. Восходящие потоки нагретого воздуха размывают облачность над ними. И даже когда в окружающих горах ненастье, над котловинами ясное небо и кружат стаи орлов в восходящих потоках воздуха. Большое влияние на климат и в целом на природные условия Тувы оказывает абсолютная высота местности. В Туве можно выделить три вертикальных климатических пояса: низкогорье (от 520 до 1000-1250 м), среднегорье (от 1000-1250 до 1900-2000 м) и высокогорье (свыше 2000 м). Восточная часть Тувы более влажна не только потому, что имеет наветренную экспозицию, но и потому, что почти вся целиком расположена в поясе среднегорья и высокогорья. Низинные равнины западной Тувы значительно суше; здесь преобладают степи, в то время как почти вся восточная часть покрыта лесами. Охарактеризовать климат поясов можно и по сезонам. Весна в низкогорье наступает, как правило, во второй декаде апреля, а в среднегорье - в первой декаде мая. Нарастание тепла идет очень интенсивно, особенно в котловинах. Средние температуры июля достигают в низкогорье 19-20°, августа + 15-18°. В среднегорье лето более короткое и прохладное. Средняя температура июля - самого теплого месяца - составляет + 13-15°. При этом максимальные температуры могут превышать +30, а минимальные - доходить до -3°. Осадки распределяются по сезонам неравномерно. Минимальное количество их (от 4 до 10% годовых) приходится на зиму и весну. В летние месяцы над Тувой получает развитие циклональная деятельность, что обусловливает максимум осадков в этот сезон. Среднегодовой уровень осадков в котловинах составляет 200-230 мм, в среднегорье - до 400 мм, причем на летние месяцы приходится 60-65% этого количества. Ненастье обычно длится не больше 1-2 дней. Затяжные дожди бывают только в восточных районах Тувы. В связи с развитием циклонов в летние месяцы увеличивается и облачность, достигая максимума в июне (60-70%). В теплый период над Тувой господствуют западные и северо-западные ветры. Средняя скорость ветра в течение лета удерживается на уровне 2-3 м/сек. Получают развитие горно-долинные ветры: днем - к вершинам, ночью - в долины. Их постоянство служит надежным признаком устойчивой ясной погоды.(Сергеев, 1986). 1.2 Рельеф Тува имеет горно-котловинный рельеф: горы занимают примерно 82 процента территории, а 18 процентов приходится на межгорные котловины (сухостепная Тувинская, полупустынная Убсу-Нурская, таежно-лесные Тоджинская и Тере-Хольская). Средняя высота котловин – 520-1200м над уровнем моря. На территории республики протекает около 8 тысяч рек, среди которых наиболее крупными являются Енисей и Хемчик, имеются 11 солено-грязевых и более 8 пресных озер общей площадью более 300 кв. км. Имеются почти все виды минеральных вод, используемых в бальнеологических целях (Сергеев, 1989) Самая примечательная особенность рельефа Тувы - его чашеобразность. Центр республики представляет собой группу котловин, окруженную со всех сторон высокими горными хребтами. Протягиваясь по границам Тувы, они образуют две, подковообразные горные дуги: Саянскую и Танну-Ольскую, обращенные выпуклостью к северу. Северная дуга состоит из сложной горной системы Западного Саяна. С востока к ним примыкает меридиональная горная цепь Чарм-Тайга, включающая в себя массивы пика Топографов (3044 м) и горы Мунку-Сасан (3126 м). На западе северная дуга смыкается с Шапшальским меридиональным хребтом (3614 м), который в юго-восточном направлении переходит в хребет Цаган-Шибэту (3495 м), уходящий в Монголию. К ним примыкает с юга хр. Лихачева (4020 м), имеющий также меридиональное направление. Таким образом, крупная подковообразная горная дуга закрывает Туву с запада, севера и востока. (Сергеев, 1989) В пределах Тувы, вдоль ее южной границы с Монголией, расположена меньшая дуга, состоящая из хребтов Западного (3061 м) и Восточного (2591 м) Танну-Ола и нагорья Сангилен (3274 м). С запада эта меньшая дуга, повторяющая форму Саянской, смыкается с хр. Цаган-Шибэту, на востоке - с хр. Хан-Тайга (2616 м), имеющим меридиональное простирание. Наличие этих двух крупных орографических дуг оказывает решающее влияние на климат, растительность и весь природный облик Тувы. Кроме того, в пределах Тувы между реками Бий-Хем и Каа-Хем протягивается в широтном направлении система хребтов Академика Обручева, которая смыкается на западе с системой Западного Саяна. В пределах Тувы известно около 45 вершин высотой более 3000 м. Большинство из них сосредоточено в юго-западной части республики. По хребтам Лихачева, Цаган-Шибэту, Танну-Ола и Сангилен проходит участок мирового водораздела между бассейном Ледовитого океана и бессточной областью Центральной Азии. Здесь также проходят южная граница сибирских таежных ландшафтов и северная граница пустынь Монголии. Здесь находится и предельная отметка, являющаяся и высшей точкой Восточной Сибири, – гора Монгун-Тайга 3976 м (рисунок 2), самая низкая точка – устье реки Хемчик 508м над уровнем моря). Территория Тувы прошла продолжительную геологическую историю и сложена комплексом разнообразных по составу и возрасту горных пород. Современный рельеф практически сформировался в четвертичный период (около 1,8 млн. лет). В результате новейших горообразовательных процессов (альпийская складчатость) древняя выровненная поверхность местами поднялась на разные высоты, и возник нынешний облик поверхности территории республики. Эти процессы сопровождались извержениями вулканов и землетрясениями. Тува - единственный в своем роде географический район на земном шаре, откуда дальше до океана, чем из любой другой точки планеты. 
Рисунок 2. Гора Монгун-Тайга 1.3 Растительные сообщества Разнообразие природных условий определило богатство ее растительного мира. Для северо-востока и востока республики характерны таежные леса, сухие степи (в Тувинской котловине) и полупустынные ландшафты (в Убсунурской котловине), субальпийские и альпийские луга, горные тундры (в высокогорье) (Сергеев, 1986). Флора и фауна Тувы поражает разнообразием, здесь много эндемичных растений. Характер растительности Тувы имеет ярко выраженную вертикальную поясность. Верхняя граница леса в северо-восточных районах (Западный и Восточный Саян) проходит на высоте 1800 м, в районе хр. Академика Обручева и Каа-Хемского нагорья - на высоте 2000 м, в хребтах Сангилен и Танну-Ола она поднимается до 2200 м, а на юго-западе Тувы, в массиве Монгун-Тайга,- даже до 2400 м. Территории, лежащие выше этой границы, заняты тундрово-луговым высокогорным ландшафтом. Здесь моховые, болотистые тундры перемещаются нагромождениями валунов и россыпями камней; отдельные пятна альпийских лугов чередуются с участками каменисто-щебнистой тундры. Значительны площади кустарников, березки круглолистной, спиреи, ивы, даурского рододендрона. У верхней границы леса к ним присоединяются низкие, отдельно стоящие кедры с флагообразными вершинами с подлеском из жимолости, можжевельника алтайского, растущих на сплошном ковре зеленого мха и лишайников (Сергеев, 1986) Леса покрывают почти половину территории Тувы. На хвойные породы приходится 95 % от всей площади лесов. Основная древесная порода – сибирская лиственница, занимающая 69 % площади и достигающая иногда гигантских размеров. Следующим по распространенности идет кедр (сосна сибирская), незначительна, но постоянна примесь ели, пихты. Изредка встречаются береза и осина. Сосна обыкновенная произрастает лишь в нескольких местах небольшими сообществами, самое крупное из которых – Балгазынский сосновый бор, расположенный примерно в 100 км. на юг от Кызыла. Считается, что в северном полушарии это самый южный сосновый бор. Около 3 млн. гектаров территории Тувы покрыто кедровыми лесами (11 % всех кедровых лесов России). Лиственница не дает большой тени, поэтому тувинские леса светлы и просторны, между мощными красноватыми стволами видно далеко. Почва в лесу покрыта, как правило, богатыми травами, которые особенно сильны на полянах (еланях). Много бурелома. Подлесок образуют бузина, шиповник, рябина, смородина, крыжовник. Особенно много смородины и крыжовника в камнепадах по склонам ущелий, на солнечной стороне. В изобилии другие ягоды: брусника, голубика, костяника, черника, земляника. Травяной ярус состоит из кислички, ириса, вейника лесного, мятлика и др. Нижняя граница леса проходит на высоте от 1100 (хр. Уюкский) до 1400 м. Если не считать безлесных горных вершин и водоемов, то свободную от леса территорию Тувы занимают степи. В основном это сухие степи, а на юге – опустыненные степи и небольшие участки песков (Сергеев, 1986). 2. ХАРАКТЕРИСТИКА ФАУНЫ ПРЯМОКРЫЛЫХ ТУВЫ Общая характеристика отряда прямокрылых: насекомые от средних до крупных размеров, как правило, с удлиненными задними ногами, имеющими мощные, приспособленные для прыжков бедра (Рисунок 3) Почти у всех форм большая переднеспинка, продолженная на боках вниз, так что образуется широкий воротничок позади головы. Голова крупная, с удлиненными антеннами, хорошо развитыми глазами и грызушим ротовым аппаратом простого типа. У многих видов крылья большие и функционируют; надкрылья (уплотненные передние крылья) всегда кожистые, а задние крылья мембранозные, веерообразные, складывающиеся в покое. У других видов крылья могут быть укорочены или полностью отсутствовать. К этому отряду относятся кобылки (Acrididae), кузнечики (Tettigonidae), сверчки (Gryllidae), медведки (Gryllotalpidae) и прыгунчики (Tetrigidae), в совокупности представляющие собой группу форм самых разных размеров, внешнего вида, окраска и образа жизни (Росс.Г и др., 1985) 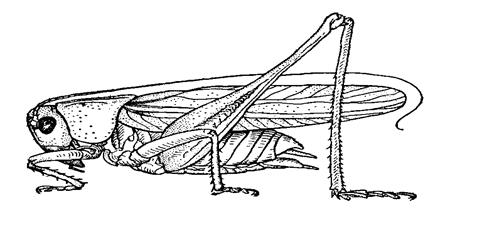
Рисунок 3. Внешнее строение прямокрылых (Тыщенко, 1986) Прямокрылые, особенно саранчовые обитают во всех степных, полупустынных и пустынных местообитаниях, однако наиболее обильны они в степях и полупустынях. В соответствии с наиболее часто встречающимися в литературе по фауне прямокрылых северной Азии взглядами на территории Тувы проходит граница между Скивской и Сахаро-Гобийской зоогеографическими подобластями Палеарктической области. Степная часть Тувы входит в Сарматская провинцию Скивской подобласти, а южная горная часть - в Монгольская провинцию Сахаро-Гобийской подобласти (Лачинский и др., 2002; Сергеев, 1986). Фауна прямокрылых Сарматской провинции насчитывает 104 вида, а Монгольской провинции – 41 вид (Лачинский и др., 2002). В настоящее время для территории Тувы и юга Сибири в литературе указывается 145 видов прямокрылых из 8 семейств (Лачинский и др., 2002; Сергеев, 1986), из них 60 видовточно отмечены на территории Тувы (Приложение). Очаги разнообразия прямокрылых в Туве являются: Центрально-Тувинская котловина, Убсунурская котловина, степная Улуг-Хемская котловина, Центральная Тува. Наиболее подробно изучены Центрально-Тувинская и Убсунурская котловина (Сергеев, 1986). Распределение прямокрылых определяется типом растительности в каждом типе растительных сообществ формируется свой комплекс видов (сообщество) прямокрылых (Сергеев, 1986). В центрально-Тувинской котловине сообщества прямокрылых существенно беднее как по видовому разнообразию так по численности, чем другие районы. На плакорах и террасах наиболее обычны степные монгольско-сибирские виды (например,Myrmeleotettix palpalis).В поймах доминируетChorthippus albomargynatus(Сергеев, 1986). Естественно, что по примыкающим к горным котловинам хребтам встречаются более разнообразные сообщества, характеризующиеся преобладанием собственно степных, северостепных и полизональных форм (например,Omocestus haemorrhoidalis) (Сергеев, 1986). В целом для сообществ прямокрылых гор Южной Сибири (даже для высокогорных) характерно преобладание видов, тяготеющих к степной и лесостепной зонам. Эндемиков, однако, почти среди них нет (Сергеев, 1986). Характер местных естественных сообществ и популяционных группировок прямокрылых позволяет им проникать почти во все пригодные для их существования агро- и уробиоландшафты. Вместе тем наблюдается определенная региональная специфика в формировании населения нарушенных местообитаний. Для Улуг-Хемской котловины для полей залежей характерно падение разнообразия на фоне такого же уровня численности, что и в зональных станциях. На пастбищах обычно наблюдается резкое повышение обилия прямокрылых при небольшом падении видового разнообразия. В качестве доминанта повсюду сохраняется плакорный комплекс саранчовых. Примеси видов, не свойственных зональным ландшафтам, почти нет. Еще резче эта тенденция выражена в горно-степном Забайкалье. Поля и залежи заселяются в первую очередь граминиколами. На орошаемые массивы, однако, заселяются мезофильные прямокрылые, встречающиеся в засушливых степях почти исключительно в пределах речных долин, т.е. очень локальными поселениями. В местные урбоценозы эти насекомые проникают незначительно, за исключениемGlyptobotrus. biguttulus, который встречается здесь даже в центральных частях городов и поселков (Сергеев, 1986). Для населения прямокрылых в нарушенных экосистемах Убсунурсой котловины свойственны в основном те же черты, что и для природных. В первую очередь это господство прямокрылых из трибыBryodemini. Вместе с тем орошаемые поля однолетних культур, и особенно приканальные полосы, в значительной степени заселяются мезофильными, как правило, в той или иной степени тяготеющими к поверхности почвы видами (Glyptobotrus biguttulus, Chorthippus fallax). На залежах различного возраста разнообразие и обилие этих видов постепенно возрастает (Сергеев, 1986). ЗАКЛЮЧЕНИЕ На сравнительно небольшой территории Тувы имеется большое разнообразие природных ландшафтных и климатических условий, определяющих большое разнообразие растительных сообществ. На территории Тувы проходит граница между Скифской и Сахаро-Гобийской зоогеографическими подобластями Палеарктической области. Многие виды прямокрылых, обитающие на территории Тувы, широко распространены на юге Сибири и севере Монголии. В настоящее время для этого региона в литературе указывается 60 видов прямокрылых из 8 семейств (Tettigonidae, Bradiporidae, Gryllidae, Trigonidiidae, Tridactylidae, Tetrigidae,Catantopidae, Trigonidiidae). СПИСОК ЛИТЕРАТУРЫ 1. Бей-Биенко Г. Я. Общая энтомология – М.: Высшая школа, 1980. – 416с. 2. Бей-Биенко.Г.А. Саранчовые фауны СССР и сопредельных стран – Новосибирск: Издательство Академии Наук СССР,1951. – 668с. 3. Бережков Р.П. Саранчовые Западной Сибири – Томск: Издательство ТГУ, 1956 – 175с. 4. Второв П.Г. Эталоны природы – М. Издательство Наука, 1983 – 203с 5. Лачининский А. В., Сергеев М. Г., Чильдебаев М. и др. Саранчовые Казахстана, Средней Азии и сопредельных территорий. - Ларами, 2002. – 387с. 6. Локвуд Дж. А. Экология насекомых в азиатских и североамериканских степях // Сибирский экологический журнал, 1997, 4 (3). С. 241-251. 7. Мищенко Л.Л. Отряд Orthoptera (Saltartoria) – Прямокрылые ( Прыгающие насекомые) // Насекомые и клещи – вредители сельскохозяйственных культур, 1972. – 282с. 8. Правдин Ф.Н. Экологическая география насекомых средней Азии. Ортоптероиды. – М.: Издательство Наука, 1978. – 271с. 9. Сергеев М. Г. География и природные ресурсы. - 1989. №2. – С.89-93. 10. Сергеев М. Г. Закономерности распространения прямокрылых насекомых азиатской части СССР: автореферат д-ра биол. наук/ М. Г. Сергеев, СПб – Н.,1991. 11. Сергеев М. Г. Закономерности распространения прямокрылых насекомых Северной Азии – Новосибирск. Издательство Наука, 1986 – 237с 12. Сергеев М. Г. Биологическое разнообразие и сообщества ортоптероидных насекомых в горах Средней и Центральной Азии // Сибирский экологический журнал №6, 1994 13. Сергеев М. Г., Копанева Л.М., Рубцов И.А., Антипанова Е.М., и др. Сибирская кобылка (Aeropus sibiricus L.) – Новосибирск: Издательство Наука, 1995. – 176с. 14. Соболев Н. Н., Омельченко Л. В. Вопросы экологии: – Новосибирск: Издательство Наука, 1981. – 121с. 15. Стебаев И. В. Саранчовые (Acridoidea) и чернотелки (Tenebrionidae) как стимуляторы микробиологических процессов в почвах сухих степей Тувинской автономной республики // Почвоведение,1964 - 95 16. Росс Г.,Росс Ч., Росс Д. Энтомология. – М.: Мир, 1985. – 572с. ПРИЛОЖЕНИЕ Таблица. Предварительный список прямокрылых Тувы. № | Вид | Русское название | Распространение | Подотряд Ensifera (Длинноусые) | СемействоTettigonidae(Кузнечики). | 1. | Poecilimon intermedius (Fieber, 1853) | Пилохвост восточный. | Центр и восток Европейской части, юг Сибири на восток до Забайкалья. | 2. | Tettigonia caudata(Charpentier, 1842) | Кузнечик хвостатый. | Центр и юг Европейской части, Кавказ, юг Сибири на восток до Прибайкалья. | 3. | Bienkoxenus beybienkoi (Stebaev, 1964) | Пустынный кузнечик Бей-Биенко. | Тува | 4. | Eulithoxenus mongolicus (Uvarov, 1928) | Кузнечик монгольский. | Тува. | 5. | Onconotus laxmanni (Pallas, 1771) | Севчук Лаксманна. | Степная зона Европейской части, Кавказ, юг Западной Сибири. | 6. | Onconotus servillei(Fischer von Waldheim, 1846) | Севчук Сервилля. | Степная зона Европейской части, Кавказ, юг Западной Сибири. | 7. | Decticus verrucivorus (Linnaeus, 1758) | Кузнечик серый. | Везде, кроме крайнего севера. | 8. | Platycleis intermedia (Serville, 1838) | Скачок пятнистый. | Юг Европейской части, Кавказ, южная Сибирь на восток до Забайкалья и Ю.Якутии. | 9. | Montana eversmanni (Kittary, 1849) | Скачок Эверсманна.=dubia(Uvarov, 1910).=falzfeini(Ramme, 1929) | .Юг и отчасти центр Европейской части, южная Сибирь на восток до Тувы. | 10. | Montanamontana(Kollar, 1833) | Скачок степной. | Европейская часть (кроме крайнего севера и северо-востока), южная Сибирь на восток до Забайкалья и Якутии. | 11. | Roeseliana fedschenkoi (Saussure, 1874) | Скачок Федченко | Европейская часть (кроме крайнего севера), юг Сибири на восток до Алтайского края. | СемействоBradiporidae(Шароголовые кузнечики) | 12. | Zichya baranovi(Bei-Bienko, 1933) | Зичия Баранова. | Тува. | 13. | Eracanthina deracanthoides (Bei-Bienko, 1933) | Гуловидка обыкновенная.=kaszabi(Bazyluk, 1970). | Тува. | СемействоGryllidae(Настоящие сверчки) | 14. | Modicogryllus frontalis (Fieber, 1844) | Сверчок лобастый. | Юг Европейской части, Кавказ, юг Западной Сибири, Тува. | СемействоTrigonidiidae(Сверкуны) | 15. | Dianemobius fascipesnigrofasciatu(Walker, 1869) | Сверкун чернополосый. | Алтай, Тува, Приамурье, Приморье, Кунашир. | Caelifera (Короткоусые) | СемействоTridactylidae(Триперсты) | 16. | Bruntridactylus tartarus (Saussure, 1874) | Триперст туркестанский. | Юг Европейской части, Предкавказье, юг Сибири на восток до Прибайкалья. | СемействоTetrigidae(Прыгунчики) | 17. | Tetrix simulans(Bei-Bienko, 1929)=amurense(Bei-Bienko, 1929) | | Юг Сибири от Алтая и Енисея до Забайкалья, Приамурье, Приморье. | 18. | Tetrix tenuicornis (Sahlberg, 1891) | Прыгунчик тонкоусый | Европейская часть (кроме крайнего севера и крайнего юга), Кавказ, Сибирь (кроме севера, на восток до Прибайкалья и Якутии). | СемействоCatantopidae(Шипогрудые саранчевые) | 19. | Zubovskya mongolica(Storozhenko, 1986). | | Тува, Ю-З. Забайкалье. | 20. | Podismapedestris(Linnaeus, 1758) | Кобылка пешая. | Номинативный подвид на севере и в центре Европейской части, на Кавказе и юге Сибири, на северо-восток до Вилюйского района Якутии. | 21. | Ognevia longipennis (Shiraki, 1910) | Кобылка древесная.=debilis(Ikonnikov, 1911) | Юг Сибири от Алтайского края до Забайкалья, Приамурье, Приморье, Сахалин. | 22. | Calliptamus abbreviatus (Ikonnikov, 1913) | Прус светлокрылый.=holoptera(Ramme, 1951).=sibiricus(Vnukovskii, 1926). | Юг Сибири от Алтая до Забайкалья, Приамурье, Приморье. | СемействоAcrididae(Настоящие саранчевые) | 23. | Psophus stridulus(Linnaeus, 1758) | Огневка треcкучая | Лесная и лесостепная зона Европейской части, Кавказ, южная Сибирь на восток до Забайкалья, Приамурье, Приморье. | 24. | Locusta migratoria (Linnaeus, 1758) | Саранча перелетная. | Центр и Юг Европейской части, Кавказ, Приморье. Как мигрант залетает на север Европейской части и в южную Сибирь. | 25. | Oedaleus asiaticus(Bei-Bienko, 1941) | . | | 26. | Bryodema gebleri(Fischer von Waldheim, 1836) | Трещотка Геблера. | Номинативный подвид (=baicalensis(Fischer von Waldheim, 1846)) на Южном Урале, Алтае, в Туве, на Восточном Саяне и в Забайкалье. Подвидmongolicum(Zubovski, 1900) на Ю-В. Алтае (Чуйская и Курайская степь) и на южных склонах хр. Танну-Ола. | 27. | Bryodemella holdereri (Krauss, 1901) | | Номинативный подвид в Забайкалье, подвидoccidentale(Bei-Bienko, 1930) на Ю-В. Алтае, в Минусинской котловине и Туве. | 28. | Bryodemella orientale(Bei-Bienko, 1930). | | России подвидsimulans(Stebaev, 1964). Тува (Убсунурская котловина). | 29. | Bryodemella tuberculata (Fabricius, 1775) | Трещотка ширококрылая. | Номинативный подвид на северо-западе и в центре Европейской части, на Южном Урале и Алтае. Подвидdilutum(Stoll, 1813) от Среднего Урала через среднюю и южную Сибирь до Колымы и Приморья. | 30. | Angaracris barabensis (Pallas, 1773) | Трещотка барабинская.=rhodopa(Fischer von Waldheim, 1836). | Южное Зауралье, юг Сибири на восток до Забайкалья. | 31. | Celes skalozubovi (Adelung, 1906) | Кобылка Скалозубова. | Номинативный подвид на Южном Урале, в южной Сибири на восток до Забайкалья, изолированно на Приленском плато (к югу от Якутска). Подвидakitanus(Shiraki, 1910) в Приморье. | 32. | Celes variabilis(Pallas, 1774) | Кобылка изменчивая. | Лесостепная и степная зона Европейской части, Кавказ, южная Сибирь на восток до Западного Саяна. | 33. | Sphingonotus beybienkoi (Mishchenko, 1936) | Пустынница Бей-Биенко. | Тува, южное Забайкалье. | 34. | Epacromius coerulipes (Ivanov, 1887) | Летунья синеногая. | Юг Европейской части, восточное Предкавказье, южная Сибирь. | 35. | Epacromius pulverulentus(Fischer von Waldheim, 1846). | | Юг Европейской части, юг Сибири на восток до Забайкалья, Приамурье, Приморье. | 36. | Stethophyma grossa (Linnaeus 1758) | Кобылка болотная большая. | Европейская часть, Кавказ, Сибирь (кроме севера), Приамурье. | 37. | Stethophyma magister(Rehn, 1902).=tsherskii(Ikonnikov, 1911). | | Юг Сибири от Хакасии до Забайкалья, Приамурье, Приморье, Ю. Курилы. | 38. | Pararcyptera microptera(Fischer von Waldheim, 1833) | Кобылка крестовая. | Номинативный подвид на юге Европейской части, на Северном Кавказе (кроме Дагестана) и на юге Западной Сибири | 39. | Arcyptera albogeniculata(Ikonnikov, 1911)=orientalis(Storozhenko, 1988). | | Тува, Забайкалье, Приамурье, Приморье. | 40. | Arcyptera fusca(Pallas, 1773) | Кобылка пестрая. | Юг Европейской части, Кавказ, юг Западной Сибири, Алтай, Прибайкалье, Якутия. | 41. | Eremippus mistshenkoi (Stebaev, 1965) | Пустынник Мищенко. | Тува (хр. Танну-Ола). | 42. | Euthystira brachyptera (Ocskay, 1826) | Зеленчук короткокрылый=longicrus(Fischer von Waldheim, 1846)=leucoptera(Fischer von Waldheim, 1846)=homoptera(Eversmann, 1848). | Европейская часть (кроме крайнего севера), Кавказ, Сибирь (кроме севера). | 43. | Mongolotettix japonicus (Bolivar, 1898) | Зеленчук полосатый. | Номинативный подвид в Приамурье и Приморье. Подвидvittatus(Uvarov, 1914) на юге Восточной Сибири. | 44. | Podismopsis altaica (Zubovski, 1900) | Короткокрылка алтайская. | Южная Сибирь от Алтая до Забайкалья. | 45. | Podismopsis poppiusi (Miram, 1908) | Короткокрылка лесная. | Номинативный подвид на севере Европейской части и Западной Сибири, в Восточной Сибири. Подвидfemoralis(Mishchenko, 1951) на Алтае (Онгудай), в Хакасии и на юге Красноярского края. | 46. | Omocestus haemorrhoidalis (Charpentier, 1825) | Травянка краснобрюхая. | Номинативный подвид в Европейской части (кроме крайнего севера), почти на всем Кавказе, в Сибири (кроме севера) на восток до Забайкалья. Подвидciscaucasicus(Mishchenko, 1951) в горной Чечне (Салги). | 47. | Omocestusrufipes(Zetterstedt, 1821) | Травянка красноногая=ventralis(Zetterstedt, 1821). | Европейская часть, Кавказ, южная Сибирь на восток до Забайкалья. | 48. | Omocestus viridulus (Linnaeus, 1758) | Травянка зеленая. | Европейская часть, Кавказ, Сибирь (кроме севера) на восток до Забайкалья. | 49. | Stenobothrus fischeri (Eversmann, 1848) | Травянка Фишера. | Юг Европейской части, Кавказ, юг Сибири на восток до Алтая, Тува. | 50. | Stenobothrus lineatus (Panzer, 1796) | Травянка толстоголовая. | Европейская часть (кроме севера), Кавказ, юг Сибири на восток до Забайкалья. | 51. | Myrmeleotettix palpalis (Zubovski, 1900) | Копьеуска восточная. | Юг Сибири от Алтая до Забайкалья. | 52. | Gomphocerus rufus(Linnaeus, 1758) | Копьеноска рыжая. | Европейская часть, Кавказ, Сибирь (кроме севера) на восток до Забайкалья. | 53. | Aeropus sibiricus(Linnaeus, 1767) | Кобылка сибирская. | Номинативный подвид на севере и востоке Европейской части, по всей Сибири на восток до северного Приамурья, Камчатки и Чукотки. Подвидcaucasicus(Mochulsky, 1840) на Кавказе. | 54. | Stauroderus scalaris(Fischer von Waldheim, 1846) | Конек темнокрылый. | Центр и юг Европейской части, Кавказ, почти вся Сибирь на восток до Якутии и Забайкалья. | 55. | Chorthippus brunneus (Thunberg, 1815) | Конек обыкновенный. | Европейская часть, Кавказ, Сибирь (кроме севера) на восток до Забайкалья, Приамурье. | 56. | Chorthippus dubius(Zubovski, 1898) | Конек узкокрылый. | Юго-восток Европейской части, юг Сибири на восток до Забайкалья. | 57. | Chorthippus hammarstroemi (Miram, 1908) | Конек сибирский. | Сибирь от Алтая до Якутии и Забайкалья, Приамурье, Приморье. | 58. | Chorthippus karelini (Uvarov, 1910) | Конек Карелина=Ch. albomarginatus karelini(Uvarov, 1910). | Юго-восток Европейской части, Тува, Прибайкалье, Забайкалье. | 59. | Chorthippus montanus(Charpentier, 1825) | Конек лесной=longicornispartim. (Latreille, 1804). | Север и центр Европейской части, южная Сибирь на восток до Верхоянска и Забайкалья, Камчатка. | 60. | Chorthippus porphyropterus (Voroncovskij, 1928)=Ch. mollis porphyropterus (Voroncovskij, 1928). | | Юг Европейской части, юг Сибири на восток до Забайкалья. |
| superbotanik.net
Реферат - Фауна совок (lepidoptera: noctuidae) юго-восточного средиземноморья: структура и ландшафтное распределение 03. 02. 05 энтомология
На правах рукописи
Кравченко Василий Дмитриевич
ФАУНА СОВОК (LEPIDOPTERA: NOCTUIDAE) ЮГО-ВОСТОЧНОГО
СРЕДИЗЕМНОМОРЬЯ: СТРУКТУРА И ЛАНДШАФТНОЕ
РАСПРЕДЕЛЕНИЕ
03.02.05 – энтомология
автореферат
диссертации на соискание ученой степени
доктора биологических наук
Москва – 2010
Работа выполнена на кафедре зоологии факультета Естественных наук им.
Дж. Визе Тель-Авивского Университета
Научный консультант: член корреспондент РАН, профессор
^ Стриганова Бeлла Рафаиловна
Официальные оппоненты: доктор биологических наук, профессор
Сачков Сергей Анатольевич
доктор биологических наук, профессор
^ Чернышев Владимир Борисович
доктор биологических наук
Синёв Сергей Юрьевич
Ведущее учреждение: Московский Педагогический
Государственный Университет,
^ Биолого-химический факультеГлава 6. ^ ВЛИЯНИЕ РЕЛЬЕФА НА РАЗНООБРАЗИЕ ВИДОВОГО СОСТАВА СОВОК
6.1. Экспозиция склонов и ветровой перенос влаги
Основное направление ветра в средиземноморской части ЮВС ориентировано со стороны моря (с запада), горные цепи вытянуты меридионально (вдоль моря) в два ряда, поэтому наветренные склоны хребтов, обращённые к морю увлажнены гораздо лучше, чем подветренные, обращённые в сторону континента. В связи с этим в меридиональном направлении наветренные склоны одного и того же хребта являются путём проникновения мезофильной фауны на юг, а подветренные склоны – путём проникновения аридной фауны на север. Для выявления влияния ветрового переноса влаги рассмотрены особенности изменений видового состава совок на профиле от Тель Авива до пустыни Бадия (рис. 5).
Показано, что на наветренных (более увлажнённых) склонах хребтов преобладают мультимодальные виды средиземноморского происхождения, а на подветренных – унимодальные виды ирано-туранского происхождения. В приморской части преобладают мультимодальные виды тропического, средиземноморского происхождения. На дне рифтовой долины в окрестностях Мёртвого моря доминируют унимодальные пустынные виды сахаро-синдского происхождения.
Рис. 5. Распределение видов соответственно их ареалогическим характеристикам и количеству периодов лёта на геоморфологическом профиле Тель Авив – Амман. Ш Р – широко распространенные виды. С – средиземноморские, И Т - ирано-туранские, С С – сахаро-синдские, Т – тропические, У – унимодальные, М – мультимодальные.
^ 6.2. Экспозиция склонов и освещённость
Высокая степень расчленённости ЮВС имеет место не только в меридиональном, но и в широтном направлении. Западные склоны горных хребтов прорезаны многочисленными глубокими каньонами, выработанными реками, стекающими к морю, и ориентированы в широтном направлении. Экспозиция этих каньонов весьма сходна. Соответственно наклону земной оси, склоны каньонов обращённые на юг, получают гораздо большее количество солнечной радиации, чем склоны, обращённые на север (в южном полушарии ситуация обратная). Различия в величине радиационного баланса сказываются на микроклимате склонов (Pavlíček at al., 2003), а соответственно и на составе фауны бабочек и времени их лёта (Kravchenko, 2002a; 2002b).
Сравнение фауны совок в двух каньонах, расположенных в Средиземноморской зоне на расстоянии более 100 км друг от друга (Nahal Keziv – в верхней Галилее и Nahal Oren – на Кармеле) показало, что видовой состав совок на склонах с одинаковой экспозицией более сходен между собой, чем видовой состав противоположных склонов в одном и том же каньоне (рис. 6).
Рис. 6. Влияние освещенности на склонах с разной экспозиции на видовой состав и численность совок. Е – тропические, СС – сахаро-синдские, ИТ – Ирано-туранские, ШР – широко распространенные, С – средиземноморские.
Видовой состав на склонах с южной экспозицией сходен (индекс сходства Чекановского = 0,74), а в сезонной динамике проявляются весенний и осенний пики видового разнообразия. Весной преобладают унимодальные средиземноморские скальные виды обычные в сходных биотопах в Италии, Греции и Турции. Осенняя фауна представлена унимодальными ирано-туранскими видами подсемейства Amphipyrinae, развивающимися на травянистой растительности. Фауна северных склонов обоих каньонов сходна (индекс = 0,78) и представлена средиземноморскими видами, развивающимися на деревьях, преимущественно видами рода Catocala. Сезонная динамика видового разнообразия на склонах обоих каньонов с северной экспозицией характеризуется только одним пиком видового разнообразия – в мае, июне.
Характерной геоморфологической особенностью строения склона каньонов, является терраса между наклонной равниной, образованной осыпанием отвесного склона (педимент) и самой отвесной скалой. Подстилающим горизонтом таких террас является водоупорный слой, способствующий накоплению конденсатной влаги с поверхности отвесной скалы. Поэтому такая терраса становится центром видового разнообразия растений и бабочек, а в жаркие летние месяцы естественным резерватом для мультивольтинных средиземноморских видов и видов с широкими палеарктическими ареалами.
^ 6.3. Вертикальная зональность
6.3.1. Западный (наветренный) склон Антиливанского хребта
Высотные пояса и растительные ассоциации для наветренного склона Антиливанского хребта выделены Шмида (Shmida, 1977), а так же Даниным и Питманом (Danin, Plitmann, 1987). В главе прослежены изменения видового состава совок с предгорий на высоте 200-300 м до высоты 2200м. Всего выявлен 291 вид совок, из них только 41 вид (14,1 %) встречался по всему склону, остальные 250 видов специфичны для отдельных биотопов.
^ 6.3.2. Сравнение западного и восточного склонов Антиливанского хребта
Высотные пояса и растительные ассоциации для восточного склона Антиливанского хребта выделены Коэном (Cohen at al., 1981). Этот склон поднимается от Сирийской пустыни и только на высоте около 500 м на нем появляется типичная степная растительность, выше которой расположен пояс вечнозелёных дубов, за которым начинается пояс трагакантов.
Здесь пояс горных лесов и кустарников совершенно не выражен. Восточный склон обследован менее детально, чем западный, однако и на нём поймано 196 видов совок. Из них только 24 вида (12,2%) встречались по всему склону, остальные были специфичными для отдельных биотопов.
Фауна предгорий, представленная преимущественно видами сахаро-синдского происхождения, по составу крайне схожа с фауной пустыни Бадия в восточной Иордании (индекс сходства Чекановского = 0,78). Фауна степей на высоте 500 – 900 м, схожа со степной фауной Иорданской долины (индекс сходства = 0,73). Фауна совок вечнозелёных дубов фактически сходна на обоих склонах, однако открытые участки в дубовых лесах на восточном склоне гораздо суше, чем на западном, поэтому здесь преобладают ирано-туранские виды (14 из 19). Над поясом вечнозелёных дубов отмечено 12 горно-степных видов, в отличие от западного склона, преимущественно ирано-туранского происхождения.
Выше по склону начинается пояс трагакантов, населённый такими же видами, как и на противоположном склоне. Участков с альпийской растительностью на этом склоне не обнаружено.
Ниже приводятся краткие эколого-географические характеристики комплексов совок отдельных местообитаний горных склонов (рис. 7).
1) В нижней части западного склона на заболоченных и луговых участках присутствуют мультимодальные луговые (полифаги) и болотные (монофаги) виды с широкими палеарктическими ареалами.
2) В приречных лесах - унимодальные зимние, или ранневесенние монофаги, развивающиеся на деревьях, таких же, как и в лесной зоне Евразии.
3) В поясе лесов вечнозелёных дубов обитают унимодальные летние монофаги средиземноморского происхождения.
Рис. 7. Комплексы совок в южной части Антиливанского хребта. 1) Заболоченные и луговые участки. 2) Приречные леса. 3) Леса из вечнозелёного дуба. 4) Открытые участки среди леса. 5) Горные леса. 6) Горные степи. 7) Участки с альпийской растительностью. 8) Трагаканты. 9) Открытые участки среди леса. 10) Степи. 11) Сирийская пустыня.
4) В открытых местообитаниях среди леса на западном склоне - унимодальные летние и осенние полифаги со средиземноморскими и широко-палеарктическими ареалами, связанные с травянистой растительностью, а на восточном склоне - унимодальные осенние полифаги на травянистой растительности с ирано-туранскими ареалами.
5) В горных лесах обитают унимодальные осенние монофаги на листопадных дубах и розоцветных, распространенные так же и на юге лесной зоны Европы.
6) В горно-степных местообитаниях западного склона - унимодальные раннелетние и осенние средиземноморские и ирано-туранские полифаги, связанные с травянистой растительностью.
7) В альпийских местообитаниях - унимодальные раннелетние полифаги с горно-средиземноморскими ареалами на травянистой растительности.
8) В сообществах трагакантов - унимодальные раннелетние и осенние подгрызающие совки со средиземноморскими и ирано-туранскими ареалами.
9) В открытых местообитаниях среди леса на восточном склоне - унимодальные осенние полифаги на травянистой растительности с ирано-туранскими ареалами.
10) В степях предгорий восточного склона - унимодальные ранневесенние и осенние полифаги с ирано-туранскими ареалами на травянистой растительности.
11) У подножья восточного склона в Сирийской пустыне - унимодальные ранневесенние (монофаги) и осенние (полифаги) с сахаро-синдскими ареалами на эфемерной растительности.
^ 6.3..3. Сравнение видового состава западных склонов Ливанского и Антиливанского хребтов
Реконструкция вертикального распределения совок на Ливанском хребте основана на анализе материалов, собранных Бетинским-Зальцем (Bytinski-Salz) в 1950 – 60-х годах в Ливане и публикаций Уилче (Wiltshire, 1940; 1952), Элисона и Уилче (Ellison and Wiltshire, 1939) и Тальхука (Talhouk, 1997). Растительные ассоциации и высотные пояса наветренного склона Ливанского хребта (по Ramadan-Jaradi & Ramadan-Jaradi, 1999) соответствуют таковым на наветренном склоне Антиливанского хребта.
Отличия между хребтами проявляются в фауне предгорий и высокогорий. На Ливанском хребте отмечено 43 вида совок, не найденных на Антиливанском хребте. В высокогорьях Ливанского хребта отмечено 14 видов, отсутствующих в высокогорьях Антиливана. Преимущественно это горно-средиземноморские и широко распространённые в Палеарктике виды. В высокогорьях Антиливана пойман 21 вид, не отмеченный на Ливанском хребте. Большинство этих видов ирано-туранского происхождения.
В предгорьях Ливанского хребта большая площадь влажных биотопов по сравнению с Антиливаном. Поэтому, в предгорьях Ливанского хребта луговых и болотных видов значительно больше, в предгорьях Антиливана. В фауне высокогорий Антиливанского хребта превалируют ирано-туранские виды (79 %), тогда как на Ливанском хребте их всего 31 %. Такая разница связана с высоким уровнем увлажнения и меньшей сезонной амплитудой температур на наветренном склоне Ливанского хребта, по сравнению с континентальными и засушливыми вершинами Антиливана.
^ 6.4. Преобладание подгрызающих совок - характерная особенность высокогорий
Характерной чертой обоих высокогорий является повышенное количество видов подгрызающих совок. Это происходит вследствие появления здесь видов специфических горных родов таких, например, как Chersotis и Dichagyris. Расцвет подгрызающих совок в горах, вероятно, связан с повышенной плотностью корней растений в почве. Стебли горных растений укорочены и прилегают к земле, а корневая система сильно развита и образует единую сеть для препятствования водному смыву на крутых горных склонах.
Глава 7. ^ ОСОБЕННОСТИ ЛАНДШАФТНО-БИОТОПИЧЕСКОГО РАСПРЕДЕЛЕНИЯ СОВОК
В ЮВС выделены три ландшафтных района (глава 1.2.) в пределах, которых описано 25 биотопов. За основу выявления отдельных биотопов принято геоботаническое описание (Danin, 1988). Эта схема дополнена нами выделением ряда местообитаний, характеризующихся наличием специфических видов совок. Для всех выделов указаны специфические виды совок, встречающиеся только в этом биотопе, а так же виды, находящиеся на окраине их ареалов и эндемики ЮВС. Рассматриваются трофическая специализация и время лёта специфических видов.
Глава 8. ^ ЦЕНТРЫ ВИДОВОГО РАЗНООБРАЗИЯ СОВОК В ЮВС
8.1. Распределение видов, находящихся на окраине своего ареала по биотопам
Поскольку ЮВС расположено на границе трёх подобластей, почти половина видов совок здесь (294/634 – 46,4 %) находится на окраине своего ареала. Для сравнения, на территории соседних стран доля таких видов гораздо ниже. Например, в Турции – 11,4%, в Ираке - 6,2%, в Саудовской Аравии и Египте – 7,8%. Согласно пограничному положению ЮВС, большинство маргинальных популяций представлено средиземноморскими видами (98 видов – 33,3 %), ирано-туранскими (93 – 31,6 %) и сахаро-синдскими (103 – 35 %). Для 86 видов в ЮВС отмечены популяции, далеко отстоящие от границ основного ареала (т.е. эксклавами).
Процент видов находящихся на окраине их ареала в различных биотопах составляет от 21,7% до 90,9% (рис. 8). Характерно, что большинство этих же видов являются специфическими для биотопов, которые они населяют. Это связано с тем, что выраженность экологических границ между различными биотопами на границе ландшафтных районов увеличивается (Safriel et al., 1994).
Анализ карты распределения биотопов с повышенным количеством видов на окраине их ареала и эксклавов показывает, что в пределах ЮВС они концентрируются либо в горных системах, либо в рифтовой долине (рис. 9). Все новые для науки виды обнаружены там же. Для многих видов описанных из ЮВС в XIX и XX веках, типовое местонахождение указано не определённо (например «Palestine»). Но для 56 видов оно известно с точностью до ближайшего населённого пункта.
Рис. 8. Количество специфических видов, видов на окраине ареала и их биогеографические характеристики. Биотопы отсортированы соответственно убыванию процента видов на окраине их ареала. Биотопы с более, 70% видов на окраине их ареала помечены жирным шрифтом. Ландшафтные районы: С – средиземноморский, ИТ – Ирано-Туранский, СС – Сахаро-Синдский.
Все эти виды так же были обнаружены либо в Ливанских горах – 16 видов, либо на склонах Антиливанских гор – 12 видов, либо в рифтовой долине и её окрестностях (Jerusalem, Jericho) – 28 видов. Несмотря на равномерное обследование территории Израиля, и Иордании в последние 18 лет, все 22 вида, описанных нами, так же были обнаружены в пределах рифтовой долины, или на хребте Антиливан.
^ 8.2.Рефугиальные комплексы
8.2.1. Горные системы
Анализ вертикального распределения бабочек в различных горных системах Центральной Европы и Средиземноморья показал, что наибольшее видовое богатство приходится на нижние или средние высоты (Claridge, Singharao, 1978; Sanchez-Rodriguez, Baz, 1995). По мере подъёма уменьшается число неспециализированных горных видов, соответственно, увеличивается количество специализированных. При этом общее количество видов в высокогорьях снижено в связи со специфичностью условий. В отличие от таких горных систем, в горах ЮВС общее количество видов бабочек в высокогорьях не только не снижается, но даже увеличивается.
Такая особенность рассматривается как следствие смещения видов из низких высотных поясов на вершины из-за климатических флуктуаций. В ЮВС этот процесс усложняется различиями между западными и восточными склонами одних и тех же горных хребтов. На западных (наветренных) склонах зональность ранжируется от предгорных лугов до пояса трагакантов и альпийской растительности, а средние высоты служат убежищем для видов со средиземноморскими или широкими палеарктическими ареалами. На восточных (подветренных) склонах зональность ранжируется от Сирийской пустыни до горных степей и фауна меняется от сахаро-синдской до ирано-туранской (рис. 7). Поэтому, на вершинах гор в полной мере представлены специализированные виды высокогорий, как средиземноморской, так и ирано-туранской фауны.
Для 73.1 % видов специализированной горной фауны вершины Ливанского и Антиливанского хребтов являются окраиной их ареала. Этот феномен проявляется и на родовом уровне. Например, все 7 отмеченных в ЮВС видов горно-средиземноморского рода Chersotis не встречаются южнее этих хребтов. Для 17 из 25 видов горного, преимущественно ирано-туранского рода Dichagyris, это также южная граница ареала. Остальные виды этого рода продвигаются на юг по вершинам сухих гор. Они отмечены на вершинах Иудейских гор, высоком Негеве (Мицпе Рамон) и 4 вида доходят вплоть до гор южного Синая (Санта Катарина – 2800 м). В зоне трагакантов в замкнутых долинах концентрируется альпийская растительность и здесь же встречается большинство специализированных горных видов рода Hadena, для которых эти долины также становятся южной окраиной ареала.
В горных степях ЮВС большая часть специфических видов так же находятся на южной окраине их ареала. Севернее, в центрах их ареалов они широко распространены в низинных травянистых биотопах, но по мере продвижения на юг поднимаются в горы. Например, это ряд обычных лесостепных европейских видов: Hadena compta, Lygephila lusoria, и даже несколько известных вредителей овощных и зерновых культур в Палеарктике: серая зерновая совка - Apamea anceps, совка восклицательная Agrotis exclamationis.
Западные склоны приморских хребтов являются южным форпостом для большинства средиземноморских моно- и олигофагов, развивающихся на деревьях. Среди них, виды связанные с розоцветными деревьями и кустарниками, как например все виды подсемейства Psaphidinae, Catocala hymenaea, Tiliacea cypreago и другие. Для видов развивающихся на листопадных дубах это также южная окраина ареала. Это виды рода Dichonia, Maraschia grisescens, Rileyiana fovea, Scotochrosta pulla и многие другие.
В приречных лесах у подножия западных склонов большинство видов так же находится на южной окраине их ареала. По большей части, это виды обычные в умеренных областях Европы и Азии, развивающиеся на тополях и ивах. Как, например виды родов Orthosia, Acronicta, Craniophora, Lithophane, а также виды рода Egira, связанные преимущественно с платанами.
По внутриконтинентальной цепи гор (от Антиливана в Сирии до гор Эдом в южной Иордании) прослеживается ещё одна граница распространения видов. Здесь исчезает ряд ирано-туранских видов, которые не переходят через рифтовую долину в Средиземноморскую (прибрежную) часть ЮВС. Например, это Eicomorpha antiqua, Cucullia hemidiaphana, Caradrina belucha и Tarachephia hueberi.
^ 8.2.2. Рифтовая долина
Рифтовая долина является северной частью Большого Африканского разлома, и опускается в районе Мёртвого моря на 420 м ниже уровня мирового океана. Множество пустынных видов в ЮВС встречаются только узкой полосой по дну этой долины. Из 55 специфических видов, населяющих эту долину, большинство так же находятся на окраине их ареала. Часть из них - псаммофилы из внутренних районов Сахары, как например виды родов Anumeta, Anydrophila, Eremodrina, или виды обычные для лёссовых почв Аравийского полуострова, как например пустынные виды рода Agrotis, Polymixis juditha, P. lea, P. aegyptiaca, Diadochia stigmatica и другие.
Своеобразной смесью различных фаун являются пресноводные оазисы в рифтовой долине. Здесь большинство специфических видов также находятся на окраине своего ареала. Часть из них – типичные обитатели оазисов в пустынях Африки и Аравийского полуострова, например: Condica palaestinensis, Arenostola deserticola, Rhabdophera arefacta, другие типичны для эфиопской фауны, как например виды рода Ulotrichopus.
^ 8.2.3. Побережье Средиземного моря
На побережье маргинальные популяции видов обнаружены в двух типах биотопов. Один из них, орошаемые парки скверы и газоны в городах и населённых пунктах, где обычны виды, интродуцированные из тропической Африки и Индии. Другой - песчаные дюны со своеобразной псаммофильной фауной распространённой от песков Андалузии вдоль южного побережья Средиземного моря.
^ 8.3. Климатические ритмы и видовое разнообразие совок
Дизъюнкция ареалов совок ЮВС возможно происходила при чередовании периодов потепления и похолодания климата в течение последних 3-5 миллионов лет (четвертичный период). Периоды наступления ледников связаны в ЮВС и Сахаре с периодами
Рис. 9. Места находок новых видов, эксклавы и вероятные пути исторических миграций различных ареалогических групп.
увеличения осадков (плювиальные периоды), а отступления ледников - с иссушением (интерплювиальные периоды) (Смит, 1990).
Чередование ряда плювиальных и интерплювиальных периодов приводило к тому, что на территорию ЮВС попадали попеременно как европейские виды (в плювиальные периоды), так и виды сахарского и эфиопского происхождения (в интерплювиальные периоды). Беспрепятственному перемещению фауны способствовало меридиональное направление горных хребтов и рифтовой долины соответствующее направлению изменения климатического градиента. При этом горы и рифтовая долина становились своеобразными ловушками для видов, у которых по ЮВС проходила граница ареала. В случае исчезновения вида в пределах его основного ареала, эти структуры становились его единственным местообитанием.
Для северных видов возможность сохранения в период иссушения предполагала смещение в более влажные и прохладные высокогорья. Горные леса, вероятно, оказались рефугиумами древнего пояса западно-палеарктических широколиственных лесов, поскольку именно ЮВС было в наименьшей степени затронуто оледенением (Абатуров, 2007).В пустынной части рифтовой долины периоды увлажнения способствовали проникновению с юга эфиопской фауны, а периоды иссушения – проникновению аравийской и сахарской фаун. Эта часть долины в продольном профиле представляет собой свод (рис. 9 «Пути перемещения различных фаун»). Южное крыло свода начинается от уровня мирового океана (северная оконечность Красного моря), северное – на 420 м ниже уровня океана (Мёртвое море), а центральная часть приподнята на 350 м над уровнем моря (в районе мошава Фаран). Поэтому, «отступление» пустынной фауны в периоды увлажнения и похолодания в рифтовой долине происходило не на юг, а на север – в более низкий и жаркий район Мёртвого моря.
Анализ фауны совок на разных крыльях свода выявил отличия по ряду псаммофильных видов совок. Только на южном крыле свода встречаются: Anumeta hilgerti, 1909, Anumeta cestis, Odontelia daphnadeparisae, Boursinia lithoxylea . Только на северном крыле свода встречаются: Scythocentropus eberti, Anydrophila stuebeli, Anumeta spilota, Anumeta henkei и ряд других.
Глава 9. ^ ИЗМЕНЕНИЯ ФАУНЫ СОВОК ЮГО-ВОСТОЧНОГО СРЕДИЗЕМНОМОРЬЯ В ТЕЧЕНИЕ ПОСЛЕДНИХ 150 ЛЕТ. РЕДКИЕ И ЛОКАЛЬНЫЕ ВИДЫ
9.1. Изменение видового состава совок в течение последних 150 лет
Сборы совок в ЮВС начались ещё в середине XIX столетия, и большая часть старинного материала хранится в музее естественной истории при Тель-авивском университете. На основании анализа коллекционного материала прослежены изменения в фауне совок, произошедшие в течение последних 150 лет. Для некоторых видов отмечено увеличение встречаемости, для других – снижение. Риск исчезновения многих популяций совок в ЮВС крайне велик в связи с возрастающей хозяйственной деятельностью человека, в особенности это относится к видам на окраине их ареала.
Соответственно типу ареала и особенностям биологии выделены 3 группы исчезнувших видов, а так же видов ставших крайне редкими.
1. Виды, встречающиеся на вершинах гор. Ряд видов, обычных в горах, в действительности не являются специализированными горными. Это средиземноморские и широко распространенные палеарктические виды, поднимающиеся в горы только на южной окраине их ареала. Такие виды встречаются в поясе горных лесов, обычно на дне каньонов. Некоторые из них не регистрировали уже много лет, как например Euxoa nigrofusca и Ochropleura leucogaster, пойманные в последний раз в 1960. Некоторые стали крайне редкими, как например - Dichagyris flammatra, Euxoa conspicua, Noctua interjecta. Серая зерновая совка была поймана в ЮВС последний раз в 1998 г. на вершинах хребта Антиливан. Исчезновение этих видов может быть связано с общим потеплением климата.
2. Луговые виды. Некоторые из них, обычные в сборах середины прошлого века, также исчезли. Так, например ^ Acontia trabealis, Abrostola clarissa уже многие годы не попадают в ловушки. Крайне редкой и локальной стала Acontia titania, что связано, вероятно, с исчезновением лугов.
3. Степные виды. Некоторые из этих видов исчезли, или стали крайне редкими. Большая их часть на склонах рифтовой долины (в верхней части Иудейской пустыни, в Иорданской долине). Это 2 вида рода Euchalcia – Е. emichi и E. аugusta, а так же Agrotis margelanoides, Metopoceras felicina, Cucullia santonici, Omphalophana anatolica и Rhyacia arenacea. По всей вероятности, исчезновение этих видов связано с сельскохозяйственным освоением этих территорий. Три степных вида были пойманы по одному экземпляру ещё в XIX веке и уже тогда были, вероятно, крайне редкими. В настоящее время судьбу их трудно оценить. Это: Pamparama acuta, Recophora beata и Cleonymia opposita. Ещё один вид - Cryphia amseli известен только в одном экземпляре из окрестностей Мёртвого моря (Иерихон).
Большинство новых видов, описанных в последние годы, по всей вероятности являются эндемиками ЮВС. Ни один из них пока не найден в соседних странах. По большей части - это редкие виды, однако есть и весьма обычные, но локальные. Это, например: Odontelia daphnadeparisae - псаммофил из пустыни Арава, 2 степных вида обычных в октябре в окрестностях Тибериатского озера: Polymixis steinhardti и Episema tamardayanae. В разделе рассматриваются особенности биологии, распространение и уровни численности этих видов.
^ 9.2. Влияние выпаса скота на видовой состав и численность совок
В последние десятилетия природа ЮВС существенно изменилась под воздействием хозяйственной деятельности человека. Исчезли, или сохранились только в заповедниках и национальных парках некоторые биотопы присущие ЮВС. Это прибрежные болота, луга, приречные леса. В связи со строительством гостиничных комплексов катастрофически сократилась площадь прибрежных песков. Из-за сельскохозяйственного освоения пустыни Арава сократилась площадь песчаных участков на дне рифтовой долины. Наиболее обычным типом воздействия на природу в ЮВС является выпас скота (прежде всего коз и овец).
Влияние выпаса скота на различные сезонно-фаунистические комплексы совок было прослежено в Иудейской пустыне. Выделено 3 комплекса. а) не специализированные мультимодальные виды; специализированные степные представлены 2-мя группами б) унимодальными весенними видами и в) унимодальными осенними. Количество мультимодальных видов на участках с выпасом скота в целом за год снизилось на 18,2% (табл. 5), а унимодальных – осенних – на 22,7%.
Таблица 5
Влияние выпаса скота на количество видов (процент снижения числа видов).
Участки
Мультимодальные
Унимодальные
Всего
Весенние
Осенние
^ Без выпаса
44
72
97
213
С выпасом
36 (18,2%)
18 (75%)
75 (22,7%)
129 (39%)
Наибольшее влияние выпас скота оказал на весенние унимодальные виды, снизив их численность на 75%. В первую очередь, пострадали виды, развивающиеся на злаках (Poaceae), это виды родов: Autophila, Leucania, Mythimna, Apamea и на гвоздичных (Caryophyllaceae) – родов Hadena, Enterpia и Sideridis.
Для территории ЮВС характерно большое количество поселений с регулярно-орошаемой садово-приусадебной растительностью. Сравнение фауны совок в поселениях и их окрестностях (табл. 6) показало, что все 40 выявленных мультимодальных вида встречаются в обоих местообитаниях, по крайней мере, весной и осенью. С июля по сентябрь – когда травянистая растительность в природе высыхает, мультивольтинные виды практически исчезают в окрестностях поселений, а в самих поселениях их численность возрастает. Количество унивольтинных осенних видов несколько выше в окрестностях, чем в самом поселении. Это происходит за счёт подгрызающих совок видов родов Xestia и Euxoa. Наибольшие отличия между местообитаниями отмечены для унивольтинных весенних видов. В поселениях не зарегистрированы виды подсемейства Oncocnemidinae - обитатели каменистых склонов и виды приречных лесов (рода Acronicta, Egira, Orthosia).
Таблица 6
Сравнение видового состава совок в поселениях и их окрестностях (процент снижения числа видов).
Участки
Мультимодальные
Унимодальные
Всего
Весенние
Осенние
Окрестности
40
39
76
155
Посёлок
40 (0%)
6 (84,6%)
58 (23,7%)
105 (32,3%)
Глава 10.^ РАСПРОСТРАНЕНИЕ И ВРЕМЯ ЛЁТА ОТДЕЛЬНЫХ ВИДОВ
СОВОК В ЮВС
В главе приводятся ареалогические и экологические характеристики для всех 634 видов ЮВС. Распространение, характерные биотопы, сроки лёта в различных биотопах, а также кормовые растения для каждого вида приводятся на основании собственных данных для Израиля, Иордании и Синая (Kravchenko, Hausmann & Müller, 2006). Аналогичные данные по Ливану и Сирии приведены на основании анализа коллекционного материала из Музея Естественной Истории в Тель Авиве, различных Европейских музеев и литературных данных.
ЗАКЛЮЧЕНИЕ
У совок имеются специальные адаптации биологии развития. Они позволяют им освоить широкий спектр экологических ниш, различающихся по гидротермике и ресурсной базе, и поддерживать в них стабильное присутствие. В ЮВС отмечено высокое фаунистическое разнообразие и плотность населения совок. Многие виды – актуальные вредители сельскохозяйственных культур. Изучение закономерностей пространственного распределения совок и особенностей циклов их развития в разных типах экосистем представляют интерес для оценки адаптационного потенциала, а также для разработки методов контроля вредителей культурных растений.
ВЫВОДЫ
1. В результате фаунистических исследований в ЮВС было выявлено 634 вида совок, относящихся к 27 подсемействам, 34 трибам и 213 родам.
2. Высокое обилие видов объясняется тем, что юго-восточное Средиземноморье расположено на стыке трёх фаунистических подобластей: Средиземноморской, Сахаро-Синдской и Ирано-Туранской. Благодаря сильной расчленённости рельефа, большой амплитуде высот, различной экспозиции склонов по отношению к солнечному свету и ветровому переносу влаги, виды с различными экологическими требованиями находят здесь условия, подходящие для их существования.
3. Выявлены четкие границы фаунистических подобластей хорошо прослеживаются в пределах юго-восточного Средиземноморья и соответствуют трём различным типам климата, трём фитогеографическим зонам с различным биогеографическим составом растений, а так же трём биогеографически различным фаунам совок. Показано, что на границах между подобластями преобладают виды совок с крайне широкими ареалами, в большинстве своём полифаги, приуроченные к нарушенным местообитаниям, что соответствует неустойчивому видовому составу растений на этих границах.
4. Соответственно пограничному положению между подобластями, в фауне ЮВС повышенное количество видов находится на окраине их ареалов (42,1%), и часто представлены популяциями, далеко отстоящими от границ их основного ареала (эксклавами). Эти же виды являются специфическими для отдельных биотопов, в связи с увеличением гетерогенности среды на границах между крупными биогеографическими выделами.
5. Повышенное количество эксклавов связано с особенностями генезиса фауны ЮВС, связанными с структурой ландшафтов: меридиональное направление гор и рифтовой долины разрешает беспрепятственное перемещение фауны в север-южном направлении и миграции между горными склонами и рифтовой долиной при вековых колебаниях климата.
6. Адаптация сезонных циклов развития совок состоит в синхронизации периода развития гусеницы с периодом массовой вегетации кормового растения. Выявлены две стратегии, позволяющие осуществить эту цель. Ранневесенние виды откладывают яйца избирательно - на свои кормовые растения в начале периода их вегетации. Это виды-специалисты, олиго- или монофаги, их пространственное распределение агрегированное, поскольку соответствует пространственному распределению их кормовых растений. Осенние виды откладывают яйца задолго до периода активной вегетации растений. Это виды-генералисты откладывающие яйца не избирательно на сухие растения. Такая стратегия предполагает полифагию, а пространственное распределение таких бабочек весьма однородно.
7. Анализ динамики разнообразия совок за последние 100-150 лет показал, что снижение числа видов происходит за счёт Средиземноморских и широко распространенных палеарктических видов, поднимающихся в горы на окраинах их ареалов, а так же за счёт уменьшения числа луговых видов. Вероятно, в обоих случаях причиной их исчезновения является глобальное потепление.
^ СПИСОК ПУБЛИКАЦИЙ ПО ТЕМЕ ДИССЕРИАЦИИ:
Публикации в периодических изданиях (выделены издания из списка ВАК РФ)
1. Кравченко В.Д., 1981. Взаимосвязь между лётом на свет и мобильностью у Helicoverpa armigera Hb. (Lepidoptera: Noctuidae). Зоологический журнал, 9 (8): 1267-1269.
2. Кравченко В.Д., 1981. Ночная активность Helicoverpa armigera Hb. Зоологический журнал, 60 (11): 1640-1644.
3. Булеза В.В., Мытус Э.Р., Ковалёва А.С., Каган М.З., Кравченко В.Д. и Апасов С.Г., 1983. Хлопковая совка (Helicoverpa armigera Hb.). Лёт самцов на компоненты полового феромона. Доклады Академии Наук СССР, 272 (1): 244-247.
4. Кравченко В.Д., 1984. Поведение хлопковой совки (Helicoverpa armigera Hb.) в естественных условиях. Зоологический журнал, 63 (5): 682-686.
5. Кравченко В.Д., 1984. Влияние освещения на ритм призывной активности Heliothis viriplaca L.: Noctuidae. Зоологический журнал, 63 (6): 937-940.
6. Кравченко В.Д., 1985. Влияние возраста на ритм репродуктивной активности у (Heliothis armigera Hb. Noctuidae). Известия Азербайджанской Академии Наук. 3, 54-59
7. Кравченко В.Д., 1986. О связи между мобильностью и лётом на свет у ночных бабочек. Зоологический журнал, 65 (6): 859-866.
8. Кравченко В.Д., 1987. Суточный ритм лёта трёх подгрызающих видов совок (Lepidoptera: Noctuidae) к синтетическим половым аттрактантам в Азербайджане. Зоологический журнал, 66 (9): 1421-1423.
9. Pavlíček, T., Chikatunov V., Kravchenko V.D., Zahradnik P. & Nevo A., 1999. New records of deathwatch beetles (Anobiidae) from Israel. Zoology in the Middle East. 17: 77-78.
10. Kravchenko V.D., Hacker H., & Nevo A., 2001. List of Noctuoidea (Lepidoptera) collected in Israel. Esperiana. 9: 459-474.
11. Hacker H., Kravchenko V.D. & Yarom I., 2001. List of Noctuoidea (Lepidoptera) collected in Arava (Israel) with faunistical and ecological comments. Esperiana. 9: 515-534.
12. Gershenson Z., Kravchenko V.D., Pavlíček T. & Nevo A. 2001: New records of Yponomeutoid moths in Israel (Lepidoptera: Yponomeutidae, Plutellidae, Argyresthiidae). Mitteilungen des Internationalen Entomologischen Vereins. 26, 3/4: 147-153.
13. Kravchenko V.D., Pavlíček T., Chikatunov V. & Nevo A., 2002, a. Seasonal and spatial distribution of butterflies (Lepidoptera: Rhopalocera) in”Evolution Canyon”, Lower Nahal Oren, Mt. Carmel, Israel. Ecologia Mediterranea. 28/1: 98-112.
14. Kravchenko V.D., Hacker H. & Nevo A., 2002, b. “Evolution Canyon” model: Interslope differences in Noctuidae (Lepidoptera) of two canyons in Carmel and Galilee mountains, Israel. Esperiana. 9: 451-458.
15. Kravchenko V.D., Hacker H. & Nevo A., 2002. “Evolution Canyon” model: Interslope differences in occurrence of Noctuidae (Lepidoptera) of two canyons in Carmel and Galilee mountains, Israel. Israel Journal of Zoology (в настоящее время - Israel journal of ecology & evolution). 48(2):178 – 183.
16. Mandelik, Y., Dayan T., Chikatunov V., & Kravchenko V.D., 2002. Measuring species diversity: sampling effort for beetles, moths and vegetation of the San-San Nature Reserve. Israel Journal of Zoology (в настоящее время - Israel journal of ecology & evolution). 48(2):174 – 186
17. Pavlíček T., Sharon D., Kravchenko V.D., Saaroni H. & Nevo A., 2003. Microclimatic inter slope differences underlying biodiversity contrasts in “Evolution Canyon”, Mt. Carmel, Israel Journal of Earth Sciences. 52: 1-9.
18. Kravchenko V.D., Fibiger M., Mooser J. & Müller G.C., 2005. The Polymixis Hübner, [1820] genus-group of Israel, with description of one new species and one new subspecies (Lepidoptera, Noctuidae). Sociedad Hispano-Luso-Americana de Lepidopterologia (SHILAP). 33 (132): 487-502.
19. Kravchenko V.D., Fibiger M., Müller G.C. & Ronkay L., 2005. The Cuculliinae (Lepidoptera: Noctuidae) of Israel. Sociedad Hispano-Luso-Americana de Lepidopterologia (SHILAP). 33 (129): 83-95.
20. Kravchenko V.D., Fibiger M., Ronkay L., Orlova O., Mooser J., Chuang Li, Müller G.C., 2005. Plusiinae (Lepidoptera: Noctuidae) of Israel. Sociedad Hispano-Luso-Americana de Lepidopterologia (SHILAP). 33 (132): 449-459.
21. Kravchenko V.D., Orlova O., Fibiger M., Ronkay L., Mooser J., Chuang Li, Müller G.C., 2005. The Heliothinae (Lepidoptera: Noctuidae) of Israel. Sociedad Hispano-Luso-Americana de Lepidopterologia (SHILAP). 33 (131): 365-374.
22. Kravchenko V.D., Müller G.C., Orlova O. & Seplyarsky V., 2005. The Catocalinae (Lepidoptera: Noctuidae) of Israel. Russian Entomological Journal. 13 (3): 175-186.
23. Witt T., Günter C., Kravchenko V.D., Miller M., Hausmann A. & Speidel W., 2005. A new Olepa species from Israel (Lepidoptea: Arctiidae). Nachrichtenblatt der Bayerischen Entomologen. 54 (3/4): 101-115.
24. Müller G.C., Kravchenko V.D., Speidel W., Hausmann A., Ortal R., Miller M., Orlova, O. & Witt T., 2005. The distribution, phenology, ecology, behaviour and conservation issues of the new Israeli tiger moth Olepa schleini (Lepidoptera: Arctiidae). Mitteilungen der Münchener Entomologischen Gesellschaft. 95: 19-30.
25. Müller G.C., Kravchenko V.D., Speid
www.ronl.ru
