
Нуклеиновые кислоты, биополимеры, состоящие из остатков фосфорной кислоты, сахаров и азотистых оснований (пуринов и пиримидинов). Имеют фундаментальное биологическое значение, поскольку содержат в закодированном виде всю генетическую информацию любого живого организма, от человека до бактерий и вирусов, передаваемую от одного поколения другому.
Нуклеиновые кислоты были впервые выделены из клеток гноя человека и спермы лосося швейцарским врачом и биохимиком Ф.Мишером между 1869 и 1871. Впоследствии было установлено, что существует два типа нуклеиновых кислот: рибонуклеиновая (РНК) и дезоксирибонуклеиновая (ДНК), однако их функции долго оставались неизвестными.
В 1928 английский бактериолог Ф.Гриффит обнаружил, что убитые патогенные пневмококки могут изменять генетические свойства живых непатогенных пневмококков, превращая последние в патогенные. В 1945 микробиолог О.Эвери из Рокфеллеровского института в Нью-Йорке сделал важное открытие: он показал, что способность к генетической трансформации обусловлена переносом ДНК из одной клетки в другую, а следовательно, генетический материал представляет собой ДНК. В 1940–1950 Дж.Бидл и Э.Тейтум из Станфордского университета (шт. Калифорния) обнаружили, что синтез белков, в частности ферментов, контролируется специфическими генами. В 1942 Т.Касперсон в Швеции и Ж.Браше в Бельгии открыли, что нуклеиновых кислот особенно много в клетках, активно синтезирующих белки. Все эти данные наводили на мысль, что генетический материал – это нуклеиновая кислота и что она как-то участвует в синтезе белков. Однако в то время многие полагали, что молекулы нуклеиновых кислот, несмотря на их большую длину, имеют слишком простую периодически повторяющуюся структуру, чтобы нести достаточно информации и служить генетическим материалом. Но в конце 1940-х годов Э.Чаргафф в США и Дж.Уайатт в Канаде, используя метод распределительной хроматографии на бумаге, показали, что структура ДНК не столь проста и эта молекула может служить носителем генетической информации.
Структура ДНК была установлена в 1953 М.Уилкинсом, Дж.Уотсоном и Ф.Криком в Англии. Это фундаментальное открытие позволило понять, как происходит удвоение (репликация) нуклеиновых кислот. Вскоре после этого американские исследователи А.Даунс и Дж.Гамов предположили, что структура белков каким-то образом закодирована в нуклеиновых кислотах, а к 1965 эта гипотеза была подтверждена многими исследователями: Ф.Криком в Англии, М.Ниренбергом и С.Очоа в США, Х.Кораной в Индии. Все эти открытия, результат столетнего изучения нуклеиновых кислот, произвели подлинную революцию в биологии. Они позволили объяснить феномен жизни в рамках взаимодействия между атомами и молекулами.
Типы и распространение. Как мы уже говорили, есть два типа нуклеиновых кислот: ДНК и РНК. ДНК присутствует в ядрах всех растительных и животных клеток, где она находится в комплексе с белками и является составной частью хромосом. У особей каждого конкретного вида содержание ядерной ДНК обычно одинаково во всех клетках, кроме гамет (яйцеклеток и сперматозоидов), где ДНК вдвое меньше. Таким образом, количество клеточной ДНК видоспецифично. ДНК найдена и вне ядра: в митохондриях («энергетических станциях» клеток) и в хлоропластах (частицах, где в растительных клетках идет фотосинтез). Эти субклеточные частицы обладают некоторой генетической автономией.
Бактерии и цианобактерии (сине-зеленые водоросли) содержат вместо хромосом одну или две крупные молекулы ДНК, связанные с небольшим количеством белка, и часто – молекулы ДНК меньшего размера, называемые плазмидами. Плазмиды несут полезную генетическую информацию, например содержат гены устойчивости к антибиотикам, но для жизни самой клетки они несущественны.
Некоторое количество РНК присутствует в клеточном ядре, основная же ее масса находится в цитоплазме – жидком содержимом клетки. Бльшую ее часть составляет рибосомная РНК (рРНК). Рибосомы – это мельчайшие тельца, на которых идет синтез белка. Небольшое количество РНК представлено транспортной РНК (тРНК), которая также участвует в белковом синтезе. Однако оба этих класса РНК не несут информации о структуре белков – такая информация заключена в матричной, или информационной, РНК (мРНК), на долю которой приходится лишь небольшая часть суммарной клеточной РНК.
Генетический материал вирусов представлен либо ДНК, либо РНК, но никогда обеими одновременно.
Общие свойства
Молекулы нуклеиновых кислот содержат множество отрицательно заряженных фосфатных групп и образуют комплексы с ионами металлов; их калиевая и натриевая соли хорошо растворимы в воде. Концентрированные растворы нуклеиновых кислот очень вязкие и слегка опалесцируют, а в твердом виде эти вещества белые. Нуклеиновые кислоты сильно поглощают ультрафиолетовый свет, и это свойство лежит в основе определения их концентрации. С этим же свойством связан и мутагенный эффект ультрафиолетового света.
Длинные молекулы ДНК хрупки и легко ломаются, например при продавливании раствора через шприц. Поэтому работа с высокомолекулярными ДНК требует особой осторожности.
Химическая структура. Нуклеиновые кислоты это длинные цепочки, состоящие из четырех многократно повторяющихся единиц (нуклеотидов). Их структуру можно представить следующим образом:
Символ Ф обозначает фосфатную группу. Чередующиеся остатки сахара и фосфорной кислоты образуют сахарофосфатный остов молекулы, одинаковый у всех ДНК, а огромное их разнообразие обусловливается тем, что четыре азотистых основания могут располагаться вдоль цепи в самой разной последовательности.
Сахаром в нуклеиновых кислотах является пентоза; четыре из пяти ее углеродных атомов вместе с одним атомом кислорода образуют кольцо. Атомы углерода пентозы обозначают номерами от 1 до 5. В РНК сахар представлен рибозой, а в ДНК дезоксирибозой, содержащей на один атом кислорода меньше. Фрагменты полинуклеотидных цепей ДНК и РНК показаны на рисунке.
Поскольку фосфатные группы присоединены к сахару асимметрично, в положениях 3 и 5, молекула нуклеиновой кислоты имеет определенное направление. Сложноэфирные связи между мономерными единицами нуклеиновых кислот чувствительны к гидролитическому расщеплению (ферментативному или химическому), которое приводит к высвобождению отдельных компонентов в виде небольших молекул.
Азотистые основания – это плоские гетероциклические соединения. Они присоединены к пентозному кольцу по положению 1. Более крупные основания имеют два кольца и называются пуринами: это аденин (А) и гуанин (Г). Основания, меньшие по размерам, имеют одно кольцо и называются пиримидинами: это цитозин (Ц), тимин (Т) и урацил (У). В ДНК входят основания А, Г, Т и Ц, в РНК вместо Т присутствует У. Последний отличается от тимина тем, что у него отсутствует метильная группа (Ch4). Урацил встречается в ДНК некоторых вирусов, где он выполняет ту же функцию, что и тимин.
Трехмерная структура. Важной особенностью нуклеиновых кислот является регулярность пространственного расположения составляющих их атомов, установленная рентгеноструктурным методом. Молекула ДНК состоит из двух противоположно направленных цепей (иногда содержащих миллионы нуклеотидов), удерживаемых вместе водородными связями между основаниями:
Водородные связи, соединяющие основания противоположных цепей, относятся к категории слабых, но благодаря своей многочисленности в молекуле ДНК они прочно стабилизируют ее структуру. Однако если раствор ДНК нагреть примерно до 60 С, эти связи рвутся и цепи расходятся – происходит денатурация ДНК (плавление).
Обе цепи ДНК закручены по спирали относительно воображаемой оси, как будто они навиты на цилиндр. Эта структура называется двойной спиралью. На каждый виток спирали приходится десять пар оснований.
Правило комплементарности. Уотсон и Крик показали, что образование водородных связей и регулярной двойной спирали возможно только тогда, когда более крупное пуриновое основание аденин (А) в одной цепи имеет своим партнером в другой цепи меньшее по размерам пиримидиновое основание тимин (Т), а гуанин (Г) связан с цитозином (Ц). Эту закономерность можно представить следующим образом:
Соответствие АТ и ГЦ называют правилом комплементарности, а сами цепи комплементарными. Согласно этому правилу, содержание аденина в ДНК всегда равно содержанию тимина, а количество гуанина – количеству цитозина. Следует отметить, что две цепи ДНК, различаясь химически, несут одинаковую информацию, поскольку вследствие комплементарности одна цепь однозначно задает другую.
Структура РНК менее упорядочена. Обычно это одноцепочечная молекула, хотя РНК некоторых вирусов состоит из двух цепей. Но даже такая РНК более гибка, чем ДНК. Некоторые участки в молекуле РНК взаимно комплементарны и при изгибании цепи спариваются, образуя двухцепочечные структуры (шпильки). В первую очередь это относится к транспортным РНК (тРНК). Некоторые основания в тРНК подвергаются модификации уже после синтеза молекулы. Например, иногда происходит присоединение к ним метильных групп.
Функция нуклеиновых кислот
Одна из основных функций нуклеиновых кислот состоит в детерминации синтеза белков. Информация о структуре белков, закодированная в нуклеотидной последовательности ДНК, должна передаваться от одного поколения к другому, и поэтому необходимо ее безошибочное копирование, т.е. синтез точно такой же же молекулы ДНК (репликация).
Репликация и транскрипция. С химической точки зрения синтез нуклеиновой кислоты – это полимеризация, т.е. последовательное присоединение строительных блоков. Такими блоками служат нуклеозидтрифосфаты; реакцию можно представить следующим образом:
Энергия, необходимая для синтеза, высвобождается при отщеплении пирофосфата, а катализируют реакцию особые ферменты – ДНК-полимеразы.
В результате такого синтетического процесса мы получили бы полимер со случайной последовательностью оснований. Однако большинство полимераз работает только в присутствии уже существующей нуклеиновой кислоты –матрицы, диктующей, какой именно нуклеотид присоединится к концу цепи. Этот нуклеотид должен быть комплементарен соответствующему нуклеотиду матрицы, так что новая цепь оказывается комплементарной исходной. Используя затем комплементарную цепь в качестве матрицы, мы получим точную копию оригинала.
ДНК состоит из двух взаимно комплементарных цепей. В ходе репликации они расходятся, и каждая из них служит матрицей для синтеза новой цепи:
Так образуются две новые двойные спирали с той же последовательностью оснований, что и у исходной ДНК. Иногда в процессе репликации происходит «сбой», и возникают мутации.
В результате транскрипции ДНК образуются клеточные РНК (мРНК, рРНК и тРНК):
Они комплементарны одной из цепей ДНК и являются копией другой цепи, за исключением того, что место тимина у них занимает урацил. Таким способом можно получить множество РНК-копий одной из цепей ДНК.
В нормальной клетке передача информации осуществляется только в направлении ДНК ДНК и ДНК РНК. Однако в клетках, инфицированных вирусом, возможны и другие процессы: РНК РНК и РНК ДНК. Генетический материал многих вирусов представлен молекулой РНК, обычно одноцепочечной. Проникнув в клетку-хозяина, эта РНК реплицируется с образованием комплементарной молекулы, на которой, в свою очередь, синтезируется множество копий исходной вирусной РНК:
Вирусная РНК может транскрибироваться ферментом обратной транскриптазой в ДНК, которая иногда включается в хромосомную ДНК клетки-хозяина. Теперь эта ДНК несет вирусные гены, и после транскрипции в клетке может появиться вирусная РНК. Таким образом, спустя длительное время, в течение которого никакого вируса в клетке не обнаруживается, он снова в ней появится без повторного заражения. Вирусы, генетический материал которых включается в хромосому клетки-хозяина, часто являются причиной рака.
Трансляция нуклеиновых кислот в белки. Генетическая информация, закодированная в нуклеотидной последовательности ДНК, переводится не только на язык нуклеотидной последовательности РНК, но и на язык аминокислот – мономерных единиц белков.
Белковая молекула – это цепочка из аминокислот. Каждая аминокислота содержит кислую карбоксильную группу –COOH и оснвную аминогруппу
–Nh3. Карбоксильная группа одной аминокислоты связывается с аминогруппой другой, образуя амидную связь, и этот процесс продолжается, пока не образуется цепь, содержащая до 1000 аминокислот.
В белках присутствует 20 разных аминокислот, от последовательности которых зависят их природа и функции. Эта последовательность определяется нуклеотидной последовательностью соответствующего гена – участка ДНК, кодирующего данный белок. Однако сама ДНК не является матрицей при синтезе белка. Сначала она транскрибируется в ядре с образованием матричной РНК (мРНК), которая диффундирует в цитоплазму, и на ней как на матрице синтезируется белок. Процесс ускоряется благодаря тому, что на каждой молекуле мРНК может одновременно синтезироваться множество белковых молекул.
Репликация нуклеиновых кислот осуществляется благодаря образованию водородных связей между комплементарными основаниями исходной и дочерней цепей. Аминокислоты не образуют водородных связей с основаниями, так что прямое копирование матрицы невозможно. Они взаимодействуют с матрицей опосредованно, через «адапторные» нуклеиновые кислоты – небольшие молекулы транспортных РНК (тРНК), состоящие примерно из 80 оснований и способные связываться с мРНК.
Каждая тРНК содержит специфическую последовательность из трех оснований, антикодон, который комплементарен группе из трех оснований, кодону, в мРНК. Антикодоны взаимодействуют с кодонами по правилу комплементарности, примерно так же, как взаимодействуют две цепи ДНК. Таким образом, последовательность оснований в мРНК определяет порядок присоединения тРНК, несущих аминокислоты. Схематически перенос информации от ДНК к белку можно представить следующим образом:
Последовательность оснований в ДНК задает порядок следования аминокислот в белке, поскольку каждая аминокислота присоединяется специфическим ферментом только к определенным тРНК, а те, в свою очередь, – только к определенным кодонам в мРНК. Комплексы тРНК-аминокислота связываются с матрицей по одному в каждый данный момент времени. Ниже перечислены основные этапы белкового синтеза (см. также рисунок).
1. Ферменты, называемые аминоацил-тРНК-синтетазами, присоединяют аминокислоты к соответствующим тРНК. Таких ферментов 20, по одному для каждой аминокислоты.
2. Молекула мРНК присоединяется своим первым кодоном к небольшой частице, называемой рибосомой. Рибосомы состоят из примерно равных количеств рРНК и белка. Структура и функция рибосом весьма сложны, но главная их задача – облегчение взаимодействия мРНК и тРНК и ускорение полимеризации аминокислот, связанных с разными тРНК.
3. тРНК, нагруженная аминокислотой, связывается с соответствующим кодоном мРНК, которая, в свою очередь, контактирует с рибосомой. Образуется комплекс рибосома-мРНК-тРНК-аминокислота.
4. мРНК, подобно ленте на конвейере, продвигается по рибосоме на один кодон вперед.
5. Следующая тРНК, нагруженная аминокислотой, присоединяется ко второму кодону.
6. Первая и вторая аминокислоты связываются между собой.
7. Первая тРНК отсоединяется от комплекса, и теперь вторая тРНК несет две аминокислоты, связанные между собой.
8. мРНК снова продвигается на один кодон вперед, и все события повторяются, а растущая аминокислотная цепь удлиняется на одну аминокислоту. Процесс продолжается, пока не будет достигнут последний, «стоп»-кодон и последняя тРНК не отделится от готовой белковой цепи. В бактериальных клетках цепь из 100–200 аминокислот собирается за несколько секунд. В животных клетках этот процесс занимает около минуты.
Генетический код. Итак, каждая аминокислота в белке опосредованно детерминируется определенным кодоном (группой из 3 оснований) в мРНК и в конечном счете в ДНК. Поскольку в нуклеиновых кислотах имеется четыре вида оснований, число возможных кодонов составляет 444 = 64. Соответствие между кодонами и аминокислотами, которые они кодируют, называется генетическим или биологическим кодом. Это соответствие было установлено опытным путем: к разрушенным клеткам добавляли синтетические полинуклеотиды известного состава и смотрели, какие аминокислоты включаются в белки. Позднее появилась возможность прямо сравнить последовательности аминокислот в вирусных белках и оснований в вирусных нуклеиновых кислотах. Чрезвычайно интересно, что генетический код, за редкими исключениями, одинаков для всех организмов – от вирусов до человека. Одно из таких исключений составляют изменения в генетическом коде, используемом митохондриями. Митохондрии это небольшие автономные субклеточные частицы (органеллы), присутствующие во всех клетках, кроме бактерий и зрелых эритроцитов. Предполагают, что когда-то митохондрии были самостоятельными организмами; проникнув в клетки, они со временем стали их неотъемлемой частью, но сохранили некоторое количество собственной ДНК и синтезируют несколько митохондриальных белков.
| Аланин | Аргинин | Аспарагин | Аспарагиновая кислота | |
| ГЦУ | ЦГУ | ГАУ | ААУ | |
| ГЦЦ | ЦГЦ | ГАЦ | ААЦ | |
| ГЦА | ЦГА | |||
| ГЦГ | ЦГГ | |||
| АГА | ||||
| АГГ | ||||
| Валин | Гистидин | Глицин | Глутаминовая кислота | |
| ГУУ | ЦАУ | ГГУ | ЦАА | |
| ГУЦ | ЦАЦ | ГГЦ | ЦАГ | |
| ГУА | ГГА | |||
| ГУГ | ГГГ | |||
| Глутамин | Изолейцин | Лейцин | Лизин | |
| ГАА | АУУ | УУА | ААА | |
| ГАГ | АУЦ | УУГ | ААГ | |
| АУА | ЦУУ | |||
| ЦУЦ | ||||
| ЦУА | ||||
| ЦУГ | ||||
| Метионин | Пролин | Серин | Тирозин | |
| АУГ | ЦЦУ | АГУ | УАУ | |
| ЦЦЦ | АГЦ | УАЦ | ||
| ЦЦА | УЦА | |||
| ЦЦГ | УЦГ | |||
| УЦУ | ||||
| УЦЦ | ||||
| Треонин | Триптофан | Фенилаланин | Цистеин | Нет |
| АЦУ | УГГ | УУУ | УГУ | УАА |
| АЦЦ | УУЦ | УГЦ | УАГ | |
| АЦА | УГА | |||
| АЦГ | ||||
| ГЕНЕТИЧЕСКИЙ СЛОВАРЬ: указаны аминокислоты, встречающиеся в белках, и соответствующие им кодоны в мРНК. «Буквы» в кодонах записаны в направлении 5' 3'. В этом же направлении идут транскрипции нуклеиновых кислот и синтез белка на матрице. «Нет» означает, что кодон не кодирует никаких аминокислот; такие кодоны называются «бессмысленными». Генетический словарь одинаков для всех живых организмов – от вирусов до человека. |
Вообще говоря, каждой аминокислоте соответствует более одного кодона. Большинство кодонов, кодирующих одну и ту же аминокислоту, имеют два одинаковых первых основания, но в трех случаях (для лейцина, серина и аргинина) имеются два альтернативных набора первых дублетов в кодонах, соответствующих одной и той же аминокислоте. Природа основания в третьем положении не столь важна; одна и та же аминокислота глицин может кодироваться по-разному: ГГУ, ГГЦ, ГГА и ГГГ. Однако кодоны для двух разных аминокислот могут иметь два одинаковых первых основания, и тогда различие между ними будет определяться природой третьего основания – пурином или пиримидином. Так, гистидин кодируется триплетами ЦАУ и ЦАЦ, а глутамин ЦАА и ЦАГ. Три кодона, УАА, УАГ и УГА, не кодируют никаких аминокислот и называются «бессмысленными».
Одна молекула ДНК кодирует много белковых цепей. Каждый отрезок, кодирующий одну цепь, называют цистроном. Начало и конец цистрона, а также граница раздела между ними помечаются с помощью своего рода знаков химической пунктуации. По крайней мере у бактерий в начале цистрона находится метиониновый кодон АУГ. Логично предположить, что первой аминокислотой в белке всегда должен быть метионин, но часто несколько первых аминокислот отщепляются ферментативно после окончания синтеза белка. Конец белковой цепи помечается одним или несколькими «бессмысленными» кодонами.
У бактерий (прокариот) практически вся ДНК кодирует какие-либо белки или тРНК. Однако у высших форм (эукариот) значительная часть ДНК состоит из простых повторяющихся последовательностей и «молчащих» генов, которые не транскрибируются в РНК и поэтому не транслируются в белки. Кроме того, исходно синтезированная мРНК содержит участки, не детерминирующие никаких белковых последовательностей. Такие участки (интроны), расположенные между кодирующими участками (экзонами), перед началом синтеза белка удаляются специальными ферментами. Почему в ДНК существуют эти казалось бы бесполезные сегменты – неясно; возможно, они выполняют регуляторные функции.
У простейшей Tetrahymena РНК сама удаляет свои интроны и соединяет свободные концы цепей, действуя как фермент по отношению к себе самой. Это единственное известное исключение из правила, согласно которому нуклеиновые кислоты не обладают ферментативной активностью.
Транспортные РНК и супрессия. Смысл информации, содержащейся в ДНК, если переводить ее на язык аминокислот, определяется как самой ДНК, так и считывающим механизмом, т.е. зависит не только от того, какие кодоны есть в ДНК и в какой последовательности они расположены, но также и от того, какие именно аминокислоты (и к каким тРНК) присоединяют аминоацил-тРНК-синтетазы. Конечно, природа синтетаз и тРНК тоже определяется ДНК, и в этом смысле ДНК является первичным детерминантом белковой последовательности. Тем не менее суммарная детерминация представляет собой функцию всей системы, поскольку результат зависит от исходных компонентов. Если бы соответствие между тРНК и аминокислотами было другим, смысл кодонов тоже изменился бы.
Известно, что мутации в ДНК изменяют считывающий механизм и в результате меняют – пусть и незначительно – смысл кодонов. Так, в бактерии Escherichia coli глициновая тРНК обычно узнает в мРНК кодон ГГА; мутация в ДНК, с которой транскрибируется эта тРНК, изменяет антикодон глициновой тРНК таким образом, что теперь он узнает кодон АГА, соответствующий аргинину, и в белковой молекуле вместо аргинина появляется глицин. Это не обязательно имеет фатальные последствия, поскольку не все аргинины кодируются триплетом АГА и есть аргининовые тРНК, по-прежнему узнающие «свои» АГА. В результате измененными оказываются не все белковые молекулы. Иногда такие мутации, изменяющие антикодон, подавляют (супрессируют) мутации в кодоне. Например, если в результате мутации глициновый кодон ГГА превращается в АГА, он все же может прочитываться как глицин, если антикодон глициновой тРНК, в свою очередь, изменился так, что эта тРНК стала узнавать АГА. В этом случае вторая «ошибка» устраняет первую.
Мутации, приводящие к изменению антикодонов, могут иметь разные последствия, поскольку один и тот же кодон может узнаваться несколькими тРНК. Вообще говоря, узнавание осуществляется благодаря комплементарности оснований кодона и антикодона, однако одно из оснований кодона может модифицироваться таким образом, что антикодон будет узнавать даже неполностью комплементарный кодон. В результате одна и та же тРНК может взаимодействовать с несколькими разными кодонами, кодирующими одну и ту же аминокислоту. Этот феномен неполного соответствия кодона и антикодона был назван Ф.Криком «шатанием».
Регуляция активности генов. Для организма было бы катастрофой, если бы во всех его клетках одновременно работали все гены и синтезировались все закодированные ими белки. Бактерии, например, должны все время приспосабливаться к условиям среды, синтезируя нужные ферменты. Все клетки высших организмов имеют один и тот же набор генов, но, к счастью, клетки мозга не продуцируют пищеварительные ферменты, а в хрусталике глаза не синтезируются мышечные белки.
Активность гена характеризуется тем, транскрибируется ли он с образованием соответствующей мРНК. ДНК длинная молекула, и в определенных ее участках имеются последовательности, называемые промоторами, которые распознаются специфическим транскрибирующим ферментом полимеразой. В этих участках и только в них начинается транскрипция, продолжаясь до тех пор, пока не достигнет последовательности оснований, означающей конец считывания.
Существуют особые репрессорные белки, которые связываются с ДНК поблизости от промотора в участке, называемом оператором. Образовавшийся комплекс блокирует транскрипцию, и мРНК не синтезируется. Таким образом, репрессорные белки являются ингибиторами транскрипции. С другой стороны, существуют небольшие молекулы, которые образуют комплекс с репрессорами и снимают их блокирующее действие на транскрипцию. Иными словами, они ингибируют ингибиторы. Так, у бактерий в норме отсутствуют ферменты, катализирующие расщепление некоторых сахаров; однако если один из этих сахаров появляется в среде, он образует комплекс с репрессором, ингибирование снимается и запускается синтез соответствующего фермента. Ферменты, синтез которых индуцируется собственными субстратами, называются индуцибельными. В ряде случаев, наоборот, репрессорный белок не блокирует транскрипцию мРНК, если он не связан с определенной молекулой. У бактерий некоторые ферменты, участвующие в синтезе определенных аминокислот, образуются только в отсутствие этих аминокислот, т.е. бактерии производят данные ферменты лишь по мере надобности. Если добавить в среду соответствующую аминокислоту, она образует комплекс с репрессором и активирует его, а тем самым ингибирует транскрипцию соответствующих генов. Уже образовавшаяся мРНК вскоре расщепляется, и синтез ферментов останавливается. Такие ферменты являются отрицально индуцибельными.
Поскольку репрессорные белки сами кодируются генами, работа которых, в свою очередь, может регулироваться другими генами, а синтез малых молекул-индукторов и гормонов также в конечном счете регулируется генами, механизмы регуляции генной активности могут быть очень сложными.
Список литературы
Ичас М. Биологический код. М., 1971
Шабарова З.А., Богданов А.А. Химия нуклеиновых кислот и их компонентов, М., 1978
Зенгер В. Принципы структурной организации нуклеиновых кислот. М., 1987
www.ronl.ru
Нуклеиновые кислоты
Нуклеиновые кислоты, биополимеры, состоящие из остатков фосфорной кислоты, сахаров и азотистых оснований (пуринов и пиримидинов). Имеют фундаментальное биологическое значение, поскольку содержат в закодированном виде всю генетическую информацию любого живого организма, от человека до бактерий и вирусов, передаваемую от одного поколения другому.
Нуклеиновые кислоты были впервые выделены из клеток гноя человека и спермы лосося швейцарским врачом и биохимиком Ф.Мишером между 1869 и 1871. Впоследствии было установлено, что существует два типа нуклеиновых кислот: рибонуклеиновая (РНК) и дезоксирибонуклеиновая (ДНК), однако их функции долго оставались неизвестными.
В 1928 английский бактериолог Ф.Гриффит обнаружил, что убитые патогенные пневмококки могут изменять генетические свойства живых непатогенных пневмококков, превращая последние в патогенные. В 1945 микробиолог О.Эвери из Рокфеллеровского института в Нью-Йорке сделал важное открытие: он показал, что способность к генетической трансформации обусловлена переносом ДНК из одной клетки в другую, а следовательно, генетический материал представляет собой ДНК. В 1940–1950 Дж.Бидл и Э.Тейтум из Станфордского университета (шт. Калифорния) обнаружили, что синтез белков, в частности ферментов, контролируется специфическими генами. В 1942 Т.Касперсон в Швеции и Ж.Браше в Бельгии открыли, что нуклеиновых кислот особенно много в клетках, активно синтезирующих белки. Все эти данные наводили на мысль, что генетический материал – это нуклеиновая кислота и что она как-то участвует в синтезе белков. Однако в то время многие полагали, что молекулы нуклеиновых кислот, несмотря на их большую длину, имеют слишком простую периодически повторяющуюся структуру, чтобы нести достаточно информации и служить генетическим материалом. Но в конце 1940-х годов Э.Чаргафф в США и Дж.Уайатт в Канаде, используя метод распределительной хроматографии на бумаге, показали, что структура ДНК не столь проста и эта молекула может служить носителем генетической информации.
Структура ДНК была установлена в 1953 М.Уилкинсом, Дж.Уотсоном и Ф.Криком в Англии. Это фундаментальное открытие позволило понять, как происходит удвоение (репликация) нуклеиновых кислот. Вскоре после этого американские исследователи А.Даунс и Дж.Гамов предположили, что структура белков каким-то образом закодирована в нуклеиновых кислотах, а к 1965 эта гипотеза была подтверждена многими исследователями: Ф.Криком в Англии, М.Ниренбергом и С.Очоа в США, Х.Кораной в Индии. Все эти открытия, результат столетнего изучения нуклеиновых кислот, произвели подлинную революцию в биологии. Они позволили объяснить феномен жизни в рамках взаимодействия между атомами и молекулами.
Типы и распространение. Как мы уже говорили, есть два типа нуклеиновых кислот: ДНК и РНК. ДНК присутствует в ядрах всех растительных и животных клеток, где она находится в комплексе с белками и является составной частью хромосом. У особей каждого конкретного вида содержание ядерной ДНК обычно одинаково во всех клетках, кроме гамет (яйцеклеток и сперматозоидов), где ДНК вдвое меньше. Таким образом, количество клеточной ДНК видоспецифично. ДНК найдена и вне ядра: в митохондриях («энергетических станциях» клеток) и в хлоропластах (частицах, где в растительных клетках идет фотосинтез). Эти субклеточные частицы обладают некоторой генетической автономией.
Бактерии и цианобактерии (сине-зеленые водоросли) содержат вместо хромосом одну или две крупные молекулы ДНК, связанные с небольшим количеством белка, и часто – молекулы ДНК меньшего размера, называемые плазмидами. Плазмиды несут полезную генетическую информацию, например содержат гены устойчивости к антибиотикам, но для жизни самой клетки они несущественны.
Некоторое
количество РНК присутствует в клеточном ядре, основная же ее масса находится в
цитоплазме – жидком содержимом клетки. Б льшую ее часть составляет рибосомная
РНК (рРНК). Рибосомы – это мельчайшие тельца, на которых идет синтез белка.
Небольшое количество РНК представлено транспортной РНК (тРНК), которая также
участвует в белковом синтезе. Однако оба этих класса РНК не несут информации о
структуре белков – такая информация заключена в матричной, или информационной,
РНК (мРНК), на долю которой приходится лишь небольшая часть суммарной клеточной
РНК.
льшую ее часть составляет рибосомная
РНК (рРНК). Рибосомы – это мельчайшие тельца, на которых идет синтез белка.
Небольшое количество РНК представлено транспортной РНК (тРНК), которая также
участвует в белковом синтезе. Однако оба этих класса РНК не несут информации о
структуре белков – такая информация заключена в матричной, или информационной,
РНК (мРНК), на долю которой приходится лишь небольшая часть суммарной клеточной
РНК.
Генетический материал вирусов представлен либо ДНК, либо РНК, но никогда обеими одновременно.
Общие свойства
Молекулы нуклеиновых кислот содержат множество отрицательно заряженных фосфатных групп и образуют комплексы с ионами металлов; их калиевая и натриевая соли хорошо растворимы в воде. Концентрированные растворы нуклеиновых кислот очень вязкие и слегка опалесцируют, а в твердом виде эти вещества белые. Нуклеиновые кислоты сильно поглощают ультрафиолетовый свет, и это свойство лежит в основе определения их концентрации. С этим же свойством связан и мутагенный эффект ультрафиолетового света.
Длинные молекулы ДНК хрупки и легко ломаются, например при продавливании раствора через шприц. Поэтому работа с высокомолекулярными ДНК требует особой осторожности.
Химическая структура. Нуклеиновые кислоты это длинные цепочки, состоящие из четырех многократно повторяющихся единиц (нуклеотидов). Их структуру можно представить следующим образом:

Символ Ф обозначает фосфатную группу. Чередующиеся остатки сахара и фосфорной кислоты образуют сахарофосфатный остов молекулы, одинаковый у всех ДНК, а огромное их разнообразие обусловливается тем, что четыре азотистых основания могут располагаться вдоль цепи в самой разной последовательности.
Сахаром в нуклеиновых кислотах является пентоза; четыре из пяти ее углеродных атомов вместе с одним атомом кислорода образуют кольцо. Атомы углерода пентозы обозначают номерами от 1 до 5. В РНК сахар представлен рибозой, а в ДНК дезоксирибозой, содержащей на один атом кислорода меньше. Фрагменты полинуклеотидных цепей ДНК и РНК показаны на рисунке.
Поскольку фосфатные группы присоединены к сахару асимметрично, в положениях 3 и 5, молекула нуклеиновой кислоты имеет определенное направление. Сложноэфирные связи между мономерными единицами нуклеиновых кислот чувствительны к гидролитическому расщеплению (ферментативному или химическому), которое приводит к высвобождению отдельных компонентов в виде небольших молекул.
Азотистые основания – это плоские гетероциклические соединения. Они присоединены к пентозному кольцу по положению 1. Более крупные основания имеют два кольца и называются пуринами: это аденин (А) и гуанин (Г). Основания, меньшие по размерам, имеют одно кольцо и называются пиримидинами: это цитозин (Ц), тимин (Т) и урацил (У). В ДНК входят основания А, Г, Т и Ц, в РНК вместо Т присутствует У. Последний отличается от тимина тем, что у него отсутствует метильная группа (Ch4). Урацил встречается в ДНК некоторых вирусов, где он выполняет ту же функцию, что и тимин.
Трехмерная структура. Важной особенностью нуклеиновых кислот является регулярность пространственного расположения составляющих их атомов, установленная рентгеноструктурным методом. Молекула ДНК состоит из двух противоположно направленных цепей (иногда содержащих миллионы нуклеотидов), удерживаемых вместе водородными связями между основаниями:

Водородные связи, соединяющие основания противоположных цепей, относятся к категории слабых, но благодаря своей многочисленности в молекуле ДНК они прочно стабилизируют ее структуру. Однако если раствор ДНК нагреть примерно до 60 С, эти связи рвутся и цепи расходятся – происходит денатурация ДНК (плавление).
Обе цепи ДНК закручены по спирали относительно воображаемой оси, как будто они навиты на цилиндр. Эта структура называется двойной спиралью. На каждый виток спирали приходится десять пар оснований.
Правило комплементарности. Уотсон и Крик показали, что образование водородных связей и регулярной двойной спирали возможно только тогда, когда более крупное пуриновое основание аденин (А) в одной цепи имеет своим партнером в другой цепи меньшее по размерам пиримидиновое основание тимин (Т), а гуанин (Г) связан с цитозином (Ц). Эту закономерность можно представить следующим образом:

Соответствие АТ и ГЦ называют правилом комплементарности, а сами цепи комплементарными. Согласно этому правилу, содержание аденина в ДНК всегда равно содержанию тимина, а количество гуанина – количеству цитозина. Следует отметить, что две цепи ДНК, различаясь химически, несут одинаковую информацию, поскольку вследствие комплементарности одна цепь однозначно задает другую.
Структура РНК менее упорядочена. Обычно это одноцепочечная молекула, хотя РНК некоторых вирусов состоит из двух цепей. Но даже такая РНК более гибка, чем ДНК. Некоторые участки в молекуле РНК взаимно комплементарны и при изгибании цепи спариваются, образуя двухцепочечные структуры (шпильки). В первую очередь это относится к транспортным РНК (тРНК). Некоторые основания в тРНК подвергаются модификации уже после синтеза молекулы. Например, иногда происходит присоединение к ним метильных групп.
Функция нуклеиновых кислот
Одна из основных функций нуклеиновых кислот состоит в детерминации синтеза белков. Информация о структуре белков, закодированная в нуклеотидной последовательности ДНК, должна передаваться от одного поколения к другому, и поэтому необходимо ее безошибочное копирование, т.е. синтез точно такой же же молекулы ДНК (репликация).
Репликация и транскрипция. С химической точки зрения синтез нуклеиновой кислоты – это полимеризация, т.е. последовательное присоединение строительных блоков. Такими блоками служат нуклеозидтрифосфаты; реакцию можно представить следующим образом:

Энергия, необходимая для синтеза, высвобождается при отщеплении пирофосфата, а катализируют реакцию особые ферменты – ДНК-полимеразы.
В результате такого синтетического процесса мы получили бы полимер со случайной последовательностью оснований. Однако большинство полимераз работает только в присутствии уже существующей нуклеиновой кислоты –матрицы, диктующей, какой именно нуклеотид присоединится к концу цепи. Этот нуклеотид должен быть комплементарен соответствующему нуклеотиду матрицы, так что новая цепь оказывается комплементарной исходной. Используя затем комплементарную цепь в качестве матрицы, мы получим точную копию оригинала.
ДНК состоит из двух взаимно комплементарных цепей. В ходе репликации они расходятся, и каждая из них служит матрицей для синтеза новой цепи:

Так образуются две новые двойные спирали с той же последовательностью оснований, что и у исходной ДНК. Иногда в процессе репликации происходит «сбой», и возникают мутации .
В результате транскрипции ДНК образуются клеточные РНК (мРНК, рРНК и тРНК):

Они комплементарны одной из цепей ДНК и являются копией другой цепи, за исключением того, что место тимина у них занимает урацил. Таким способом можно получить множество РНК-копий одной из цепей ДНК.
В нормальной клетке передача информации осуществляется только в направлении ДНК ДНК и ДНК РНК. Однако в клетках, инфицированных вирусом, возможны и другие процессы: РНК РНК и РНК ДНК. Генетический материал многих вирусов представлен молекулой РНК, обычно одноцепочечной. Проникнув в клетку-хозяина, эта РНК реплицируется с образованием комплементарной молекулы, на которой, в свою очередь, синтезируется множество копий исходной вирусной РНК:

Вирусная РНК может транскрибироваться ферментом обратной транскриптазой в ДНК, которая иногда включается в хромосомную ДНК клетки-хозяина. Теперь эта ДНК несет вирусные гены, и после транскрипции в клетке может появиться вирусная РНК. Таким образом, спустя длительное время, в течение которого никакого вируса в клетке не обнаруживается, он снова в ней появится без повторного заражения. Вирусы, генетический материал которых включается в хромосому клетки-хозяина, часто являются причиной рака.
Трансляция нуклеиновых кислот в белки. Генетическая информация, закодированная в нуклеотидной последовательности ДНК, переводится не только на язык нуклеотидной последовательности РНК, но и на язык аминокислот – мономерных единиц белков.
Белковая
молекула – это цепочка из аминокислот. Каждая аминокислота содержит кислую
карбоксильную группу –COOH и оснвную аминогруппу
–Nh3. Карбоксильная группа одной аминокислоты связывается с аминогруппой другой, образуя амидную связь, и этот процесс продолжается, пока не образуется цепь, содержащая до 1000 аминокислот .
В белках присутствует 20 разных аминокислот, от последовательности которых зависят их природа и функции. Эта последовательность определяется нуклеотидной последовательностью соответствующего гена – участка ДНК, кодирующего данный белок. Однако сама ДНК не является матрицей при синтезе белка. Сначала она транскрибируется в ядре с образованием матричной РНК (мРНК), которая диффундирует в цитоплазму, и на ней как на матрице синтезируется белок. Процесс ускоряется благодаря тому, что на каждой молекуле мРНК может одновременно синтезироваться множество белковых молекул.
Репликация нуклеиновых кислот осуществляется благодаря образованию водородных связей между комплементарными основаниями исходной и дочерней цепей. Аминокислоты не образуют водородных связей с основаниями, так что прямое копирование матрицы невозможно. Они взаимодействуют с матрицей опосредованно, через «адапторные» нуклеиновые кислоты – небольшие молекулы транспортных РНК (тРНК), состоящие примерно из 80 оснований и способные связываться с мРНК.
Каждая тРНК содержит специфическую последовательность из трех оснований, антикодон, который комплементарен группе из трех оснований, кодону, в мРНК. Антикодоны взаимодействуют с кодонами по правилу комплементарности, примерно так же, как взаимодействуют две цепи ДНК. Таким образом, последовательность оснований в мРНК определяет порядок присоединения тРНК, несущих аминокислоты. Схематически перенос информации от ДНК к белку можно представить следующим образом:

Последовательность оснований в ДНК задает порядок следования аминокислот в белке, поскольку каждая аминокислота присоединяется специфическим ферментом только к определенным тРНК, а те, в свою очередь, – только к определенным кодонам в мРНК. Комплексы тРНК-аминокислота связываются с матрицей по одному в каждый данный момент времени. Ниже перечислены основные этапы белкового синтеза (см. также рисунок).
1. Ферменты, называемые аминоацил-тРНК-синтетазами, присоединяют аминокислоты к соответствующим тРНК. Таких ферментов 20, по одному для каждой аминокислоты.
2. Молекула мРНК присоединяется своим первым кодоном к небольшой частице, называемой рибосомой. Рибосомы состоят из примерно равных количеств рРНК и белка. Структура и функция рибосом весьма сложны, но главная их задача – облегчение взаимодействия мРНК и тРНК и ускорение полимеризации аминокислот, связанных с разными тРНК.
3. тРНК, нагруженная аминокислотой, связывается с соответствующим кодоном мРНК, которая, в свою очередь, контактирует с рибосомой. Образуется комплекс рибосома-мРНК-тРНК-аминокислота.
4. мРНК, подобно ленте на конвейере, продвигается по рибосоме на один кодон вперед.
5. Следующая тРНК, нагруженная аминокислотой, присоединяется ко второму кодону.
6. Первая и вторая аминокислоты связываются между собой.
7. Первая тРНК отсоединяется от комплекса, и теперь вторая тРНК несет две аминокислоты, связанные между собой.
8. мРНК снова продвигается на один кодон вперед, и все события повторяются, а растущая аминокислотная цепь удлиняется на одну аминокислоту. Процесс продолжается, пока не будет достигнут последний, «стоп»-кодон и последняя тРНК не отделится от готовой белковой цепи. В бактериальных клетках цепь из 100–200 аминокислот собирается за несколько секунд. В животных клетках этот процесс занимает около минуты.
Генетический код. Итак, каждая аминокислота в белке опосредованно детерминируется определенным кодоном (группой из 3 оснований) в мРНК и в конечном счете в ДНК. Поскольку в нуклеиновых кислотах имеется четыре вида оснований, число возможных кодонов составляет 444 = 64. Соответствие между кодонами и аминокислотами, которые они кодируют, называется генетическим или биологическим кодом. Это соответствие было установлено опытным путем: к разрушенным клеткам добавляли синтетические полинуклеотиды известного состава и смотрели, какие аминокислоты включаются в белки. Позднее появилась возможность прямо сравнить последовательности аминокислот в вирусных белках и оснований в вирусных нуклеиновых кислотах. Чрезвычайно интересно, что генетический код, за редкими исключениями, одинаков для всех организмов – от вирусов до человека. Одно из таких исключений составляют изменения в генетическом коде, используемом митохондриями. Митохондрии это небольшие автономные субклеточные частицы (органеллы), присутствующие во всех клетках, кроме бактерий и зрелых эритроцитов. Предполагают, что когда-то митохондрии были самостоятельными организмами; проникнув в клетки, они со временем стали их неотъемлемой частью, но сохранили некоторое количество собственной ДНК и синтезируют несколько митохондриальных белков.
|
Аланин |
Аргинин |
Аспарагин |
Аспарагиновая кислота |
|
|
ГЦУ |
ЦГУ |
ГАУ |
ААУ |
|
|
ГЦЦ |
ЦГЦ |
ГАЦ |
ААЦ |
|
|
ГЦА |
ЦГА |
|
|
|
|
ГЦГ |
ЦГГ |
|
|
|
|
|
АГА |
|
|
|
|
|
АГГ |
|
|
|
|
Валин |
Гистидин |
Глицин |
Глутаминовая кислота |
|
|
ГУУ |
ЦАУ |
ГГУ |
ЦАА |
|
|
ГУЦ |
ЦАЦ |
ГГЦ |
ЦАГ |
|
|
ГУА |
|
ГГА |
|
|
|
ГУГ |
|
ГГГ |
|
|
|
Глутамин |
Изолейцин |
Лейцин |
Лизин |
|
|
ГАА |
АУУ |
УУА |
ААА |
|
|
ГАГ |
АУЦ |
УУГ |
ААГ |
|
|
|
АУА |
ЦУУ |
|
|
|
|
|
ЦУЦ |
|
|
|
|
|
ЦУА |
|
|
|
|
|
ЦУГ |
|
|
|
Метионин |
Пролин |
Серин |
Тирозин |
|
|
АУГ |
ЦЦУ |
АГУ |
УАУ |
|
|
|
ЦЦЦ |
АГЦ |
УАЦ |
|
|
|
ЦЦА |
УЦА |
|
|
|
|
ЦЦГ |
УЦГ |
|
|
|
|
|
УЦУ |
|
|
|
|
|
УЦЦ |
|
|
|
Треонин |
Триптофан |
Фенилаланин |
Цистеин |
Нет |
|
АЦУ |
УГГ |
УУУ |
УГУ |
УАА |
|
АЦЦ |
|
УУЦ |
УГЦ |
УАГ |
|
АЦА |
|
|
|
УГА |
|
АЦГ |
|
|
|
|
|
ГЕНЕТИЧЕСКИЙ СЛОВАРЬ: указаны аминокислоты, встречающиеся в белках, и соответствующие им кодоны в мРНК. «Буквы» в кодонах записаны в направлении 5' 3' . В этом же направлении идут транскрипции нуклеиновых кислот и синтез белка на матрице. «Нет» означает, что кодон не кодирует никаких аминокислот; такие кодоны называются «бессмысленными». Генетический словарь одинаков для всех живых организмов – от вирусов до человека. |
||||
Вообще говоря, каждой аминокислоте соответствует более одного кодона. Большинство кодонов, кодирующих одну и ту же аминокислоту, имеют два одинаковых первых основания, но в трех случаях (для лейцина, серина и аргинина) имеются два альтернативных набора первых дублетов в кодонах, соответствующих одной и той же аминокислоте. Природа основания в третьем положении не столь важна; одна и та же аминокислота глицин может кодироваться по-разному: ГГУ, ГГЦ, ГГА и ГГГ. Однако кодоны для двух разных аминокислот могут иметь два одинаковых первых основания, и тогда различие между ними будет определяться природой третьего основания – пурином или пиримидином. Так, гистидин кодируется триплетами ЦАУ и ЦАЦ, а глутамин ЦАА и ЦАГ. Три кодона, УАА, УАГ и УГА, не кодируют никаких аминокислот и называются «бессмысленными».
Одна молекула ДНК кодирует много белковых цепей. Каждый отрезок, кодирующий одну цепь, называют цистроном. Начало и конец цистрона, а также граница раздела между ними помечаются с помощью своего рода знаков химической пунктуации. По крайней мере у бактерий в начале цистрона находится метиониновый кодон АУГ. Логично предположить, что первой аминокислотой в белке всегда должен быть метионин, но часто несколько первых аминокислот отщепляются ферментативно после окончания синтеза белка. Конец белковой цепи помечается одним или несколькими «бессмысленными» кодонами.
У бактерий (прокариот) практически вся ДНК кодирует какие-либо белки или тРНК. Однако у высших форм (эукариот) значительная часть ДНК состоит из простых повторяющихся последовательностей и «молчащих» генов, которые не транскрибируются в РНК и поэтому не транслируются в белки. Кроме того, исходно синтезированная мРНК содержит участки, не детерминирующие никаких белковых последовательностей. Такие участки (интроны), расположенные между кодирующими участками (экзонами), перед началом синтеза белка удаляются специальными ферментами. Почему в ДНК существуют эти казалось бы бесполезные сегменты – неясно; возможно, они выполняют регуляторные функции.
У простейшей Tetrahymena РНК сама удаляет свои интроны и соединяет свободные концы цепей, действуя как фермент по отношению к себе самой. Это единственное известное исключение из правила, согласно которому нуклеиновые кислоты не обладают ферментативной активностью.
Транспортные РНК и супрессия. Смысл информации, содержащейся в ДНК, если переводить ее на язык аминокислот, определяется как самой ДНК, так и считывающим механизмом, т.е. зависит не только от того, какие кодоны есть в ДНК и в какой последовательности они расположены, но также и от того, какие именно аминокислоты (и к каким тРНК) присоединяют аминоацил-тРНК-синтетазы. Конечно, природа синтетаз и тРНК тоже определяется ДНК, и в этом смысле ДНК является первичным детерминантом белковой последовательности. Тем не менее суммарная детерминация представляет собой функцию всей системы, поскольку результат зависит от исходных компонентов. Если бы соответствие между тРНК и аминокислотами было другим, смысл кодонов тоже изменился бы.
Известно, что мутации в ДНК изменяют считывающий механизм и в результате меняют – пусть и незначительно – смысл кодонов. Так, в бактерии Escherichia coli глициновая тРНК обычно узнает в мРНК кодон ГГА; мутация в ДНК, с которой транскрибируется эта тРНК, изменяет антикодон глициновой тРНК таким образом, что теперь он узнает кодон АГА, соответствующий аргинину, и в белковой молекуле вместо аргинина появляется глицин. Это не обязательно имеет фатальные последствия, поскольку не все аргинины кодируются триплетом АГА и есть аргининовые тРНК, по-прежнему узнающие «свои» АГА. В результате измененными оказываются не все белковые молекулы. Иногда такие мутации, изменяющие антикодон, подавляют (супрессируют) мутации в кодоне. Например, если в результате мутации глициновый кодон ГГА превращается в АГА, он все же может прочитываться как глицин, если антикодон глициновой тРНК, в свою очередь, изменился так, что эта тРНК стала узнавать АГА. В этом случае вторая «ошибка» устраняет первую.
Мутации, приводящие к изменению антикодонов, могут иметь разные последствия, поскольку один и тот же кодон может узнаваться несколькими тРНК. Вообще говоря, узнавание осуществляется благодаря комплементарности оснований кодона и антикодона, однако одно из оснований кодона может модифицироваться таким образом, что антикодон будет узнавать даже неполностью комплементарный кодон. В результате одна и та же тРНК может взаимодействовать с несколькими разными кодонами, кодирующими одну и ту же аминокислоту. Этот феномен неполного соответствия кодона и антикодона был назван Ф.Криком «шатанием».
Регуляция активности генов. Для организма было бы катастрофой, если бы во всех его клетках одновременно работали все гены и синтезировались все закодированные ими белки. Бактерии, например, должны все время приспосабливаться к условиям среды, синтезируя нужные ферменты. Все клетки высших организмов имеют один и тот же набор генов, но, к счастью, клетки мозга не продуцируют пищеварительные ферменты, а в хрусталике глаза не синтезируются мышечные белки.
Активность гена характеризуется тем, транскрибируется ли он с образованием соответствующей мРНК. ДНК длинная молекула, и в определенных ее участках имеются последовательности, называемые промоторами, которые распознаются специфическим транскрибирующим ферментом полимеразой. В этих участках и только в них начинается транскрипция, продолжаясь до тех пор, пока не достигнет последовательности оснований, означающей конец считывания.
Существуют особые репрессорные белки, которые связываются с ДНК поблизости от промотора в участке, называемом оператором. Образовавшийся комплекс блокирует транскрипцию, и мРНК не синтезируется. Таким образом, репрессорные белки являются ингибиторами транскрипции. С другой стороны, существуют небольшие молекулы, которые образуют комплекс с репрессорами и снимают их блокирующее действие на транскрипцию. Иными словами, они ингибируют ингибиторы. Так, у бактерий в норме отсутствуют ферменты, катализирующие расщепление некоторых сахаров; однако если один из этих сахаров появляется в среде, он образует комплекс с репрессором, ингибирование снимается и запускается синтез соответствующего фермента. Ферменты, синтез которых индуцируется собственными субстратами, называются индуцибельными. В ряде случаев, наоборот, репрессорный белок не блокирует транскрипцию мРНК, если он не связан с определенной молекулой. У бактерий некоторые ферменты, участвующие в синтезе определенных аминокислот, образуются только в отсутствие этих аминокислот, т.е. бактерии производят данные ферменты лишь по мере надобности. Если добавить в среду соответствующую аминокислоту, она образует комплекс с репрессором и активирует его, а тем самым ингибирует транскрипцию соответствующих генов. Уже образовавшаяся мРНК вскоре расщепляется, и синтез ферментов останавливается. Такие ферменты являются отрицально индуцибельными.
Поскольку репрессорные белки сами кодируются генами, работа которых, в свою очередь, может регулироваться другими генами, а синтез малых молекул-индукторов и гормонов также в конечном счете регулируется генами, механизмы регуляции генной активности могут быть очень сложными.
Список литературы
Ичас М. Биологический код. М., 1971
Шабарова З.А., Богданов А.А. Химия нуклеиновых кислот и их компонентов, М., 1978
Зенгер В. Принципы структурной организации нуклеиновых кислот. М., 1987
www.referatmix.ru
Содержание
1. Общие сведения о нуклеиновых кислотах и история их открытия
Нуклеиновые кислоты, биополимеры, состоящие из остатков фосфорной кислоты, сахаров и азотистых оснований (пуринов и пиримидинов).
Нуклеиновые кислоты играют главную роль в передаче наследственных признаков (генетической информации) и управлении процессом биосинтеза белка.
Открытие нуклеиновых кислот связано с именем молодого врача из города Базеля (Швейцария) Фридриха Мишера. После окончания медицинского факультета Мишер был послан для усовершенствования и работы над диссертацией в Тюбинген (Германия) в физиолого-химическую лабораторию, возглавляемую Ф. Гоппе-Зейлером. Тюбингенская лаборатория в то время была известна ученому миру. Пройдя практику по органической химии, Мишер приступил к работе в биохимической лаборатории. Ему было поручено заняться изучением химического состава гноя. Молодой ученый не возражал против предложенной темы, так как считал лейкоциты, присутствующие в гное, одними из самых простых клеток.
Путём многочисленных опытов он получил из гнойных клеток вещество ядерного происхождения. Мишер был уверен именно в ядерном его источнике. Поэтому он начал более тщательное выделение ядер. В то время еще никто в биохимических лабораториях не пытался выделить ядра или какие-либо другие субклеточные компоненты, так что и здесь он был пионером.
Продолжив дальше очищать ядро от других клеточных фрагментов, он получил странное вещество. Оно не разлагалось протеолитическими ферментами, значит, не являлось белком. Отсутствие растворимости в горячем спирте указывало на то, что это вещество не являлось и фосфолипидом. По-видимому, оно относилось к новому классу биохимических соединений.
Но Мишер с большой горячностью настаивал на точности своих результатов и добивался разрешения опубликовать их в печати. Тогда Гоппе-Зейлер решил проверить данные Мишера лично. Он и два его ассистента (одним из них был русский химик Любавин) в течение года шаг за шагом прошли все этапы аналитической работы Мишера и полностью подтвердили его данные, выделив нуклеин из клеток крови и из дрожжей.
В 1871 г. работа Мишера вместе с подтверждающими ее контрольными работами Гоппе-Зейлера и его ассистентов увидела свет. Существование нуклеина как специфического ядерного вещества стало научным фактом. Вскоре методика Мишера была применена для выделения нуклеина из различных тканей.
В 1879 немецкий химик К. А. Коссель открыл в нуклеине соединение желтого цвета, которое оказалось гуанином, ранее выделенным из перуанского гуано — помета птиц, ценного азотного удобрения. Впоследствии он же выделил тимин клеток вилочковой железы, или тимуса, быка (отсюда название), цитозин (от греч. cytos — клетка) и аденин (от греч. aden — железа). Русский химик Ф. Левен установил, что, кроме тетрады аденин, гуанин, тимин и цитозин, нуклеин содержит еще и фосфорную кислоту и сахар дезоксирибозу. Термин «нуклеиновые кислоты» был предложен в 1889: нуклеиновыми они были названы потому, что впервые были открыты в ядрах клеток, а кислотами — из-за наличия в их составе остатков фосфорной кислоты. Позже было показано, что нуклеиновые кислоты построены из большого числа нуклеотидов (от нескольких десятков до сотен миллионов). В состав каждого нуклеотида входит азотистое основание, углевод (пентоза) и фосфорная кислота.
Сам же термин «нуклеиновые кислоты» был предложен в 1889 году: нуклеиновыми они были названы потому, что впервые были открыты в ядрах клеток, а кислотами — из-за наличия в их составе остатков фосфорной кислоты. Позже было показано, что нуклеиновые кислоты построены из большого числа нуклеотидов (от нескольких десятков до сотен миллионов). В состав каждого нуклеотида входит азотистое основание, углевод (пентоза) и фосфорная кислота.
Впоследствии было установлено, что существует два типа нуклеиновых кислот: рибонуклеиновая (РНК) и дезоксирибонуклеиновая (ДНК), однако их функции долго оставались неизвестными.
В 1928 английский бактериолог Ф. Гриффит обнаружил, что убитые патогенные пневмококки могут изменять генетические свойства живых непатогенных пневмококков, превращая последние в патогенные. В 1945 микробиолог О. Эвери из Рокфеллеровского института в Нью-Йорке сделал важное открытие: он показал, что способность к генетической трансформации обусловлена переносом ДНК из одной клетки в другую, а следовательно, генетический материал представляет собой ДНК. В 1940—1950 Дж. Бидл и Э. Тейтум из Станфордского университета (шт. Калифорния) обнаружили, что синтез белков, в частности ферментов, контролируется специфическими генами. В 1942 Т. Касперсон в Швеции и Ж. Браше в Бельгии открыли, что нуклеиновых кислот особенно много в клетках, активно синтезирующих белки. Все эти данные наводили на мысль, что носителем генетического материала являются нуклеиновые кислоты и что они каким-то образом участвуют в синтезе белков. Однако в то время многие полагали, что молекулы нуклеиновых кислот, несмотря на их большую длину, имеют слишком простую периодически повторяющуюся структуру, чтобы нести достаточно информации и служить основой генетической информации. Но в конце 1940-х годов Э. Чаргафф в США и Дж. Уайатт в Канаде, используя метод распределительной хроматографии на бумаге, показали, что структура ДНК не столь проста и эта молекула может служить носителем генетического материала.
Структура ДНК была установлена в 1953 М. Уилкинсом, Дж. Уотсоном и Ф. Криком в Англии. Это фундаментальное открытие позволило понять, как происходит удвоение (репликация) нуклеиновых кислот. Вскоре после этого американские исследователи А. Даунс и Дж. Гамов предположили, что структура белков каким-то образом закодирована в нуклеиновых кислотах, а к 1965 эта гипотеза была подтверждена многими исследователями: Ф. Криком в Англии, М. Ниренбергом и С. Очоа в США, Х. Кораной в Индии. Все эти открытия, результат столетнего изучения нуклеиновых кислот, произвели подлинную революцию в биологии. Они позволили объяснить феномен жизни в рамках взаимодействия между атомами и молекулами.
нуклеиновая кислота нуклеотид структура
2. Нахождение нуклеиновых кислот в природе
Нуклеиновые кислоты в природе встречаются во всех живых клетках. Живые клетки, за исключением сперматозоидов, в норме содержат значительно больше рибонуклеиновой, чем дезоксирибонуклеиновой кислоты. На методы выделения дезоксирибонуклеиновых кислот оказало большое влияние то обстоятельство, что, тогда как рибонуклеопротеиды и рибонуклеиновые кислоты растворимы в разбавленном (0,15 М) растворе хлористого натрия, дезоксирибонуклеопротеидные комплексы фактически в нем нерастворимы.
Поэтому гомогенизированный орган или организм тщательно промывают разбавленным солевым раствором, из остатка с помощью крепкого солевого раствора экстрагируют дезоксирибонуклеиновую кислоту, которую осаждают затем добавлением этанола.
В клетках эукариот (например, животных или растений) ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органоидах (митохондриях и пластидах). В клетках прокариотических организмов (бактерий и архей) кольцевая или линейная молекула ДНК, так называемый нуклеотид, прикреплена изнутри к клеточной мембране. У них и у низших эукариот (например, дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами. Кроме того, одно — или двухцепочечные молекулы ДНК могут образовывать геном ДНК-содержащих вирусов.
3. Строение нуклеиновых кислот
Нуклеиновые кислоты представляют собой биополимеры. Их макромолекулы состоят из не однократно повторяющихся звеньев, которые представлены нуклеотидами. И их логично назвали полинуклеотидами. Одной из главных характеристик нуклеиновых кислот является их нуклеотидный состав. В состав нуклеотида (структурного звена нуклеиновых кислот) входят три составные части:
1) Азотистое основание (нуклеиновое основание). Так в химии НК называют входящие в их состав гетероциклические соединения, среди которых различают структуры пиримидинового и пуринового рядов. В нуклеиновых кислотах содержатся основания 4-х разных видов: два из них относятся к классу пуринов и два — к классу пиримидинов. Азот, содержащийся в кольцах, придает молекулам основные свойства. В качестве заместителей в гетероциклическом ядре они содержат либо оксо — (урацил, тимин), либо аминогруппу (аденин), либо одновременно обе эти группы (цитозин, гуанин). Для них принято буквенное обозначение, составленное из первых букв их латинских названий.
НК различаются входящими в них гетероциклическими основаниями: урацил входит только в РНК, а тимин — в ДНК.
2) Моносахарид — D-рибоза или 2-дезокси-D-рибоза. Сахар, входящий в состав нуклеотида, содержит пять углеродных атомов, т. е. представляет собой пентозу. В зависимости от вида пентозы, присутствующей в нуклеотиде, различают два вида нуклеиновых кислот — рибонуклеиновые кислоты (РНК), которые содержат рибозу, и дезоксирибонуклеиновые кислоты (ДНК), содержащие дезоксирибозу.
3) Остаток фосфорной кислоты. Нуклеиновые кислоты являются кислотами потому, что в их состав входит остаток фосфорной кислоты.
Нуклеозиды — это гетероциклические соединения, образованные N-гликозидной связью между азотистым основанием и моносахаридом (рибозой или дезоксирибозой)
В зависимости от природы углеводного остатка различают рибонуклеозиды и дезоксирибонуклеозиды. Более часто употребляются названия, производимые от тривиального названия соответствующего нуклеинового основания с суффиксами — идин у пиримидиновых и — озин у пуриновых нуклеозидов.
Являясь N-гликозидами, нуклеозиды устойчивы к гидролизу в слабощелочной среде, но легко расщепляются в кислой. Пуриновые нуклеозиды гидролизуются легко, пиримидиновые — труднее.
Антибиотики-нуклеозиды.
В клетках в свободном состоянии содержатся некоторые нуклеозиды, не являющиеся компонентами нуклеиновых кислот. Эти нуклеозиды обладают антибиотической активностью и приобретают все большее значение при лечении злокачественных новообразований. Известно несколько десятков таких нуклеозидов, выделенных из микроорганизмов, а также из растительных и животных тканей.
Нуклеозиды-антибиотики отличаются от обычных нуклеозидов некоторыми деталями строения либо углеводной части, либо гетероциклического основания. По-видимому, это позволяет им выступать в роли антиметаболитов. Нуклеозидные антибиотики пиримидинового ряда часто подобны цитидину, пуринового ряда — аденозину.
Нуклеотиды.
Нуклеотимды — фосфорные эфиры нуклеозидов, нуклеозидфосфаты.
В природе наиболее распространены нуклеотиды, являющиеся в-N-гликозидами пуринов или пиримидинов и пентоз — D-рибозы или D-2-дезоксирибозы. В зависимости от структуры пентозы различают рибонуклеотиды и дезоксирибонуклеотиды, которые являются мономерами молекул сложных биологических полимеров (полинуклеотидов) — соответственно РНК или ДНК.
Таким образом, структура нуклеотида образована двумя связями: N-гликозидной связью между азотистым основанием и моносахаридом (рибоза или дезоксирибоза) и сложноэфирной связью между моносахаридом и остатком фосфорной кислоты.
3. Номенклатура нуклеотидов
Для нуклеотидов используют два вида названий. Одно включает наименование нуклеозида с указанием положения в нем фосфатного остатка, например, аденозин-3'-фосфат, уридин-5'-фосфат; другое строится с добавлением сочетания — иловая кислота к названию остатка пиримидинового основания, например, 5'-уридиловая кислота, или пуринового основания, например 3'-адениловая кислота.
Используя принятый для нуклеозидов однобуквенный код, 5'-фосфаты записывают с добавлением латинской буквы «р» перед символом нуклеозида, 3'-фосфаты — после символа нуклеозида. Аденозин-5'-фосфат обозначается рА, аденозин-3'-фосфат — Ар и т. п. Эти сокращенные обозначения используют для записи последовательности нуклеотидных остатков в нуклеиновых кислотах.
По отношению к свободным нуклеотидам в биохимической литературе широко используют их названия как монофосфатов с отражением этого признака в сокращенном коде, например АМР (или АМФ) для аденозин-5'-фосфата и т. д.
4. Структура нуклеиновых кислот
4.1 Первичная структура
В полинуклеотидных цепях нуклеотидные звенья связаны через фосфатную группу. Фосфатная группа образует две сложноэфирные связи: с С-3' предыдущего и с С-5' последующего нуклеотидных звеньев. Каркас цепи состоит из чередующихся пентозных и фосфатных остатков, а гетероциклические основания являются «боковыми» группами, присоединенными к пентозным остаткам. Нуклеотид со свободной 5'-ОН группой называют 5'-концевым, а нуклеотид со свободной З'-ОН группой — З'-концевым.
На рисунке приведено строение произвольного участка цепи ДНК, включающего четыре нуклеиновых основания. Легко представить, какое множество сочетаний можно получить путем варьирования последовательности четырех нуклеотидных остатков. Принцип построения цепи РНК такой же, как и у ДНК, с двумя исключениями: пентозным остатком в РНК служит D-рибоза, а в наборе гетероциклических оснований используется не тимин, а урацил.
Первичная структура нуклеиновых кислот определяется последовательностью нуклеотидных звеньев, связанных ковалентными связями в непрерывную цепь полинуклеотида.
Для удобства записи первичной структуры существует несколько способов сокращений. Один из них заключается в использовании ранее приведенных сокращенных названий нуклеозидов. Например, показанный на рисунке фрагмент цепи ДНК может быть записан как d (ApCpGpTp.) или d (A-C-G-T.). Часто букву d опускают, если очевидно, что речь идет о ДНК.
Важной характеристикой нуклеиновых кислот служит нуклеотидный состав, т. е. набор и количественное отношение нуклеотидных компонентов. Нуклеотидный состав устанавливают, как правило, путем исследования продуктов гидролитического расщепления нуклеиновых кислот.
ДНК и РНК различаются поведением в условиях щелочного и кислотного гидролиза. ДНК устойчивы к гидролизу в щелочной среде. РНК легко гидролизуются в мягких условиях в щелочной среде до нуклеотидов, которые, в свою очередь, способны в щелочной среде отщеплять остаток фосфорной кислоты с образованием нуклеозидов. Нуклеозиды в кислой среде гидролизуются до гетероциклических оснований и углеводов.
4.2 Вторичная структура ДНК
Структура двойной спирали ДНК была предложена Френсисом Криком и Джеймсом Уотсоном в 1953 году на основании рентгеноструктурных данных, полученных Морисом Уилкинсом и Розалинд Франклин, и «правил Чаргаффа», согласно которым в каждой молекуле ДНК соблюдаются строгие соотношения, связывающие между собой количество азотистых оснований разных типов. К 1953 году было известно, что ДНК состоит из 4 нуклеотидов, а каждый из них — из одного азотистого основания, 5-углеродного сахара дезоксирибозы и остатка фосфорной кислоты. Не было понятно, как эти части соединяются в молекулы ДНК.
Они создали модели нуклеотидов и попытались построить из них двойную спираль. Это получилось только после того, когда они догадались перевернуть одну из цепей, так что те шли навстречу друг другу — антипараллельно. Такая модель показывает, как способность ДНК к воспроизводству своей структуры (свойство, лежащее в основе передачи признаков при делении клеток) объясняется ее химическим строением: при копировании ДНК порядок чередования нуклеотидов в одной цепи однозначно задает порядок чередования нуклеотидов в другой цепи, строящейся на первой, как на матрице.
Позже предложенная Уотсоном и Криком модель строения ДНК была доказана, а их работа отмечена Нобелевской премией по физиологии и медицине 1962 г.
Таким образом, двойная спираль правозакрученная, полинуклеотидные цепи в ней антипараллельны, т. е. если одна из них ориентирована в направлении 3'> 5', то вторая — в направлении 5'> 3'. Поэтому на каждом из концов молекулы ДНК расположены 5'-конец одной цепи и 3'-конец другой цепи.
Все основания цепей ДНК расположены внутри двойной спирали, а пентозофосфатный остов — снаружи. Полинуклеотидные цепи удерживаются относительно друг друга за счёт водородных связей между комплементарными пуриновыми и пиримидиновыми азотистыми основаниями, А и Т (две связи) и между G и С (три связи). При таком сочетании каждая пара содержит по три кольца, поэтому общий размер этих пар оснований одинаков по всей длине молекулы. Водородные связи при других сочетаниях оснований в паре возможны, но они значительно слабее. Последовательность нуклеотидов одной цепи полностью комплементарна последовательности нуклеотидов второй цепи. Поэтому, согласно правилу Чаргаффа (Эрвин Чаргафф в 1951 г. установил закономерности в соотношении пуриновых и пиримидиновых оснований в молекуле ДНК), число пуриновых оснований (А + G) равно числу пиримидиновых оснований (Т + С).
Комплементарые основания уложены в стопку в сердцевине спирали. Между основаниями двухцепочечной молекулы в стопке возникают гидрофобные взаимодействия, стабилизирующие двойную спираль.
Такая структура исключает контакт азотистых остатков с водой, но стопка оснований не может быть абсолютно вертикальной. Пары оснований слегка смещены относительно друг друга. В образованной структуре различают две бороздки — большую, шириной 2,2 нм, и малую, шириной 1,2 нм. Азотистые основания в области большой и малой бороздок взаимодействуют со специфическими белками, участвующими в организации структуры хроматина.
Т.е. под вторичной структурой понимают пространственную организацию полинуклеотидной цепи. Согласно модели Уотсона-Крика молекула ДНК состоит из двух полинуклеотидных цепей, правозакрученных вокруг общей оси с образованием двойной спирали. Пуриновые и пиримидиновые основания направлены внутрь спирали. Между пуриновым основанием одной цепи и пиримидиновым основанием другой цепи возникают водородные связи. Эти основания составляют комплементарные пары.
Водородные связи образуются между аминогруппой одного основания и карбонильной группой другого — NH. O=C-, а также между амидным и иминным атомами азота — NH. N — Например, как показано ниже, между аденином и тимином образуются две водородные связи, и эти основания составляют комплементарную пару, т. е. аденину в одной цепи будет соответствовать тимин в другой цепи. Другую пару комплементарных оснований составляют гуанин и цитозин, между которыми возникают три водородные связи.
Водородные связи между комплементарными основаниями — один из видов взаимодействий, стабилизирующих двойную спираль. Две цепи ДНК, образующие двойную спираль, не идентичны, но комплементарны между собой. Это означает, что первичная структура, т. е. нуклеотидная последовательность, одной цепи предопределяет первичную структуру второй цепи.
5. Функции нуклеиновых кислот
5.1 Функции ДНК
1. Важнейшая биологическая функция ДНК — генетическая, т. е. ДНК является носителем генетической информации.
2. ДНК способна передавать генетическую информацию в ряду поколений посредством и-РНК.
3. Регулирует процессы биосинтеза белка,
В клетках эукариот ДНК располагается главным образом в ядре в виде набора хромосом. Бактериальная (прокариоты) ДНК обычно представлена одной кольцевой молекулой ДНК, расположенной в неправильной формы образовании в цитоплазме, называемым нуклеотидом. Генетическая информация генома состоит из генов. Ген — единица передачи наследственной информации и участок ДНК, который влияет на определённую характеристику организма. Ген содержит открытую рамку считывания, которая транскрибируется, а также регуляторные последовательности, например, промотор и энхансер, которые контролируют экспрессию открытых рамок считывания.
Функции хранения и передачи информации обеспечиваются биосинтезом белка, посредством и-РНК и т-РНК.
5.2 Функции РНК
Существуют три типа РНК, каждый из которых выполняет свою особую роль в синтезе белка.
1. Матричная РНК переносит генетический код из ядра в цитоплазму, определяя таким образом синтез разнообразных белков.
2. Транспортная РНК переносит активированные аминокислоты к рибосомам для синтеза полипептидных молекул.
3. Рибосомная РНК в комплексе примерно с 75 разными белками формирует рибосомы — клеточные органеллы, на которых происходит сборка полипептидных молекул.
Матричная РНК (информационная) представляет собой длинную одноцепочечную молекулу, присутствующую в цитоплазме. Эта молекула РНК содержит от нескольких сотен до нескольких тысяч нуклеотидов РНК, образующих кодоны, строго комплементарные триплетам ДНК. Она переносит генетическую информацию через мембрану ядра к месту синтеза белка на рибосоме. Переписывание (перекодирование) информации с ДНК на РНК происходит посредством транскрипции.
Еще один тип РНК, играющий важнейшую роль в синтезе белка, называют транспортной РНК, поскольку он транспортирует аминокислоты к строящейся молекуле белка. Каждая транспортная РНК специфически связывается только с одной из 20 аминокислот, составляющих белковые молекулы. Транспортные РНК действуют как переносчики специфических аминокислот, доставляя их к рибосомам, на которых происходит сборка полипептидных молекул. Каждая специфическая транспортная РНК распознает «свой» кодон матричной РНК, прикрепившейся к рибосоме, и доставляет соответствующую аминокислоту на соответствующую позицию в синтезируемой полипептидной цепи. Цепь транспортной РНК гораздо короче матричной РНК, содержит всего около 80 нуклеотидов и упакована в форме клеверного листа. На одном конце транспортной РНК всегда находится аденозинмонофосфат (АМФ), к которому через гидроксильную группу рибозы прикрепляется транспортируемая аминокислота. Транспортные РНК служат для прикрепления специфических аминокислот к строящейся полипептидной молекуле, поэтому необходимо, чтобы каждая транспортная РНК обладала специфичностью и в отношении соответствующих кодонов матричной РНК. Код, посредством которого транспортная РНК распознает соответствующий кодон на матричной РНК, также является триплетом и его называют антикодоном. Антикодон располагается примерно посередине молекулы транспортной РНК. Во время синтеза белка азотистые основания антикодона транспортной РНК прикрепляются с помощью водородных связей к азотистым основаниям кодона матричной РНК. Таким образом, на матричной РНК выстраиваются в определенном порядке одна за другой различные аминокислоты, формируя соответствующую аминокислотную последовательность синтезируемого белка.
Рибосомные РНК содержат 3000−5000 нуклеотидов; молекулярная масса — 1 000 000−1 500 000. На долю рРНК приходится 80−85% от общего содержания РНК в клетке. В комплексе с рибосомными белками рРНК образует рибосомы — органоиды, осуществляющие синтез белка. В эукариотических клетках синтез рРНК происходит в ядрышках.
Функции рРНК:
1) необходимый структурный компонент рибосом и, таким образом, обеспечение функционирования рибосом;
2) обеспечение взаимодействия рибосомы и тРНК;
3) первоначальное связывание рибосомы и кодона-инициатора иРНК и определение рамки считывания,
4) формирование активного центра рибосомы.
Список использованной литературы
1. Ленинджер А. Основы биохимии: Учебник. — М.: Мир, 1985.
2. Тюкавкина Н. А., Бауков Ю. И. Биоорганическая химия: Учебник. — М.: Медицина, 1991.
3. Остерман Л. А. Методы исследования белков и нуклеиновых кислот. — М.: Наука, 1981.
4. Шабарова 3.А., Богданов А. А., Химия нуклеиновых кислот и их компонентов, М., 1978.
5. Зенгер В. Принципы структурной организации нуклеиновых кислот. М., 1987.
6. Сайт: http: //ru. wikipedia. org/wiki/%C2%F2%EE%F0%E8%F7%ED%E0%FF_%F1%F2%F0%F3%EA%F2%F3%F0%E0
7. Сайт: http: //slovari. yandex. ru/нуклеиновые%20кислоты/БСЭ/Нуклеиновые%20кислоты/
Показать Свернутьwestud.ru
Нуклеиновые кислоты, биополимеры, состоящие из остатков фосфорной кислоты, сахаров и азотистых оснований (пуринов и пиримидинов). Имеют фундаментальное биологическое значение, поскольку содержат в закодированном виде всю генетическую информацию любого живого организма, от человека до бактерий и вирусов, передаваемую от одного поколения другому.
Нуклеиновые кислоты были впервые выделены из клеток гноя человека и спермы лосося швейцарским врачом и биохимиком Ф.Мишером между 1869 и 1871. Впоследствии было установлено, что существует два типа нуклеиновых кислот: рибонуклеиновая (РНК) и дезоксирибонуклеиновая (ДНК), однако их функции долго оставались неизвестными.
В 1928 английский бактериолог Ф.Гриффит обнаружил, что убитые патогенные пневмококки могут изменять генетические свойства живых непатогенных пневмококков, превращая последние в патогенные. В 1945 микробиолог О.Эвери из Рокфеллеровского института в Нью-Йорке сделал важное открытие: он показал, что способность к генетической трансформации обусловлена переносом ДНК из одной клетки в другую, а следовательно, генетический материал представляет собой ДНК. В 1940–1950 Дж.Бидл и Э.Тейтум из Станфордского университета (шт. Калифорния) обнаружили, что синтез белков, в частности ферментов, контролируется специфическими генами. В 1942 Т.Касперсон в Швеции и Ж.Браше в Бельгии открыли, что нуклеиновых кислот особенно много в клетках, активно синтезирующих белки. Все эти данные наводили на мысль, что генетический материал – это нуклеиновая кислота и что она как-то участвует в синтезе белков. Однако в то время многие полагали, что молекулы нуклеиновых кислот, несмотря на их большую длину, имеют слишком простую периодически повторяющуюся структуру, чтобы нести достаточно информации и служить генетическим материалом. Но в конце 1940-х годов Э.Чаргафф в США и Дж.Уайатт в Канаде, используя метод распределительной хроматографии на бумаге, показали, что структура ДНК не столь проста и эта молекула может служить носителем генетической информации.
Структура ДНК была установлена в 1953 М.Уилкинсом, Дж.Уотсоном и Ф.Криком в Англии. Это фундаментальное открытие позволило понять, как происходит удвоение (репликация) нуклеиновых кислот. Вскоре после этого американские исследователи А.Даунс и Дж.Гамов предположили, что структура белков каким-то образом закодирована в нуклеиновых кислотах, а к 1965 эта гипотеза была подтверждена многими исследователями: Ф.Криком в Англии, М.Ниренбергом и С.Очоа в США, Х.Кораной в Индии. Все эти открытия, результат столетнего изучения нуклеиновых кислот, произвели подлинную революцию в биологии. Они позволили объяснить феномен жизни в рамках взаимодействия между атомами и молекулами.
Типы и распространение. Как мы уже говорили, есть два типа нуклеиновых кислот: ДНК и РНК. ДНК присутствует в ядрах всех растительных и животных клеток, где она находится в комплексе с белками и является составной частью хромосом. У особей каждого конкретного вида содержание ядерной ДНК обычно одинаково во всех клетках, кроме гамет (яйцеклеток и сперматозоидов), где ДНК вдвое меньше. Таким образом, количество клеточной ДНК видоспецифично. ДНК найдена и вне ядра: в митохондриях («энергетических станциях» клеток) и в хлоропластах (частицах, где в растительных клетках идет фотосинтез). Эти субклеточные частицы обладают некоторой генетической автономией.
Бактерии и цианобактерии (сине-зеленые водоросли) содержат вместо хромосом одну или две крупные молекулы ДНК, связанные с небольшим количеством белка, и часто – молекулы ДНК меньшего размера, называемые плазмидами. Плазмиды несут полезную генетическую информацию, например содержат гены устойчивости к антибиотикам, но для жизни самой клетки они несущественны.
Некоторое количество РНК присутствует в клеточном ядре, основная же ее масса находится в цитоплазме – жидком содержимом клетки. Бльшую ее часть составляет рибосомная РНК (рРНК). Рибосомы – это мельчайшие тельца, на которых идет синтез белка. Небольшое количество РНК представлено транспортной РНК (тРНК), которая также участвует в белковом синтезе. Однако оба этих класса РНК не несут информации о структуре белков – такая информация заключена в матричной, или информационной, РНК (мРНК), на долю которой приходится лишь небольшая часть суммарной клеточной РНК.
Генетический материал вирусов представлен либо ДНК, либо РНК, но никогда обеими одновременно.
Общие свойства
Молекулы нуклеиновых кислот содержат множество отрицательно заряженных фосфатных групп и образуют комплексы с ионами металлов; их калиевая и натриевая соли хорошо растворимы в воде. Концентрированные растворы нуклеиновых кислот очень вязкие и слегка опалесцируют, а в твердом виде эти вещества белые. Нуклеиновые кислоты сильно поглощают ультрафиолетовый свет, и это свойство лежит в основе определения их концентрации. С этим же свойством связан и мутагенный эффект ультрафиолетового света.
Длинные молекулы ДНК хрупки и легко ломаются, например при продавливании раствора через шприц. Поэтому работа с высокомолекулярными ДНК требует особой осторожности.
Химическая структура. Нуклеиновые кислоты это длинные цепочки, состоящие из четырех многократно повторяющихся единиц (нуклеотидов). Их структуру можно представить следующим образом:
Символ Ф обозначает фосфатную группу. Чередующиеся остатки сахара и фосфорной кислоты образуют сахарофосфатный остов молекулы, одинаковый у всех ДНК, а огромное их разнообразие обусловливается тем, что четыре азотистых основания могут располагаться вдоль цепи в самой разной последовательности.
Сахаром в нуклеиновых кислотах является пентоза; четыре из пяти ее углеродных атомов вместе с одним атомом кислорода образуют кольцо. Атомы углерода пентозы обозначают номерами от 1 до 5. В РНК сахар представлен рибозой, а в ДНК дезоксирибозой, содержащей на один атом кислорода меньше. Фрагменты полинуклеотидных цепей ДНК и РНК показаны на рисунке.
Поскольку фосфатные группы присоединены к сахару асимметрично, в положениях 3 и 5, молекула нуклеиновой кислоты имеет определенное направление. Сложноэфирные связи между мономерными единицами нуклеиновых кислот чувствительны к гидролитическому расщеплению (ферментативному или химическому), которое приводит к высвобождению отдельных компонентов в виде небольших молекул.
Азотистые основания – это плоские гетероциклические соединения. Они присоединены к пентозному кольцу по положению 1. Более крупные основания имеют два кольца и называются пуринами: это аденин (А) и гуанин (Г). Основания, меньшие по размерам, имеют одно кольцо и называются пиримидинами: это цитозин (Ц), тимин (Т) и урацил (У). В ДНК входят основания А, Г, Т и Ц, в РНК вместо Т присутствует У. Последний отличается от тимина тем, что у него отсутствует метильная группа (Ch4). Урацил встречается в ДНК некоторых вирусов, где он выполняет ту же функцию, что и тимин.
Трехмерная структура. Важной особенностью нуклеиновых кислот является регулярность пространственного расположения составляющих их атомов, установленная рентгеноструктурным методом. Молекула ДНК состоит из двух противоположно направленных цепей (иногда содержащих миллионы нуклеотидов), удерживаемых вместе водородными связями между основаниями:
Водородные связи, соединяющие основания противоположных цепей, относятся к категории слабых, но благодаря своей многочисленности в молекуле ДНК они прочно стабилизируют ее структуру. Однако если раствор ДНК нагреть примерно до 60 С, эти связи рвутся и цепи расходятся – происходит денатурация ДНК (плавление).
Обе цепи ДНК закручены по спирали относительно воображаемой оси, как будто они навиты на цилиндр. Эта структура называется двойной спиралью. На каждый виток спирали приходится десять пар оснований.
Правило комплементарности. Уотсон и Крик показали, что образование водородных связей и регулярной двойной спирали возможно только тогда, когда более крупное пуриновое основание аденин (А) в одной цепи имеет своим партнером в другой цепи меньшее по размерам пиримидиновое основание тимин (Т), а гуанин (Г) связан с цитозином (Ц). Эту закономерность можно представить следующим образом:
Соответствие АТ и ГЦ называют правилом комплементарности, а сами цепи комплементарными. Согласно этому правилу, содержание аденина в ДНК всегда равно содержанию тимина, а количество гуанина – количеству цитозина. Следует отметить, что две цепи ДНК, различаясь химически, несут одинаковую информацию, поскольку вследствие комплементарности одна цепь однозначно задает другую.
Структура РНК менее упорядочена. Обычно это одноцепочечная молекула, хотя РНК некоторых вирусов состоит из двух цепей. Но даже такая РНК более гибка, чем ДНК. Некоторые участки в молекуле РНК взаимно комплементарны и при изгибании цепи спариваются, образуя двухцепочечные структуры (шпильки). В первую очередь это относится к транспортным РНК (тРНК). Некоторые основания в тРНК подвергаются модификации уже после синтеза молекулы. Например, иногда происходит присоединение к ним метильных групп.
Функция нуклеиновых кислот
Одна из основных функций нуклеиновых кислот состоит в детерминации синтеза белков. Информация о структуре белков, закодированная в нуклеотидной последовательности ДНК, должна передаваться от одного поколения к другому, и поэтому необходимо ее безошибочное копирование, т.е. синтез точно такой же же молекулы ДНК (репликация).
Репликация и транскрипция. С химической точки зрения синтез нуклеиновой кислоты – это полимеризация, т.е. последовательное присоединение строительных блоков. Такими блоками служат нуклеозидтрифосфаты; реакцию можно представить следующим образом:
Энергия, необходимая для синтеза, высвобождается при отщеплении пирофосфата, а катализируют реакцию особые ферменты – ДНК-полимеразы.
В результате такого синтетического процесса мы получили бы полимер со случайной последовательностью оснований. Однако большинство полимераз работает только в присутствии уже существующей нуклеиновой кислоты –матрицы, диктующей, какой именно нуклеотид присоединится к концу цепи. Этот нуклеотид должен быть комплементарен соответствующему нуклеотиду матрицы, так что новая цепь оказывается комплементарной исходной. Используя затем комплементарную цепь в качестве матрицы, мы получим точную копию оригинала.
ДНК состоит из двух взаимно комплементарных цепей. В ходе репликации они расходятся, и каждая из них служит матрицей для синтеза новой цепи:
Так образуются две новые двойные спирали с той же последовательностью оснований, что и у исходной ДНК. Иногда в процессе репликации происходит «сбой», и возникают мутации.
В результате транскрипции ДНК образуются клеточные РНК (мРНК, рРНК и тРНК):
Они комплементарны одной из цепей ДНК и являются копией другой цепи, за исключением того, что место тимина у них занимает урацил. Таким способом можно получить множество РНК-копий одной из цепей ДНК.
В нормальной клетке передача информации осуществляется только в направлении ДНК ДНК и ДНК РНК. Однако в клетках, инфицированных вирусом, возможны и другие процессы: РНК РНК и РНК ДНК. Генетический материал многих вирусов представлен молекулой РНК, обычно одноцепочечной. Проникнув в клетку-хозяина, эта РНК реплицируется с образованием комплементарной молекулы, на которой, в свою очередь, синтезируется множество копий исходной вирусной РНК:
Вирусная РНК может транскрибироваться ферментом обратной транскриптазой в ДНК, которая иногда включается в хромосомную ДНК клетки-хозяина. Теперь эта ДНК несет вирусные гены, и после транскрипции в клетке может появиться вирусная РНК. Таким образом, спустя длительное время, в течение которого никакого вируса в клетке не обнаруживается, он снова в ней появится без повторного заражения. Вирусы, генетический материал которых включается в хромосому клетки-хозяина, часто являются причиной рака.
Трансляция нуклеиновых кислот в белки. Генетическая информация, закодированная в нуклеотидной последовательности ДНК, переводится не только на язык нуклеотидной последовательности РНК, но и на язык аминокислот – мономерных единиц белков.
Белковая молекула – это цепочка из аминокислот. Каждая аминокислота содержит кислую карбоксильную группу –COOH и оснвную аминогруппу
–Nh3. Карбоксильная группа одной аминокислоты связывается с аминогруппой другой, образуя амидную связь, и этот процесс продолжается, пока не образуется цепь, содержащая до 1000 аминокислот.
В белках присутствует 20 разных аминокислот, от последовательности которых зависят их природа и функции. Эта последовательность определяется нуклеотидной последовательностью соответствующего гена – участка ДНК, кодирующего данный белок. Однако сама ДНК не является матрицей при синтезе белка. Сначала она транскрибируется в ядре с образованием матричной РНК (мРНК), которая диффундирует в цитоплазму, и на ней как на матрице синтезируется белок. Процесс ускоряется благодаря тому, что на каждой молекуле мРНК может одновременно синтезироваться множество белковых молекул.
Репликация нуклеиновых кислот осуществляется благодаря образованию водородных связей между комплементарными основаниями исходной и дочерней цепей. Аминокислоты не образуют водородных связей с основаниями, так что прямое копирование матрицы невозможно. Они взаимодействуют с матрицей опосредованно, через «адапторные» нуклеиновые кислоты – небольшие молекулы транспортных РНК (тРНК), состоящие примерно из 80 оснований и способные связываться с мРНК.
Каждая тРНК содержит специфическую последовательность из трех оснований, антикодон, который комплементарен группе из трех оснований, кодону, в мРНК. Антикодоны взаимодействуют с кодонами по правилу комплементарности, примерно так же, как взаимодействуют две цепи ДНК. Таким образом, последовательность оснований в мРНК определяет порядок присоединения тРНК, несущих аминокислоты. Схематически перенос информации от ДНК к белку можно представить следующим образом:
Последовательность оснований в ДНК задает порядок следования аминокислот в белке, поскольку каждая аминокислота присоединяется специфическим ферментом только к определенным тРНК, а те, в свою очередь, – только к определенным кодонам в мРНК. Комплексы тРНК-аминокислота связываются с матрицей по одному в каждый данный момент времени. Ниже перечислены основные этапы белкового синтеза (см. также рисунок).
1. Ферменты, называемые аминоацил-тРНК-синтетазами, присоединяют аминокислоты к соответствующим тРНК. Таких ферментов 20, по одному для каждой аминокислоты.
2. Молекула мРНК присоединяется своим первым кодоном к небольшой частице, называемой рибосомой. Рибосомы состоят из примерно равных количеств рРНК и белка. Структура и функция рибосом весьма сложны, но главная их задача – облегчение взаимодействия мРНК и тРНК и ускорение полимеризации аминокислот, связанных с разными тРНК.
3. тРНК, нагруженная аминокислотой, связывается с соответствующим кодоном мРНК, которая, в свою очередь, контактирует с рибосомой. Образуется комплекс рибосома-мРНК-тРНК-аминокислота.
4. мРНК, подобно ленте на конвейере, продвигается по рибосоме на один кодон вперед.
5. Следующая тРНК, нагруженная аминокислотой, присоединяется ко второму кодону.
6. Первая и вторая аминокислоты связываются между собой.
7. Первая тРНК отсоединяется от комплекса, и теперь вторая тРНК несет две аминокислоты, связанные между собой.
8. мРНК снова продвигается на один кодон вперед, и все события повторяются, а растущая аминокислотная цепь удлиняется на одну аминокислоту. Процесс продолжается, пока не будет достигнут последний, «стоп»-кодон и последняя тРНК не отделится от готовой белковой цепи. В бактериальных клетках цепь из 100–200 аминокислот собирается за несколько секунд. В животных клетках этот процесс занимает около минуты.
Генетический код. Итак, каждая аминокислота в белке опосредованно детерминируется определенным кодоном (группой из 3 оснований) в мРНК и в конечном счете в ДНК. Поскольку в нуклеиновых кислотах имеется четыре вида оснований, число возможных кодонов составляет 444 = 64. Соответствие между кодонами и аминокислотами, которые они кодируют, называется генетическим или биологическим кодом. Это соответствие было установлено опытным путем: к разрушенным клеткам добавляли синтетические полинуклеотиды известного состава и смотрели, какие аминокислоты включаются в белки. Позднее появилась возможность прямо сравнить последовательности аминокислот в вирусных белках и оснований в вирусных нуклеиновых кислотах. Чрезвычайно интересно, что генетический код, за редкими исключениями, одинаков для всех организмов – от вирусов до человека. Одно из таких исключений составляют изменения в генетическом коде, используемом митохондриями. Митохондрии это небольшие автономные субклеточные частицы (органеллы), присутствующие во всех клетках, кроме бактерий и зрелых эритроцитов. Предполагают, что когда-то митохондрии были самостоятельными организмами; проникнув в клетки, они со временем стали их неотъемлемой частью, но сохранили некоторое количество собственной ДНК и синтезируют несколько митохондриальных белков.
| Аланин | Аргинин | Аспарагин | Аспарагиновая кислота | |
| ГЦУ | ЦГУ | ГАУ | ААУ | |
| ГЦЦ | ЦГЦ | ГАЦ | ААЦ | |
| ГЦА | ЦГА | |||
| ГЦГ | ЦГГ | |||
| АГА | ||||
| АГГ | ||||
| Валин | Гистидин | Глицин | Глутаминовая кислота | |
| ГУУ | ЦАУ | ГГУ | ЦАА | |
| ГУЦ | ЦАЦ | ГГЦ | ЦАГ | |
| ГУА | ГГА | |||
| ГУГ | ГГГ | |||
| Глутамин | Изолейцин | Лейцин | Лизин | |
| ГАА | АУУ | УУА | ААА | |
| ГАГ | АУЦ | УУГ | ААГ | |
| АУА | ЦУУ | |||
| ЦУЦ | ||||
| ЦУА | ||||
| ЦУГ | ||||
| Метионин | Пролин | Серин | Тирозин | |
| АУГ | ЦЦУ | АГУ | УАУ | |
| ЦЦЦ | АГЦ | УАЦ | ||
| ЦЦА | УЦА | |||
| ЦЦГ | УЦГ | |||
| УЦУ | ||||
| УЦЦ | ||||
| Треонин | Триптофан | Фенилаланин | Цистеин | Нет |
| АЦУ | УГГ | УУУ | УГУ | УАА |
| АЦЦ | УУЦ | УГЦ | УАГ | |
| АЦА | УГА | |||
| АЦГ | ||||
| ГЕНЕТИЧЕСКИЙ СЛОВАРЬ: указаны аминокислоты, встречающиеся в белках, и соответствующие им кодоны в мРНК. «Буквы» в кодонах записаны в направлении 5' 3'. В этом же направлении идут транскрипции нуклеиновых кислот и синтез белка на матрице. «Нет» означает, что кодон не кодирует никаких аминокислот; такие кодоны называются «бессмысленными». Генетический словарь одинаков для всех живых организмов – от вирусов до человека. |
Вообще говоря, каждой аминокислоте соответствует более одного кодона. Большинство кодонов, кодирующих одну и ту же аминокислоту, имеют два одинаковых первых основания, но в трех случаях (для лейцина, серина и аргинина) имеются два альтернативных набора первых дублетов в кодонах, соответствующих одной и той же аминокислоте. Природа основания в третьем положении не столь важна; одна и та же аминокислота глицин может кодироваться по-разному: ГГУ, ГГЦ, ГГА и ГГГ. Однако кодоны для двух разных аминокислот могут иметь два одинаковых первых основания, и тогда различие между ними будет определяться природой третьего основания – пурином или пиримидином. Так, гистидин кодируется триплетами ЦАУ и ЦАЦ, а глутамин ЦАА и ЦАГ. Три кодона, УАА, УАГ и УГА, не кодируют никаких аминокислот и называются «бессмысленными».
Одна молекула ДНК кодирует много белковых цепей. Каждый отрезок, кодирующий одну цепь, называют цистроном. Начало и конец цистрона, а также граница раздела между ними помечаются с помощью своего рода знаков химической пунктуации. По крайней мере у бактерий в начале цистрона находится метиониновый кодон АУГ. Логично предположить, что первой аминокислотой в белке всегда должен быть метионин, но часто несколько первых аминокислот отщепляются ферментативно после окончания синтеза белка. Конец белковой цепи помечается одним или несколькими «бессмысленными» кодонами.
У бактерий (прокариот) практически вся ДНК кодирует какие-либо белки или тРНК. Однако у высших форм (эукариот) значительная часть ДНК состоит из простых повторяющихся последовательностей и «молчащих» генов, которые не транскрибируются в РНК и поэтому не транслируются в белки. Кроме того, исходно синтезированная мРНК содержит участки, не детерминирующие никаких белковых последовательностей. Такие участки (интроны), расположенные между кодирующими участками (экзонами), перед началом синтеза белка удаляются специальными ферментами. Почему в ДНК существуют эти казалось бы бесполезные сегменты – неясно; возможно, они выполняют регуляторные функции.
У простейшей Tetrahymena РНК сама удаляет свои интроны и соединяет свободные концы цепей, действуя как фермент по отношению к себе самой. Это единственное известное исключение из правила, согласно которому нуклеиновые кислоты не обладают ферментативной активностью.
Транспортные РНК и супрессия. Смысл информации, содержащейся в ДНК, если переводить ее на язык аминокислот, определяется как самой ДНК, так и считывающим механизмом, т.е. зависит не только от того, какие кодоны есть в ДНК и в какой последовательности они расположены, но также и от того, какие именно аминокислоты (и к каким тРНК) присоединяют аминоацил-тРНК-синтетазы. Конечно, природа синтетаз и тРНК тоже определяется ДНК, и в этом смысле ДНК является первичным детерминантом белковой последовательности. Тем не менее суммарная детерминация представляет собой функцию всей системы, поскольку результат зависит от исходных компонентов. Если бы соответствие между тРНК и аминокислотами было другим, смысл кодонов тоже изменился бы.
Известно, что мутации в ДНК изменяют считывающий механизм и в результате меняют – пусть и незначительно – смысл кодонов. Так, в бактерии Escherichia coli глициновая тРНК обычно узнает в мРНК кодон ГГА; мутация в ДНК, с которой транскрибируется эта тРНК, изменяет антикодон глициновой тРНК таким образом, что теперь он узнает кодон АГА, соответствующий аргинину, и в белковой молекуле вместо аргинина появляется глицин. Это не обязательно имеет фатальные последствия, поскольку не все аргинины кодируются триплетом АГА и есть аргининовые тРНК, по-прежнему узнающие «свои» АГА. В результате измененными оказываются не все белковые молекулы. Иногда такие мутации, изменяющие антикодон, подавляют (супрессируют) мутации в кодоне. Например, если в результате мутации глициновый кодон ГГА превращается в АГА, он все же может прочитываться как глицин, если антикодон глициновой тРНК, в свою очередь, изменился так, что эта тРНК стала узнавать АГА. В этом случае вторая «ошибка» устраняет первую.
Мутации, приводящие к изменению антикодонов, могут иметь разные последствия, поскольку один и тот же кодон может узнаваться несколькими тРНК. Вообще говоря, узнавание осуществляется благодаря комплементарности оснований кодона и антикодона, однако одно из оснований кодона может модифицироваться таким образом, что антикодон будет узнавать даже неполностью комплементарный кодон. В результате одна и та же тРНК может взаимодействовать с несколькими разными кодонами, кодирующими одну и ту же аминокислоту. Этот феномен неполного соответствия кодона и антикодона был назван Ф.Криком «шатанием».
Регуляция активности генов. Для организма было бы катастрофой, если бы во всех его клетках одновременно работали все гены и синтезировались все закодированные ими белки. Бактерии, например, должны все время приспосабливаться к условиям среды, синтезируя нужные ферменты. Все клетки высших организмов имеют один и тот же набор генов, но, к счастью, клетки мозга не продуцируют пищеварительные ферменты, а в хрусталике глаза не синтезируются мышечные белки.
Активность гена характеризуется тем, транскрибируется ли он с образованием соответствующей мРНК. ДНК длинная молекула, и в определенных ее участках имеются последовательности, называемые промоторами, которые распознаются специфическим транскрибирующим ферментом полимеразой. В этих участках и только в них начинается транскрипция, продолжаясь до тех пор, пока не достигнет последовательности оснований, означающей конец считывания.
Существуют особые репрессорные белки, которые связываются с ДНК поблизости от промотора в участке, называемом оператором. Образовавшийся комплекс блокирует транскрипцию, и мРНК не синтезируется. Таким образом, репрессорные белки являются ингибиторами транскрипции. С другой стороны, существуют небольшие молекулы, которые образуют комплекс с репрессорами и снимают их блокирующее действие на транскрипцию. Иными словами, они ингибируют ингибиторы. Так, у бактерий в норме отсутствуют ферменты, катализирующие расщепление некоторых сахаров; однако если один из этих сахаров появляется в среде, он образует комплекс с репрессором, ингибирование снимается и запускается синтез соответствующего фермента. Ферменты, синтез которых индуцируется собственными субстратами, называются индуцибельными. В ряде случаев, наоборот, репрессорный белок не блокирует транскрипцию мРНК, если он не связан с определенной молекулой. У бактерий некоторые ферменты, участвующие в синтезе определенных аминокислот, образуются только в отсутствие этих аминокислот, т.е. бактерии производят данные ферменты лишь по мере надобности. Если добавить в среду соответствующую аминокислоту, она образует комплекс с репрессором и активирует его, а тем самым ингибирует транскрипцию соответствующих генов. Уже образовавшаяся мРНК вскоре расщепляется, и синтез ферментов останавливается. Такие ферменты являются отрицально индуцибельными.
Поскольку репрессорные белки сами кодируются генами, работа которых, в свою очередь, может регулироваться другими генами, а синтез малых молекул-индукторов и гормонов также в конечном счете регулируется генами, механизмы регуляции генной активности могут быть очень сложными.
Список литературы
Ичас М. Биологический код. М., 1971
Шабарова З.А., Богданов А.А. Химия нуклеиновых кислот и их компонентов, М., 1978
Зенгер В. Принципы структурной организации нуклеиновых кислот. М., 1987
www.ronl.ru
Виды нуклеиновых кислот. Нуклеиновые кислоты — фосфорсодержащие биополимеры живых организмов, обеспечивающие хранение и передачу наследственной информации. Они были открыты в 1869 г. швейцарским биохимиком Ф. Мишером в ядрах лейкоцитов, сперматозоидов лосося. Впоследствии нуклеиновые кислоты обнаружили во всех растительных и животных клетках, вирусах, бактериях и грибах.
В природе существует два вида нуклеиновых кислот — дезок-сирибонуклеиновые (ДНК) и рибонуклеиновые (РНК). Различие в названиях объясняется тем, что молекула ДНК содержит пяти-углеродный сахар дезоксирибозу, а молекула РНК— рибозу. В настоящее время известно большое число разновидностей ДНК и РНК, отличающихся друг от друга по строению и значению в метаболизме.
ДНК находится преимущественно в хромосомах клеточного ядра (99% всей ДНК клетки), а также в митохондриях и хлоропластах. РНК входит в состав рибосом; молекулы РНК содержатся также в цитоплазме, матриксе пластид и митохондрий.
Нуклеотиды — структурные компоненты нуклеиновых кислот. Нуклеиновые кислоты представляют собой биополимеры, мономерами которых являются нуклеотиды.
Нуклеотиды —сложные вещества. В состав каждого нуклео-тида входит азотистое основание, пятиуглеродный сахар (рибоза или дезоксирибоза) и остаток фосфорной кислоты.
Существует пять основных азотистых оснований: аденин, гуанин, урацил, тимин и цитозин. Первые два являются пуриновыми; их молекулы состоят из двух колец, первое содержит пять членов, второе — шесть. Следующие три являются пиримидинами и имеют одно пятичленное кольцо.
Вот как выглядит, например, формула тимидилового нуклеотида(тимидин):
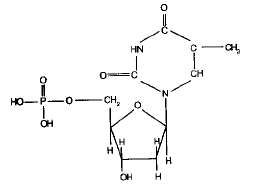
Названия нуклеотидов происходят от названия соответствующих азотистых оснований; и те и другие обозначаются заглавными буквами: аденин — аденилат (А), гуанин — гуанилат (Г), цитозин — цитидилат (Ц), тимин — тимидилат (Т), урацил — уридилат (У).
Количество нуклеотидов в молекуле нуклеиновых кислот бывает разным — от 80 в молекулах транспортных РНК до нескольких сотен миллионов у ДНК.
ДНК. Молекула ДНК состоит из двух полинуклеотидных, спирально закрученных относительно друг друга цепочек.
В состав нуклеотидов молекулы ДНК входят четыре вида азотистых оснований: аденин, гуанин, тимин и цитоцин. В полинук-леотидной цепочке соседние нуклеотиды связаны между собой ковалентными связями, которые образуются между фосфатной группой одного нуклеотида и З'-гидроксильной группой пентозы другого. Такие связи называются фосфодиэфирными. Фосфатная группа образует мостик между З'-углеродом одного пентоз-ного цикла и 5-углеродом следующего. Остов цепей ДНК образован, таким образом, сахарофосфатными остатками (рис. 1.2).
Хотя в состав ДНК входит четыре типа нуклеотидов, благодаря различной последовательности их расположения в длинной цепочке достигается огромное разнообразие этих молекул.
Полинуклеотидная цепь ДНК закручена в виде спирали наподобие винтовой лестницы и соединена с другой, комплементарной ей цепью с помощью водородных связей, образующихся между адени-ном и тимином (две связи), а также гуанином и цитозином (три связи). Нуклеотиды А и Т, Г и Ц называются комплементарными.
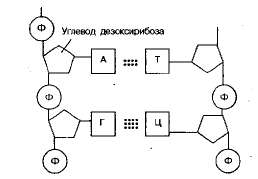
Рис 1.2. Фрагмент молекулы ДНК (между А—Т— две водородные связи; между Г—Ц — три водородные связи).
В результате у всякого организма число адениловых нуклеотидов равно числу тимидиловых, а число гуаниловых — числу цитидиловых. Эта закономерность получила название «правило Чаргаффа». Благодаря этому свойству последовательность нуклеотидов в одной цепи определяет их последовательность в другой. Такая способность к избирательному соединению нуклеотидов называется комплемен-тарностью, и это свойство лежит в основе образования новых молекул ДНК на базе исходной молекулы (репликации, т. е. удвоения).
Цепи в молекуле ДНК противоположно направлены (антипа-раллелъностъ). Так, если для одной цепи мы выбираем направление от З'-конца к 5'-концу, то вторая цепь с таким направлением будет ориентирована противоположно первой — от 5-конца к З'-концу, иначе говоря, «голова» одной цепи соединяется с «хвостом» другой и наоборот.
Впервые модель молекулы ДНК была предложена в 1953 г. американским ученым Дж. Уотсоном и англичанином Ф. Криком на основе данных Э. Чаргаффа о соотношении пуриновых и пиримидиновых оснований молекул ДНК и результатов рентге-но-структурного анализа, полученных М. Уилкинсом и Р. Франклин. За разработку двухспиральной модели молекулы ДНК Уот-сон, Крик и Уилкинс были удостоены в 1962 г. Нобелевской премии.
ДНК — самые крупные биологические молекулы. Их длина составляет от 0,25 (у некоторых бактерий) до 40 мм (у человека). Это значительно больше самой крупной молекулы белка, которая в развернутом виде достигает длины не более 100—200 нм. Масса молекулы ДНК составляет 6x10-12 г.
Диаметр молекулы ДНК 2 нм, шаг спирали 3,4 нм; каждый виток спирали содержит 10 пар нуклеотидов. Спиральная структура поддерживается многочисленными водородными связями, возникающими между комплементарными азотистыми основаниями, и гидрофобными взаимодействиями. Молекулы ДНК эука-риотических организмов линейны. У прокариот ДНК, напротив, замкнута в кольцо и не имеет ни 3-, ни 5-концов.
При изменении условий ДНК, подобно белкам, может под-. вергаться денатурации, которая называется плавлением. При постепенном возврате к нормальным условиям ДНК ренатурирует.
Функцией ДНК является хранение, передача и воспроизведение в ряду поколений генетической информации. В ДНК любой клетки закодирована информация обо всех белках данного организма, о том, какие белки, в какой последовательности и в каком количестве будут синтезироваться. Последовательность аминокислот в белках записана в ДНК так называемым генетическим (триплетным) кодом.
Основным свойством ДНК является ее способность к репликации.
Репликация — это процесс самоудвоения молекул ДНК, происходящий под контролем ферментов. Репликация осуществляется перед каждым делением ядра. Начинается она с того, что спираль ДНК временно раскручивается под действием фермента ДНК-полимеразы. На каждой из цепей, образовавшихся после разрыва водородных связей, по принципу комплементарности синтезируется дочерняя цепь ДНК. Материалом для синтеза служат свободные нуклеотиды, которые есть в ядре (рис. 1.3).

Рис. 1.3.. Схема репликации ДНК
Таким образом, каждая полинуклеотидная цепь выполняет роль матрицы для новой комплементарной цепи (поэтому процесс удвоения молекул ДНК относится к реакциям матричного синтеза). В результате получается две молекулы ДНК, у каждой из которых ' одна цепь остается от родительской молекулы (половина), а другая — вновь синтезированная. Причем одна новая цепь синтезируются сплошной, а вторая — сначала в виде коротких фрагментов, которые затем сшиваются в длинную цепь специальным ферментом—ДНК-лигазой. В результате репликации две новые молекулы ДНК представляют собой точную копию исходной молекулы.
Биологический смысл репликации заключается в точной передаче наследственной информации от материнской клетки к дочерним, что и происходит при делении соматических клеток.
РНК. Строение молекул РНК во многом сходно со строением молекул ДНК. Однако имеется и ряд существенных отличий. В молекуле РНК вместо дезоксирибозы в состав нуклеотидов входит рибоза, вместо тимидилового нуклеотида (Т) — уридило-вый (У). Главное отличие от ДНК состоит в том, что молекула РНК представляет собой одну цепь. Однако ее нуклеотиды способны образовывать водородные связи между собой (например, в молекулах тРНК, рРНК), но в этом случае речь идет о внутри-цепочечном соединении комплементарных нуклеотидов. Цепочки РНК значительно короче ДНК.
В клетке существует несколько видов РНК, которые различаются по величине молекул, структуре, расположению в клетке и функциям:
Информационная (матричная) РНК(иРНК). Этот вид наиболее разнороден по размерам и структуре. иРНК представляет собой незамкнутую полинуклеотидную цепь. Она синтезируется в ядре при участии фермента РНК-полимеразы, комплементарна участку ДНК, на котором происходит ее синтез. Несмотря на относительно низкое содержание (3—5% РНК клетки), она выполняет важнейшую функцию в клетке: служит в качестве матрицы для синтеза белков, передавая информацию об их структуре с молекул ДНК. Каждь|й белок клетки кодируется специфической иРНК, поэтому число их типов в клетке соответствует числу видов белков.
Рибосомная РНК (рРНК). Это одноцепочечные нуклеиновые кислоты, образующие в комплексе с белками рибосомы — орга-неллы, на которых происходит синтез белка. Рибосомные РНК синтезируются в ядре. Информация об их структуре закодирована в участках ДНК, которые расположены в области вторичной перетяжки хромосом. Рибосомные РНК составляют 80% всей РНК клетки, поскольку в клетке имеется огромное количество рибосом. Рибосомные РНК обладают сложной вторичной и третичной структурой, образуя петли на комплементарных участках, что приводит к самоорганизации этих молекул в сложное по форме тело. В состав рибосом входит три типа рРНК у прокариот и четыре типа рРНК у эукариот. 3. Транспортная (трансферная) РНК(тРНК). Молекула тРНК состоит в среднем из 80 нуклеотидов. Содержание тРНК в клетке — около 15% всей РНК. Функция тРНК — перенос аминокислот к месту синтеза белка. Число различных типов тРНК в клетке невелико (20—60). Все они имеют сходную пространственную организацию. Благодаря внутрицепочечкым водо-родным связям молекула тРНК приобретает характерную вторичную структуру, называемую клеверным листам. Трехмерная же модель тРНК выглядит несколько иначе. В тРНК выделяют четыре петли: акцепторную (служит местом присоединения аминокислоты), антикодоновую (узнает кодон в иРНК в процессе трансляции) и две боковые.
Источник : Н.А. Лемеза Л.В.Камлюк Н.Д. Лисов "Пособие по биологии для поступающих в ВУЗы"
Список литературы
Для подготовки данной работы были использованы материалы с сайта http://biology.asvu.ru
Дата добавления: 07.06.2008
www.km.ru
Содержание
1. История открытия и названия нуклеиновых кислот
2. Нахождение нуклеиновых кислот в природе
3. Получение нуклеиновых кислот
4. Химические свойства нуклеиновых кислот
5. Применение нуклеиновых кислот
6. Занимательные факты о нуклеиновых кислотах
1. История открытия нуклеинов и их названия
Открытие нуклеиновых кислот связано с именем молодого врача изгорода Базеля (Швейцария) Фридриха Мишера. После окончания медицинского факультета Мишер был послан для усовершенствования и работы над диссертацией в Тюбинген (Германия) в физиолого-химическую лабораторию, возглавляемуюФ. Гоппе-Зейлером. Тюбингенская лаборатория в то время была известна ученому миру. Пройдя практику по органической химии, Мишер приступил к работе в биохимической лаборатории. Ему было поручено заняться изучением химического состава гноя. Молодой ученый не возражал против предложенной темы, так как считал лейкоциты, присутствующие в гное, одними из самых простых клеток.
Путём многочисленных опытов он получил из гнойных клеток вещество ядерного происхождения. Мишер был уверен именно в ядерном его источнике. Поэтому он начал более тщательное выделение ядер. В то время еще никто в биохимических лабораториях не пытался выделить ядра или какие-либо другие субклеточные компоненты, так что и здесь он был пионером.
Продолжив дальше очищать ядро от других клеточных фрагментов, он получил странное вещетво. Оно не разлагалось протеолитическими ферментами, значит, не являлось белком. Отсутствие растворимости в горячем спирте указывало на то, что это вещество не являлось и фосфолипидом. По-видимому, оно относилось к новому классу биохимических соединений.
Но Мишер с большой горячностью настаивал на точности своих результатов и добивался разрешения опубликовать их в печати. Тогда Гоппе-Зейлер решил проверить данные Мишера лично. Он и два его ассистента (одним из них был русский химик Любавин) в течение года шаг за шагом прошли все этапы аналитической работы Мишера и полностью подтвердили его данные, выделив нуклеин из клеток крови и из дрожжей.

В 1871 г.работа Мишера вместе с подтверждающими ее контрольными работами Гоппе-Зейлера и его ассистентов увидела свет.Существование нуклеина как специфического ядерного вещества стало научным фактом. Вскоре методика Мишера была применена для выделения нуклеина из различных тканей.
Термин«нуклеиновые кислоты» был предложен в 1889: нуклеиновыми они были названы потому, что впервые были открыты в ядрах клеток, а кислотами — из-за наличия в их составе остатков фосфорной кислоты. Позже было показано, что нуклеиновые кислоты построены из большого числа нуклеотидов (от нескольких десятков до сотен миллионов). В состав каждого нуклеотида входит азотистое основание, углевод (пентоза) и фосфорная кислота.
2. Нахождение нуклеиновых кислот в природе
Нуклеиновые кислоты в природе встречаются во всех живых клетках. Живые клетки, за исключением сперматозоидов, в норме содержат значительно больше рибонуклеиновой, чем дезоксирибонуклеиновой кислоты. На методы выделения дезоксирибонуклеиновых кислот оказало большое влияние то обстоятельство, что, тогда как рибонуклеопротеиды и рибонуклеиновые кислоты растворимы в разбавленном (0,15 М) растворе хлористого натрия, дезоксирибонуклеопротеидные комплексы фактически в нем нерастворимы.
Поэтому гомогенизированный орган или организм тщательно промывают разбавленным солевым раствором, из остатка с помощью крепкого солевого раствора экстрагируют дезоксирибонуклеиновую кислоту, которую осаждают затем добавлением этанола.
В клетках эукариот (например, животных или растений) ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органоидах (митохондриях и пластидах). В клетках прокариотических организмов (бактерий и архей) кольцевая или линейная молекула ДНК, так называемый нуклеотид, прикреплена изнутри к клеточной мембране. У них и у низших эукариот (например, дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами. Кроме того, одно- или двухцепочечные молекулы ДНК могут образовывать геном ДНК-содержащих вирусов.
3. Получение нуклеиновых кислот
В клетках нуклеиновые кислоты связаны с белками, образуя нуклеопротеиды. Выделение нуклеиновых кислот сводится к очистке их от белков. Для этого препараты, содержащие нуклеиновые кислоты, обрабатывают ПАВ и экстрагируют белки фенолом. Послед, очистка и фракционирование нуклеиновых кислот проводятся с помощью ультрацентрифугирования, различных видов жидкостной хроматографии и гель - электрофореза. Для получения индивидуальных нуклеиновых кислот обычно используют различные варианты последнего метода.
Современные методы химического синтеза нуклеиновых кислот позволяют получать крупные фрагменты ДНК, в том числе целые гены. Методические основы химически - ферментативных методов синтеза ДНК разработаны X. Кораной.
Они включают:
- химический синтез комплементарных, взаимоперекрывающихся олигонуклеотидов, из которых затем в результате комплементационных взаимодействий выстраиваются дуплексы - фрагменты молекулы синтезируемой ДНК с несовпадающими разрывами в обеих цепях;
- соединение (лигирование) таких олигонуклеотидов в составе дуплекса с помощью фермента Т4 ДНК-лигазы. Сборку протяженных ДНК из синтетически однотяжевых олигонуклеотидов проводят в несколько этапов. Сначала собирают небольшие дуплексы с "липкими" концами (однотяжевыми комплементарными участками), из которых затем последовательно формируют более протяженные структуры. Таким образом могут быть получены искусственные фрагменты ДНК большой длины и с любой нуклеотидной последовательностью. С помощью генетической инженерии возможно клонирование (получение в индивидуальном виде и размножение) искусственных ДНК.
Несмотря на малую эффективность этого метода, были синтезированы олигонуклеотиды, содержащие до 16 звеньев, из которых были собраны первые синтетические гены. Фосфодиэфирный метод образования межнуклеотидных связей, использованный Кораной, имеет историческое значение. Однако разработанные им приемы введения и избирательные удаления защитных групп широко используются в других методах синтеза нуклеиновых кислот.
Важным шагом в совершенствовании синтеза олигонуклеотидов явиласьразработка так называемого фосфотриэфирного метода. Образующийся динуклеотид после частичного деблокирования фосфата конденсируют аналогичным образом с другими динуклеотидом и т.д. Применение этого способа, в котором используют защиту фосфатной группы, позволило значительно сократить время синтеза и повысить выходы олигонуклеотидов.
Параллельно этим методам, которые осуществляют в растворах, разрабатывались твердофазные способы синтеза нуклеиновых кислот. В последнем случае процесс проводят в двухфазной системе; нуклеозидный компонент связан ковалентно с нерастворимым полимером, а нуклеотидный компонент и необходимые реагенты находятся в растворе.
Обычно в этом случае на первой стадии нуклеозид присоединяют с помощью "якорной" группы к нерастворимому полимеру. Затем его 5'-гидроксильную группу деблокируют и конденсируют с нуклеотидным компонентом. У образующегося полностью защищенного динуклеозидмонофосфата деблокируют защитную группу в положении 5' и присоединяют следующему нуклеотид и т.д.
Наиболее распространенные методы твердофазного синтеза олигонуклеотидов основаны на использовании нуклеотидного компонента, содержащегоР(III).В так называемом амидофосфитном способе нуклеотидным компонентом является эфир 3'-амидофосфита дезоксинуклеозида. Достаточно устойчивые амидофосфиты при протонировании в присутствии тетразола превращаются в сильные фосфорилирующие агенты. После завершения синтеза удаляют защитные группы с межнуклеотидных фосфатов, отделяют олигонуклеотид от носителя, деблокируют группы Nh3 гетероциклов. Липофильную группу (МеО)2Тr удаляют после первого хроматографического разделения.
Стандартность операций в твердофазном синтезе олигонуклеотидов явилась основой для автоматизации процесса. Принцип работы автомата-синтезатора основан на подаче в реактор с помощью насоса (под контролем микропроцессора) защищенных нуклеотидных компонентов реагентов и растворителей по заданной программе в колонку, содержащую полимерный носитель с закрепленным на нем первым нуклеозидом. После окончания синтеза и отделения полностью защищенного олигонуклеотида от полимерного носителя проводят деблокирование, очистку и анализ синтезированных фрагментов ДНК. Так, с помощью гидрофосфорильного метода вавтомате - синтезаторе за несколько часов получают 30-40-звенные олигонуклеотиды; возможен синтез более чем 100-звенных фрагментов ДНК.Разработаны синтезаторы, позволяющие проводить одновременно синтез несколько олигонуклеотидов.
Синтез олигорибонуклеотидов ферментативным путем осуществляют обычно с использованием рибонуклеаз или полинуклеотидфосфорилаз.
В качестве нуклеотидного и нуклеозидного компонента применяют мономеры или олигонуклеотиды. Эту реакцию используют для синтеза ди-, три- и тетрарибонуклеотидов. При увеличении длины олигорибонуклеотида начинает преобладать обратная реакция (гидролиз олигонуклеотида).
Химический синтез олигорибонуклеотидов проводят в основном с использованием тех же приемов, как и при синтезе ДНК.
4. Химические свойства нуклеиновых кислот
Нуклеиновые кислоты:
- хорошо растворимы в воде
- практически не растворимы в органических растворителях.
- очень чувствительны к действию температуры и критических значений уровня pH.
- молекулы ДНК с высокой молекулярной массой, выделенные из природных источников, способны фрагментироваться под действием механических сил, например при перемешивании раствора.
- нуклеиновые кислоты фрагментируются ферментами — нуклеазами.
Химические свойства РНК.
Напоминают свойства ДНК, однако наличие дополнительных групп ОН в рибозе и меньшее (в сравнении с ДНК) содержание стабилизированных спиральных участковделает молекулы РНК химически более уязвимыми. При действии кислот или щелочей основные фрагменты полимерной цепи Р(О)-О-СН2 легко гидролизуются, группировки А, У, Г и Ц отщепляются легче. Если нужно получить мономерные фрагменты, сохранив при этом химически связанные гетероциклы, используют деликатно действующие ферменты, называемые рибонкулеазами.
Химические свойства ДНК.
В воде ДНК образует вязкие растворы, при нагревании таких растворов до 60°С или при действии щелочей двойная спираль распадается на две составляющие цепи, которые вновь могут объединиться, если вернуться к исходным условиям. В слабокислых условиях происходит гидролиз, в результате частично расщепляются фрагменты – Р-О-СН2- с образованием фрагментов – Р-ОН и НО-СН2 , соответственно результате образуются мономерные, димерные (сдвоенные) или примерные (утроенные) кислоты, представляющие собой звенья, из которых была собрана цепь ДНК.
Участие ДНК и РНК в синтезе белков – одна из основных функций нуклеиновых кислот. Белки – важнейшие компоненты каждого живого организма. Мышцы, внутренние органы, костная ткань, кожный и волосяной покров млекопитающих состоят из белков. Это полимерные соединения, которые собираются в живом организме из различных аминокислот. В такой сборке управляющую роль играют нуклеиновые кислоты, процесс проходит в две стадии, причем на каждой из них определяющий фактор –взаимоориентация азотсодержащих гетероциклов ДНК и РНК.
Основная задача ДНК – хранить записанную информацию и предоставлять в тот момент, когда начинается синтез белков. В связи с этим понятна повышенная химическая устойчивость ДНК в сравнении с РНК.
Природа позаботилась о том, чтобы сохранить по возможности основную информацию неприкосновенной
5. Применение нуклеиновых кислот
Последнее десятилетие характеризуется интенсивным развитием технологий, которые ориентированы на создание устройств, позволяющих получать информацию о свойствах различных сред (объектов) в форме электрического сигнала. В сенсорных технологиях чувствительный элемент способен "узнать" исследуемое вещество среди множества родственных и преобразовать полученную информацию о его присутствии в ответ, фиксируемый в цифровой или аналоговой форме. Наибольшее развитие имеют аналитические устройства, использующие в качестве узнающего элементабиомакромолекулы - биосенсоры.
Принцип действия биодатчиков, использующих частицы жидкокристаллической дисперсии, состоит в следующем: азотистые основания в молекулах ДНК, фиксированных в структуре холестерической жидкокристаллической дисперсии, тем или иным способом "узнают" молекулы биологически активного соединения (БАС) и "адресуют" их в определенные места на поверхности ДНК. Образование комплекса "ДНК-БАС" приводит к появлению первичного (в частности, оптического) сигнала. Пространственная структура холестерика многократно усиливает генерируемый в системе первичный сигнал и делает видимыми результаты действия биологически активного соединения на ДНК: в спектре кругового дихроизма появляется аномальная полоса (полосы) в области поглощения биологически активного соединения. Амплитуда этой полосы пропорциональна концентрации биологически активного соединения, а знак полосы несет информацию о способе ориентации его молекул по отношению к парам оснований ДНК.
В последние годы возрос интерес к иммуностимуляторам. Впервые нуклеиновые кислоты стали применять в 1882 году по инициативе Горбачевского при инфекционных заболеваниях стрепто - и стафилококкового происхождения. В 1911 году Черноруцкий установил, что под влиянием дрожжевой нуклеиновой кислоты увеличивается количество иммунных тел.
Нуклеинат натрия:увеличивает фагоцитарную активность, активирует поли- и мононуклеары, увеличивает эффективность тетрациклинов при смешанной инфекции, вызванной стафилококком и синегнойной палочкой. При профилактическом введении нуклеинат натрия обусловливает и противовирусный эффект, так как обладает интерфероногенной активностью.
Нуклеинат натрия ускоряет формирование прививочного иммунитета, увеличивает его качество, позволяет уменьшить дозу вакцины. Этот препарат оказывает позитивный эффект при лечении больных с хроническим паротитом, язвенной болезнью, различными формами пневмонии, хроническим воспалением легких, бронхиальной астмой. Нуклеинат натрия увеличивает содержание РНК и белка в макрофагах в 1,5 раза и гликогена в 1,6 раза, увеличивает активность лизосомальных ферментов, следовательно, увеличивает завершенность фагоцитоза макрофагами. Препарат увеличивает содержание у человека лизоцима и нормальных антител, если их уровень был снижен.
Особое место среди препаратов нуклеиновых кислот занимаетиммунная РНК макрофагов, которая представляет собой информационную РНК, которая вносит в клетку фрагмент антигена. То есть, идет неспецифическая стимуляция иммунокомпетентных клеток нуклеотидами.
Неспецифическими стимуляторами являются синтетические двухцепочечные полинуклеотиды, которые стимулируют антителообразование, увеличивают антигенный эффект неиммуногенных доз антигена, обладающего антивирусными свойствами, связанными с интерфероногенной активностью. Их механизм действия сложен и недостаточно выяснен. Двунитчатая РНК включается в систему регуляции синтеза белка в клетке, активно взаимодействуя с клеточной мембраной.
Но высокая стоимость препаратов, недостаточная их эффективность, наличие побочных явлений (тошнота, рвота, снижение артериального давления, увеличение температуры тела, нарушение функций печени, лимфопения - из-за прямого токсического действия на клетки), отсутствие схем использованияделают применение препаратов ограниченным.
6. Занимательные факты
- Почти полвека тому назад был открыт принцип структурной (молекулярной) организации генного вещества – дезоксирибонуклеиновой кислоты (ДНК). Структура ДНК дала ключ к механизму точного воспроизведения генного вещества. Так возникла новая наука – молекулярная биология.
- Накопление знаний о генетическом коде, нуклеиновых кислотах и биосинтезе белков привело к утверждению принципиально новой идеи о том, что все начиналось вовсе не с белков, а с РНК.
- Известно, что рибонуклеиновая кислота является основным переносчиком генетической информации от ДНК к белку. Поэтому многие заболевания связаны именно с неправильной передачей этой информации.
- Достаточно неожиданно обнаружилось, что во внеклеточных жидкостях организма находится весьма заметное количество нуклеиновых кислот. До сих пор не понятно, как они туда попадают. Самым простым было бы предположить, что нуклеиновые кислоты оказываются во внеклеточном пространстве при гибели клеток. Однако, имеются факты, противоречащие этому предположению.
- Расшифровка структуры ДНК (1953 г.) стала одним из поворотных моментов в истории биологии. За выдающийся вклад в это открытие Фрэнсису Крику, Джеймсу Уотсону, Морису Уилкинсу была присуждена Нобелевская премия по физиологии и медицине 1962 г.
superbotanik.net
