Наука биология включает в себя массу различных разделов, более мелких, но очень важных, специализирующихся на каких-то конкретных проблемах дисциплин. Это делает ее столь обширной и глобально значимой для человечества, что переоценить ее влияние просто невозможно.
Одной из таких важных наук стала эмбриология. Это достаточно старая дисциплина, понятие о которой и историю формирования мы и рассмотрим в данной статье.
Эмбриология - это не просто биологическая дисциплина. Это целая наука, которая занимается изучением образования, развития и формирования эмбрионов живых существ с момента появления половых клеток и их слияния до появления на свет нового организма.
Все эти процессы очень важны, жизненно необходимо их правильное и нормальное протекание. Поэтому цель, которую ставит перед собой данная наука - изучить все вопросы и механизмы, связанные с зародышами, их жизнью, образованием и развитием.
Исходя из поставленной цели, задачами эмбриологии являются следующие пункты.
Таким образом, становится понятно, что же такое эмбриология. Это узко специализированная наука о внутриутробном развитии эмбрионов от момента их образования и до выхода на свет. А также изучение вопросов, связанных с процессами гаметогенеза, то есть формирования половых клеток.
Значение слова "эмбриология" достаточно простое. Ведь на латыни слово "зародыш" произносится как embryon, а вторая часть слова logos - учение. Вот и получается, что в названии науки отражен весь ее глубокий смысл, кратко выражен предмет изучения.
Во всех современных толковых словарях значение слова "эмбриология" схоже. Оно практически такое же, как и в переводе с латинского. Добавить что-то новое сложное. Что значит эмбриология? Во всех источниках ответ один - наука о предзародышевом и эмбриональном развитии животных, человека и растений.
Свое начало история эмбриологии берет еще с древности. Одним из первых об исследованиях в этой области заговорил Аристотель. Его наблюдения заключались в исследовании формирования зародыша куриного яйца. Так было положено начало развития рассматриваемой науки.
Позже, уже к XVI-XVII столетиям ученые, которые были представителями данной дисциплины, разделились на два лагеря по теоретическим взглядам на вопросы формирования зародышей, и вообще происхождения новых организмов.
Так, существовали:
Суть первой заключается в следующем: все структуры будущего организма не развиваются со временем, а уже существуют в очень уменьшенном виде либо в яйцеклетке (овисты), либо в сперматозоиде (анималькулисты). А с течением жизни и развитием зародыша они просто увеличиваются в размерах за счет получаемых питательных веществ.
Такие взгляды были, конечно, ошибочными. Однако именно они просуществовали практически до середины XIX века. Приверженцами данных взглядов среди ученых разных временных периодов были:
Вторая теория в истории развития эмбриологии, которой придерживалось также значительное количество светлых голов разного времени, называется эпигенезом. Сторонники ее считали, что организм начинает свое развитие только после попадания половых клеток друг в друга. При этом в образующемся зародыше нет ничего готового. Структуры, будущие органы формируются постепенно, из внутренних тканей.
Представителями, которые придерживались данных взглядов, были:
В противостоянии этих двух лагерей копились многочисленные данные эмбриологии, ведь учеными постоянно проводились исследования, эксперименты, собирался теоретический материал.
Начиная с середины XIX века, по взглядам преформистов были нанесены сокрушительные удары благодаря следующим открытиям.
Результатом стало постепенное формирование науки такой, какой мы видим ее сегодня. Большой вклад в развитие дисциплины внесли следующие ученые XIX-XX века:
Основные разделы рассматриваемой науки можно обозначить следующими пунктами.
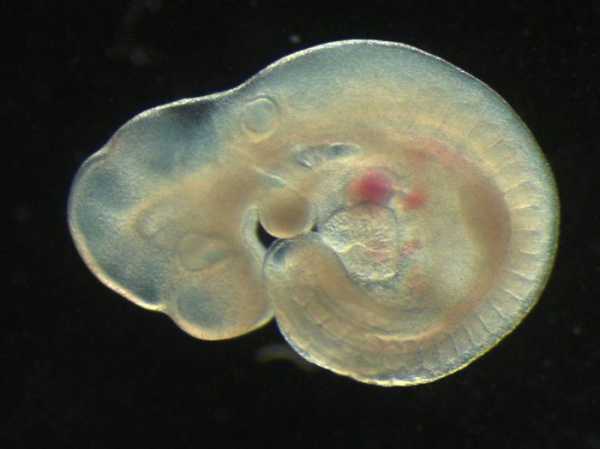
По типу исследуемых организмов эмбриология также подразделяется на:
У каждого раздела есть свои цели, задачи и объекты исследования, которые имеют большое теоретическое и практическое значение в понимании механизмов жизнедеятельности. Эмбриология животных - очень значимый раздел науки в сельском хозяйстве, животноводстве.
Общая эмбриология занимается исследованием и сравнительным анализом эмбрионов всех организмов на разных эволюционных этапах развития планеты. В результате получается множество фактического материала, доказывающего единство происхождения всего живого на нашей планете.
В область исследования данной дисциплины входит изучение процессов гаметогенеза. Данные эмбриологии имеют важное значение в вопросах здоровья будущего поколения, поэтому этой науке уделяется особенное внимание.
Основной метод сопоставления данных в этой дисциплине - анализ. Сравнительная эмбриология занимается изучением эмбрионов животных, растений или человека с целью выяснения схожести или истоков развития.
Основоположником ее стал Карл Бэр, открывший яйцеклетку человека и сформулировавший первый закон о зародышах. Большой вклад в развитие знаний дисциплины был внесен Геккелем. Его биогенетический закон был универсальным долгое время. Сравнительная эмбриология копила доказательства, подтверждающие эту особенность.

Если говорить проще, то суть сводилась к следующему: каждый эмбрион в процессе своего развития проходит множество стадий. Все они в совокупности являются повторением общего течения эволюции, которую проходили все организмы во время формирования живых существ на планете.
Отсюда такая схожесть в строении зародышей у всех классов животных: рыб, амфибий, пресмыкающихся, птиц и млекопитающих. Однако, по современным данным, закон Геккеля универсальным не является. Ведь он не объясняет, почему так сильно различаются между собой личинки насекомых и их взрослые особи, особенно когда речь идет о неполном превращении.

Еще одним пунктом, который тщательно изучается эмбриологами, являются мутации. Так, было доказано, что чем раньше возникают хромосомные неполадки, тем больший эффект их будет во внешнем проявлении после формирования организма. То есть чем более поздняя стадия подвергнется мутации, тем меньше это будет заметно фенотипически у взрослой особи.
Данный раздел имеет важное значение в развитии сельского хозяйства. Предметом изучения являются стадии формирования животных эмбрионов. Они следующие:
То есть животная эмбриология - это то же самое, что и все остальные ее разделы, только более узкоспециализированная на объекте изучения область. Она также рассматривает мутации в процессе эмбриогенеза, законы и механизмы их формирования, ищет пути предотвращения и решения различных проблем. Например, заболеваний животных организмов.
Это имеет большое значение для птицеводства, скотоводства, разведения рыбы, решения ветеринарных вопросов и проблем осеменения животных.
Самым глобальным достижением современности, которое смогла дать человеку эмбриология, является прогнозирование бесплодия и детальное наблюдение за всеми этапами формирования человеческих эмбрионов. Ведь это позволяет либо избежать рождения обреченных на генетические заболевания детей, либо исправить медицинским вмешательством грядущие мутационные перемены.

Сегодня каждая беременная женщина находится под тщательным наблюдением врачей, которые при помощи специального оборудования могут контролировать и прогнозировать любые ситуации в развитии зародыша.
Главные заслуги данной науки еще, конечно, впереди. Ведь развитие технических средств не стоит на месте, и современные технологии позволяют вмешиваться в течение практически всех известных жизненных процессов.
В будущем возможно открытие таких процессов на стадии эмбрионального развития, которые помогут избежать заболеваний плода, устранят явление бесплодия и избавят людей от множества насущных проблем.
fb.ru
ЭМБРИОЛОГИЯ
ОСНОВЫ ЭМБРИОЛОГИИ
СВЯЗЬ ИНДИВИДУАЛЬНОГО И ИСТОРИЧЕСКОГО РАЗВИТИЯ ОРГАНИЗМА
Эмбриология (от греч. embryon — зародыш, logos— учение) — наука о зародыше, о закономерностях его развития.
Медицинская эмбриология изучает закономерности развития зародыша человека, структурные, метаболические и функциональные особенности плацентарного барьера (система мать — плацента — плод), причины возникновения уродств и других отклонений от нормы, а также механизмы регуляции эмбриогенеза. Пути и методы влияния на эмбриогенез исследуются главным образом в условиях эксперимента на животных, а также в клинических условиях при патологии беременности. Одним из актуальных аспектов современной эмбриологии является изучение источников и механизмов развития тканей (гистогенез).
В понятие эмбриогенеза включают период от момента оплодотворения до рождения (для живородящих животных), вылупле-ния из яиц (для яйцекладущих), окончания метаморфоза (для животных с личиночной стадией развития).
Эмбриогенез— часть индивидуального развития, т. е. онтогенеза. Он тесно связан с прогенезом (развитие и созревание половых клеток) и ранним постэмбриональным периодом.
Актуальными задачами эмбриологии являются изучение влияния различных эндогенных и экзогенных факторов, роли микроокружения на развитие и строение половых клеток, развитие и взаимоотношение тканей, органов и систем, исследование механизмов, контролирующих репродуктивную функцию и обеспечивающих гомеостаз зародышей человека и млекопитающих, и других факторов, изучение критических периодов развития. Частным, но очень важным вопросом современной эмбриологии является культивирование яйцеклеток, зародышей и имплантация их в матку. Знание условий и факторов оплодотворения и эмбрионального развития позволяет врачам решать такие практически важные проблемы, как искусственное оплодотворение женщин при бесплодных браках, цитодиагностика патологии беременности и др.
Изучению эмбриогенеза человека предшествует краткое изложение основ сравнительной эмбриологии, так как в ходе исторического развития млекопитающих сложились основные этапы, последовательность и закономерности эмбриогенеза.
Процесс эмбрионального развития человека является результатом длительной эволюции и в определенной степени отражает черты развития других форм животного мира. Некоторые ранние стадии развития человека очень сходны с аналогичными стадиями эмбриогенеза более низко организованных хордовых животных.
Идея связи индивидуального и исторического развития была обоснована в начале XIX столетия. В частности, К. Бэр, изучая в сравнительном аспекте развитие некоторых позвоночных, пришел к заключению, что у большой группы животных на ранних стадиях развития проявляется больше сходства, чем частных, индивидуальных различий. По мере увеличения сроков эмбрионального развития это частное, индивидуальное вырисовывается все более отчетливо (закон зародышевого сходства). Ф. Мюллер, изучая развитие личиночных стадий ракообразных, также обнаружил сходство некоторых личиночных форм с вымершими ракообразными. Ч. Дарвин, придавая большое значение явлениям зародышевого сходства, считал это одним из доказательств общности происхождения животного мира.
В конце 60-х годов XIX столетия Э. Геккель сформулировал биогенетический закон, согласно которому индивидуальное развитие зародыша есть сжатое, сокращенное повторение исторического развития, иначе онтогенез повторяет в краткой форме филогенез. Идея биогенетического закона сыграла большую роль в развитии не только эмбриологии, но и эволюционного учения. Вместе с тем формулировка биогенетического закона не отражает влияние факторов окружающей среды, экологических условий, которые имеются в действительности и оказывают влияние на эмбриогенез. А. Н. Северцов, продолживший в 20—30-е годы XX столетия разработку биогенетического закона, пришел к заключению, что эволюционный процесс совершается не путем накопления изменений признаков взрослых животных, как считали Ч. Дарвин и Э. Геккель, а путем суммирования изменений, появляющихся у зародышей (теория филэмбриогенеза). Биологически важная перемена в условиях существования (среда) данного вида животных согласно воззрениям А. Н. Северцова, является стимулом к изменению его организации; характер же изменения среды, количественное и качественное соотношение между изменением среды и морфофункциональными изменениями организма определяет направление, в котором будет происходить эволюция изменяющегося вида в данную эпоху.
ОСНОВЫ СРАВНИТЕЛЬНОЙ ЭМБРИОЛОГИИ Прогенез
Половые клетки (гаметы)
Зрелые половые клетки в отличие от соматических содержат одиночный (гаплоидный) набор хромосом. Все хромосомы гаметы, за исключением одной половой, называются аутосомами. В муж-
ских половых клетках у млекопитающих содержатся половые хромосомы либо X, либо Y, в женских половых клетках — только хромосома X. Дифференцированные гаметы обладают невысоким уровнем метаболизма и неспособны к размножению.
Мужские половые клетки
Мужские половые клетки — сперматозоиды, или спермин, развиваются в очень большом количестве: выделяющаяся при эякуляции семенная жидкость содержит несколько миллионов сперматозоидов. Они невелики по размерам. У человека их размер достигает 70 мкм. Сперматозоиды обладают способностью к активному движению. Скорость их движения у человека 30— 50 мкм/с. Мужские половые клетки имеют жгутиковую форму.
Строение. В сперматозоиде различают головку и хвост (рис. 23). Головка сперматозоида (caput spermatozoidi) включает небольшое плотное ядро, окруженное тонким слоем цитоплазмы. Ядра сперматозоидов характеризуются высоким содержанием нуклеопрота-минов и нуклеогистонов. Передняя половина ядра покрыта плоским мешочком, составляющим "чехлик" сперматозоида. В нем у переднего полюса располагается акросома (от греч. acron — верхушка, soma—тело). Чехлик и акросома являются производными комплекса Гольджи. Акросома содержит набор ферментов, среди которых важное место принадлежит гиалуронидазе и про-теазам, способным растворять оболочки, покрывающие яйцеклетку. Важно отметить, что у высших позвоночных способность к оплодотворению (явление капацитации сперматозоидов) приобретается постепенно, по мере их продвижения по репродуктивному женскому тракту.
За головкой имеется кольцевидное сужение. Головка так же, как и хвостовой отдел, покрыта клеточной мембраной.
Хвостовой отдел (flagellum) сперматозоида состоит из связующих, промежуточных, главной и терминальной частей.
В связующей части (pars conjungens) или шейке (cervix) располагаются центриоли — проксимальная и дистальная, от которой начинается осевая нить (ахопета). Промежуточная часть (pars intermedia) содержит 2 центральных и 9 пар периферических микротрубочек', окруженных расположенными по спирали митохондриями (митохондриальное влагалище — vagina mitochond-rialis). Именно митохондрии обеспечивают энергией двигательную активность сперматозоидов, нарушение которой нередко связано с поражением процесса энергообразования в митохондриях. Движения хвостового жгутика бичеобразны. Они обусловлены последовательным изменением белков микротрубочек (динеин и др.). Эти белки обладают АТФ-азной активностью и расщепляют АТФ,
У многих животных между центральными и периферическими микротрубочками имеется еще 9 одиночных фибрилл.
М
Рис. 23. Строение мужских половых клеток.
А -- спермин в двух плоскостях; Б — ультрамикроскопическое строение спермиев; В — фрагмент главного отдела хвоста: Г —' фрагмент дистального отдела хвоста. / _ головка: // — хвост; а — связующий отдел (шейка): б—промежуточный отдел; а— главный отдел; г — дистальный отдел; / — цитолемма; 2 — акросома; 2а — акросо-мальный пузырек; } — ядро; 4 - прокси-мальная центриоль; S — дистальная цент-риоль; и — митохондрии; 7 — осевая нить: S — циркулярные фибриллы; 9 — периферические микротрубочки: 10 — центральные микротрубочки.
вырабатываемую митохондриями. Освобождаемая при этом энергия используется для сокращения белков и обеспечения подвижности спермиев в жидкой среде. Среди факторов, влияющих на скорость движения, большое значение имеют степень зрелости спермиев, температура и рН среды.
Главная часть (pars principalis) по строению напоминает ресничку. Окружена тонкофибриллярным влагалищем (vagina tibrosa). Терминальная, или конечная, часть (pars terminalis) содержит единичные сократительные филаменты.
Сперматозоиды животных отличаются друг от друга соотношением указанных отделов и главным образом формой головки. Продолжительность жизни и оплодотворяющая способность сперматозоидов после эякуляции в определенных оптимальных условиях неодинаковы у различных животных. У млекопитающих они варьируют от нескольких часов до нескольких суток. В кислой
среде сперматозоиды быстро утрачивают способность к движению, оплодотворению и склеиваются. Способность к оплодотворению зависит также от концентрации сперматозоидов в семенной жидкости, продолжительности их пребывания в эякуляте и др.
Женские половые клетки. Классификация
Яйцеклетки, или овоциты (от лат. ovum — яйцо), созревают в неизмеримо меньшем количестве, чем сперматозоиды. Для некоторых млекопитающих количество созревающих в течение всей жизни яйцеклеток исчисляется сотнями. У других позвоночных их может быть гораздо больше (например, у рыб и амфибий). Как правило, яйцеклетки имеют шаровидную форму, больший объем цитоплазмы, чем у спермиев, они не обладают способностью самостоятельно передвигаться.
Характерным для яйцеклеток является наличие желтка (lecithos) (белково-липидных включений) в цитоплазме. В зависимости от количества желтка размеры яйцеклеток колеблются от нескольких микрометров до нескольких сантиметров (например, яйцеклетки птиц, акуловых рыб). Яйцеклетки классифицируют на безжелтковые (алецитальные), маложелтковые (олиголеци-тальные) и многожелтковые (полилецитальные). Маложелтковые яйцеклетки подразделяются на первичные (у примитивных хордовых, например ланцетника) и в торичн ы е (у млекопитающих и человека). Количество желтка в цитоплазме находится в прямой зависимости от условий развития животного (во внешней или внутренней среде) и продолжительности развития во внешней среде (рис. 24).
Как правило, в маложелтковых яйцеклетках желточные включения (гранулы, пластинки) распределены равномерно, поэтому они называются еще изолецитальными (греч. isos—равный). У большинства полилецитальных яйцеклеток желток в большей или меньшей степени сосредоточен у одного полюса (вегетативного), а орга-неллы—у противоположного (анимального). Такие яйцеклетки называются телолецитальными (греч. thelos—конец), а если желдгок находится в центре клетки — центролецитальными. Среди телолецитальных различают умеренно телолецитальные — мезо-лецитальные (например, у амфибий) и резко телолецитальные (например, у птиц).
У животных, ведущих наземное существование, организация яйцеклеток сложная. В частности, у пресмыкающихся и птиц яйцеклетки резко телолецитальны. Размеры яйцеклетки большие. Наземное развитие привело к возникновению вторичных и третичных оболочек, предохраняющих яйцеклетку от повреждающего действия механических, температурных и других факторов окружающей среды (пресмыкающиеся, птицы).
У плацентарных млекопитающих в связи с внутриутробным развитием и питанием за счет материнского организма отпала необходимость создания сколько-нибудь значительных запасов желтка
Рис. 24. Строение женских половых клеток. А — различные типы яйцеклеток: я — первично изолецитальная у ланцетника; б — умеренно телолецитальная у лягушки; в — резко телолецитальная у птицы; г — вторично изолеци-талькая у человека; 7 — ядро; 2 — цитоплазма; 3 — желточные зерна; 4 — желточные пластинки (масштаб не соблюден). Б—схема микроскопического (а) и ультрамикроскопического (б) строения яйцеклетки: 1 — ядро; 2 — цитоплазма с желточными включениями; 3 — кортикальные гранулы; 4 — цитолемма; 5 — микроворсинки цитолеммы; 6 — блестящая оболочка; 7 — фолликулярные клетки с отростками, формирующими лучистый венец.
в яйцеклетке. Поэтому вторично в эволюции появились маложелтковые яйцеклетки. Исключением являются яйцеклетки представителей примитивных млекопитающих (клоачные, отчасти сумчатые). Эти животные сохраняют многие черты своих предков — пресмыкающихся, в том числе и резко телолецитальные яйцеклетки. Яйцеклетка плацентарных млекопитающих относительно небольшая, диаметром 50—150 мкм, окружена прозрачной зоной (zona pellucida) и слоем фолликулярных клеток, принимающих участие в ее питании (см. рис. 24).
Строение. Яйцеклетка содержит ядро, цитоплазму (ооплазму), включающую в том или ином количестве питательный материал —
желток и оболочки. Все яйцеклетки имеют цитолемму (оволем-му), или первичную оболочку, а многие еще окружены вторичной (углеводно-белковой) и некоторые — третичной (скорлуповой, подскорлуповой) оболочками. Строение яйцеклеток характеризуется полярностью, которая выражена тем сильнее, чем больше желтка в клетке, например, у птиц. Та часть яйцеклетки, в которой накапливается желток, составляет вегетативный полюс, а противоположная, куда смещается ядро, — анимальный. Поверхность яйцеклетки покрыта микроворсинками.
Ядро женской половой клетки имеет гаплоидный набор хромосом. В период роста ооцита в ядре происходят интенсивные синтетические процессы амплификации генов синтеза РНК — образование многочисленных копий с тех участков хромосомной ДНК, которые кодируют рибосомную РНК. Копии ДНК замыкаются в кольца и смещаются к периферии ядра. На них возникают новые копии ДНК, которые в виде ядрышек выходят в цитоплазму, где становятся центрами усиленного синтеза рРНК и иРНК. Большая часть копий ДНК оказывается заблокированной белковыми молекулами (информосомы) до наступления оплодотворения.
Особенностью ооцитов является накопление огромных запасов химических компонентов аппарата трансляции: рибосом, иРНК, тРНК, количество которых в сотни и тысячи раз может превысить содержание их в соматических клетках. В цитоплазме яйцеклеток накапливаются также запасы разнообразных белков: гистонов, структурных белков рибосом, тубулина, липофосфопро-теидов желтка.
Среди органелл в яйцеклетках разных животных хорошо развита эндоплазматическая сеть. Количество митохондрий умеренно. Комплекс Гольджи на ранних стадиях развития яйцеклетки располагается около ядра, а в ходе созревания яйцеклетки смещается на периферию цитоплазмы. Здесь располагаются небольшие кортикальные гранулы (granula corticalia), содержащие гликозаминогли-каны. В цитоплазме яйцеклеток млекопитающих постоянно выявляются мультивезикулярные тельца. Из включений ооплазмы особого внимания заслуживает желток — питательный материал, во многом определяющий характер эмбриогенеза. Желток выявляется в виде гранул или более крупных шаров и пластинок, образованных фосфолипидами, протеинами и углеводами. Структурной единицей желтка является комплекс липовителлина (липопротеида) и фос-фовитина (фосфопротеина). Каждая пластинка состоит из более плотной центральной и более рыхлой периферической зон, снаружи ограничена осмиофильной мембраной. Плотная зона образована молекулами фосфовитина, имеет вид кристаллической решетки. Образуется желток при непосредственном участии эндоплазматиче-ской сети и комплекса Гольджи.
В процессе роста и созревания яйцеклеток в яичнике они окружаются слоем плоских или кубических клеток, называемых фолликулярными. За счет деятельности ооцита и фолликулярных клеток вокруг яйцеклетки образуется зона, богатая гликозаминогликанами.
У млекопитающих она называется прозрачной зоной (zona pellu-cida).
Фолликулярные клетки посылают через прозрачную зону длинные отростки, направленные к ооциту. В свою очередь цитолемма ооцита имеет микроворсинки, располагающиеся между отростками фолликулярных клеток (см. рис. 24, Б). Фолликулярные клетки выделяют вещества, которые поглощаются яйцеклеткой и способствуют ее росту. Фолликулярный эпителий выполняет также защитную функцию.
Эмбриогенез
Развитие зародыша происходит стадийно с постепенными качественными и количественными изменениями. Различают следующие стадии: оплодотворе ни е, дробление и образование бластулы, гаструляция и д ифференцировк а зародышевых листков с образованием зачатков тканей (г и с-т о гене з), органов (органогенез) и систем органов (с и стемогене з) плода.
Оплодотворение
Оплодотворение (fertilisatio) — слияние мужской и женской половых клеток, в результате чего восстанавливается диплоид-ный набор хромосом, характерный для данного вида животных, и возникает качественно новая клетка — зигота (оплодотворенная яйцеклетка или одноклеточный зародыш).
В зиготе масса ядра увеличивается вдвое, а объем цитоплазмы практически остается тот же, особенно при оплодотворении телоцитальных яйцеклеток. Оплодотворению предшествует о с е-менение— излияние семенной жидкости в половые пути при внутреннем оплодотворении или в среду, где находятся яйцеклетки, при наружном оплодотворении.
В процессе оплодотворения различают три фазы: 1) дистант-ное взаимодействие и сближение гамет; 2) контактное взаимодействие и активизация яйцеклетки; 3) вхождение сперматозоида в яйцо и последующее слияние — сингамия.
Первая фаза— дистантное взаимодействие — обеспечивается совокупностью ряда неспецифических факторов, повышающих вероятность столкновения половых клеток. Важную роль в этом играют химические вещества, вырабатываемые половыми клетками, — гамоны: гиногамоны (1, II), вырабатываемые яйцеклетками и андрогамоны (1, II), продуцируемые спермиями. Гиногамоны 1 (низкомолекулярные вещества небелковой природы, выделяемые яйцеклетками, активизируют движение спермиев. Гиногамоны II (фертилизины) — видоспецифические белки, вызывающие склеивание спермиев при их реакции с комплементарным андрогамоном II, встроенным в цитолему спермия. Склеивание
спермиев предохраняет яйцеклетку от проникновения многих спермиев.
Андрогамоны 1 — антагонисты гиногамонов 1 — вещества небелковой природы, подавляют подвижность спермиев.
Вторая фаза— контактное взаимодействие гамет и проникновение спермия в яйцеклетку — осуществляется с помощью акросомы и ее ферментов спермолизинов. Плазматические мембраны в месте контакта половых клеток сливаются и происходит плазмогамия — объединение цитоплазм обеих гамет.
У млекопитающих при оплодотворении в яйцеклетку проникает лишь один сперматозоид. Такое явление называют моноспер-мией. У беспозвоночных животных, рыб, хвостатых амфибий, рептилий и птиц возможна полиспермия, когда в яйцеклетку проникает несколько сперматозоидов, однако сливается с ядром яйцеклетки ядро только одного спермия. Оплодотворению способствуют тысячи других принимающих участие в осеменении сперматозоидов. Ферменты, выделяемые из акросом, — спермолизины (трипсин, гиалуронидаза), разрушают лучистый венец, расщепляют гликозаминогликаны вторичной (блестящей) оболочки яйцеклетки. Отделяющиеся фолликулярные клетки склеиваются в конгломерат, который вслед за яйцеклеткой перемещается по трубе благодаря мерцанию ресничек эпителиальных клеток слизистой оболочки.
Третья фаза. В ооплазму проникает головка и промежуточная часть хвостового отдела. После вхождения сперматозоида на периферии ооплазмы происходит уплотнение ее (кортикальная реакция) и образуется оболочка оплодотворения (рис. 25).
Как показано на беспозвоночных, механизм кортикальной реакции включает: приток ионов натрия через участок мембраны сперматозоида, встроенный в поверхность яйцеклетки после завершения акросомальной реакции. В результате отрицательный мембранный потенциал клетки становится слабоположительным. Приток ионов натрия обусловливает высвобождение ионов кальция из внутриклеточных депо и увеличение его содержания в цитоплазме яйцеклетки. Вслед за этим начинается экзоцитоз кортикальных гранул. Освобождающиеся из них протеолитические ферменты разрывают связи между блестящей оболочкой (или желточной оболочки у амфибий и птиц) и плазмолеммой яйцеклетки, а также между спермиями и прозрачной оболочкой. Кроме того, выделяется гликопротеид, связывающий воду и привлекающий ее в пространство между плазмолеммой и блестящей оболочкой. Вследствие этого формируется перивителлиновое пространство. Наконец, выделяется фактор, способствующий затвердению прозрачной оболочки и образованию из нее оболочки оплодотворения ("membrana fertilisationis).
Кортикальная реакция — один из механизмов, препятствующий проникновению в яйцеклетку других сперматозоидов. Проникновение сперматозоида через несколько минут значи-
тельно усиливает процессы внутриклеточного обмена, что связано с активизацией ферментативных систем яйцеклетки, в частности окислительно-восстановительных, а позднее — белковых синтезов.
Зигота. Ооплазматическая сегрегация. Образование мужского и женского пронуклеусов
Вслед за проникновением спермия в яйцеклетку и усилением окислительно-восстановительных реакций начинается интенсивное перемещение составных частей цитоплазмы (ооплазмы) с образованием зон повышенной концентрации желточных и пигментных гранул, органелл, что носит название о о плазмат и-ческой сегрегации. Методом маркировки установлено, что в ходе дальнейшего развития каждый участок оплодотворенной яйцеклетки даст начало определенной структуре зародыша.
04
Такие участки называются презумптивными (от лат. praesumptio — предположение, основанное на вероятности).
Попавшая в яйцеклетку головка спермия поворачивается на 180°, ядро постепенно набухает, округляется, хроматин разрыхляется и оно превращается в мужской пронуклеус. Центриоли, внесенные мужской половой клеткой, становятся при этом центром движения внутри оплодотворенной яйцеклетки (зиготы).
Ядро женской половой клетки, имеющее также гаплоидный набор хромосом, набухает, превращается в женский пронуклеус. Пронуклеусы сближаются. При этом в них происходит реплика-ция ДНК. В конце сближения происходит спирализация хромосом, образование метафазной пластинки из двух гаплоидных про-нуклеусов. Объединение двух пронуклеусов —синкарион (от греч. sin — связь, karyon — ядро) — приводит к восстановлению характерного для данной особи животного или человека диплоидного набора хромосом. Таким образом, зигота приобретает гены, унаследованные от обоих родителей. В реализации наследственной информации, кроме ядер, половых клеток, важная роль принадлежит цитоплазме клетки. Об этом свидетельствуют эксперименты с пересадкой ядер соматических клеток в яйцеклетку. При этом пол развивающегося организма зависит от половых хромосом. При слиянии яйцеклетки со сперматозоидом, несущим хромосому X, образуется женская особь, а при слиянии со сперматозоидом, имеющим хромосому Y, — мужская особь.
Дробление
Дробление (fissio) — последовательное митотическое деление зиготы на клетки (бластомеры) бс? последующего роста их до размеров материнской.
Вследствие фактического отсутствия интерфазы g[-периода, во время которого происходит рост клеток, образовавшиеся в результате деления клетки гораздо меньше материнской, поэтому и величина зародыша в целом в этот период независимо от составляющих его клеток не превышает величину исходной клетки — зиготы. Все это позволило назвать описываемый процесс дроблением, измельчением, а клетки, образующиеся в результате дробления, — бластомерами (от греч. blastos — зачаток, meros — часть).
На ранних стадиях все бластомеры сохраняют способность к развитию при определенных условиях в самостоятельный организм, или, как принято говорить, они тотипотентны. Дробление (уменьшение размеров бластомеров) продолжается до тех пор, пока не восстановится характерное для соматических клеток данного вида животного соотношение ядра и цитоплазмы. После этого наступает дерепрессия синтеза белка, и каждая дочерняя клетка увеличивается до размеров материнской. Дробление зародыша происходит неодинаково у различных
9S
позвоночных, что определяется прежде всего количеством и характером распределения желтка в яйцеклетке.
Существует определенный строгий порядок появления борозд дробления. Борозды и плоскости попеременно проходят через анимальный и вегетативный полюса клетки (меридианное направление), поперечно (широтные) или параллельно поверхности (тангенциальные). Чем больше желтка в яйцеклетке у различных видов животных, тем менее полно и менее равномерно происходит дробление (рис. 26).
Первично олиголецитальные изолецитальные яйцеклетки дробятся полно и равномерно. В мезолецитальных яйцеклетках дробление полное, но неравномерное, так как в вегетативной части, где сосредоточен желток, дробление происходит медленнее, чем на анимальном полюсе, и неполно. В резко телолецитальных яйцеклетках дробление частичное — меробластическое. Например, у птиц дробится лишь часть яйцеклетки у анимального полюса'. Для вторично олиголецитальных, изолецитальных яйцеклеток плацентарных млекопитающих и человека характерно полное, или г о лобластическо е, асинхронное, неравномерное дробление. Дробление происходит во время движения зиготы по яйцеводу, причем количество бластомеров нарастает в неправильном и притом у различных животных неодинаковом порядке (2, 3, 5, 10, 13, 17 и т. д.). В результате дробления образуется многоклеточный зародыш, сначала в форме плотного скопления клеток (мору ла), а затем в виде пузырька с небольшой полостью—бластоциста (бластула) (см. рис. 26).
Дробление зародышей требует соблюдения оптимальных условий среды (химический состав, осмотическое давление, температура, содержание кислорода и др.) Зародыши обладают высокой чувствительностью к химическим, физическим и другим повреждающим факторам, которые могут привести к мутациям.
Бластула ("blastula) имеет стенку — бластодерму и полость — бластоцель, заполненную жидкостью — продуктом секреции бластомеров. В бластодерме различают крышу, образовавшуюся за счет раздробившегося материала анимального полюса, дно — из материала вегетативного полюса и краевую зону, расположенную между ними.
При полном равномерном дроблении (например, ланцетника) бластула имеет однослойную бластодерму, а бластоцель находится в центре. Такая бластула называется целобластулой. В результате полного неравномерного дробления (минога, лягушка) образуется бластула с многослойной бластодермой и эксцентрично расположенным бластоцелем — амфибластула. Крыша такой бластулы, состоящая из мелких бластомеров, сравнительно тонкая,
studfiles.net
Эмбриология
План:
1.История развития и значение эмбриологии.
2. Развитие и строение половых клеток.
3. Оплодотворение.
4. Ранние этапы эмбриогенеза. Дробление.
5. Гаструляция и закладка осевых органов.
6.Формирование внезародышевых органов.
7.Эмбриогенез птиц.
8. Эмбриогенез млекопитающих.
Эмбриология (embryon- зародыш, logos- учение)- наука о закономерностях развития организма животных от момента оплодотворения яйцеклетки и образования зиготы до рождения или вылупления из яйца.
Начало развития эмбриологии было положено в Греции более 2-х тысяч лет назад. Впервые Гиппократ описал развитие зародыша в курином яйце и пытался понять процесс развития эмбриона у млекопитающих.
Позднее Аристотель достаточно полно описал процесс развития у млекопитающих внутренних органов в эмбриогенезе. Описал функции плаценты и пуповины. Им впервые было выявлено, что в начальный период развития в организме появляются общие черты, свойственные животным вообще, а позднее формируются частные признаки, характерные данному типу, или виду животных.
С изобретением микроскопа в 17 веке Левенчук открыл спермии, а Грааф описал фолликулы в яичнике, приняв их за яйцеклетку. И только спустя 150 лет были обнаружены внутри фолликулов яйцеклетки.
В дальнейшем в развитие эмбриологии внесли вклад многие ученые, в том числе и работавшие в России (Вольф, Пандр, Бер, Ковалевский, Северцов, Боголюбский и др.)
Особенно бурное развитие получила эмбриология в последние 50 лет в связи с использованием современных методов исследования (электронной микроскопии, гистохимии, гистоавторадиографии, микрохирургии, культуры тканей и др.
Достижения современной эмбриологии нашли широкое применение в практике животноводства и ветеринарии.Это искусственное осеменение животных, стимуляция многоплодия, трансплантация эмбрионов. Генно- инженерные манипуляции позволили ученым получить животное от соматической клетки (Овечка Долли в Шотландии).
Знание эмбриологии позволяет ветеринарным врачам выяснять причины бесплодия и других вопросов акушерства, что необходимо для эффективного лечения животных, повышения их плодовитости и тем самым ускорения воспроизводства животных.
Развитие и строение половых клеток.
Процесс развития половых клеток называют прогенезом. Он происходит в половых железах (гонадах)- яичниках и семенниках. Родоначальником половых клеток являются гаметобласты.
Они формируются у животного уже в зародышевый период развития из клеток первичной эктодермы в стенке желточного мешка.
Отсюда с током крови гаметоблаты мигрируют в зачатки половых желез, где в последствии из них развиваются спермии и яйцеклетки. Процесс образования спермиев называется сперматогенезом, а яйцеклеток- овогенезом.
Сперматогенез- происходит в извитых канальцах семенников и завершается в половых путях самца.
Весь процесс включает 4 последовательных периода: размножение, рост, созревание и формирование.
В период размножения первичные половые клетки (сперматогонии) усиленно делятся путем митоза и располагаются по периферии семенного канальца. Они связаны между собой цитоплазматическими мостиками, формируя синцитий. Затем отдельные сперматогонии перестают делиться и вступают во второй период- роста.
Период роста. В этот период сперматогонии увеличиваются в объеме, в их цитоплазме происходит активный синтез нуклеиновых кислот, белков, углеводов, увеличивается ядро, количество митохондрий. Они становятся сперматоцитами 1-го порядка и вступают в тесный контакт с поддерживающими клетками Сертоли (сустентоциты), выполняющими опорную, трофическую и защитную функции.
В период созревания происходит мейоз состоящий из двух последовательных делений:
1)редукционное, при котором из сперматоцитов 1-го порядка образуются сперамтоциты 2-го порядка с гаплоидным набором хромосом.
2)Сперматоциты 2-го порядка сразу же вступают во 2-ое (эквационное деление). В результате которого из каждого сперматоцита 2-го порядка образуется два сперматида, имеющие также гаплоидный набор хромосом. Таким образом из каждого сперматоцита 1-го порядка образуется 4 сперматида. Половые Х и У хромосомы расходятся по разным сперматидам, в связи с чем два сперматида имеют Х- хромосомы, а 2- У хромосомы (пол зависит от спермия).
4-ый период формирования характеризуется перегруппировкой органелл и изменением ядра. Ядро уплотняется и становится головкой будущего спермия. Над ядром в зоне комплекса Гольджи образуется уплотненная гранула (акробласт), который увеличиваясь в размере покрывает ядро в виде чехлика, образуя мембранную шапочку. Внутри этого чехлика из аппарата Гольджи формируется акросома, уплотненное тельце, богатое ферментами гиалуронидазой и трипсином (протеазы). Ферменты играют важную роль в оплодотворении. К противоположному от ядра полюсу перемещается центросома, в которой различают проксимальную и дистальную центриоли.
Они располагаются в области шейки спермия. Дистальная центриоль делится на две части. От передней отрастает осевая нить хвостика, состоящая из микротрубочек (9*2+2). Задняя часть образует колечки и сползает по осевой нити хвостика, увлекая за собой цитоплазму, митохондрии и капельки гликогена.
Миттохондрии и гликоген располагаются в начальном отделе хвостика. Цитоплазма, смещаясь по хвостику тонким слоем одевает главную его часть. Конец хвостика покрыт только плазмолеммой.
Клетка продолжает удлиняться и приобретает вид спермия. (Демонстрация схемы строения спермия и рисунок на доске).
Таким образом спермии имеют жгутиковую форму. Подвижность хвостика обусловленна осевой нитью, являющейся производной и продолжением центриоли.
Комплекс трубочек, образующих осевую нить называют аксонемой (9*2+2).
Исследования спермы очень важны в зоотехнии и ветеринарии. В одном эякуляре от 20 до 200 млн. спермиев.
С целью успешного оплодотворения путем искусственного осеменения изучают густоту спермиев, их форму и подвижность.
Создаются банки спермиев от ценных племенных производителей.
Ср. р-р сперматозоидов- 70 мкм движутся со скоростью 50 мкм в 1 час.
Овогенез.
Развитие женских половых клеток происходит в яичниках и завершается в яйцеводе.
В отличие от сперматогенеза, овогенез включает 3 периода: размножения, роста, созревания. Период размножения протекает во время внутриутробного развития самки и завершается в течении первых месяцев после рождения.
Гаметобласты усиленно делятся и превращаются в овогонии с диплоидным набором хромосом. Некоторые овогонии прекращают делиться и вступают в период роста. Они быстро увеличиваются в размерах. В них происходит активный синтез нуклеиновых кислот, белков, углеводов. Увеличивается количество органелл и овогонии превращаются в овоциты 1-го порядка. Вначале они окружаются одним слоем фолликулярных клеток и овоцит 1-го порядка называется первичным фолликулом. Затем он покрывается несколькими слоями фолликулярных клеток и становится вторичным фолликулом.
Вещество, выделяемое фолликулярными клетками и самим овоцитом образует вокруг овоцита 1-го порядка блестящую оболочку (зону пеллюцида).
Фолликулярные клетки доставляют овоциту белки, липиды, углеводы для синтеза желтка.
Вблизи цитолеммы в овоците формируется кортикальный слой, состоящий из плотных гранул с ферментами, которые играют важную роль в оплодотворении.
Стадия роста отличается длительностью и сложностью процессов, происходящих в яйцеклетке.
До полового созревания самки активно идет формирование рибосом, синтетического аппарата. Затем с наступлением половой зрелости самки, накапливается большое количество желточных гранул. В состав желточных гранул входит фосфолитин (белок, насыщенный фосфором) и липовитилин (белок в сочетании с липидами).
В блестящем слое с помощью фолликулярных клеток и самой яйцеклетки накапливаются гликопротеиды. Ученые обнаружили три их формы, одна из которых притягивает сперматозоиды, т.к. у спермиев есть рецепторы, которые реагируют на эту форму гликопротеида и они устремляются к яйцеклетке, окружая её в огромном количестве (до 4 тысяч).
От соматических клеток яйцеклетка отличается:
1. Гораздо большими размерами (130-200 мкм).
2. Наличием большого количества желточных гранул.
3. Наличием кортикального слоя под плазмолеммой.
4. Наличием большого запаса РНК, тубулина, липофосфопротеидов и др.
5. В процессе созревания исчезают центриоли.
6. В женских яйцеклетках только Х- хромосома.
7. Наличием трех оболочек: плазмолеммы, блестящей и фолликулярной (лучистый венец).
8. В ядре хорошо выражено ядрышко и поровый комплекс.
9. Гаплоидным набором хромосом.
По количеству желтка различают яйцеклетки:
1) Олигоцитальные- с малым количеством желтка ( у ланцетника и млекопитающих).
2) Полилецитальные- с большим количеством желтка (у пресмыкающихся и птиц).
По характеру распределения желтка:
1) Изолицитальные- с равномерным распределением (первично- у ланцетника, вторично- у млекопитающих).
2) Телолицетальные- с локализацией желтка у одного полюса (у птиц и пресмыкающихся).
Верхняя часть яйцеклеток называется аномальным полюсом, а нижняя- вегетативным. Желток скапливается в вегетативном полюсе.
Как только накопится достаточное количество желтка рост ооцита 1-го порядка прекращается, а фолликул продолжает расти. В нем образуется полость, заполненная жидкостью, содержащей женские половые гормоны (эстроген), и он становится пузырчатым. Затем разрывается и яйцеклетка выходит в яйцевод.
3 стадия созревания- у большинства млекопитающих начинается в яичнике, т.е. до овуляции и заканчивается в яйцеводе. Проникшие в половые пути самки спермии стимулируют процесс мейоза. Иногда он завершается когда спермий уже внедрился в цитоплазму яйцеклетки.
В результате двух последовательных делений (редукционного и эквационного) образуется один овоцит 2-го порядка с гаплоидным набором хромосом и 3 направительных тельца.
Оплодотворение.
Это процесс слияния мужской и женской половых клеток, в результате которого образуется зигота с диплоидным объединенным набором хромосом, которая дает начало новому организму.
Оплодотворение протекает в 4 стадии:
1. Сближение половых клеток.
2. Проникновение спермиев в блестящую оболочку.
3. Проникновение в цитоплазму.
4. Слияние ядер половых клеток (синкарион).
1)Движению спермиев к яйцеклетке способствуют колебательные движения хвостика, реотаксис, способность двигаться против тока жидкости и присасывающее действие гладкой мускулатуры матки и яйцеводов, выделяющиеся яйцеклеткой вещества (гинагомоны), наличие в блестящей оболочке специфических рецепторов (гликопротеидов- ZP3). У спермиев есть рецепторы, которые реагируют на эти гликопротеиды и они во множестве (до 4-5 тыс.) окружают яйцеклетку.
2)Выделяющееся яйцеклеткой вещество- фиртилизин вызывает акросомальную реакцию. Плазмолемма головки спермия сливается с наружной мембраной акросомы и ферменты (гиалуронидаза) изливаются наружу, разрушают блестящую оболочку и спермии проникают внутрь самой яйцеклетки. В момент прикосновения спермия к плазмолемме яйцеклетки на её поверхности образуется выпячивание (бугорок оплодотворения).
3) Далее наступает 3-я стадия оплодотворения. Приблизившись к поверхности яйцеклетки, один из спермиев прикасается к её цитолемме боковой поверхностью головки. В месте контакта мембрана спермия и яйцеклетки частично разрушаются. В результате головка, шейка и начальная часть хвостика проникают в цитоплазму яйцеклетки.
Как только спермий проникнет в цитоплазму, приходят в действие кортикальные пузырьки. Они сливаются с плазмолеммой и изливают ферменты в привитилиновое пространство (область между плазмолеммой и блестящей оболочкой). Ферменты воздействуют на гликопротеиды (рецепторы привлекшие спермии) и в результате изменяют их химическую структуру и блестящая оболочка не пропускает другие спермии, становится оболочкой оплодотворения, хотя по виду остается прежней.
4) Слияние двух половых клеток. Головка проникшего в цитоплазму спермия постепенно набухает, округляется, превращаясь в гаплоидное круглое ядро или мужской пронуклеус, который вместе со своей центросомой продвигается к центру яйца. Сюда же после завершения мейоза движется и ядро яйца- женский пронуклеус. Происходит слияние пронуклеусов (синкарион) и удваивается число хромосом. Женская и мужская наследственность объединились и дали начало новому организму.
Центриоли, внесенные спермием расходятся, образуя веретено деления. Ядерная оболочка исчезает, отцовские и материнские хромосомы формируют звезду первого митотического деления оплодотворенной яйцеклетки и начинается дробление- первый этап эмбриогенеза.
Зигота в течении нескольких дней остается окруженной блестящей оболочкой, поэтому дробление не приводит к увеличению объема зиготы.
Дочерние клетки не расходятся. А тесно прилегают друг к другу. Размеры клеток с увеличением их количества уменьшаются, поэтому процесс и называется дроблением.
Деление клеток проходит быстро одно за другим, интерфазы практически нет, поэтому дочерние клетки не дорастают до размеров материнской и становятся все мельче и мельче.
Образующиеся в результате дробления клетки называются бластомерами.
Процесс дробления (с 1 по 4 суток) осуществляется в яйцеводах по пути следования яйца в матку.
У разных животных зиготы дробятся неодинаково. Тип дробления определяется количеством и распределением желтка. Чем больше в цитоплазме желточных гранул, тем медленнее делится соответствующая часть зиготы. Желток задерживает процесс дробления. При большом содержании желтка деление определенной части зиготы становится невозможным, поэтому дробление бывает полным и неполным.
| Дробление | ||
| Полное Неполное (у птиц) | ||
| Равномерное (у ланцетника) | Неравномерное (у млекопитающих) | |
| Целобластула | Морула | Дискобластула |
| Бластоциста |
При полном дроблении принимает участие весь материал зиготы, при неполном та часть, где нет желтка.
1) Полное дробление бывает равномерным, если бластомеры одинаковой величины и неравномерным- бластомеры разной величины.
Полное равномерное дробление характерно для ланцетника. При этом образуются одинаковые бластомеры, которые располагаются в один слой и образуют зародыш в виде пузырька с с полостью, который называется целобластулой.
2) Полное галобластическое неравномерное асинхронное дробление характерно для млекопитающих с олигоцетальными мезолецитальными яйцеклетками.
В результате дробления у млекопитающих образуется вначале плотный шар, называемый морулой, а затем зародышевый пузырек с небольшой полостью, называемой бластоцистой или серобластулой.
3) Неполное (частичное) дробление свойственно яйцеклеткам птиц.
Дробление происходит только в анимальном полюсе. Образующиеся бластомеры формируют дискобластулу.
Таким образом первый этап эмбриогенеза- дробление заканчивается формированием бластулы.
Гаструляция и закладка осевых органов.
Вслед за дробленим начинается следующий этап эмбриогенеза- гаструляция с образованием зародышевых листков. Зародышевые листки располагаются строго закономерно: эктодерма- наружный листок, энтодерма- внутренний, мезодерма занимает промежуточное положение. В основе гаструляции лежат следующие процессы: размножение, рост, перемещение, взаимодействие клеток и их дифференцировка.
Дифференцировкой называется ряд морфологических и гистохимических изменений в клетках.
Перераспределение клеточного материала при гаструляции происходит 4 способами:
1) инвагинация- впячивание (у ланцетника)
2) Обрастание- эпиболия (у земноводных)
3) Расслоение- деламинация (у млекопитающих
4) Выселение- иммиграция и птиц)
В процессе гаструляции происходит также и формирование мезенхимы- эмбриональной соединительной ткани. Мезенхима состоит из клеток, выселившихся в основном из мезодермы. Клетки мезенхимы заполняют пространство между зародышевыми листками и формирующимися органами.
Из зародышевых листков и мезенхимы происходит формирование всех тканей и органов, т.е. гистогенез и органогенез развивающегося организма.
В начале закладываются осевые зачатки органов- нервная трубка, хорда, кишечная трубка. Хорда- это скелетный тяж, который располагается вдоль тела животного. Позже на месте хорды у птиц и млекопитающих развивается позвоночник.
Нервная трубка формируется над хордой из первичной эктодермы. В начале из изменившихся клеток образуется пластинка, затем желобок и трубка. По бокам формируются нервные валики. Из клеток трубки и валиков формируются ткани и все органы нервной системы, а также органы чувств.
Остальная зародышевая эктодерма является источником развития кожного эпителия зародыша.
Из второго зародышевого листка- энтодермы, образуется кишечная трубка и хорда. Из кишечной трубки развивается эпителий органов пищеварения, органов дыхания и некоторых органов эндокринной системы.
Мезодерма дифференцируется на
1) сомиты, лежащие по бокам нервной трубки и хорды (дерматом, миотом, склеротом)
2) сегментные ножки, (нефрогонотомы) и спланхнотомы, боковые пластинки (висцеральная и париетальная).
Из дерматома развиваются соединительно- тканная часть кожи, из миотома скелетная мускулатура, из склеротома- хрящи и скелет.
Из нефрогонотома развиваются эпителиальные зачатки почек и половых желез.
Из листков спланхнотома образуется эпителий серозных оболочек брюшины, плевры, перикарда.
Из мезенхимы развивается кровь и лимфа, кроветворные органы, сосуды, собственно соединительная ткань, гладкая мышечная ткань.
В процессе органогенеза взаимосвязь и взаимопроникновение зародышевых листков настолько тесные, что в образовании каждого органа принимают участие клеточные элементы почти всех зародышевых листков.
В процессе эмбриогенеза признаки, характерные для животного, появляются постепенно. Вначале у животных разных типов сходен характер возникновения и расположения осевых органов. Затем становятся видны черты, присущие классу, позже отряду, роду, виду, породе и, наконец, индивидууму.
В стадии гаструляции и образования осевых органов происходит формирование внезародышевых органов, которые обеспечивают условия для нормального развития эмбриона. Они развиваются из внезародышевой части зародышевых листков. Внезародышевые органы функционируют только в период эмбрионального развития, поэтому их называют провизорными, т.е. непостоянными.
Они окружают зародыш в виде оболочек. К ним относятся:
1. Желточный мешок образуется энтодермальным и висцеральным листками. Выполняет трофическую, кроветворную и дыхательную функцию.
2. Амнион образован эктодермой и мезодермой, создает вокруг зародыша водную среду, выполняет защитную функцию и участвует в питании плода у птиц.
3. Аллантоис- образован энтодермой и висцеральным листком мезодермы. Вырастает из стенки задней кишки. Это орган газообмена и выделения.
4. Серозная оболочка (у птиц) образуется из эктодермы и париетального листка мезодермы одновременно с амниотической оболочкой. Участвует в снабжении эмбриона кислородом и кроме того выполняет защитную функцию.
5. Хорион- (у млекопитающих) развивается из трофобласта и внезародышевой мезодермы. Мезодерма подрастает к трофобласту и образует вместе с ним вторичные ворсинки и трофобласт превращается в хорион.
6. Плацента- врастая в слизистую оболочку хорион вместе с ней образует плаценту. Функция трофическая, дыхательная, выделительная, гормональная, защитная.
Эмбриогенез птиц.
В яйце птиц собственно яйцеклеткой является желток с зародышевым диском. По количеству желтка яйцеклетка полилецитальная по распределению, резко телолицетальная.
Снаружи желток покрыт первичной тонкой оболочкой. После овуляции (выхода из яичника) вторичная фолликулярная оболочка утрачивается. Яйцеклетка попадает в яйцевод, где оплодотворяется и на неё по мере продвижения начинают наслаиваться третичные оболочки- белковая, подскорлуповая и скорлупа.
В белке формируются два плотных тяжа(халаза) из пучков белково- углеводных нитей, которые поддерживают желток в центральном положении. Подскорлуповая оболочка состоит из двух листков, которые расщепляются на тупом конце, образуя воздушную камеру. Подскорлуповая оболочка состоит из белкового волокнистого материала, скорлупа- из более грубых волокон, пропитаных солями Са.
Спермии в половых путях самки живут и оплодотворяют яйцеклетку в течении 30 дней. В верхней трети яйцевода яйцеклетка оплодотворяется и превращается в зиготу, дробление которой происходит только на анимальном полюсе.
В результате формируется дискобластула и начинается процесс гаструляции. Таким образом, оплодотворенные и снесенные яйца имеют уже двухслойный зародыш. До инкубации или насиживания эмбриогенез приостанавливается. Через 12ч инкубации в центре зародышевого диска образуется зародышевый щиток- утолщенный участок зародышевого диска. Он представлен светлой зоной в которой формируется зародыш и темной зоной, окружающей светлую в виде плотного кольца, из которой в дальнейшем развиваются внезародышевые органы.
В светлой зоне зародышевого щитка клетки энергично передвигаются (мигрируют) двумя потоками в каудальном направлении, затем, соединяясь, передвигаются по средней линии вперед, формируя первичную полоску в форме утолщенного клеточного валика. Затем в середине этой полоски образуется углубление (первичная борозда). На конце которой формируетс утолщение (гензеновский узел).
Клеточные потоки из гензеновского узелка уходят вперед и вглубь и начинают формировать осевые органы птицы. Разрастается хордальный отросток, затем по сторонам от него образуются клеточные тяжи, растущие вперед и в стороны, вклиниваясь между эктодермой и энтодермой, формируя мезодерму- третий зародышевый листок. Из эктодермы формируется нервная трубка.
Таким образом, ранний зародыш состоит из эктодермы, энтодермы, нервной трубки, хорды и мезодермы. Он лежит, распластавшись на поверхности желточного мешка, который окружен энтодермой и висцеральным листком мезодермы.
Из мезодермы в стенкам желточного мешка образуются клетки крови и кровеносные сосуды и он становится аппаратом питания, выполняя трофическую и кроветворную функции.
Из внезародышевых листков стенках раннего зародыша сначала образуется туловищная, а затем амниотическая складки. Туловищная складка приподнимает зародыш над желтком и замыкает энтодерму в кишечную трубку.
Амниотическая складка разрастаясь над зародышем и смыкаясь краями формирует две плодовые оболочки- серозную (из наружной части складки) и амнион (из внутренней части складки).
Серозная оболочка подрастает под скорлупу и участвует в снабжении эмбриона кислородом и в минеральном обмене. С помощью многочисленных ворсинок клетки серозной оболочки контактируют со скорлупой. Они секретируют соляную кислоту, которая вводится в канальцы скорлупы и растворяет соли кальция. Попадая в кровяное русло они приносятся в тело зародыша и участвуют в формировании скелета.
Амнион окружает только зародыш и создает для него водную среду.
Серозная оболочка, разрастаясь окружает и белок, который начинает поступать по серозно- амниотическому каналу в полость амниона на питание зародыша. Эктодерма амниона продуцирует амниотическую жидкость, которая заполняет амниотическую полость, создавая для развивающегося зародыша самую благоприятную водную среду. Амнион выполняет защитную функцию, он сглаживает удары и создает для эмбриона возможность подвижности.
Аллантоис- образуется после заверщения формирования кишечника на вентральной поверхности задней кишки. Он состоит из энтодермы и висцерального листка мезодермы. Аллантоис постепенно заполняет все щели между амнионом, желточным мешком и серозной оболочкой, срастаясь с мезодермальным листком серозы.
В этом участке формируется густая сеть кровеносных сосудов, что способствует увеличению снабжения организма кислородом.
Таким образом, аллантоис, прилегая к скорлупе участвует в газообмене и выполняет функцию выделительного ргана, т.к. в нем скапливаются продукты обмена.
Развитие птиц протекает стадийно. Наиболее важными периодами в развитии птиц являются следующие:
1. Латеральное питание (первые 30-36 часов). Материалом для питания служит желток Латебры, содержащий белок, соли и воду. Источником энергии является гликоген. Отсутствует кровообращение, зародыш в кислороде почти не нужнается.
2. Желточное питание и наличие желточного круга кровообращения (с 30-36 до 7-8 дней инкубации). К 30-му часу закладывается сердце и желточный мешок, в стенках которого развиваются кровеносные сосуды. Запас гликогена исчезает. В организм поступает кислород, что облегчает использование белков и жиров. Развивается нервная система, начинается биение сердца, сокращение мускулатуры тела. Закладка печени обуславливает синтез мочевины, поэтому продукты распада белков становятся менее вредными.
3. Дыхание кислородом и питание белками (с 7-8 до 18-19 дней). Интенсивно развивается и функционирует аллантоис, в стенке которого густая сеть кровеносных сосудов. Прилегая к серозной оболочке, аллантоис обеспечивает снабжение кислородом. Растворяется скорлупа и возрастает интенсивность минерального обмена, усиливается освоение жиров.
4. Потребление кислорода воздуха из воздушной камеры. (18-19 дней до наклева). Переход на легочное дыхание. Начинает функционировать малый круг кровообращения, появляется артериальная кровь. Аллантоис подвергается обратному развитию и цыпленок испытывает недостаток в кислороде и проклевывает скорлупу.
5. Стадия вылупления. Питание остатком желтка, который втягивается в полость кишки. Цыпленок освобождается от скорлупы.
2-ая классификация: зародышевый период (8 дней), предплодный (8-13 дней (наряду с желтком питание кишечное), плодный период (13-20), вылупление (20-21).
Яйцеклетка млекопитающего в отличие от птиц содержит мало желтка (олиголицетальная и вторично изолицетальная) и дробление зиготы полное неравномерное. В результате получаются мелкие и крупные бластомеры. Мелкие выселяются под самую блестящую оболочку, формируя трофобласт, а крупные собираются в кучку, формируя внутри зародышевый узелок или эмбриобласт. Затем с помощью трофобласта внутри зародышевого пузырька начинает накапливаться жидкость, оттесняя эмбриобласт вверх к трофобласту и распластывая его в один слой. Этот слой крупных клеток представляет собой зародышевый диск у млекопитающих, из которого формируются все органы и ткани животного и некоторые внезародышевые органы. Трофобласт выполняет вспомогательную роль. С его помощью зародыш начинает получать питательные вещества из стенки матки и зародышевый пузырек быстро разрастается.
В середине зародышевого диска образуется уплотненная темная зона (зародышевый щиток) из которого развивается тело зародыша, из окружающих светлых клеток формируются внезародышевые органы. В зародышевом щитке начинают происходить те же процессы, что и у птиц (формирование первичной полоски и гензеновского узелка и т.д.). Аналогично формируются и внезародышевые органы.
Только желточный мешок к концу зародышевого периода (от 23 до 80 дней) редуцируется. Но за короткий период эмбриогенеза он выполняет очень важные функции: трофическую, дыхательную, формирует сосудистую сеть, выполняет функцию кроветворения, является местом образования половых клеток, вырабатывает гормоны. .
Аллантоис у млекопитающих не разрастается так сильно, как у птиц и имеет вид длинного канатика, которых с остатком желточного мешка является основой для пупочного канатика. Пупочный канатик образуется из стенок сильно разрастающегося амниона. В процессе роста стенки амниона приближаются к хориону. В том месте, где расположен аллантоис и остатки желточного мешка, стенки амниона сближаясь, зажимают аллантоис, окружают его по всей длине со всех сторон. Так возникает пупочный канатик, на котором как бы в подвешенном состоянии плавает в околоплодной жидкости зародыш.
Типы плацент по характеру строения и взаимоотношения между ворсинками хориона и тканями слизистой оболочки матки:
1) эпителиохориальные (у свиней)- ворсинки хориона контактируют с эпителием маточных желез.
2) Десмохориальные (у жвачных)- ворсинки хориона разрушают эпителий и контактируют с соединительной тканью слизистой матки.
3) Эндотелиохориальные ( у хищных)- ворсинки контактируют с эндотелием сосудов матки.
4) Гемохориальные (у приматов и человека)- ворсины разрушают стенки сосудов и контактируют непосредственно с материнской кровью.
По форме:
1)Диффузная плацента- почти вся поверхность пузыря у зародыша равномерно покрыта ворсинками. Хорион всей своей поверхностью прилегает к стенке матки (у свиньи, верблюда, лошади).
3) Котиледонная плацента- ворсины хориона собраны в группы- котиледоны, между которыми поверхность пузыря гладкая, без ворсинок (у жвачных).
4) Поясная плацента- хорион с разветвленными ворсинами имеет форму широкого пояса, опоясывающего плодный пузырь (у хищников).
5) Дискоидальная- хорион с разветвленными ворсинками в виде диска окружает пуповину.
Этапы эмбриогенеза млекопитающих.
1. Зародышевый период (от дробления до начала органогенеза). У свиней 23, у овец 30, у коров 35, у лошадей 85 дней. В конце этого этапа редуцируется желточный мешок.
2. Предплодный период (происходит активный органогенез. Полное развитие плаценты). У свиньи до 38 дня, у овцы до 45, у коров до 60 дней, у кобылы до 160 дней.
3. Плодный период ( совершенствование структуры и становление функций органов, активный рост плода). У овец до 145-150 дней, у свиней 113-114 дней, у коров 275 дней, у лошадей до 330 дней.
kursak.net
Эмбриогенез человека — это часть его индивидуальногоразвития, онтогенеза. Он тесно связан с прогенезом (образованием половых клеток иранним постэмбриональным развитием. Эмбриология человека изучает процессразвития человека, начиная с оплодотворения и до рождения. Эмбриогенез человека, продолжающийся в среднем 280суток (10 лунных месяцев ), подразделяется натри периода: начальный (первая неделя развития), зародышевый(вторая-восьмая недели), и плодный (с девятойнедели до рождения ребенка). В курсе эмбриологиичеловека на кафедре гистологии более подробно изучаются ранние стадии развития.
В процессеэмбриогенеза можно выделить следующие основные стадии:
1. Оплодотворение~слияние женской и мужской половых клеток. В результате образуется новый одноклеточный организм-зигота.
2. Дробление.Серия быстроследующих друг за другом делений зиготы. Эта стадиязаканчивается образованием многоклеточногозародыша, имеющего у человека форму пузырька-бластоцисты, соответствующей бластуле других позвоночных.
3. Гаструляция.В результате деления, дифференцировки, взаимодействияи перемещения клеток зародыш становится многослойным. Появляются зародышевыелистки эктодерма, энтодерма и мезодерма, несущие в себе накладкиразличных тканей и органов.
4. Гистогенез, органогенез, системогенез.В ходе дифференцировки зародышевых листков образуются зачаткитканей, формирующие органы и системы организма человека.
Половыеклетки. Зрелые половые клетки гаметы, вотличие от соматических содержат гаплоидный набор хромосом (23 хромосомы учеловека). Мужские половые клеткиназываются сперматозоидами или спермиями, женские — яйцеклетками. Все хромосомы гамет называются аутосомамиза исключениемодной — половой. В женских половых клетках содержатся Х-хромосомы.Мужские половые клетки бывают двух типов — одни спермии содержат Х-хромосому, а другие У-хромосому, Мужские половые клетки человека имеют размеры 70 мкм. Развиваются исозревают они в яичках мужчины в больших количествах.В 3 мл эиякулята всреднем содержится 350 млн. спермиев. Мужские половые клетки очень подвижны, особенно с У-хромосомой. За 1,5-2 часа они могут достигать маточной трубы,где происходит созревание женской половой клетки и оплодотворение. Спермии сохраняют оплодотворяющую способностьв половых путях женщиныдвое суток. Мужские половыеклетки состоят из головки и хвостика, вкотором различают связующую (или шейку), промежуточную (тело), главную и терминальные части. В головке расположеноплотное ядро, окруженноенебольшим ободком цитоплазмы. Спереди ядро покрыто плоскиммешочком- “чехликом>>. в котором у переднего полюса
расположена акросома. Чехликс хромосомой является производным комплекса Гольджи.В акросоме содержитсянабор ферментов, среди которых гиалуронидаза и протеазы, способные растворять оболочки яйцеклетки, В связующей части спермия в цитоплазмерасполагаются проксимальнаяцентриоль и дистальная, от которой начинаетсяосевая нить, аксонема. В промежуточном отделе (теле) осевая нить (2 центральныхи 9 пар периферических трубочек) окружена расположеннымипо спирали митохондриями, обеспечивающимиэнергетику спермия. Главнаячасть хвостика по строению напоминаетресничку, окруженную тонкофибриллярнымвлагалищем. В терминальнойчасти хвостика содержатся единичные сократительныефибриллы.
Женские половыеклетки,яйцеклетки, классифицируются по количеству и расположению желтка, находящегося в их цитоплазме. Количество желтка зависит от условий и продолжительности развития эмбриона,
ТИПЫ ЯЙЦЕКЛЕТОК
1. Алецитальная (безжелтковая).
2. Олиголецитальная (маложелтковая),в них желток равномерно распределен по цитоплазме, поэтому их называют изолецитальными. Среди них различают первично изолецитальные (у ланцетника)и вторично изолецитальные(у млекопитающих нчеловека),
3.Полилецитальные (многожелтковые)
Желток в этихяйцеклетках может быть сосредоточен в центре — это центролецитальные клетки.Среди телолецитальных яйцеклеток в свою очередь различают умереннотелолецитальные или мезолецитальные со среднимсодержанием желтка(у амфибий) и резко телолецитальные, перегруженные желтком откоторого свободна лишь небольшая часть анимального полюса (у птиц )
Созреваниеяйцеклетки и ее оплодотворениепроисходит в маточных трубах. Яйцеклетка человека неможет самостоятельно передвигаться. Она имеетдиаметр до 130 мкм, окружена прозрачной (блестящей оболочкой)и слоем фолликулярных клеток. В яйцеклетке большое количество РНК, хорошо развита эндоплазматическаясеть. Небольшоеколичествожелтковых зерен достаточно яйцеклетке для питания в течение12-24 часовпосле овуляции, затем она погибает, или происходит оплодотворение и меняется источник питания.
воплодотворении различают три фазы.
1. Дистантное взаимодействие, в котором важную роль играютхимические вещества гиногамоны1 и II яйцеклетки и андрогомоны 1 и II спермиев. Гиногамоны1 активизируют двигательную активность снермиев, а андрогамоны1. напротив, подавляют. Гиногамоны II (фертилизины) вызывают склеиваниеспермиевпри взаимодействиис андрогамоном II, встроенным в цитолеммуспермия и предотвращают проникновениемногих сперматозоидов в яйцеклетку.
2. Контактное взаимодействие половыхклеток. Под влиянием сперматолизиновакросомы спермиев происходит слияние плазматическихмембран и плазмогамия — объединение цитоплазмы контактирующих гамет,
3. Третьяфаза — это проникновение в ооплазму (цитоплазму яйцеклетки) спермияс последующей кортикальной реакцией — уплотнением периферическойчасти ооплазмы иформированием оболочки оплодотворения.
Различают оплодотворение наружное (например, уамфибий) и внутреннее (у птиц, млекопитающих, человека), а также полиспермное,когда в яйцеклетку проникают несколько спермиев (например, у птиц) и моноспермное (у млекопитающих, человека).
Оплодотворение у человекавнутреннее, моноспермное. Оно происходит в ампулярной части маточной трубы. Яйцеклеткаокружается многочисленными спермиями.которые биением своих жгутиков заставляют вращаться яйцеклетку.Происходит капацитация — активация спермиевпод влиянием слизистого секрета железистых клеток яйцевода и акросомальнаяреакция выделениегиалуронидазы и трипсина из акросомы спермия. Онирасщепляют блестящую оболочку и контакты междуфолликулярными клетками, и спермий проникает в яйцеклетку. Сближаются ядра- пронуклеусы яйцеклетки и спермия,образуется синкарион. Далее пронуклеусысливаются и формируется зигота — новый одноклеточный организм, в которыйобъединялась материнская и отцовская наследственность.Пол ребенка определяется комбинацией половых хромосом в зиготе и зависит от половыххромосом отца. Аномальный кариотипприводит к патологии развития.
Дробление зиготы начинаетсяк концу первых сутокв яйцеводах по мере продвижения оплодотворенной яйцеклеткик матке и заканчивается в матке. Дробление зависит от типа яйцеклетки,от количества желтка и его распределения. Различают следующие типы дробления:
1. Полное, равномерное(у первично изолецитальных яйцеклетокланцетника, Полностью дробится зигота на равныечасти — бластомеры.
2. Полное, неравномерное(у мезолецитальныхяйцеклеток амфибий). Зигота дробитсяполностью, но бластомерыобразуются неодинаковые (мелкие на анимальном полюсе и крупные на вегетативном, где сосредоточен желток).
3. Частичное или меробластическое(у полилецитальных яйцеклеток птиц).Дробится лишь часть анимальногополюса яйцеклетки, свободного от желтка.
4.Полное ,неравномерное, асинхронное (у вторично изолецитальных яйцеклеток плацентарных млекопитающих и человека).
Дроблениехарактеризуется появлениемборозд дробления: меридианных широтных и тангенциальных, параллельныхповерхности дробления. Чем больше желтка содержитяйцеклетка, тем менее полно и равномерно происходит дробление. В результате дроблениязародыш становится многоклеточным — бластулой. Бластула имеет стенку — бластодерму, состоящуюиз клеток — бластомерови полость — бластоцель, заполненную жидкостью,продуктом секреции бластомеров.В бластодермеразличают крышу, образовавщуюсяза счет анимального полюса, дно — из материала вегетативного полюса и краевуюзону, расположенную междуними. У ланцетника при полном равномерном дроблении образуется шарообразная бластула — с однослойной бластодермой(только меридианные и широтные борозды) и с центральнорасноложеннымбластоцелем — целобластула. У лягушек в результате полного неравномерногодробления (все три типа борозд дробления) образуется бластула смногослойной бластодермой эксцентрично расположеннымбластоцелем — это амфибластула. У птиц и пресмыкающихсяс резко телолецитальными яйцеклетками дробитсялишь часть анимального полюса, свободного от желтка,и образуется дискобластула с щелевидным бластоцелем между бластомерами в областианимального полюсаи нераздробленным желтком. Умлекопитающих и человека с вторично изолецитальными яйцеклетками дроблениеполное (дробится без остатка вся зигота), асинхронное (количествобластомеровнарастает в неправильном и особом порядке уразных животных (у человека 2, 3, 4, 5, 7), неравномерное(образуется два типа бластомеров).Одни бластомерытемные, крупные, медленнодробящиеся — это эмбриобласт.Из него образуется тело зародыша и все внезародышевые органы, кроме трофобласта.Второй тип бластомеровпредставлен мелкими, светлыми, быстроделящимися клетками — это трофобласт, связывающийзародыш с организмом матери и обеспечивающийего трофику. Светлые бластомеры обрастают кучкутемных бластомеров и дробящийся зародыш приобретает вид плотногошара — морулы через50-60 часов, На третьи сутки начинаетсяформирование бластоцисты — полого пузырька, образованного снаружи трофобластом и заполненного жидкостью, с эмбриобластом в видеузелка клеток, прикрепленнымизнутри к трофобластуна одном полюсе бластоцисты. Бластоциста поступает вматку на 5 сутки и свободно в ней раполагается. Происходитподготовка к имплантации. В трофобластестановится больше лизосом, у трофобластапоявляются выросты. Зародышевый узелок, уплощаясь, преобразуетсяв зародышевый щиток, подготавливаясь к первойфазе гаструляции
С седьмыхсуток начинается имплантация — внедрение бластоцисты в стенку матки,при котором зародыш полностьюпогружается в слизистую оболочку матки, аслизистая оболочка срастается над зародышем (интерстициальнаяимплантация). В имплантации различают двестадии: адгезия(прилипание) и инвазия (проникновение). На образующихсяворсинках-выростах трофобласта формируются два слоя: цитотрофобласт — внутренний и наружный — симпластотрофобласт, продуцирующий протеолитические ферменты, подплавляющиеслизистую матки. Так в матке появляется имплантационнаяямка, куда проникает бластоциста.Гистиотрофный типпитания за счет потребления продуктов распада материнских тканей в первые две недели сменяется на гематрофный тип — непосредственно изматеринской крови. Имплантация являетсякритическим периодом в эмбриогенезе человека.
Гаструляциятакже являетсякритическим периодом в развитии. Она приводит к образованию многослойногозародыша (гаструла), Способыобразования гаструлы различны:
1. Инвагинация-впячивание(уланцетника).
2. Эпиболия-обрастание(у амфибий эпиболия идет совместно счастичной инвагинацией).
3. Деляминация — расщепление(у птиц, млекопитающих, человека).
4. Иммиграция — выселение, перемещение(у птиц,млекопитающих, человека).
У человека гаструляция протекает вдве фазы: первая (7-е сутки) — путем деляминации эмбриобласта образуютсядва листка: наружный — эпибласт ивнутренний — гипобласт.Вторая стадия (14-15 сутки) происходит как и у птиц с образованием первичнойполоски и первичного узелка путем перемещения, иммиграции клеточных масс, что в итогеприводит к формированию мезодермы и хорды. Междудвумя стадиями гаструляцииобразуются внезародышевые органы: амниотический, желточный пузырьки и хорион,обеспечивающие условия для развития зародыша исоставляющие одну из особенностей развитиячеловека. У семисуточного зародыша из зародышевого щитка выселяются отростчатые клетки — (внезародышевая мезодерма),которая участвует в образовании амниона вместе с эктодермой, желточного мешка вместе с энтодермой и хориона вместе с трофобластомна второй неделе развития человека. К IIсуткам внезародышевая мезодерма заполняет полость бластоцисты,подрастает к трофобласту,формируя хорион. В выросты трофобластаврастает внезародышеваямезодерма, а позднее прорастают и кровеносные сосуды — так образуются ворсинкихориона. Последние приконтакте с эндометрием матки будут формироватьплаценту. На 13-14сутки у эмбриона человека — два листка: эпибласт (первичная эктодерма) и гипобласт(первичная энтодерма), и два пузырька — амниотический и желточный.Дно амниотическогопузырька (эпибласт)и крыша желточного (гипобласт) образуют вместе зародышевый щиток. Тяж внезародышевой мезодермы амниотическая или зародышевая ножкаприкрепляет к хориону два пузырька: амниотическийи желточный
После второйстадии гаструляциина 15-17 сутки в амниотическуюножку врастает пальцевидный вырост из заднего отдела кишечной трубки — аллантоис, по которому растут сосуды к хориону. У 17-ти суточного эмбриона уже сформированы тризародышевых листка, внезародышевые органы, ипроисходит дифференцировказародышевых листков и закладка осевых основных зачатков органов.
ДИФФЕРЕНЦИРОВКАЗАРОДЫШЕВЫХ ЛИСТКОВ.
Дифференцировка — это изменения в структуре клеток, связанные со специализацией их функций и обусловленные активностьюопределенных генов. Различают 4 этапа дифференцировки:
1. Оотипическаядифференцировка на стадии зиготы представленапредположительными,презумптивнымизачатками - участками оплодотворенной яйцеклетки.
2. Бластомернаядифференцировка на стадиибластулы заключается в появлении неодинаковых бластомеров (например, бластомерыкрыши, дна краевых зон у некоторых животных).
3. Зачатковаядифференцировка на стадииранней гаструлыВозникают обособленные участки — зародышевые листки.
4. Гистогенетическаядифференцировка на стадии поздней гаструлы. В пределах одного листка появляются зачатки различныхтканей (например, в сомитах мезодермы). Из тканейформируются зачатки органов и систем. В процессегаструляции, дифференцировкизародышевых листков появляются осевой комплексзачатков органов.
Зародышевыелистки дифференцируются у большинства позвоночных одинаково, Приэтом каждый листок дифференцируется в определенномнаправлении. Из первичнойэктодермы образуется нервная трубка, ганглиозные пластинки, плакоды, кожная эктодерма,прехордальная пластинка и внезародышевая эктодерма. Первичная энтодерма является источникомзародышевой кишечной энтодермы и внезародышевой(желточной). При дифференцировкемезодермы возникают три части: в дорсальном отделе появляются (1) сомиты, за ним следуют (2) сегментные ножки (нефротомы), из которых образуется эпителий почек игонад. Вентральная мезодерма не сегментируетсяи формирует (3) спланхнотом расщепляющийсяна два листка: париетальный, сопровождающий эктодерму,и висцеральный, прилежащий к энтодерме. Между листками возникает целомическая полость, из листков спланхнотомаобразуется эпителий серозных оболочек — брюшины. плевры,перикарда. Далее в теле сомитадифференцируется из наружной его части дерматом- (источник дермы кожи),из центральной — миотом(зачаток скелетной мышечной ткани) и из внутренней склеротом(зачаток скелетных соединительных тканей — костей и хрящей). В процессе дифференцировкизародышевых листков мезодермы у зародыша появляетсямезенхима.
На 20-21сутки у эмбриона человека образуются туловищные складки, обособляющие тело зародыша человека от внезародышевыхорганов и окончательно формируются осевые зачатки органов: хорда, из эктодермы- нервная трубка, замыкающаяся к 25 суткам. Формируется кишечная трубка.Мезодерма зародыша дифференцируется на сомиты (сомитный период), нефротом и спланхнотом с париетальными висцеральными листками. В теле сомита различают: дерматом, миотом и склеротом. В период дифференцировкимезодермы изо всех трех зародышевых листков, но преимущественно из мезодермы, появляются мезенхима зародыша — отростчатыеклетки, эмбриональный зачаток многих тканей и органов всех видов соединительнойткани (отсюда ее часто называют эмбриональной соединительной тканью), а также гладкомышечной ткани, микрооглии сосудов, крови, лимфы,кроветворных органов. Ко второму месяцу у эмбриона человека произошел начальныйгисто — и органогенези имеются закладки почти всех органов, К концу 8-й недели эмбриогенеза заканчиваетсязародышевый период развития и начинается плодный.
Ранние стадии развития человека имеют ряд особенностей:1. Асинхронный тип полного неравномерногодробления с образованием “темных" и “светлых” бластомеров; 2. Интерстициальныйтип имплантации. 3. Наличие двух фаз гаструляции — деляминациии иммиграции, между которыми бурно развиваются внезародышевыеорганы; 4. Раннее обособление и формирование внезародышевых органов; 5. Раннее образование амниотического пузырька без амниотических складок; 6.Сильное развитие амниона, хориона и слабое-желточного меткаи аллантоиса.
Внезародышевые органы (провизорные,временные или зародышевые оболочки), обеспечивающиеразвитие зародыша. В эволюции появляются впервые у рыб (желточный мешок). У птиц имеются следующиевнезародышевые органы: амнион, сероза, желточный мешок и аллантоис. Амнион — воднаяоболочка, серозная — орган дыхания. Образуютсяэти две оболочки у птицпутем смыкания амниотических складок. Желточный мешок выполняет уптиц трофическую и кроветворную функции, а аллантоис -орган выделения и газообмена у птиц.
Вэмбриогенезе человека образуется пять внезародышевых органов: амнион, желточный мешок, хорион,формирующий плаценту и аллантоис.Амнион, создающийводную среду у человека, образуется без амниотических складок. Желточный мешок у человекапрактически утрачиваеттрофическую и выполняет в основномкроветворную функцию и образования первичных половых клеток. Аллантоис.редуцирующийся на втором месяце является проводником кровеносных сосудов к хориону.Хорошо развитый хорион у человекаформирует плаценту, за счет которой устанавливается связь зародыша и матери.
Плацента,обеспечивающая связь зародыша с организмомматери, выполняет многочисленные функции: трофическую, дыхательную, выделительную, эндокринную,защитную, депонирующую. По морфологическим признакамразличают четыре типа плаценты: эпителиохориальные, десмохориальные,эндотелиохориальныеи гемохориальные. Эпителиохориальные диффузные плаценты(у дельфинов, свиней, лошадей) характеризуются врастанием ворсинок хориона вматочные железы. В десмохориальныхмножественныхплацентах (у корой,овец) ворсинки хориона, разрушая эпителии маточных желез, врастаютв подлежащую соединительнуюткань эндометрия матки. Эндотелиохориальный поясной тип плаценты характерен для хищников(кошки, волки, куницы, лисы), Хориальныеворсинки у подобного типа плацент разрушают эпителий,соединительную ткань и контактируют сэндотелием сосудов эндометрия матки. Гемохориальныйтип плаценты (например, у летучих мышей, приматов, человека)характеризуется разрушением стенок сосудов эндометрия матки ворсинками хориона инепосредственным контактом их с материнской кровью. С рождением, новорожденные,имеющие плацентыпервых двух типов, способны к самостоятельному питанию ипередвижению. тогда как новорожденные с двумя последними типами плацент после рождения долгое время не способнысамостоятельно питаться.
Плацента человека гемохориальнаядискоидальнаяворсинчатая плацента выполняет многочисленныефункции, обеспечивающие рост и развитие эмбриона за счет организма матери. В плаценте выделяют две части: зародышевую или плодную(детскую) и материнскую или маточную. Плодная часть образуется ветвистымхорионом, покрытым амниотической оболочкой, а материнская базальной пластинкой — видоизмененной базальной частью эндометрия. Развитие плаценты происходит параллельно началу формирования зачатковорганов: с 3 по 6 недели (критический период в эмбриогенезе человека) изаканчивается в конце 3-го месяца беременности. К этому времени плодная часть плаценты состоит из плотной соединительно-тканной хориальной пластинки с отходящими от нееветвящимися ворсинками хориона, погруженными в лакуны с материнской кровью. Хориальная пластинкасверху покрыта частью амниотическойоболочки.
После оплодотворения слизистую оболочку матки называют децидуальной, отпадающей и вней выделяют 3 части: основную отпадающую, где произошла имплантация между эмбрионом и мышечной оболочкой матки: вторую часть — сумочную отпадающую. отделяющую зародыш от полостиматки и третьючасть — пристеночнуюотпадающую, остальную часть децидуальнойоболочки. Ворсинки хориона, обращенные к основной отпадающей, сильноразрастаются и ветвятся — это ветвистый(пышный хорион). В этой области и формируется плацента: за счет ветвистогохориона — ее плодная часть, а за счет основной отпадающей -- ее материнскаячасть. В области пристеночнойи сумочной отпадающих ворсинки хориона вдальнейшем вообще исчезают (гладкий хорион). Хориальные ворсинкисостоят из эмбриональной волокнистой соединительно-тканной стромы с сосудами. Клеточный и волокнистыйсостав этой соединительной ткани, вязкостьосновного вещества (содержание гиалуроновойи хондроитин — серной кислоты, с которыми связана регуляция проницаемости ворсинокплаценты) изменяется со сроком беременности. С поверхности, соединительно-тканнаястрома ворсинок наранних сроках беременности покрыта трофобластическимэпителием, имеющим клеточное строение. Он представлен однослойнымэпителием — цитотрофобластом, постепенноредуцирующимся со второго месяца эмбриогенеза. На поверхности цитотрофобластапоявляется наружныйслой — синцитиотрофобласт — многоядерная структура с большим количеством протеолитических и окислительных ферментов. Вконце беременности синцитиотрофобласт также подвергаетсяраспаду и местами на поверхности ворсинок возникает фибриноподобная оксифильная масса (фибриноид Лангханса).
Материнская частьплаценты представлена базальной пластинкой (глубокие неразрушенныечасти отпадающей оболочки вместе с трофобластом), соединительно-тканными септами, отходящимиот базальной пластинки исрастающиеся с ворсинкамихориона. Это так называемые якорные или стволовыеворсинки делят плаценту на дольки-котиледоны. Также в материнской части плаценты имеются лакуны с материнскойкровью и ворсинками хориона (конечные разветвления стволовых ворсинок). Базальный слой эндометрия — глубокий слойслизистойоболочки матки содержит в своей соединительной ткани крупные децидуальные клетки с оксифильнойцитоплазмой, богатой включениямигликогена, округлыми ядрами, и четкими клеточными границами. В базальной пластинке в области прикрепленияякорных ворсинок нередко имеются скопления базофильных клеток периферического цитотрофобласта.На поверхности базальнойпластинки, обращенной к ворсинкам иногдаформируется аморфная оксифильная субстанция (фибриноид Рора), которая вместе с трофобластическимиклеткамибазальной пластинки обеспечивает иммунологический гомеостазсистемы мать — плод. Часть основной отпадающейоболочки по краю плацентарного диска награнице гладкого и ветвистого хориона плотно прирастает к хориону и не разрушается, образуя замыкательную пластинку, препятствующуюистечению крови из лакун.
Кровь материи плода,циркулируя по самостоятельным системам никогда не смешивается благодаря наличию гемоплацентарного (гомохориального) барьера, разделяющего кровотокплода от кровотока матери.Гемоплацентарный барьер состоит из эндотелия с базальноймембраной сосудов плода, окружающей эти сосудысоединительно-тканной стромы хориальных ворсин и их эпителия (цитотрофобласт, синцитиотрофобласт)и фибриноида. Эмбрион выделяет в кровь матери углекислый газ и продукты обмена и получает из крови матери кислород, воду, питательныевещества, витамины, гормоны, иммуноглобулины,а также лекарственные вещества, алкоголь, никотин, вирусы.
Пупочный канатикразвивается восновном из мезенхимы амниотической ножки и представляет собой упругое соединительно-тканноеобразование с сосудами, а также с остатками желточного стебелька и аллантоиса, снаружи покрытое амниотической оболочкой. В его студенистой, слизистой соединительно-тканной основе(вартониев студень)проходят пупочныеартерии и пупочная вена, обеспечивающие обменные процессыэмбриона.
Система мать — плод, развивающаясяпри беременности,состоит из организма матери и плода, связанныхмежду собой плацентой. Основными механизмами,обеспечивающими взаимодействие в системе мать - плод являются нейрогуморальныемеханизмы матери и плода: рецепторные, регуляторные,исполнительные. Эти механизмы направлены на созданиеоптимальных условий для развития плода. Приэтом особо важная роль принадлежит плаценте,аккумулирующей и синтезирующейвещества, гормоны, необходимые для развития плода,и осуществляющей гуморальные и нервные связи между плодом иматерью. Гуморальные связи осуществляются нетолько через плаценту,
www.ronl.ru
Как и многие другие естественные науки, эмбриология зародилась в античности. В трудах Аристотеля имеются довольно подробные описания развития куриного зародыша. В это же время возникли и две основные точки зрения на процессы развития - преформизм и эпигенез . Эти два взгляда на развитие полностью сформировались к XVII в., и между ними началась борьба. Тогда в связи с появлением микроскопа стали накапливаться фактические данные о строении зародышей и процессах развития разных организмов.
Голландец Ян Сваммердам изучил развитие и метаморфоз насекомых. Его соотечественник Антони ван Левенгук открыл партеногенез у тлей и наблюдал сперматозоиды человека. Итальянец Марчелло Мальпиги переисследовал развитие куриного эмбриона и изучил микроскопическую анатомию многих органов и тканей животных и растений. Большинство сделанных наблюдений истолковывались их авторами с позиций преформизма (предсуществования). Согласно этой точке зрения, в процессе развития не возникает ничего действительно нового, все части зародыша в уже сформированном виде присутствуют в яйце, и если их нельзя увидеть, то только из-за малых размеров; в дальнейшем происходит лишь их рост. Так, Сваммердам считал, что внутри яйца бабочки спрятана гусеница, внутри гусеницы - куколка, а внутри куколки - взрослая бабочка. Отметим, что с точки зрения преформистов казалось маловероятным самозарождение организмов. Ясно, что если зародыш предсуществует, он может находится только либо в яйцеклетке, либо в сперматозоиде. Поэтому преформисты разделились на два лагеря - овистов (от лат. ovum - яйцо) и анималькулистов (от лат. animal-culum - "зверек", одно из названий сперматозоида). При этом оказывалось совершенно непонятным, как же передаются потомкам признаки второго родителя.
Доведенные до логического завершения взгляды преформистов привели к появлению "теории вложения", одним из авторов которой был математик Г.Лейбниц . Если в яйцеклетке есть полностью сформированный зародыш, то в его яичниках должны быть сформированные яйцеклетки, в них - зародыши следующего поколения, и так до бесконечности. С точки зрения современной науки эти взгляды, конечно, совершенно нелепы.
Согласно точки зрения, получившей название эпигенеза, в ходе развития происходит постепенное усложнение формы; из бесструктурного, однородного материала яйца возникают различающиеся части зародыша. Автором термина "эпигенез" был У.Гарвей , открывший круги кровообращения и выдвинувший в своей книге "О произрождении животных" знаменитый принцип "Все (живое) из яйца". Гарвей писал: "Ни одна часть плода не существует в яйце актуально, но все части находятся в нем потенциально". Яйцо не содержит органов зародыша, а лишь "потенции" - задатки их развития.
В XVIII в. продолжал господствовать преформизм. Однако в 1759 г. в работе малоизвестного тогда ученого Каспара Фридриха Вольфа появились факты, прямо противоречащие преформистским воззрениям. Вольф доказывал, что такие органы куриного зародыша, как нервная трубка и кишечник, первоначально закладываются в виде плоских пластов, а лишь затем сворачиваются в трубки. Таким образом, в ходе развития происходит формообразование. Однако даже эти четкие наблюдения отвергались преформистами.
В первой половине XIX в. факты, противоречащие преформизму, уже стали общепризнанными. Так, было твердо установлено, что все органы позвоночных развиваются из трех первичных зародышевых пластов, или листков (см. " Органогенез ").
Согласно современной точке зрения, истина лежит где-то посередине между преформизмом и эпигенезом. Высказывание Гарвея оказалось гениальным предвидением. Генетическая программа развития, определяющая его ход, полностью "преформирована" в оплодотворенной яйцеклетке. Кроме того, и цитоплазма яйца не бесструктурна - в разных ее участках содержатся вещества, предопределяющие ход развития частей зародыша (см. " Оплодотворение "), но, конечно, никаких структур будущего зародыша в яйце нет.
В первой половине XIX в. большие успехи делали систематика и сравнительная анатомия. Одним из главных методов в биологии стал сравнительный метод. Под влиянием этих успехов зародилась сравнительная эмбриология. Наиболее важные результаты этого направления эмбриологии были получены российским ученым Карлом Бэром . Изучив эмбриональное развитие всех классов позвоночных, Бэр установил, что на некоторой довольно ранней стадии все эти зародыши наиболее сходны между собой, а различия приобретаются в ходе дальнейшего развития. Это позволило Бэру сформулировать закон зародышевого сходства: зародыши животных, относящихся к одному типу, более сходны, чем взрослые формы. Признаки типа появляются в ходе развития раньше, чем признаки класса, отряда и т.д.; последними появляются видовые признаки.
Новый импульс это направление исследований получило после выхода в свет книги Ч.Дарвина "О происхождении видов" и быстрого распространения идей эволюционизма. Зародышевое сходство стали рассматривать как показатель общности происхождения, а изучение эмбрионального развития - как один из главных способов изучения филогении (путей эволюционного развития) организмов. Э.Геккель сформулировал биогенетический закон : "Онтогенез есть быстрое и краткое повторение филогенеза (исторического развития вида)". Геккель утверждал, что филогенез есть причина онтогенеза: идивидуальное развитие полностью обусловлено историей развития вида. В дальнейшем эти взгляды были частично отвергнуты наукой, а частично видоизменены и дополнены.
Особенно большой вклад в эволюционную эмбриологию внесли отечественные ученые А.О.Ковалевский и И.И.Мечников . Ковалевский изучил раннее развитие ланцетника - представителя хордовых - и доказал его сходство с развитием асцидий и некоторых других беспозвоночных. Мечников изучил раннее развитие примитивных многоклеточных ( губок и кишечнополостных ) и доказал сходство ранних стадий развитая всех многоклеточных животных.
Увлечение восстановлением хода эволюции на основе эмбриологических данных привело к тому, что эмбриология во многом утратила самостоятельность, став частью сравнительной анатомии и теории эволюции. Однако в конце XIX в. вновь усилился интерес к непосредственным причинам развития, его механизмам. Новому направлению одним из его основателей, Вильгельмом Ру , было дано название механики развития. В основу этого подхода Ру положил причинно-аналитический метод, согласно которому, чтобы понять механизмы развития, надо разбить его на отдельные звенья, выяснить механизмы каждого звена и причинно-следственные связи между ними. Ру подчеркивал, что сделать это можно только путем экспериментов. Например, следовало выяснить, может ли каждая часть зародыша развиваться самостоятельно или требует воздействий извне. Для этого можно изолировать часть зародыша, пересадить ее в другой участок и т.д. Он утверждал также, что процессы развития нельзя объяснить без знания законов химии и физики. Эти законы лежат в основе функционирования живых клеток, а развитие, в свою очередь, складывается из небольшого числа разных форм клеточной активности - роста, деления, миграции и т. д.
medbiol.ru
Эмбриология человекаЭмбриология человека, изучение развития человеческого организма от момента образования одноклеточной зиготы, или оплодотворенного яйцеклетки, до рождения ребенка. Эмбриональное (внутриутробное) развитие человека длится примерно 265–270 дней. В течение этого времени из исходной одной клетки образуется более 200 миллионов клеток, а размеры эмбриона увеличивается от микроскопического до полуметрового. В целом развитие человеческого эмбриона можно разделить на три стадии. Первая – это период от оплодотворения яйцеклетки до конца второй недели внутриутробной жизни, когда развивающийся эмбрион (зародыш) внедряется в стенку матки и начинает получать питание от матери. Вторая стадия длится с третьей до конца восьмой недели. В течение этого времени формируются все основные органы и эмбрион приобретает черты человеческого организма. По окончании второй стадии развития он уже называется плодом. Протяженность третьей стадии, называемой иногда фетальной (от лат. fetus – плод), – от третьего месяца до рождения. На этой заключительной стадии завершается специализация систем органов и плод постепенно приобретает способность существовать самостоятельно. Половые клетки и оплодотворение У человека зрелая половая клетка (гамета) – это сперматозоид у мужчины, яйцеклетка (яйцо) у женщины. Перед слиянием гамет с образованием зиготы эти половые клетки должны сформироваться, созреть и затем встретиться. Половые клетки человека по структуре сходны с гаметами большинства животных. Принципиальное отличие гамет от остальных клеток организма, называемых соматическими, заключается в том, что гамета содержит только половину от числа хромосом соматической клетки. В половых клетках человека их 23. В процессе оплодотворения каждая половая клетка привносит в зиготу свои 23 хромосомы, и таким образом зигота имеет 46 хромосом, т.е. двойной их набор, как это присуще всем соматическим клеткам человека. Будучи сходны по главным структурным признакам с соматическими клетками, сперматозоид и яйцеклетка в то же время высоко специализированы для своей роли в репродукции. Сперматозоид – небольшая и очень подвижная клетка . Яйцеклетка, напротив, неподвижна и гораздо крупнее (почти в 100 000 раз), чем сперматозоид. Бльшую часть ее объема составляет цитоплазма, содержащая запасы питательных веществ, необходимые эмбриону в начальный период развития . Для оплодотворения необходимо, чтобы яйцеклетка и сперматозоид достигли стадии зрелости. Более того, яйцеклетка должна быть оплодотворена в течение 12 часов после выхода из яичника, в противном случае она погибает. Человеческий сперматозоид живет дольше, около суток. Быстро двигаясь с помощью своего кнутообразного хвоста, сперматозоид достигает соединенного с маткой протока – маточной (фаллопиевой) трубы, куда попадает из яичника и яйцеклетка. Обычно это занимает менее часа после совокупления. Считается, что оплодотворение происходит в верхней трети маточной трубы. Несмотря на то, что в норме эякулят содержит миллионы сперматозоидов, только один проникает в яйцеклетку, активируя цепочку процессов, приводящих к развитию эмбриона. В силу того, что сперматозоид весь целиком проникает в яйцеклетку, мужчина привносит потомку, помимо ядерного, и некоторое количество цитоплазматического материала, в том числе центросому – небольшую структуру, необходимую для клеточного деления зиготы. Сперматозоид определяет и пол потомка. Кульминацией оплодотворения считается момент слияния ядра сперматозоида с ядром яйцеклетки. Дробление и имплантация После оплодотворения зигота постепенно спускается по маточной трубе в полость матки. В этот период, в течение примерно трех дней, зигота проходит стадию клеточного деления, известную как дробление. При дроблении число клеток увеличивается, но общий их объем не меняется, так как каждая дочерняя клетка мельче, чем исходная. Первое дробление происходит примерно через 30 часов после оплодотворения и дает две совершенно одинаковые дочерние клетки. Второе дробление наступает через 10 часов после первого и приводит к образованию четырехклеточной стадии. Примерно через 50–60 часов после оплодотворения достигается стадия т.н. морулы – шара из 16 и более клеток. По мере продолжения дробления наружные клетки морулы делятся быстрее, чем внутренние, в результате наружный клеточный слой (трофобласт) отделяется от внутреннего скопления клеток (т.н. внутренней клеточной массы), сохраняя с ними связь только в одном месте. Между слоями образуется полость, бластоцель, которая постепенно заполняется жидкостью. На этой стадии, наступающей через три–четыре дня после оплодотворения, дробление заканчивается и эмбрион называют бластоцистой, или бластулой. В течение первых дней развития, эмбрион получает питание и кислород из секрета (выделений) маточной трубы. Примерно через пять–шесть дней после оплодотворения, когда бластула находится уже в матке, трофобласт образует пальцевидные ворсинки, которые, энергично двигаясь, начинают внедряться в ткань матки. В то же время, по-видимому, бластула стимулирует выработку ферментов, способствующих частичному перевариванию слизистой (эндометрия) матки. Примерно на 9–10 день эмбрион имплантируется (врастает) в стенку матки и оказывается полностью окруженным ее клетками; с имплантацией эмбриона прекращается менструальный цикл. В дополнение к своей роли в имплантации, трофобласт участвует также в образовании хориона – первичной мембраны, окружающей эмбрион. В свою очередь хорион содействует образованию плаценты, губчатой по структуре мембраны, через которую эмбрион в дальнейшем получает питание и выводит продукты обмена. Эмбриональные зародышевые листки Эмбрион развивается из внутренней клеточной массы бластулы. По мере увеличения давления жидкости внутри бластоцеля клетки внутренней клеточной массы, которая становится компактной, формируют зародышевый щиток, или бластодерму. Зародышевый щиток разделяется на два слоя. Один из них становится источником трех первичных зародышевых листков: эктодермы, энтодермы и мезодермы. Процесс обособления сначала двух, а затем и третьего зародышевого листка (т.н. гаструляция) знаменует превращение бластулы в гаструлу. Зародышевые листки вначале различаются лишь по расположению: эктодерма – самый наружный слой, энтодерма – внутренний, а мезодерма – промежуточный. Формирование трех зародышевых листков завершается примерно через неделю после оплодотворения. Постепенно, шаг за шагом, каждый зародышевый листок дает начало определенным тканям и органам. Так, эктодерма формирует наружный слой кожи и ее производные (придатки) – волосы, ногти, кожные железы, выстилку ротовой полости, носа и заднего прохода, – а также всю нервную систему и рецепторы органов чувств, например сетчатку глаза. Из энтодермы образуются: легкие; выстилка (слизистая оболочка) всего пищеварительного тракта, кроме рта и заднего прохода; некоторые примыкающие к этому тракту органы и железы, такие, как печень, поджелудочная железа, тимус, щитовидная и паращитовидные железы; выстилка мочевого пузыря и мочеиспускательного канала. Мезодерма – источник системы кровообращения, выделительной, половой, кроветворной и иммунной систем, а также мышечной ткани, всех типов опорно-трофических тканей (скелетной, хрящевой, рыхлой соединительной и т.д.) и внутренних слоев кожи (дермы). Полностью развившиеся органы обычно состоят из нескольких типов тканей и поэтому связаны своим происхождением с разными зародышевыми листками. По этой причине проследить участие того или иного зародышевого листка можно только в процессе формирования ткани. Внезародышевые оболочки Развитие эмбриона сопровождается образованием нескольких оболочек, окружающих его и отторгаемых при рождении. Самая наружная из них – уже упоминавшийся хорион, производное трофобласта. Он соединен с эмбрионом с помощью телесного стебелька из соединительной ткани, происходящей из мезодермы. Со временем стебелек удлиняется и образует пупочный канатик (пуповину), соединяющий эмбрион с плацентой. Плацента развивается как специализированный вырост плодных оболочек. Ворсинки хориона прободают эндотелий кровеносных сосудов слизистой оболочки матки и погружаются в кровяные лакуны, заполненные кровью матери. Таким образом, кровь плода отделена от крови матери лишь тонкой наружной оболочкой хориона и стенками капилляров самого зародыша, т.е. непосредственного смешения кр...
www.tnu.in.ua
Лекция № 1: Предмет, история развития и методы исследования в эмбриологии.
Название эмбриология происходит от греческих слов эмбрион - зародыш и логос - учение. Это название не соответствует содержанию современной науки. Эмбриология действительно описывает и выясняет всех процессов зародышевого развития - от оплодотворения яйца сперматозоидом до вылупления зародыша из яйцевых оболочек у яйцекладущих животных или до выхода его из материнского организма у живородящих. Однако эмбриология изучает и предзародышевый период - формирование половых клеток. Эмбриология изучает и так называемый постэмбриональный период. У млекопитающих животных некоторые системы органов (например, половая система, эндокринные железы) приобретают дефинитивные, т.е. окончательные, свойственные взрослому состоянию, структуры и функции спустя тот или иной промежуток времени после рождения. Зародыши многих животных, освобождаясь от яйцевых оболочек, имеют строение, мало сходное со строением взрослых организмов; у них развиваются провизорные (временные) органы, обеспечивающие их самостоятельное существование. Такие зародыши и личинки живут в совершенно отличной от имаго внешней среде и имеют специальные приспособления к этой среде. Впоследствии совершается метаморфоз, в ходе которого личиночные органы преобразуются, прежде чем достигнут своего дефинитивного состояния.
Таким образом, эмбриология занимается изучением индивидуального развития организма. Предметом ее исследований являются и регенерация, и бесполое размножение. Эмбриология изучает и патологические явления - причины нарушения нормального эмбрионального развития, возникновения уродств, причины нарушения нормальных процессов развития и жизни тканей и органов, Некоторые эмбриологические школы исследуют в своем аспекте причины возникновения опухолей.
Краткая история эмбриологии
Зачатки эмбриологических знаний о млекопитающих и птицах были уже в древнем Египте, Вавилонии, Ассирии, Индии и Китае.
Первые регулярные знания в области эмбриологии связывают с именем Гиппократа (460 - 370 до н.э.). Гиппократ предвосхитил идею преформации: «Все части зародыша образуются в одно и то же время. Согласно этой теории, каждый зародыш является уже вполне сформированным, имеющим все части организма, которому остается только расти. Выражаясь современным языком, в зародыше преобразованы и преформированы все особенности будущего организма, происходит лишь рост без дифференциации. Наиболее крайние преформисты представляли себе, что каждый организм, в том числе и человеческий, заключает в себе громадное количество поколений зародышей, вложенных друг в друга в теле прародителей со времени сотворения мира. Эта идея
господствовало в течение 17 - 18 веков - теория преформизма.
Рис. 1. Гомункулус – миниатюрный индивидуум, расположенный в головке сперматозоида, который использует яйцеклетку для питания в процессе своего роста.
Между представителями двух направлений преформистов велись горячие споры. Анималькулисты, или сперматики, например А.В. Левенгук, описывал “семенных зверушек” (сперматозоидов) использующих запасы питательных веществ яйцеклетки для своего роста. Овисты думали, что зародыши в миниатюрном виде находятся не в мужском семени, а в яйцеклетке, и семя, входящее в яйцо при оплодотворении составляет питательный материал зародыша. Сторонниками преформизма были выдающиеся ученые ХVII и ХVIII вв. А. Левенгук, Я. Сваммердам, М. Мальпиги, А. Галлер, Ш. Бонне.
В 1V в. до н.э. работал другой величайший ученый античной древности -Аристотель (384 - 322 г.г. до н.э.). Аристотель сформулировал впервые теорию эпигенеза, значительно более соответствующую современной эмбриологии, Он, однако, внес в нее идеалистическое содержание.
Важнейшей вехой в истории эмбриологии считается 1759 г. В этот год была опубликована диссертация «Теория развития» двадцатишестилетнего Каспара Фридриха Вольфа, ставшего впоследствии академиком Петербургской Академии наук. В диссертации Фридрих Вольф впервые попытался объяснить возникновение новых растительных клеток при росте. Считал, что из уже имеющихся клеток-мешочков выдавливается жидкое вещество в виде капельки, поверхность капли затвердевает и капля превращается в новую клетку.
Вольф обосновал эпигенез, проследил развитие зародыша курицы, опровергнув преформизм. Большая заслуга Вольфа состояла в том, что он показал всю несостоятельность и абсурдность представлений преформистов о наличии в половой клетке готового организма, показав, что органы в эмбриогенезе возникают заново.
Весь XVI11 век прошел под знаком борьбы двух теорий развития. Явное торжество преформистских представлений тормозило разработку того прогрессивного начало, которое было заложено в теории эпигенеза. Накапливаемый фактический материал не получил должного признания: слишком низким был теоретический уровень науки.
Описание полного индивидуального развития - онтогенеза организма, начиная с яйцеклетки, впервые дал Карл Бэр (1792 - 1876). Он продолжил работу Вольфа на цыпленке и на основании полученных им фактов подтвердил некоторые выводы своего предшественника.
Исследования Бэра привели его к выводу, что развитие состоит в постепенном усложнении более простых структур.
Большая заслуга Бэра - открытие им яйца млекопитающих и человека. До него за яйцеклетку принимали так называемый графов пузырек - довольно значительных размеров образование, наполненное жидкостью, в стенке которого и находится яйцо.
Сравнивая развитие некоторых позвоночных, Бэр обратил внимание на то, что их зародыши обнаруживают между собой большее сходство, чем взрослые животные. При этом он отметил, что, чем моложе сравниваемые эмбриональные стадии, тем значительнее сходство. Закономерность, открытая Бэром, известна как явление зародышевого сходства.
Своим возникновением и развитием современная эволюционная эмбриология связана работами великих русских ученых А.О. Ковалевского (1840 - 1901) и И.И. Мечникова (1845- 1916).
Труды Ковалевского имели решающее значение для установления родственных связей между некоторыми группами животных. В этом отношении особенно большую роль сыграли его работы по изучению ланцетника и оболочников. Исследовав ранние стадии развития этих животных, А.О. Ковалевский доказал родство их с позвоночными и принадлежность к одному типу хордовых. Факты, добытые ученым, впервые наметили прямую связь между беспозвоночными, разделенными до того, казалось, непреодолимой пропастью.
Изучая эмбриональные стадии позвоночных и особенно мало изученных беспозвоночных, И.И. Мечников и А.О. Ковалевский показали, что развитие почти всех многоклеточных происходит через стадию образования трех зародышевых листков. Последние у животных сходны не только по способу происхождения, но также и по тем производным, которые дает каждый из них.
В четкой форме вопрос о связи эмбрионального развития с эволюцией впервые поставил Ф. Мюллер. Он сделал вывод о том, что в эмбриональном развитии наблюдается краткое повторение длительного исторического развития. Эта мысль была целиком принята Э. Геккелем и, подтвержденная новыми данными, получила более широкое обобщение в основном биогенетическом законе. Этот закон в наиболее общей формулировке гласит, что в своем индивидуальном развитии (онтогенезе) организм повторяет в краткой, сжатой форме историю своего вида (филогенез).
Экспериментальная эмбриология
Вильгельму Ру принадлежит честь основания экспериментального направления в эмбриологии, Он разрушал раскаленной иглой один из первых двух бластомеров лягушки. Из оставшегося бластомера развивалась половина зародыша. Такое же частичное развитие было обнаружено и в опытах на дробящихся яйцах некоторых других животных. Дефектные зародыши наблюдались при изоляции бластомеров асцидии, маллюсков, лошадиной аскариды, гребневиков и др.
Нарушения в развитии при изоляции бластомеров или даже отдельных частей яйца Ру объяснял предопределением в яйце частей будущего организма. Яйцо представляло как бы мозаику из зачатков органов, изъятие части мозаики вызывало отсутствие определенных органов. Одновременно с ним некоторые другие ученые стали использовать различные экспериментальные методики в своих исследованиях. Вскоре начали экспериментировать Г. Дриш, Ж. Леб и многие другие. Г. Дриш, которому эмбриология обязана выдающимися экспериментами по изоляции бластомеров.
Для прослеживания судьбы тех или иных бластомеров, для изучения передвижения клеточного материала в ходе развития важное значение имела разработанная В. Фогтом методики меток, наносимых витальным красителем на отдельные части зародыша. Эта методика дала возможность выяснить процессы гаструляции у амфибий и других животных.
Исключительное значение, притом все возрастающее в наше время, имела разработка методик культивирования тканей и зачатков органов вне организма, соответствующие способы хирургии, набор питательных сред, способы их стерилизации. Однако честь открытия метода тканевых культур принадлежит Р. Г. Гаррисону.
Наибольшее влияние на экспериментальную эмбриологию в XX в. оказала школа Ганса Шпемана, предложившего свою теорию индивидуального развития и разработавшего прекрасные методики микрохирургии на зародышах: снятие оболочек яиц животных, пересадка частей одного зародыша другому, изготовление благоприятной жидкой среды для развития и др. Шпеману и его ученикам удалось установить взаимозависимость частей развивающегося зародыша.
Одна из самых плодотворных теорий развития, которая объединяет усилия эмбриологов на протяжении всего XX в. и по настоящее время, - теория эмбриональной индукции.
Экспериментальная разработка будущей теории началась с разнообразных опытов по пересадке закладок у ранних зародышей амфибий в лаборатории Ганса Шпемана.
Немецкий ученый Г. Шпеман впервые установил, что закладка нервной системы у земноводных связана с материалом хорды, который, перемещаясь внутрь зародыша, располагается под дорзальной эктодермой, развивающейся в нервную систему. Материал хорды, который определяет закладку центральной нервной системы, был назван Шпеманом организационным центром.
Наличие формообразовательных влияний было установлено и при развитии ряда других органов. Впервые это было показано на примере развития глаза. Оказалось, у большинства исследованных животных при удалении зачатка глаза до его контакта с покрывающей эктодермой хрусталик не развивается.
Формообразовательное влияние при развитии глаза не является односторонним. Хрусталик со своей стороны действует на мозг.
Взаимодействие частей зародыша, в результате которого определяется развитие органов, получило название индукции, а сами части, определяющие развитие, - индукторов.
Исключительная роль в развитии эволюционной эмбриологии принадлежит отечественным эмбриологам Д.П. Филатову и П.П. Иванову. Разработали свои методики микрохирургии и положили начало сравнительной экспериментальной эмбриологии.
Современная эмбриология ставит своей задачей управление развитием организмов, Осуществление этой задачи возможно при условии тесной связи эмбриологии с другими науками, в первую очередь с гистологией и цитологией. Эмбриология должна быть теснейшим образом связана с генетикой и цитогенетикой. Тесная связь эмбриологии с экологическими науками проявляется при изучении влияния внешней среды на развитие организмов.
studfiles.net