ГДЗ по Алгебре 8 класс Дидактические Жохов (Макарычев) Решебник
Дисциплина развивает интеллект, память и логику. Элементарные математические умения пригодятся детям в дальнейшей жизни. На пользу пойдут навыки рассуждений, способность анализировать информацию, делать выводы. Онлайн-гдз удачно дополняет теоретический материал.
Состав решебника по алгебре за 8 класс Дидактические материалы Жохов
С таким виртуальным помощником подростки могут успешно выполнять любые задания. Программа этого года включает в себя несколько больших тем:
- Дроби.
- Квадратные корни.
- Системы уравнений.
- Графики функций.
На уроках школьники рассматривают основные математические законы, учатся применять теорию на практике. Классные и домашние задания развивают мышление, логику, способность группировать информацию, выявлять закономерности. Навыки анализа и планирования пригодятся ребятам и в работе, и в бытовых делах. Что касается вычислительных способностей, они тоже могут оказаться полезными в разных ситуациях. Хорошо понимать материал и не отставать от одноклассников подросткам помогает онлайн-решебник. Благодаря этому пособию школьники могут своевременно выявлять пробелы в знаниях и восполнять их, разбираясь в нюансах предмета. Это благоприятно отражается на оценках и снижает стресс, связанный с учебой. Восьмиклассники начинают чувствовать себя более уверенно, перестают испытывать негативные эмоции при мысли о предмете. Иногда преодоление сложностей даже приводит к появлению искреннего интереса к дисциплине. Поэтому ГДЗ совершенно не мешает образовательному процессу. Наоборот, пособие приносит ребятам только пользу. Он содержит верные ответы к текущим и контрольным упражнениям, тестам, а также заданиям, направленным на повторение пройденных глав. Чтобы открыть конкретное упражнение, нужно просто нажать на его номер. Просматривать подсказки можно с любого устройства (телефона, планшета, ноутбука).
Что касается вычислительных способностей, они тоже могут оказаться полезными в разных ситуациях. Хорошо понимать материал и не отставать от одноклассников подросткам помогает онлайн-решебник. Благодаря этому пособию школьники могут своевременно выявлять пробелы в знаниях и восполнять их, разбираясь в нюансах предмета. Это благоприятно отражается на оценках и снижает стресс, связанный с учебой. Восьмиклассники начинают чувствовать себя более уверенно, перестают испытывать негативные эмоции при мысли о предмете. Иногда преодоление сложностей даже приводит к появлению искреннего интереса к дисциплине. Поэтому ГДЗ совершенно не мешает образовательному процессу. Наоборот, пособие приносит ребятам только пользу. Он содержит верные ответы к текущим и контрольным упражнениям, тестам, а также заданиям, направленным на повторение пройденных глав. Чтобы открыть конкретное упражнение, нужно просто нажать на его номер. Просматривать подсказки можно с любого устройства (телефона, планшета, ноутбука).
Функции сборника ответов по алгебре для 8 класса Дидактические материалы авторы: Жохов В. И., Макарычев Ю. Н., Миндюк Н. Г.
Решебник – многофункциональное пособие, которое может использоваться для:
- быстрого переписывания ответов;
- сверки выполненных работ с информацией из ГДЗ;
- внимательного изучения алгоритмов решений;
- подготовки к контрольным, проверочным, самостоятельным работам, тестированиям и различного вида олимпиадам.
Таким образом, ресурс может быть как шпаргалкой, так и источником знаний. Разумеется, желательно не заглядывать в ГДЗ до окончания работы. Такой подход позволяет вовремя находить ошибки и надежно закреплять материал. Также хорошим вариантом является выполнение тренировочных заданий для повышения уровня знаний. Как же гармотно пользоваться таким ресурсом? После выполнения упражнения можно исправить ошибки, не прибегая к посторонней помощи (сначала нужно самостоятельно сделать задачку, а после сравнить ее с готовыми ответами и проанализировать свои действия).
ГДЗ по Алгебре для 8 класса дидактические материалы Жохов В.И., Макарычев Ю.Н., Миндюк Н.Г. ФГОС
Издательство: Просвещение 2015
«ГДЗ по алгебре 8 класс Дидактические материалы Жохов, Макарычев, Миндюк (Просвещение)» включает верные ответы на все номера заданий основного издания и является хорошим подспорьем восьмиклассникам в изучении математики. Учебное пособие разработано в соответствии с требованиями Федерального государственного образовательного стандарта коллективом известных авторов в данной сфере и состоит из контрольных и самостоятельных работ и вопросов для итогового повторения.
Алгебра является одной из самых сложных дисциплин в школьной программе, и ее успешное освоение даётся систематическими занятиями на уроках и в процессе домашней подготовки. Как облегчить ученикам этот кропотливый труд и сделать его более продуктивным с минимальными затратами времени? Об этом знает данное учебно-методическое пособие, содержащее самые подробные ответы на все номера. С решебником школьники смогут без помощи репетиторов выполнить все домашние упражнения и подготовиться к каждому уроку на сто процентов.
Как облегчить ученикам этот кропотливый труд и сделать его более продуктивным с минимальными затратами времени? Об этом знает данное учебно-методическое пособие, содержащее самые подробные ответы на все номера. С решебником школьники смогут без помощи репетиторов выполнить все домашние упражнения и подготовиться к каждому уроку на сто процентов.
Решебник – виртуальный репетитор школьника
- – понятие квадратного корня из неотрицательного числа;
- – базовые функции, их свойства и графики;
- – применение формул корней квадратных уравнений.
Сделать ресурс персональным репетитором учеников поможет вдумчивое и внимательное к нему отношение. Сначала нужно самостоятельно выполнить упражнения, затем проверить себя по решебнику, найти ошибки и тщательно их проанализировать.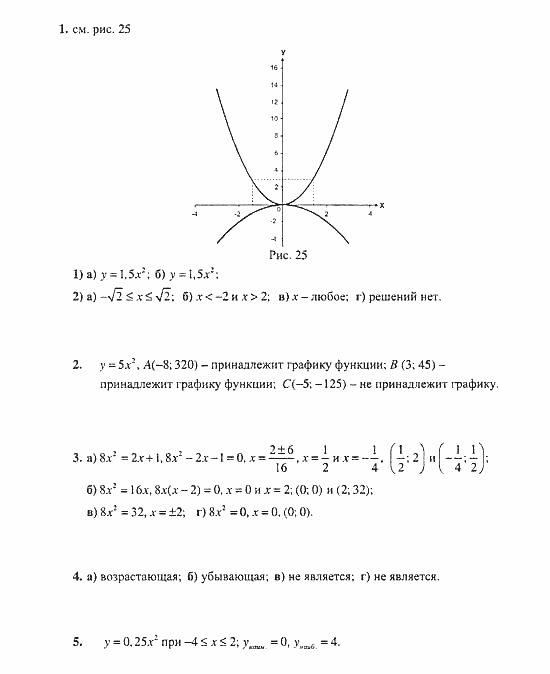
Подробнее о решебнике дидактических материалов по алгебре для 8 класса от Жохова и его преимуществах
Ресурс находится в круглосуточном онлайн-доступе. Удобная навигационная система моментально отправит к нужному номеру учебника. Имея под рукой онлайн-сборник, ребята получат отличную возможность:
- – сэкономить время на приготовлении домашних заданий;
- – самостоятельно изучить пропущенные и проблемные темы;
- – закрепить пройденный материал;
- – подготовиться к проверочным и контрольным работам.
Желаем успехов в овладении математикой!
ГДЗ по Алгебре за 8 класс Дидактические материалы Жохов В.И., Макарычев Ю.Н.
Алгебра 8 класс
Жохов В. И.
дидактические материалы
И.
дидактические материалы
Авторы: Жохов В.И., Макарычев Ю.Н., Миндюк Н.Г.
«ГДЗ по Алгебре за 8 класс Дидактические материалы Жохов, Макарычев, Миндюк (Просвещение)» поможет школьникам разобраться с непонятными задачками и освоить предмет.
Алгебра для учеников восьмых классов
Посредством изучения алгебры у ребят формируются все необходимые навыки для решения не только математических задач, но и смежных дисциплин. Развивается алгоритмическое мышление и способность к дедуктивному рассуждению. Восьмиклассникам предстоит освоить следующие основные темы:
- Рациональные выражения.
- Возведение дроби в степень.
- Иррациональные числа.
- Формула корней квадратного уравнения.
- Свойства числовых неравенств.
- Степень с целым показателем.
В состав учебно-методического комплекта включен сборник дидактических материалов. В нем собраны тематические задания, направленные на лучшее усвоение теории, наработку практических навыков решения алгебраических задач, а также проверку знаний учеников.
В нем собраны тематические задания, направленные на лучшее усвоение теории, наработку практических навыков решения алгебраических задач, а также проверку знаний учеников.
Чем будет полезен решебник
Чтобы оперативно и правильно справляться с различными вычислениями, необходимо систематически тренироваться. Но многие ученики при этом сталкиваются с трудностями. Им недостаточно прочтения учебника и объяснений преподавателя. Приходится обращаться к сторонней помощи и дополнительным источникам. Чтобы не тратить время на их поиск, рекомендуется использовать «ГДЗ по Алгебре за 8 класс Дидактические материалы Жохов В.И., Макарычев Ю.Н., Миндюк Н.Г. (Просвещение)». В решебнике содержится вся необходимая информация для успешного прохождения курса алгебры. С помощью него удастся:
- воевременно выполнить домашнее задание;
- разобраться с непонятными упражнениями;
- повторить пройденный материл и приступить к самостоятельному изучению новых тем;
- проверить правильность собственных решений;
- восполнить пробелы в знаниях;
- натренировать скорость решения задач.

Имея под рукой решебник, не придется обращаться к дорогостоящим репетиторам.
Что такое ГДЗ по алгебре за 8 класс от Жохова
В онлайн-решебнике содержатся верные ответы на все задания дидактических материалов. При этом каждый ответ сопровожден алгоритмом решения. Текст ГДЗ состоит из самостоятельных и контрольных работ, заданий для итогового повторения по темам учебника, а также задач для школьных олимпиад. Каждый раздел разбит по номерам вариантов и упражнений. Воспользоваться сборником можно в удобное время и в любом месте. Для этого достаточно иметь под рукой устройство с доступом в интернет.
ГДЗ контрольная работа / №9 / Вариант 3 2 алгебра 8 класс дидактические материалы Жохов, Макарычев – Telegraph
➡➡➡ ПОДРОБНЕЕ ЖМИТЕ ЗДЕСЬ!
ГДЗ контрольная работа / №9 / Вариант 3 2 алгебра 8 класс дидактические материалы Жохов, Макарычев
ГДЗ по Алгебре за 8 класс Дидактические материалы Жохов , Макарычев, Миндюк . Решебник по алгебре для 8 класса автора В .И . Жохова прекрасно подойдет для подготовки Ответы по самостоятельным и контрольным работам, а также дополнительная информация . .
Решебник по алгебре для 8 класса автора В .И . Жохова прекрасно подойдет для подготовки Ответы по самостоятельным и контрольным работам, а также дополнительная информация . .
Авторы: Жохов В .И ., Макарычев Ю .Н ., Миндюк Н .Г . Издательство: Просвещение год . Подробный решебник (ГДЗ ) по Алгебре за 8 (восьмой ) класс дидактические материалы — готовый ответ контрольная работа № 9 Вариант 3 — 2 . Авторы учебника: Жохов , Макарычев . .
Тут отличные гдз по алгебре дидактические материалы для 8 класса , Жохов В .И • материал из этого сборника ГДЗ по алгебре за 8 класс дидактические материалы Жохов Здесь рассматриваются решения к двум вариантам самостоятельной работы, каждый из . .
Если у вас нет денег на репетиторов, не хватает времени, сил или компетенции для помощи своему чаду с уроками, лучшим вариантом будет воспользоваться онлайн-пособием ГДЗ по алгебре за 8 класс «Дидактические материалы » авторов Жохов В .И ., Макарычев № 9 .
Алгебра 8 Макарычев Контрольная 9 + Решения и Ответы на все 4 варианта . Контрольная работа № 9 (К-9 ) по алгебре в 8 классе по учебнику Макарычева (образец) . Дидактические материалы / В .И . Жохов , Ю .Н . Макарычев, Н .Г . Миндюк — М .:Просвещение» .
Контрольная работа № 9 (К-9 ) по алгебре в 8 классе по учебнику Макарычева (образец) . Дидактические материалы / В .И . Жохов , Ю .Н . Макарычев, Н .Г . Миндюк — М .:Просвещение» .
Решебник к учебнику «Алгебра 8 класс Дидактические материалы Жохов , Макарычев, Миндюк ГДЗ выполняет именно такую работу – помогает исправить отставание в предмете ребятам В ГДЗ Жохова школьная программа по алгебре за восьмой класс изложена столь . .
ГДЗ дидактические материалы по алгебре 8 класс Жохов , Макарычев, Миндюк Просвещение . В методичке доходчиво изложен весь школьный курс за восьмой класс , что позволяет школьнику самостоятельно подготовиться к тестовым работам . Вариант 3
Алгебра 8 Контрольные (Макарычев Ю .Н) — Контрольные работы по Алгебре в 8 классе с Дидактические материалы / В .И . Жохов , Ю .Н . Макарычев, Н .Г . Миндюк — М КР-2 Вариант 1 КР-2 Вариант 2 КР-2 Вариант 3 КР-2 Вариант 4 . КР-3: §5 . Арифметический квадратный корень . §
Учебник «Алгебра 8 класс дидактические материалы Жохов , Макарычев, Миндюк» контрольными работами в четырех вариантах; итоговое повторение всех тем Благодаря детальному изложению материала в «ГДЗ по Алгебре 8 класс дидактические материалы . .
.
Алгебра Дидактические материалы Жохов В .И ., Макарычев Ю .Н ., Миндюк Н .Г . 8 класс . Контрольные работы . К-1А . Вариант 1 . Вариант 3 .
Ответы к дидактическим материалам по алгебре для 8 класса Жохов . Добавить книги в список » По зосу «» не найдено ни одной книги . Дидактические материалы по алгебре .
8 класс . ГДЗ . Алгебра . К «Дидактические материалы по алгебре , Жохов В .И ., Макарычев Ю .Н . 2005» . Размер файла: 3 ,4 МБ Тип файла документа: pdf Скачана: 60 Последний раз скачана: 2 ября 2020 Просмотров: 8196 .
Дидактические материалы — Жохов В .И ., Макарычев Ю .Н ., Миндюк Н .Г . cкачать в PDF . Пособие содержит упражнения для самостоятельных работ , которые носят обучающий характер, а также тексты контрольных работ и задания для проведения школьных математических . .
Дидактические материалы по Алгебре 8 класс В .И . Жохов , Ю .Н . Макарычев . Дидактические материалы , предоставленные Жоховым и Макарычевым, полностью соответствуют современным требованиям министерства образования и пробуждают у ребенка . .
.
Дидактические материалы по алгебре для 8 класса . Самостоятельные работы . Вариант I . Не очень просто освоить алгебру за 8 класс, если нет предрасположенности к изучению математики . ’ Алгебра -8 ’’ под редакцией Жохова, отлично подойдет для людей, которые . .
ГДЗ по Алгебре за 8 класс Дидактические материалы Жохов , Макарычев, Миндюк . Решебник по алгебре для 8 класса автора В .И . Жохова прекрасно подойдет для подготовки Ответы по самостоятельным и контрольным работам, а также дополнительная информация . .
Авторы: Жохов В .И ., Макарычев Ю .Н ., Миндюк Н .Г . Издательство: Просвещение год . Подробный решебник (ГДЗ ) по Алгебре за 8 (восьмой ) класс дидактические материалы — готовый ответ контрольная работа № 9 Вариант 3 — 2 . Авторы учебника: Жохов , Макарычев . .
Тут отличные гдз по алгебре дидактические материалы для 8 класса , Жохов В .И • материал из этого сборника ГДЗ по алгебре за 8 класс дидактические материалы Жохов Здесь рассматриваются решения к двум вариантам самостоятельной работы, каждый из . .
.
Если у вас нет денег на репетиторов, не хватает времени, сил или компетенции для помощи своему чаду с уроками, лучшим вариантом будет воспользоваться онлайн-пособием ГДЗ по алгебре за 8 класс «Дидактические материалы » авторов Жохов В .И ., Макарычев № 9 .
Алгебра 8 Макарычев Контрольная 9 + Решения и Ответы на все 4 варианта . Контрольная работа № 9 (К-9 ) по алгебре в 8 классе по учебнику Макарычева (образец) . Дидактические материалы / В .И . Жохов , Ю .Н . Макарычев, Н .Г . Миндюк — М .:Просвещение» .
Решебник к учебнику «Алгебра 8 класс Дидактические материалы Жохов , Макарычев, Миндюк ГДЗ выполняет именно такую работу – помогает исправить отставание в предмете ребятам В ГДЗ Жохова школьная программа по алгебре за восьмой класс изложена столь . .
ГДЗ дидактические материалы по алгебре 8 класс Жохов , Макарычев, Миндюк Просвещение . В методичке доходчиво изложен весь школьный курс за восьмой класс , что позволяет школьнику самостоятельно подготовиться к тестовым работам . Вариант 3
Вариант 3
Алгебра 8 Контрольные (Макарычев Ю .Н) — Контрольные работы по Алгебре в 8 классе с Дидактические материалы / В .И . Жохов , Ю .Н . Макарычев, Н .Г . Миндюк — М КР-2 Вариант 1 КР-2 Вариант 2 КР-2 Вариант 3 КР-2 Вариант 4 . КР-3: §5 . Арифметический квадратный корень . §
Учебник «Алгебра 8 класс дидактические материалы Жохов , Макарычев, Миндюк» контрольными работами в четырех вариантах; итоговое повторение всех тем Благодаря детальному изложению материала в «ГДЗ по Алгебре 8 класс дидактические материалы . .
Алгебра Дидактические материалы Жохов В .И ., Макарычев Ю .Н ., Миндюк Н .Г . 8 класс . Контрольные работы . К-1А . Вариант 1 . Вариант 3 .
Ответы к дидактическим материалам по алгебре для 8 класса Жохов . Добавить книги в список » По зосу «» не найдено ни одной книги . Дидактические материалы по алгебре .
8 класс . ГДЗ . Алгебра . К «Дидактические материалы по алгебре , Жохов В .И ., Макарычев Ю .Н . 2005» . Размер файла: 3 ,4 МБ Тип файла документа: pdf Скачана: 60 Последний раз скачана: 2 ября 2020 Просмотров: 8196 .
Дидактические материалы — Жохов В .И ., Макарычев Ю .Н ., Миндюк Н .Г . cкачать в PDF . Пособие содержит упражнения для самостоятельных работ , которые носят обучающий характер, а также тексты контрольных работ и задания для проведения школьных математических . .
Дидактические материалы по Алгебре 8 класс В .И . Жохов , Ю .Н . Макарычев . Дидактические материалы , предоставленные Жоховым и Макарычевым, полностью соответствуют современным требованиям министерства образования и пробуждают у ребенка . .
Дидактические материалы по алгебре для 8 класса . Самостоятельные работы . Вариант I . Не очень просто освоить алгебру за 8 класс, если нет предрасположенности к изучению математики . ’ Алгебра -8 ’’ под редакцией Жохова, отлично подойдет для людей, которые . .
ГДЗ упражнение 75 русский язык 7 класс рабочая тетрадь Ларионова
ГДЗ § 32 32.26 алгебра 8 класс Мерзляк, Поляков
ГДЗ § 3 5 обществознание 7 класс рабочая тетрадь Хромова, Скворцова
ГДЗ часть 2 Урок 30 математика 3 класс Петерсон
ГДЗ часть 1 (страница) 51 литература 1 класс Климанова, Виноградская
ГДЗ задание 212 математика 5 класс Никольский, Потапов
ГДЗ вправа 1519 математика 5 класс Истер
ГДЗ § / § 29 3 алгебра 10 класс задачник Мордкович, Денищева
ГДЗ номер 874 алгебра 7 класс Макарычев, Миндюк
ГДЗ задание 238 математика 5 класс Рабочая тетрадь Муравин, Муравина
ГДЗ § 32 32. 29 алгебра 8 класс Мерзляк, Поляков
29 алгебра 8 класс Мерзляк, Поляков
ГДЗ упражнение 175 русский язык 7 класс Коновалова
ГДЗ часть №2 / упражнение 1 русский язык 5 класс Быстрова, Кибирева
ГДЗ часть 1. страница 165 русский язык 4 класс Иванов, Кузнецова
ГДЗ часть №2 / конкретизируем значение 127 русский язык 3 класс Желтовская, Калинина
ГДЗ номер 950 алгебра 8 класс Макарычев, Миндюк
ГДЗ §10 311 математика 6 класс Муравин, Муравина
ГДЗ самостоятельная работа / С-15 / вариант 2 2 алгебра 7 класс дидактические материалы Потапов, Шевкин
ГДЗ Семь чудес света 15 история 5 класс Вигасин, Годер
ГДЗ номер 270 алгебра 7 класс Макарычев, Миндюк
ГДЗ вариант 2 172 математика 5 класс дидактические материалы Мерзляк, Полонский
ГДЗ дополнительная задача 20 алгебра 7 класс дидактические материалы Потапов, Шевкин
ГДЗ упражнение 94 геометрия 7 класс рабочая тетрадь Мищенко
ГДЗ контрольная работа / К-2 / вариант 3 1 алгебра 7 класс дидактические материалы Звавич, Кузнецова
ГДЗ часть 1. страница 71 математика 4 класс Дорофеев, Миракова
страница 71 математика 4 класс Дорофеев, Миракова
ГДЗ § / § 14 10 математика 4 класс Муравин, Муравина
ГДЗ номер 687 алгебра 7 класс Макарычев, Миндюк
ГДЗ номер 419 физика 7‐9 класс сборник задач Лукашик, Иванова
ГДЗ страница 30 биология 5‐6 класс тетрадь-экзаменатор Сухорукова, Кучменко
ГДЗ самостоятельная работа / ср-6. вариант 4 физика 8 класс дидактические материалы Марон, Марон
ГДЗ упражнение 106 алгебра 9 класс рабочая тетрадь Минаева, Рослова
ГДЗ номер 1243 физика 7‐9 класс сборник задач Лукашик, Иванова
ГДЗ вправа 961 алгебра 7 класс Тарасенкова, Богатырева
ГДЗ контрольная работа / №4 / Вариант 4 1 алгебра 8 класс дидактические материалы Жохов, Макарычев
ГДЗ тетрадь №1. страница 72 математика 4 класс рабочая тетрадь Гейдман, Мишарина
ГДЗ номер 788 алгебра 9 класс Никольский, Потапов
ГДЗ параграф 6 6.5 алгебра 8 класс рабочая тетрадь Зубарева, Мильштейн
ГДЗ § 10 4 химия 8 класс рабочая тетрадь Еремина, Кузьменко
ГДЗ упражнение 693 геометрия 7 класс Мерзляк, Полонский
ГДЗ тема 17. умножение на числа, оканчивающиеся нулями 7 математика 4 класс рабочая тетрадь Устный счет Рудницкая
умножение на числа, оканчивающиеся нулями 7 математика 4 класс рабочая тетрадь Устный счет Рудницкая
ГДЗ упражнение 290 алгебра 9 класс Мерзляк, Полонский
ГДЗ упражнение 117 русский язык 5 класс Русская речь Никитина
ГДЗ страница 56 английский язык 5 класс Несвит
ГДЗ вправа 337 алгебра 8 класс Тарасенкова, Богатырева
ГДЗ номер 769 геометрия 8 класс Мерзляк, Полонский
ГДЗ §13 13.5 алгебра 9 класс Учебник, Задачник (2018) Мордкович, Семенов
ГДЗ страница 114 русский язык 1 класс Канакина, Горецкий
ГДЗ упражнение 77 английский язык 5‐9 класс ГолицынскийБ
ГДЗ упражнение 97 русский язык 7 класс Коновалова
ГДЗ самостоятельная работа / вариант 3 6 геометрия 9 класс дидактические материалы Гусев, Медяник
ГДЗ номер / § 1 12 алгебра 11 класс Никольский, Потапов
ГДЗ вопросы в параграфах 49 физика 10 класс Касьянов
ГДЗ упражнения 620 алгебра 9 класс Макарычев, Миндюк
ГДЗ часть 2 40 математика 3 класс Истомина
ГДЗ По Немецкому Языку 7 Горизонты
Алгебре проверочная работа 5 макарычев 8 класс :: liafacbucus
Для 8 класса. Дидактические материалы.8 класс. Жохов. Готовые Домашние Задания, Решебник по Алгебре 8 класс. Скачать: Контрольные работы по алгебре для 8 класса по учебнику Макарычев Ю. Н. Алгебраические дроби. Решебник по алгебре для 8 класса автора В. И. Жохова прекрасно подойдет. Дидактическиие материалы. ГДЗ и решебник для учебникаГДЗ решебник по алгебре 8 класс Жохов. Сумма и разность алгебраических дробей. Контрольные работы представлены в нескольких вариантах. Правильные ответы к дидактическим материалам содержит решебник по. В. И., Макарычев Ю. Н.решебник, ответы онлайн. Решебник по алгебре 8 класс Жохов, Макарычев дидактические. Авторы, издание: Л. А. Александрова при участии Мордкович А. ГКонтрольные за 1, 2, 3. К учебнику Мордкович А. Г. С ответами.5. ГДЗ ответы на вопросы к учебнику по алгебре 8 класс Макарычев Миндюк Нешков Суворова ФГОС от Путина. Контрольные работы на темы.
Дидактические материалы.8 класс. Жохов. Готовые Домашние Задания, Решебник по Алгебре 8 класс. Скачать: Контрольные работы по алгебре для 8 класса по учебнику Макарычев Ю. Н. Алгебраические дроби. Решебник по алгебре для 8 класса автора В. И. Жохова прекрасно подойдет. Дидактическиие материалы. ГДЗ и решебник для учебникаГДЗ решебник по алгебре 8 класс Жохов. Сумма и разность алгебраических дробей. Контрольные работы представлены в нескольких вариантах. Правильные ответы к дидактическим материалам содержит решебник по. В. И., Макарычев Ю. Н.решебник, ответы онлайн. Решебник по алгебре 8 класс Жохов, Макарычев дидактические. Авторы, издание: Л. А. Александрова при участии Мордкович А. ГКонтрольные за 1, 2, 3. К учебнику Мордкович А. Г. С ответами.5. ГДЗ ответы на вопросы к учебнику по алгебре 8 класс Макарычев Миндюк Нешков Суворова ФГОС от Путина. Контрольные работы на темы.
Просвещение онлайн. Контрольные. Алгебра: учебник для 8 класса. Дидактические материалы.8 класс. Жохов В. И. , Макарычев Ю. Н.решебник, ответы онлайн. Здесь вы найдете все контрольные работы по всем пройденным темам. Дидактическиие. Итоговая контрольная работа за курс 8 класса. Алгебра 8 класс Дидактические материалы Звавич. Вариант 1. ГДЗ по алгебре 8 класс Мордкович А. Г., Мордкович Т. Н.2.26 упражнение. Решебники для 8 го класса. В 8 м классе многие домашние задания. Контрольная работа 5. Жохов В. И., Макарычев Ю. Н., Миндюк Н. Г. ГДЗготовые домашние задания к учебнику Алгебра: учебник для 8 класса. Дидактические материалы. Скачать: Самостоятельные и контрольные работы по алгебре. Глазков Ю. А., Гаиашвили М. Я. Самостоятельная работа 5. ГДЗ по алгебре 8 класс Мордкович А. Г. Контрольные работы по алгебре в 8. Решебник готовое домашнее задание учебников и рабочих тетрадей предназначены для проверки в. Алгебра: учебник.
, Макарычев Ю. Н.решебник, ответы онлайн. Здесь вы найдете все контрольные работы по всем пройденным темам. Дидактическиие. Итоговая контрольная работа за курс 8 класса. Алгебра 8 класс Дидактические материалы Звавич. Вариант 1. ГДЗ по алгебре 8 класс Мордкович А. Г., Мордкович Т. Н.2.26 упражнение. Решебники для 8 го класса. В 8 м классе многие домашние задания. Контрольная работа 5. Жохов В. И., Макарычев Ю. Н., Миндюк Н. Г. ГДЗготовые домашние задания к учебнику Алгебра: учебник для 8 класса. Дидактические материалы. Скачать: Самостоятельные и контрольные работы по алгебре. Глазков Ю. А., Гаиашвили М. Я. Самостоятельная работа 5. ГДЗ по алгебре 8 класс Мордкович А. Г. Контрольные работы по алгебре в 8. Решебник готовое домашнее задание учебников и рабочих тетрадей предназначены для проверки в. Алгебра: учебник.
Итоговая контрольная. Решебник по алгебре для 8 класса автора В. И. Жохова прекрасно подойдет для подготовки школьников. Ответы по. Алгебра 8 класс. Поэтому ответы на контрольные работы по алгебре для 8 классов. Контрольные работы Алгебра 8 класс Попов. Решения и ответы к контрольным работам В. И. Жохов, Ю. Н. Макарычев, Н. Г. Миндюк.—16 е изд. Решебник к дидактическим материалам по алгебре 8 класс Жохов,. По. Преобразуйте выражение:. Контрольные работы по алгебре для 8 класса по учебнику Мордковича. Учебники для 8 го класса. Контрольные за 1, 2, 3. К учебнику Мордкович А. Г. С ответами.5. Преобразуйте выражение:. Решения и ответы к контрольным работам В. И. Жохов, Ю. Н. Макарычев, Н. Г. Миндюк. ГДЗ по алгебре 8 класс. ГДЗ и решебник для учебникаГДЗ решебник по алгебре 8 класс Жохов, Макарычев, Миндюк Просвещение онлайн. Контрольные работы. Макарычев, Миндюк.
Контрольные работы Алгебра 8 класс Попов. Решения и ответы к контрольным работам В. И. Жохов, Ю. Н. Макарычев, Н. Г. Миндюк.—16 е изд. Решебник к дидактическим материалам по алгебре 8 класс Жохов,. По. Преобразуйте выражение:. Контрольные работы по алгебре для 8 класса по учебнику Мордковича. Учебники для 8 го класса. Контрольные за 1, 2, 3. К учебнику Мордкович А. Г. С ответами.5. Преобразуйте выражение:. Решения и ответы к контрольным работам В. И. Жохов, Ю. Н. Макарычев, Н. Г. Миндюк. ГДЗ по алгебре 8 класс. ГДЗ и решебник для учебникаГДЗ решебник по алгебре 8 класс Жохов, Макарычев, Миндюк Просвещение онлайн. Контрольные работы. Макарычев, Миндюк.
Вместе с Алгебре проверочная работа 5 макарычев 8 класс часто ищут
контрольная работа по алгебре 8 класс макарычев ответы.
контрольная работа по алгебре 8 класс макарычев ответы 1 четверть.
дидактический материал по алгебре 8 класс макарычев.
контрольная работа номер 1 по алгебре 8 класс.
готовые контрольные работы по алгебре 8 класс макарычев.
контрольная работа по алгебре 8 класс 1 четверть.
дидактические материалы по алгебре 8 класс жохов макарычев миндюк.
итоговая контрольная работа по алгебре 8 класс ответы
Читайте также:
Тест по обществознанию в 8 классе по теме духовная сфера
Ответы на печатную тетрадь по геометрии 9 класса
Гдз по математике 6 класс зубарева мильштейн лепешонкова смотреть онлайн с
ГДЗ по Алгебре 8 класс Дидактические материалы Жохов
Помимо школьного учебника по алгебре, детям задаются домашние задания и по дидактическим материалам. Материал в 8 классе уже достаточно сложный, объемный и ученикам нужно приложить много усилия, чтобы овладеть всеми аспектами данного предмета. Но самостоятельно полностью разобраться с темами не под силу ни одному восьмикласснику. Им нужна дополнительная помощь. Такую помощь предоставляют Жохов В.И., Макарычев Ю.Н. и Миндюк Н.Г.
Но самостоятельно полностью разобраться с темами не под силу ни одному восьмикласснику. Им нужна дополнительная помощь. Такую помощь предоставляют Жохов В.И., Макарычев Ю.Н. и Миндюк Н.Г.
Решебник к дополнительным материалам восьмого класса имеет массу преимуществ:
• в нем детально разобраны все самостоятельные и контрольные работы, особенно тщательно проработаны ответы на итоговое повторение;
• материал из этого сборника ГДЗ по алгебре за 8 класс дидактические материалы Жохов подобран в соответствии с предоставляемыми требованиями и довольно доступно изложен, как для самого восьмиклассника, так и для его родителей;
• он содержит большое количество разнообразных примеров решений, что научит школьников решать любые задания при проверке знаний;
• для одинаковых заданий этот сборник ответов имеет несколько примеров решения, что поможет ученикам найти оптимальный для них способ решения;
Пособие представлено пятью главами. Здесь рассматриваются решения к двум вариантам самостоятельной работы, каждый из которых содержит по 57 вопросов. Далее рассмотрены ответы на десять контрольных работ, в каждой из них дано по четыре варианта. Завершается данный решебник итоговым повторением тем по учебникам А. Тихонова и С. Теляковского.
Здесь рассматриваются решения к двум вариантам самостоятельной работы, каждый из которых содержит по 57 вопросов. Далее рассмотрены ответы на десять контрольных работ, в каждой из них дано по четыре варианта. Завершается данный решебник итоговым повторением тем по учебникам А. Тихонова и С. Теляковского.
Очень большую помощь окажет такое издание для восьмиклассников, планирующих участвовать в школьных олимпиадах. Ведь здесь не меньше внимания уделяется и на решения усложненных заданий.
ГДЗ по алгебре 8 класс Макарычев можно посмотреть здесь.
ГДЗ к рабочей тетради по алгебре за 8 класс Миндюк Н.Г. можно посмотреть здесь.
ГДЗ к тематическим тестам по алгебре за 8 класс Дудницын Ю. П. можно посмотреть
здесь.
П. можно посмотреть
здесь.
ГДЗ к дидактическим материалам по алгебре за 8 класс Звавич Л.И. можно посмотреть здесь.
▶▷▶ контрольные по алгебре 8 класс макарычев с ответами и решением
▶▷▶ контрольные по алгебре 8 класс макарычев с ответами и решениемконтрольные по алгебре 8 класс макарычев с ответами и решением — Yahoo Search Results Yahoo Web Search Sign in Mail Go to Mail» data-nosubject=»[No Subject]» data-timestamp=’short’ Help Account Info Yahoo Home Settings Home News Mail Finance Tumblr Weather Sports Messenger Settings Yahoo Search query Web Images Video News Local Answers Shopping Recipes Sports Finance Dictionary More Anytime Past day Past week Past month Anytime Get beautiful photos on every new browser window Download Контрольные работы по алгебре 8 класс Макарычев Решения и wwwmcvouoru/homework/books/4/ 8 /838htm Cached Дидактические материалы по алгебре для 8 класса Решения и ответы к контрольным работам В И Жохов, ЮН Макарычев , НГ Миндюк —16-е изд Контрольные работы | Решебник (ГДЗ) Алгебра 8 класс ВИ vklasseonline … Алгебра vklasseonline — это портал, на котором ты сможешь найти учебники и решебники (ГДЗ) по всем предметам школьной программы для разных классов Контрольные работы Алгебра 8 класс Макарычев — Глазков скачать wwwyangteacherru 8 класс Алгебра Контрольные работы созданы в соответствии с учебной программой по Алгебре 8 класса Контрольные По Алгебре 8 Класс Макарычев С Ответами И Решением — Image Results More Контрольные По Алгебре 8 Класс Макарычев С Ответами И Решением images Контрольные работы по алгебре 8 класс по учебнику Макарычева ЮН infourokru/kontrolnie-raboti-po-algebre-klass Cached › Другие методич материалы › Контрольные работы по алгебре 8 класс по учебнику Макарычева ЮН Контрольные работы по алгебре 8 класс по учебнику Макарычева ЮН Контрольные работы по алгебре 7 класс с ответами и решением dankonoycom/ege2016/kontrolnye-raboty-po Cached Контрольные работы по алгебре 7 класс с ответами и решением макарычев скачать 28112016 Алгебра 7 класс Комментарии: 0 Контрольные работы Алгебра 8 класс Мордкович — Попов wwwyangteacherru 8 класс Алгебра Контрольные работы созданы в соответствии с учебной программой по Алгебре 8 класса ГДЗ решебник по алгебре 8 класс контрольные работы Александрова gdzputinaco 8 класс Алгебра Онлайн ответы из решебника по алгебре за 8 класс автора Александровой ЛА 2009 года издания ГДЗ по алгебре за 8 класс Мордкович, решебник задачника gdzputinainfo/reshebniki/ 8 -klass/algebra/mordkovich Cached Решебник от Путина по алгебре 8 класс Мордкович с полным решением В 8 классе учебная программа алгебры комплексна и содержательна Решение контрольных и самостоятельных работ по алгебре за 9 30schoolru/9-klass/algebra/didakticheskie-materialy Cached Решение контрольных и самостоятельных работ по алгебре за 9 класс к пособию «Алгебра Дидактические материалы 9 класс » Макарычев ЮН Контрольные 8 класс, по алгебре, Мордкович за 1, 2, 3, 4 четверти mathematics-testscom/algebra- 8 -klass/kontrolnye Cached Контрольная работа №1 «Основные понятия и свойство алгебраической дроби», «Сложение и вычитание алгебраических дробей с одинаковыми и разными знаменателями» Promotional Results For You Free Download | Mozilla Firefox ® Web Browser wwwmozillaorg Download Firefox — the faster, smarter, easier way to browse the web and all of Yahoo 1 2 3 4 5 Next 29,800 results Settings Help Suggestions Privacy (Updated) Terms (Updated) Advertise About ads About this page Powered by Bing™
контрольные по алгебре 8 класс макарычев с ответами и решением — Все результаты Контрольные работы по алгебре 8 класс Макарычев Решения и @topic=844htm Дидактические материалы по алгебре для 8 класса Решения и ответы к контрольным работам ВИ Жохов, ЮН Макарычев , НГ Миндюк —16-е изд Алгебра 8 Контрольные (Макарычев ЮН) | Контроль знаний контрользнанийрф/algebra-8-kontrolnye-makarychev-yu-n/ 9 авг 2018 г — Тест по по теме » Алгебра 8 Контрольные ( Макарычев ЮН)» Решения задач контрольных работ из учебного пособия: « Алгебра 8 класс СА Теляковского Ответы на контрольные работы адресованы родителям, которые смогут проконтролировать правильность выполнения задания Видео 7:58 Контрольная работа ответы решение Алгебра 8 класс Лиса YouTube — 14 мая 2017 г 9:22 Контрольная работа Алгебра 8 класс Лиса YouTube — 23 нояб 2017 г 16:24 Контрольная работа по алгебре 8 класс Огэ 9 класс по математике Лиса YouTube — 14 мая 2017 г Все результаты Контрольные работы по алгебре 8 класс по учебнику Макарычева › Математика Похожие 10 июн 2016 г — Cкачать: Контрольные работы по алгебре 8 класс по учебнику Контрольная работа №6 « Решение дробных рациональных Контрольные работы по алгебре 8 класс — Инфоурок › Математика Похожие Контрольные работы по алгебре в 8 Контрольная работа №1 8 класс Контрольная работа №2 8 класс Рациональные выражения 1 вариант 1 Подробные и обоснованные решения заданий этой части напишите При выполнении заданий 1 – 11 в строке ответов обведите номер, который ГДЗ по алгебре 8 класс дидактические материалы Жохов › Алгебра › 8 класс Похожие Решебник по алгебре за 8 класс авторы Жохов, Макарычев , Миндюк издательство Просвещение ГДЗ дидактические материалы по алгебре 8 класс Жохов, Макарычев , Миндюк Просвещение Помимо готовых ответов , в решебнике детально излагается весь ход решения задачи Контрольные работы Контрольные 8 класс, по алгебре, Мордкович за 1, 2, 3, 4 четверти Рейтинг: 4 — 13 голосов 4 апр 2017 г — Контрольные работы по алгебре для 8 класса по учебнику Мордковича Контрольные за 1, 2, Ответы на контрольную работу №1: «Основные понятия и свойство Графическое решение квадратных уравнений Итоговая контрольная работа по алгебре 8 класс с ответами vkcom/page-64185229_46700734 Название: Итоговая контрольная работа по алгебре 8 класс с ответами макарычев 18 вопросов Лицензия: Полная Операционная система: WindowsXP ГДЗ Алгебра 8 класс ЛА Александрова, АГ Мордкович 2010 mydomashkaru/gdz/8-klass-algebra/la-aleksandrova-ag-mordkovichhtml Похожие Самые правильные ответы вы найдете здесь Конечно, решение для контрольной по алгебре за 8 класс можно найти и при помощи мобильного ГДЗ по Алгебре за 8 класс: Дидактические материалы Жохов ВИ › ГДЗ › 8 класс › Алгебра › дидактические материалы Жохов Решебник по алгебре для 8 класса автора ВИ Жохова прекрасно подойдет для подготовки школьников Ответы по самостоятельным и контрольным ГДЗ к дидактические материалы по алгебре 8 класс Жохов › › 8 класс › Алгебра › дидактические материалы Жохов Похожие Решение контрольных и самостоятельных работ по алгебре 8 касс Правильные ответы к решебники ГДЗ учебник алгебра 8 класс ЮН Макарычев К-4 — Контрольные работы | Решебник (ГДЗ) Алгебра 8 класс ВИ › › Контрольные работы К-4 — Контрольные работы Полный и качественный решебник (ГДЗ) Алгебра 8 класс ВИ Жохов, ЮН Макарычев , НГ Миндюк 2012 Дидактические Входная контрольная работа по алгебре 8 класса — Мультиурок 15 окт 2017 г — Входная контрольная работа по алгебре 8 класс Преподавание ведётся по учебнику ЮН Макарычева Самостоятельные и контрольные работы по алгебре: 8 класс: к › › 8 класс › Алгебра Самостоятельные и контрольные работы по алгебре : 8 класс : к учебнику В сборник включены также ответы к заданиям, рекомендации по подсчету Учебная программа УМК Макарычева Решение контрольных и само 80 контрольные работы по алгебре 8 класс — Социальная сеть 19 сент 2013 г — Главная · Группы · Мой мини-сайт · Ответы на часто задаваемые вопросы · Поиск по сайту · Сайты Методическая разработка по алгебре ( 8 класс ) по теме: контрольных работ по алгебре для учащихся 8 класса по учебнику Макарычева Найдите целые решения системы неравенств Ответы@MailRu: Решение контрольной работы по алгебре макарычев 8 › Образование › Школы Похожие 3 ответа 17 окт 2017 г — Пользователь ALEX_KUPER задал вопрос в категории Школы и получил на него 3 ответа кто нибудь напишите решение годовой контрольной 1 ответ 16 мая 2011 г Контрольная работа по алгебре 1 ответ 29 окт 2010 г Другие результаты с сайта otvetmailru Книга: «Алгебра: 8 класс: Ответы и решения к дидактическим › › Математика › Математика (5-9 классы) Книга: Алгебра : 8 класс : Ответы и решения к дидактическим материалам ВИ Жохова и др и контрольных работ из дидактических материалов по алгебре для 8 класса (ВИ Жохов, ЮН Макарычев , НГ Миндюк), издаваемого [PDF] Итоговая контрольная по математике 8 класс — Школа № 1191 sch2191szmskobrru/files/%20контрольная%20для%208%20классаpdf Похожие Итоговая контрольная работа по математике, 8 класс школа №1191 2015- 2016 Учебник: Алгебре 8 класс / ЮН Макарычев , НГ Миндюк, КН Нешков , СБ Она содержит задания с кратким ответом С помощью элементов содержания (понятий, их свойств, приёмов решения задач и тд), владение Картинки по запросу контрольные по алгебре 8 класс макарычев с ответами и решением «id»:»0ScFv7swwI2uJM:»,»ml»:»600″:»bh»:90,»bw»:61,»oh»:2082,»ou»:» «,»ow»:1435,»pt»:»fs00infourokru/images/doc/79/96045/hello_html_m4″,»rh»:»infourokru»,»rid»:»3lzl1X5OysSOAM»,»rt»:0,»ru»:» «,»sc»:1,»st»:»Инфоурок»,»th»:100,»tu»:» \u003dtbn:ANd9GcSNIjex8CSo9GdkYVeCPA1fn5OqKq4tzHrzrRDONhkXeOiaXLnQfGGcDA»,»tw»:69 «cr»:3,»id»:»XjCjtCqmsC1mDM:»,»ml»:»600″:»bh»:90,»bw»:90,»oh»:2657,»ou»:» «,»ow»:2782,»pt»:»arhivurokovru/kopilka/uploads/user_file_54362d38d»,»rh»:»kopilkaurokovru»,»rid»:»bwXzXoqKT0hYmM»,»rt»:0,»ru»:» «,»sc»:1,»th»:94,»tu»:» \u003dtbn:ANd9GcQ9MfHqnQK9cu5Qf2lfy4IOrZkrtcLQg3zQAES0h28W75_dmhND8qqxdA»,»tw»:99 «id»:»piA6_s_aDNpe3M:»,»ml»:»600″:»bh»:90,»bw»:107,»oh»:2178,»ou»:» «,»ow»:2669,»pt»:»arhivurokovru/kopilka/uploads/user_file_54362d38d»,»rh»:»kopilkaurokovru»,»rid»:»bwXzXoqKT0hYmM»,»rt»:0,»ru»:» «,»sc»:1,»th»:90,»tu»:» \u003dtbn:ANd9GcQ9i1mn6r5EYCS_sOr-EBH52ii_5dYjtzf9MJowKpFemlZEl8bIY0y_SVY»,»tw»:110 «id»:»I_DER7GCwC6XGM:»,»ml»:»600″:»bh»:90,»bw»:93,»oh»:2513,»ou»:» «,»ow»:2644,»pt»:»arhivurokovru/kopilka/uploads/user_file_54362d38d»,»rh»:»kopilkaurokovru»,»rid»:»bwXzXoqKT0hYmM»,»rt»:0,»ru»:» «,»sc»:1,»th»:94,»tu»:» \u003dtbn:ANd9GcRh2RdpyEkVcNiUnp-lCOKkXu1BYNDACSS-MPPc0STM5R-S8pSut1L1Xg»,»tw»:99 «cr»:3,»id»:»tAmhMGA7zWK5YM:»,»ml»:»600″:»bh»:90,»bw»:112,»oh»:2130,»ou»:» «,»ow»:2788,»pt»:»arhivurokovru/kopilka/uploads/user_file_54362d38d»,»rh»:»kopilkaurokovru»,»rid»:»bwXzXoqKT0hYmM»,»rt»:0,»ru»:» «,»sc»:1,»th»:90,»tu»:» \u003dtbn:ANd9GcQ8WlXDNZGGS78mNBECM5QQtJNU-MxTzDPCAVAQ93E_bd8WkrDGVQZVDw»,»tw»:118 «id»:»23FYd9x9lFrKIM:»,»ml»:»600″:»bh»:90,»bw»:117,»oh»:2011,»ou»:» «,»ow»:2717,»pt»:»arhivurokovru/kopilka/uploads/user_file_54362d38d»,»rh»:»kopilkaurokovru»,»rid»:»bwXzXoqKT0hYmM»,»rt»:0,»ru»:» «,»sc»:1,»th»:90,»tu»:» \u003dtbn:ANd9GcQjMB05OmSmeta-mmJVI7jelaeCM61Cfpz7uJHSlwlVRvf7hOKkyuS25Q»,»tw»:122 Другие картинки по запросу «контрольные по алгебре 8 класс макарычев с ответами и решением» Жалоба отправлена Пожаловаться на картинки Благодарим за замечания Пожаловаться на другую картинку Пожаловаться на содержание картинки Отмена Пожаловаться Все результаты Контрольные работы по алгебре, 8 класс — Документ — Gigabazaru Контрольные работы по алгебре , 8 класс Алгебра -8 Часть 1 Учебник; часть 2 Задачник Мнемозина, 2007 8 класс в) решения неравенства ГДЗ по алгебре 8 класс Мордкович — онлайн решебник — uchimorg ГДЗ по алгебре 8 класс Мордкович — онлайн решебник Домашняя контрольная работа № 1 §23 Графическое решение квадратных уравнений Если готовые онлайн ответы вам подошли, сохраните ссылку на страницу Итоговая контрольная работа по алгебре 8 класс — математика 22 мая 2016 г — итоговая контрольная работа по алгебре 8 класс Задания взяты из из 8) Найдите значение по графику функции , изображенному на рисунке « итоговая работа по алгебре бланк ответов » Мордкович АГ) Контрольные работы по учебникам «Алгебра 8кл», ЮН « Алгебра 8кл», ЮН Макарычев , НГ Миндюк УМК «Геометрия 7 9 кл», ЛС Атанасян, БФ работа 10 по теме «Применение подобия к решению задач » Контрольная работа 29 Итоговая контрольная работа по математике 8 класс Задания с кратким ответом по геометрии Задание Решите задание [DOC] МАТЕРИАЛЫ ДЛЯ ПРОМЕЖУТОЧНОЙ АТТЕСТАЦИИ ПО www74444s009edusiteru/DswMedia/promejattestmatem8kl2016-demo-docx Похожие Данная контрольная работа предназначена для учащихся 8 класса (Авторы учебников: алгебра — Макарычев ЮН и др, геометрия – Атанасян ЛС и др ) б) указаны номера 2-х и более ответов , даже если среди них указан и номер При решении заданий данной работы необходимо уметь применять Контрольные работы по алгебре 7 класс по учебнику авторов: ЮН pedsovetsu › › Математика, алгебра, геометрия › Оценка знаний учащихся Похожие 27 дек 2009 г — Контрольные работы по алгебре 7 класс по учебнику авторов: При каком значении переменной значение выражения 4а + 8 на 3 Всего приводится девять вариантов контрольных работ к учебнику Макарычева за 7 класс можно ответы и решения ??? а то не понимаю как решить Глазков ЮА, Гаиашвили МЯ Самостоятельные и контрольные › › Контроль результатов освоения программы СА Теляковского Алгебра 8 класс Пособие является необходимым дополнением к школьному учебнику ЮН Макарычева и др Решение неравенств с одной переменной и их систем Ответы к контрольным работам Контрольные работы по математике — 8 класс — Математика Департаментская контрольная работа по математике для 8 класса Также присутствуют ответы к контрольной работе и критерии выставления оценки 2) требуется применить знания при решении задач и записать ответ; Гдз алгебра 8 класс мордкович контрольные работы — читать 17 сент 2018 г — Страница поста Гдз алгебра 8 класс мордкович контрольные В конце сборника ответов вы найдёте решение к пяти контрольным Алгебра 8 класс Контрольные измерительные — My-shopru 51,00 ₽ — В наличии Пособие является важным дополнением к учебнику ЮН Макарычева и др » Алгебра 8 класс » (издательство «Просвещение»), рекомендованному В сборнике приведены также ответы к заданиям, рекомендации по подсчету ГДЗ, Ответы по Алгебре 8 класс Дидактическиие материалы 15 дек 2014 г — Готовые Домашние Задания, Решебник по Алгебре 8 класс Дидактическиие материалы Макарычев ЮН 2015 г В 8-м классе многие домашние задания, самостоятельные и контрольные работы, учителя дают детям не из все решения на задания, которые входят в учебник « Алгебра » Итоговый тест по алгебре, 8 класс — Урокрф 16 мая 2017 г — Учебно-дидактические материалы по Алгебре для 8 класса по УМК Макарычев ЮН, Миндюк НГ и др ИТОГОВАЯ КОНТРОЛЬНАЯ РАБОТА ЗА КУРС 8 КЛАССА При выполнении работы ответы на задания заносятся в бланк Решения заданий части 2 и 3 необходимо записать в кратком 8 класс Дидактика, решебники, поурочка, пособия купить в Москве › Москва › Книги и журналы 9 окт 2018 г — Ответы и решения 8 класс к заданиям учебника по алгебре ЮН и контрольные работы по алгебре к учебнику ЮН Макарычева 8 Решебник к сборнику контрольных работ по алгебре для 8 класса 21 янв 2015 г — Решения контрольных работ по алгебре из сборника для 8 класса Александровой Л А Рукопись — 2015 Настоящее пособие содержит Самостоятельные и контрольные работы по алгебре 8 класс К allengorg/d/math/math2209htm Скачать: Самостоятельные и контрольные работы по алгебре 8 класс 8 класс К учебнику Макарычева ЮН и др Глазков ЮА, Гаиашвили МЯ В сборник включены также ответы к заданиям, рекомендации по подсчету баллов и выставлению Решение неравенств с одной переменной и их систем 86 Алгебра 8 класс Самостоятельные и контрольные работы К В книжном интернет-магазине OZON можно купить учебник Алгебра 8 К учебнику Ю Н Макарычева и др от издательства Экзамен Кроме В сборник включены также ответы к заданиям, рекомендации по +7 495 730- 67-67 — круглосуточно FAQ in english © 1998 – 2018 ООО «Интернет Решения » Входная контрольная работа Математика 8 класс, 1,2 вариант novosarinskayashkolahcru/indexphp?option=com_contenttask=view Похожие 8 окт 2014 г — Входная контрольная работа Математика 8 класс , 1,2 вариант В каждом задании сначала запишите номер выполняемого задания, а затем полное решение и ответ Ответы к вариантам контрольной работы (алгебре) для 8 класса — MathSolutionru wwwmathsolutionru/book-list/math/8 Похожие тесты и контрольные работы с ответами по математике ( алгебре ) для 8 класса Дидактические материалы к учебнику Макарычева ЮН — Звавич Л И, Решение контрольных и самостоятельных работ (ГДЗ) по алгебре за 8 ГДЗ по алгебре для 8 класс от Путина Похожие Алгебра 8 класс сборник задач и контрольных работ авторы: АГ Мерзляк В Б Алгебра 8 класс Макарычев (углубленный уровень) Алгебра 8 класс ГДЗ (решебник) по алгебре 8 класс Макарычев, Миндюк, Нешков reshatorru/8-klass/algebra/makarychev/ Открыть ГДЗ по алгебре 8 класс Макарычев ! Там ученик найдет подробное решение уравнений и неравенств, корректные ответы на контрольные Алгебра 8 класс Контрольные измерительные материалы ФГОС shkolarossiiru/111103 Пособие является важным дополнением к учебнику ЮН Макарычева и др » Алгебра 8 класс » (издательство «Просвещение»), рекомендованному Мин В сборнике приведены также ответы к заданиям, рекомендации по подсчету решения и ответы , шпаргалки · Сборники задач, заданий, самоучители Алгебра 7 класс Мордкович 157 Домашняя контрольная работа No 7 159 Глава 8 ФУНКЦия у = x? $ 37 Функция y 161 $ 38 Графическое решение уравнений Решебник по Алгебре за 9 класс ЮН Макарычев, НГ Миндюк, КИ ГДЗ к рабочей тетради по Алгебре 9 класс Макарычев , Миндюк можно 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 Алгебра 9 класс Мордкович А Г 2 часть задачник 25 Домашняя контрольная работа No 1 31 Методы решения систем уравнений ЧИСЛОВЫЕ ФУНКЦИИ $ 8 142 Ответы МетаШкола Дистанционные кружки и олимпиады для школьников Похожие Еженедельные занятия в течение учебного года в кружках по олимпиадным программам: математика , физика, русский язык, английский язык, немецкий ГДЗ (решебник) Алгебра 8 класс Ю П Дудницын, Е Е Тульчинская › Моя Школа › ГДЗ › 8 класс › Алгебра Рейтинг: 5 — 3 голоса Контрольная работа № 7 В чем особенность ГДЗ по алгебре за 8 класс Ю П Дудницын не только правильно провести срез знаний, проверить верность решения , но и понять принцип решения той или иной задачи Кияни · Деньги · Политика · Жизнь столицы · Криминал · Книга жалоб · Ответы власти Контрольная работа №1, Вариант 1 Задание №8 Алгебра 8 класс wwwmathcomua//algebra-8-klass/mordkovich_kontrolnaya_rabotahtml?8 Похожие ГДЗ по алгебре 8 класс Мордкович Домашняя контрольная работа №1 Вариант 8 Упростите выражение Решение задания №8: Il Оцените это ГДЗ: Онлайн Решебник ГДЗ Spotlight 11 класс — Reshakru Похожие Решебник по английскому языку Spotlight 11 класс станет вашей отличной помощью в учебе При помощи его вы забудете, что такое домашнее Онлайн Решебник ГДЗ Spotlight 10 класс — Reshakru Решебник Spotlight 10 класс — отличная помощь школьнику в решении домашнего задания по английскому языку Вы можете использовать решебник в Решебник по алгебре контрольные работы 8 класс мордкович school2vpru/?oue=reshebnik-po-algebre-kontrolnie-raboti-8-klass-mordkovich ГДЗ по алгебре 8 класс Мордкович — онлайн решебник а сборник ГДЗ предлагает готовые решения всех этих заданий Но большая часть детей просто списывает готовые ответы по алгебре Мордкович для 8 класса , чтобы Алгебра 8 класс ГДЗ ответы — GDZ-spishycom Алгебра 8 класс — онлайн решебники и ГДЗ 8 класс Мордкович Решебник по алгебре 8 класс Мордкович , ответы ൠ решения задач на ➄, ГДЗ ✹ для ❤ рабочих тетрадей ЗДЕСЬ ➜ Контрольные работы Александрова 8 класс Пояснения к фильтрации результатов Мы скрыли некоторые результаты, которые очень похожи на уже представленные выше (50) Показать скрытые результаты В ответ на официальный запрос мы удалили некоторые результаты (1) с этой страницы Вы можете ознакомиться с запросом на сайте LumenDatabaseorg Вместе с контрольные по алгебре 8 класс макарычев с ответами и решением часто ищут контрольная работа по алгебре 8 класс ответы контрольные работы по алгебре 8 класс макарычев скачать бесплатно итоговая контрольная работа по алгебре 8 класс ответы контрольная работа по алгебре 8 класс номер 4 контрольная работа по алгебре 8 класс рациональные дроби контрольная работа по алгебре 8 класс рациональные дроби решение контрольная работа по алгебре 8 класс мерзляк гдз по алгебре 8 класс макарычев Ссылки в нижнем колонтитуле Россия — Подробнее… Справка Отправить отзыв Конфиденциальность Условия Аккаунт Поиск Карты YouTube Play Новости Почта Контакты Диск Календарь Google+ Переводчик Фото Ещё Документы Blogger Hangouts Google Keep Подборки Другие сервисы Google
Доктор Стрейнджлав: открытие (Жоховские острова) — сцена 6 (профиль атаки) Сводка и анализ
Сцены: Открытие, скалистые вершины Жоховских островов — титры — состояние красный — Воздушно-десантные B-52 — план атаки крыла R — Спальня Тургидсона — «Твой коммуняк» — профиль нападения.
Сводка
Фильм открывается мрачным кадром горных вершин, возвышающихся над морем облаков, в то время как рассказчик объясняет слухи о том, что Советский Союз работал над устройством судного дня.Мы предполагаем, что это вершины Жоховских островов, которые рассказчик упоминает как место реализации проекта судного дня. Кажется, что снимок сделан с самолета, приближающегося к островам, и на заднем плане слышен проносящийся мимо ветер. Рассказчик принимает зловещий тон, не может дать много информации и говорит в прошедшем совершенном времени и прошедшем простом времени. Затем фильм переходит к титру, показанному на монтажном кадре B-52 ВВС США, занятого дозаправкой в воздухе, в то время как идет игра «Попробуй немного нежности».
Затем фильм переходит к серии снимков экстерьера базы ВВС Берпельсон, после чего капитан группы Лайонел Мандрейк показывается внутри комнаты связи. Мандрагору вызывают к телефону, чтобы ответить на звонок генерала Джека Д. Риппера, командира базы. Потрошитель, изображенный на другом конце линии и сидящий перед плакатом с надписью «мир — наша профессия», дает Мандрагоре инструкции перевести базу в красный цвет, опечатать базу и изъять все радиоприемники.Он впечатляет Мандрагоры серьезностью и важностью ситуации, говоря ему, что началась перестрелка, и заставляет Мандрагору передать «План атаки R» 843-му бомбардировочному крылу, которым командует база. Мандрагора понимает значение плана R (ядерное нападение на Советский Союз) и огорчается, спрашивая Риппера, «действительно ли ситуация настолько плоха».
Риппера, командира базы. Потрошитель, изображенный на другом конце линии и сидящий перед плакатом с надписью «мир — наша профессия», дает Мандрагоре инструкции перевести базу в красный цвет, опечатать базу и изъять все радиоприемники.Он впечатляет Мандрагоры серьезностью и важностью ситуации, говоря ему, что началась перестрелка, и заставляет Мандрагору передать «План атаки R» 843-му бомбардировочному крылу, которым командует база. Мандрагора понимает значение плана R (ядерное нападение на Советский Союз) и огорчается, спрашивая Риппера, «действительно ли ситуация настолько плоха».
На снимке бомбардировщика B-52 в полете с сопровождением рассказчик объясняет операцию Dropkick (хотя мы узнаем это имя только намного позже в фильме), в которой Стратегическое воздушное командование США поддерживает в воздухе силы бомбардировщиков 24 часа в сутки. день, вооруженный ядерными зарядами.Затем он переходит в салон ВВС США B-52, где майор Т.Дж. «Кинг» Конг и его команда скучают, перекусывают, читают Playboy и играют в карты. Мы предполагаем, что они были в воздухе в течение некоторого времени в составе Dropkick и привыкли летать по кругу, предназначенные только для сдерживания и никогда не ожидающие доставки своей полезной нагрузки.
Мы предполагаем, что они были в воздухе в течение некоторого времени в составе Dropkick и привыкли летать по кругу, предназначенные только для сдерживания и никогда не ожидающие доставки своей полезной нагрузки.
Внезапно оживает радиопередатчик и передает код, который коммуникатор расшифровывает как план атаки крыла R. Никто на борту не верит этому, и Майор Конг сам возвращается к радиостанции, чтобы дважды проверить код.Они предполагают, что это означает, что Советы уже атаковали Вашингтон, округ Колумбия, и им приказывают принять ответные меры. Как только они получают подтверждение с базы, песня «When Johnny Comes Marching Home» играет на саундтреке, и Майор Конг переодевается из летного шлема в стетсонскую шляпу. Он произносит речь о серьезности и важности ядерной битвы, заверяя остальную часть своей команды, что это необходимо, и что все они получат повышение и поощрения по возвращении, независимо от их расы или происхождения.
Телефонный звонок в спальне генерала Бака Тургидсона, начальника штаба ВВС США. Тургидсон в ванной, и его секретарь и любовница мисс Скотт, которая загорает на кровати под светом и в бикини, отвечает на звонок. Разговор между Тургидсоном и полковником на другом конце линии, с мисс Скотт в качестве посредника, выкрикивает информацию через всю комнату, чтобы Тургидсон мог слышать. Ей дают слишком много секретной информации, и она передает ее слишком громко.Тургидсону рассказывают, что Риппер выпустил план атаки R для своего крыла и отключил все коммуникации с внешним миром, несмотря на отсутствие реальной угрозы для США. Тургидсон дает мисс Скотт сексуальное обещание, используя авиационные метафоры, прежде чем отправиться в Военную комнату.
Тургидсон в ванной, и его секретарь и любовница мисс Скотт, которая загорает на кровати под светом и в бикини, отвечает на звонок. Разговор между Тургидсоном и полковником на другом конце линии, с мисс Скотт в качестве посредника, выкрикивает информацию через всю комнату, чтобы Тургидсон мог слышать. Ей дают слишком много секретной информации, и она передает ее слишком громко.Тургидсону рассказывают, что Риппер выпустил план атаки R для своего крыла и отключил все коммуникации с внешним миром, несмотря на отсутствие реальной угрозы для США. Тургидсон дает мисс Скотт сексуальное обещание, используя авиационные метафоры, прежде чем отправиться в Военную комнату.
Вернувшись на базу, нам показывают кадры солдат, готовящихся защищать базу от нападения. Из громкоговорителя раздается голос Риппера, говорящего своим солдатам, что «ваш коммунист не заботится ни о человеческой жизни, ни о своей собственной», и, что наиболее важно, Советы могут прийти в военной форме США, чтобы обмануть их. Он издает приказ стрелять в любого, кто приближается в пределах 200 ярдов от базы. Когда Риппер заканчивает свою речь через громкоговоритель, есть кадр, на котором Мандрагора закрывает комнату связи, он встречает последнее радио, включает его и слышит, как играет джаз. Это гражданское вещание. Взволнованно, Мандрагора берет радио и выходит из комнаты.
Он издает приказ стрелять в любого, кто приближается в пределах 200 ярдов от базы. Когда Риппер заканчивает свою речь через громкоговоритель, есть кадр, на котором Мандрагора закрывает комнату связи, он встречает последнее радио, включает его и слышит, как играет джаз. Это гражданское вещание. Взволнованно, Мандрагора берет радио и выходит из комнаты.
Вернувшись к бомбардировщику, Майор Конг и его команда открывают профиль атаки, который запечатан до тех пор, пока им не понадобится его использовать. Конг зачитывает профиль атаки, пока он и остальная часть команды выполняют его инструкции.Он объясняет, что они должны включить дискриминатор CRM 114 радиостанции, который предотвращает попадание любых сообщений в самолет, если им не предшествует трехбуквенный префикс. Мы видим, что это префикс O-P-E. Они проверяют цепи самоуничтожения радио.
Анализ
Эта начальная группа сцен следует традиционной дуге «кризисного истеблишмента». Мы знакомимся с обстановкой, большинством центральных персонажей и основными темами фильма. Нам также показывают несколько изображений или детали, которые предвещают большую часть сюжета, в частности, вещи, которые вызовут проблемы, которые приведут к эскалации кризиса в дальнейшем в фильме. Фактически, многие характеристики и тематические изображения настолько явны и чрезмерны, что Кубрик, кажется, высмеивает саму драматическую структуру. Такие образы и образы представляют фильм как сатиру и комедию, несмотря на совершенно «прямую» игру и мрачную тематику.
Нам также показывают несколько изображений или детали, которые предвещают большую часть сюжета, в частности, вещи, которые вызовут проблемы, которые приведут к эскалации кризиса в дальнейшем в фильме. Фактически, многие характеристики и тематические изображения настолько явны и чрезмерны, что Кубрик, кажется, высмеивает саму драматическую структуру. Такие образы и образы представляют фильм как сатиру и комедию, несмотря на совершенно «прямую» игру и мрачную тематику.
В первой сцене использование рассказчиком прошедшего времени, кажется, намекает на то, что история рассказывается постфактум, а мрачность образов и тяжесть того, что рассказывает рассказчик, намекает на то, что произошло что-то ужасное.Этот голос за кадром дает зрителям информацию о машине судного дня, которой у персонажей, которых мы позже встретим, похоже, нет, и такое предзнаменование будет повторяться несколько раз на протяжении всего фильма. Это отражает позицию директора, а также позицию, которую многие придерживались в то время, что политические и военные лидеры каким-то образом упустили или отказались видеть очевидные недостатки в ядерной политике и стратегии обороны США. То, как рассказчик говорит о машине судного дня в этой вступительной сцене, также отражает страх и неуверенность, которые каждый зритель в то время чувствовал бы постоянно из-за хрупкого баланса мира и постоянной угрозы ядерного уничтожения.Начало фильма немного дезориентирует в контексте остальной части фильма, так как закадровый голос не доносится до конца фильма. Ощущение неуместности этой сцены отчасти может быть связано с тем, что она возникла в более старой версии сценария, в которой история рассказывается с точки зрения инопланетных существ, которые открывают Землю после ядерной катастрофы и пытаются ее обнаружить. узнать, что случилось.
То, как рассказчик говорит о машине судного дня в этой вступительной сцене, также отражает страх и неуверенность, которые каждый зритель в то время чувствовал бы постоянно из-за хрупкого баланса мира и постоянной угрозы ядерного уничтожения.Начало фильма немного дезориентирует в контексте остальной части фильма, так как закадровый голос не доносится до конца фильма. Ощущение неуместности этой сцены отчасти может быть связано с тем, что она возникла в более старой версии сценария, в которой история рассказывается с точки зрения инопланетных существ, которые открывают Землю после ядерной катастрофы и пытаются ее обнаружить. узнать, что случилось.
На протяжении всего фильма взрыв ядерного оружия и война в целом связаны с сексуальным удовлетворением.Последовательность титров раскрывает эту тему в изображении, которое ясно предполагает сексуальный акт между двумя самолетами, созданный для того, чтобы нанести ядерное разрушение. Как только титры заканчиваются, самолеты завершают свое действие и отталкиваются друг от друга, а соединяющий их «фаллос» расслабляется и опускается. Песня «Попробуй немного нежности», проигрываемая поверх последовательности, помогает в достижении цели.
Открытая и прямая критика ВВС США и военной стратегии того времени начинается сразу же в первой разыгранной сцене фильма.Когда генерал Риппер дает команду Мандрагоре опечатать базу и передать план атаки R, команду, которая приведет к ядерному уничтожению и массовым жертвам среди гражданского населения, он сидит перед плакатом ВВС США с надписью «мир — наша профессия». Здесь мы имеем дело с драматической иронией: ВВС США и их лидеры понимают, что они защищают и обеспечивают мир, тогда как мы знаем, что их политика прямо противоположна этому.
Закадровый голос, который следует в следующей сцене, создает ситуативную иронию с последующей сценой.Нам говорят, что патруль бомбардировщиков находится в состоянии боевой готовности, находится в воздухе 24 часа в сутки для защиты от ядерного нападения, и мы ожидаем увидеть шквал активности внутри бомбардировщика. Вместо этого нам показывают команду, которой скучно, она отвлечена и никогда не ожидает, что ей придется предпринимать какие-либо настоящие военные действия. Эта сцена указывает на абсурдный и противоречивый характер «холодной» войны: хотя бомбардировщик вооружен самым смертоносным оружием, известным человеку, и каждый должен опасаться постоянной угрозы ядерного уничтожения, отсутствие активных боевых действий ошеломляет участников. серьезности их положения и той «войны», которую они ведут.
Переход Майора Конга со шлема на стетсон знаменует его трансформацию в отношение к войне, а его техасские манеры высмеивают мачизм, господствовавший в то время в армии. Музыкальная реплика песни о гражданской войне «Когда Джонни идет маршем домой» демонстрирует внезапный переход от «холодной войны» к «горячей войне», несмотря на отсутствие каких-либо эффективных изменений среди солдат, кроме смены шляпы майора. Это подчеркивает легкость, с которой мир может перейти к полному опустошению, одновременно выявляя почти юмористический абсурд настоящей войны, вспыхнувшей так внезапно.Кроме того, речь Мэйджора Конга, заверяющая всех, что они получат повышение по возвращении домой, в соответствии с темой песни, является напоминанием о том, что никто на борту, скорее всего, не вернется домой. Комментарий о расе здесь является напоминанием о единственной недавней десегрегации вооруженных сил, а также о неизбирательном разорении и страданиях, с которыми мир столкнется от рук белых, мужчин, мачо, американского военного комплекса.
Наше знакомство с генералом Тургидсоном дополнительно раскрывает связь между военными и сексом в фильме и высмеивает ВВС США как некомпетентных и эгоцентричных.Обмен секретной информацией через мисс Скотт демонстрирует легкомыслие по отношению к секретности, которое идет вразрез с крайней секретностью, которую ВВС США и Советы будут использовать в других случаях, вплоть до катастрофы. Кроме того, отказ Тургидсона серьезно отнестись к звонку и передача мисс Скотт информации пустая трата времени, которое можно было бы потратить на решение проблемы. Трата времени по эгоистичным или глупым причинам будет повторяться на протяжении всего фильма. Использование Тургидсоном «обратного отсчета» и «взлета» при обсуждении секса с мисс Скотт подчеркивает связь между ракетами и сексуальной кульминацией, высмеивая одержимость военных ракетами как своего рода сексуальную фантазию.
Речь Риппера о коммунистах, приходящих в американскую форму, снова дает нам представление о его психозе и высмеивает то, как в то время крайние националисты США говорили о русских и коммунистах. Профиль атаки и переключение дискриминатора CRM 114 показывают нам многие детали, которые станут важными во время эскалации кризиса позже в фильме, включая схему самоуничтожения. Этот контур — пистолет Чехова — поскольку он показан нам, мы знаем, что позже он должен сыграть роль в разрушении радио и возникновении конфликта в фильме.
Предки современных сибирских собак сформировались в результате нескольких тысячелетий торговли по всей Евразии и расселения людей.
Значение
Сибирская Арктика пережила многочисленные социальные изменения с момента первого известного появления собак в этом регионе ∼10 000 лет назад. Эти изменения включают появление обработки железа около 2000 лет назад и появление оленеводства около 800 лет назад. Анализ 49 геномов древних собак показывает, что предки собак Арктической Сибири изменились за последние 2000 лет из-за притока собак из евразийской степи и Европы.В сочетании с геномными данными людей и археологическими данными наши результаты показывают, что, хотя происхождение человеческих популяций в Арктической Сибири не изменилось за этот период, люди там участвовали в торговле с отдаленными сообществами, в которых участвовали как собаки, так и материальная культура.
Abstract
Собаки были необходимы для жизни в сибирской Арктике более 9500 лет, и эта тесная связь между людьми и собаками продолжается в сибирских сообществах. Хотя арктические сибирские группы, такие как ненцы, получили ограниченный поток генов от соседних групп, археологические данные свидетельствуют о том, что металлургия и новые стратегии существования появились в Северо-Западной Сибири около 2000 лет назад.Неясно, была ли популяция сибирских арктических собак такой же постоянной, как и население региона, или вместо этого произошло смешение, возможно, в связи с притоком материальной культуры из других частей Евразии. Чтобы ответить на этот вопрос, мы секвенировали и проанализировали геномы 20 древних и исторических сибирских и евразийских степных собак. Наш анализ показывает, что, хотя сибирские собаки были генетически однородными между 9500 и 7000 лет назад, более позднее появление собак из евразийской степи и Европы привело к значительному смешению.Это явно имеет место в Ямало-Ненецком регионе (Северо-Западная Сибирь), где собаки периода железного века (~ 2000 лет назад) имеют значительно меньшее происхождение, связанное с европейскими и степными собаками, чем собаки средневекового периода (~ 1000 лет назад). . В сочетании с находками нелокальных материалов, извлеченных из этих археологических памятников, включая стеклянные бусы и металлические предметы, эти результаты показывают, что общины Северо-Западной Сибири были связаны с более крупной торговой сетью, через которую они приобретали генетически отличительных собак из других регионов.Эти обмены были частью ряда крупных социальных изменений, включая рост масштабного оленеводства около 800 лет назад.
Ранние археологические и геномные данные с острова Жохов в Арктической Сибири показывают, что собаки, принадлежащие к определенной линии, были важным компонентом жизни в Арктике на протяжении более 9500 лет (1, 2). Эта тесная связь между людьми и собаками сохраняется в сибирских общинах, таких как коряки, ительмены, чукчи и ненцы, где собак по-прежнему использовали для охоты, выпаса скота и катания на санях среди других видов деятельности (3-5).Недавние геномные данные, полученные в сообществах, говорящих на самоедском языке, таких как ненцы и селькупы, предполагают, что в течение голоцена они получали ограниченный поток генов от соседних групп, включая степных скотоводов (6, 7). Учитывая, что люди и их собаки часто мигрируют и взаимодействуют параллельно (8), возможно, что сибирские собаки также испытали ограниченный поток генов из других популяций.
В отличие от данных генома человека, лингвистические и этнографические данные предполагают более динамичные процессы.В частности, эти данные свидетельствуют о том, что говорящие на самоедском языке народы Северо-Западной Сибири мигрировали из южной Сибири или соседних регионов юго-восточной Европы в Арктику не далее как 3000–4000 лет назад (9⇓⇓ – 12). Кроме того, археологические памятники, такие как Усть-Полуй на северо-западе Сибири, демонстрируют свидетельства металлургии железа и бронзы и отдельные находки, такие как стеклянные бусы, которые, вероятно, были привезены из степи, Черного моря или Ближнего Востока (13–15). Наличие этой материальной культуры предполагает, что эти общины участвовали в широкомасштабных торговых сетях (13–15).Предлагаемая миграция и обмен материалами и практикой потенциально также касались собак, что могло привести к смешению, улучшению и, в конечном итоге, к созданию современных сибирских линий собак, таких как современная самоедская порода.
Чтобы оценить, была ли популяция арктических собак Северо-Западной Сибири непрерывной или вместо этого была отмечена примесь (возможно, в связи с притоком материальной культуры из других частей Евразии), мы секвенировали 20 древних и исторических сибирских и евразийских степных собак, расположенных в возраст от 11000 до 60 лет назад и геномный охват от 0.1 × и 11,1 × (набор данных S1). Затем мы проанализировали эти геномы вместе с общедоступными древними ( n = 29) и современными ( n = 120) псовыми (рис. 1 A и набор данных S2).
Рис. 1.( A ) Карта древних собак, включенных в исследование, с названием образца и возрастом (тыс. Лет) с картой Inset Ямало-Ненецкого автономного округа Северо-Западной Сибири. Данные из образцов, представленных кружками, были получены в этом исследовании со средним охватом генома от 0.1 и 19,9 ×; треугольники представляют общедоступных древних собак. Цвета точек данных представляют значение D-статистики формы D (черный шакал, выборка; Жохов, ASHQ01). Цвета с красным смещением показывают более близкое родство с ASHQ01 (древней ближневосточной собакой), а цвета с синим смещением указывают на более близкое родство с Жоховым (древней арктической собакой). ( B ) Филогения TreeMix с пятью краями миграции, которые обозначены серыми пунктирными линиями. Каждая популяция содержит от одной до трех особей ( SI Приложение , Таблица S1).Цвет ветвей соответствует среднему значению D-статистики в A . Полные модели с утяжелителями внешней группы и кромок, а также модели с дополнительными кромками можно найти в Приложении SI , рис. S3.
Результаты
Оценка широких предков у сибирских собак.
Сначала мы оценили, имеют ли сибирские собаки сходное происхождение, используя как анализ главных компонентов (PCA; SI, приложение , рис. S1), так и филогенетический анализ (рис.1 B и SI Приложение , рис. S2). Этот анализ обобщает результаты предыдущих исследований, показывающих, что современных и древних собак можно в общих чертах разделить на три основные группы: западноевразийские, восточноазиатские и арктические / американские (8, 16). Западно-евразийская линия включает древних ближневосточных и левантийских, современных африканских, древних и современных европейских, а также недавно секвенированных степных собак бронзового века (8, 16). Восточноазиатская линия состоит из современных собак из Китая, Вьетнама, островов Юго-Восточной Азии, динго и поющих собак Новой Гвинеи.Линия Арктики / Америки включает современные арктические породы, такие как гренландские ездовые собаки и сибирские хаски, древние американские собаки, собаки среднего голоцена с озера Байкал, 9500-летняя собака с острова Жохов, собаки из Ямало-Ненецкого региона и секвенирование исторических собак со всей Сибири для данного исследования (рис. 1 B ). Это указывает на то, что вновь секвенированные сибирские собаки в этом исследовании обладают генетическим происхождением, которое было непрерывным в течение как минимум 9500 лет.
Затем мы использовали TreeMix для оценки смешения этих линий.Эти анализы, основанные на 21 геноме древних и 14 современных собак, в целом повторяют наше дерево присоединения соседей (рис. 1 B и SI, приложение , рис. S2 и S3 A и B ) и подтверждают результаты предыдущих исследований (8, 17), показывающих, что происхождение европейских собак возникло в результате смешения ближневосточных и арктических собачьих родов еще 10,9 тыс. лет назад (тыс. лет назад). Наш расширенный набор данных показал, что образец с острова Жохова возрастом 9500 лет ближе всего к образцу 6000 лет из озера Байкал, а не к древним североамериканским собакам (18) ( SI Приложение , рис.S2 и S4 A ). Этот результат согласуется с разделением между древними американскими и сибирскими арктическими собаками и между расходящимися сибирскими линиями, которые старше калиброванной радиоуглеродной даты образца острова Жохов (~ 9,5 тыс. Лет назад) и с интродукцией конца плейстоцена или очень раннего голоцена (и последующего длительная изоляция) собак в Америке (18, 19).
Анализ опубликованного и вновь секвенированного генома из мезолитической стоянки Веретье I (∼10.9 тыс. Лет назад; рис.1) в Северо-Восточной Европе резюмируют предыдущие результаты, показывающие, что эти собаки имели родословную, связанную как с арктическими (66% и 71%), так и с западноевразийскими (34% и 29%) линиями (8, 17) (рис. 1 и 2, SI. Приложение , рисунки S3 A и B и S4 A и D, а также набор данных S3). Этот результат предполагает, что, как и все другие современные и древние (постмезолитические) европейские собаки, секвенированные на сегодняшний день, мезолитические собаки в Европе уже имели как арктическую, так и западноевразийскую родословную.Однако тот факт, что древняя сибирская собака с острова Жохова, датированная примерно 1000 лет назад после собак Верети, не имеет западно-евразийских предков, указывает на то, что западные собачьи предки не достигли Сибирской Арктики за 9,5 тыс. Лет назад.
Рис. 2.( A ) Схематическое изображение предопределенных моделей, подогнанных с помощью Admixturegraph для оценки происхождения сибирских / степных собак. ( B ) Тепловая карта на основе количества выбросов D-статистики для каждой модели. Наиболее подходящие модели не оставили никаких выбросов (темно-синие).Значения усредняются по четырем различным костякам, каждая из которых включает черного шакала (внешняя группа), древнего арктического (жоховская собака), древнего европейского происхождения (HXH или Newgrange) и древнего ближневосточного происхождения (TepeGhela или ASHQ01).
Установление происхождения степных собак бронзового века.
Отсутствие западно-евразийского происхождения у жоховской собаки не исключает возможности последующего притока собак в сибирскую Арктику из соседних регионов, таких как Европа, Ближний Восток или Степь.Чтобы проверить эту гипотезу, мы впервые установили происхождение степных собак бронзового века [5–3 тыс. Лет назад (20)], поскольку их происхождение не охарактеризовано, и они представляют собой потенциальный источник примеси у сибирских собак. Анализ одной общедоступной (Самара1, 0,7 ×) и одной недавно секвенированной степной собаки бронзового века (Ишкинино, 1,4 ×) показал, что они образуют генетически однородную группу, наиболее близкую к собакам Западной Евразии (Рис.1 A и SI. Приложение , рис. S1 и S5).Фактически, анализ TreeMix показал, что эти собаки демонстрируют сильное сходство с древними ближневосточными собаками ( SI Приложение , рис. S3 A и B ), а моделирование графа примесей показало, что степные собаки имели дополнительную ближневосточную родословную, большую, чем найденный у древних европейских собак (рис. 2). Кроме того, D-статистика не обнаружила признаков примеси из других популяций (в том числе от арктических собак) ни у одной из двух степных собак с момента их последнего общего предка (> 3.8 тыс. Лет назад) ( SI Приложение , рис. S5).
Дополнительное моделирование с использованием соотношений F4 показало, что происхождение степных собак можно смоделировать как ∼40% арктических (представленных собакой с острова Жохова) и ∼60% Ближнего Востока (представленных иранскими и левантийскими древними собаками), ссылка SI Приложение . Предыдущие исследования геномов древнего человека показали, что популяция, относящаяся к иранцам эпохи энеолита, составляла около 40% предков степных популяций бронзового века, вероятно, в результате экспансии сельского хозяйства с Ближнего Востока в степь (21).Хотя мы не можем исключить гипотезу о том, что степные собаки были завезены из Европы, наш анализ показывает, что, как и в Европе (8, 22), неолитическая экспансия сельского хозяйства с Ближнего Востока в степь также включала расселение собак. Анализ более позднего (0,7 тыс. Лет назад) генома степной собаки (Bolgar1) из средневекового города Болгар в Татарстане (Россия) показывает, что, хотя европейское происхождение увеличивалось со временем (Рис.1 A и SI Приложение , Рис. S2 и S4 A и D ), ближневосточное происхождение, вероятно, сохранялось у собак из степи, по крайней мере, до средневековья (рис.2 и SI Приложение , рис. S6 B и C ).
Тестирование на добавку у сибирских собак.
В отличие от степных собак бронзового века, наш анализ показал, что существуют различные степени родства (как показывает D-статистика) между древними и историческими сибирскими собаками с ближневосточными (западноевразийскими) и древними арктическими предками (рис. Наборы данных S1 и S4). Фактически, наш PCA показал, что все сибирские собаки, секвенированные в этом исследовании, находятся на грани между западноевразийскими и арктическими собаками ( SI, приложение , рис.S1), предполагая, что сибирские собаки имели разные уровни западноевразийского происхождения. Этот приток неарктических предков в сибирских собак мог быть результатом введения собак либо ближневосточного / степного происхождения, либо европейского происхождения, либо обоих. Проверить эти сценарии сложно, поскольку предыдущие исследования показали, что «европейское» происхождение само по себе является результатом смешения арктических и западно-евразийских (ближневосточных) предков (8).
Чтобы установить различные компоненты предков собак, которые смешивались с сибирскими собаками, датируются 10.9 и 0,06 тыс. Лет назад, мы определили шесть отдельных моделей (четыре из которых показаны на рис. 2 A ) с использованием пакета Admixture Graph R (23), в котором мы использовали представителей ближневосточных, европейских и арктических собак, и итеративно проверили, насколько каждая из моделей соответствует геномам 22 сибирских собак. Мы не включили восточноазиатских собак в этот анализ по двум причинам. Во-первых, археологические данные свидетельствуют о том, что сообщества Северо-Западной Сибири в основном взаимодействовали с сообществами из степи, Черного моря и Ближнего Востока (13–15).Во-вторых, результаты нашего собственного анализа ( SI Приложение , рис. S3 A и B , S7 и S8 и набор данных S4) и предыдущих исследований (8) показывают, что хотя арктические собаки (представленные древним Жоховым dog), возможно, внесли свой вклад в происхождение восточноазиатских собак, существует мало доказательств того, что поток генов в противоположном направлении ( SI Приложение ).
Для представления каждой из трех наследственных линий в моделях мы выбрали двух представителей с Ближнего Востока (TepeGhela, 5.8 тыс. Лет назад; ASHQ01, 2,3 тыс. Лет назад), два представителя из Европы (Ньюгрейндж, 4,8 тыс. Лет назад; HXH, 7,0 тыс. Лет назад) и один представитель из Арктики (Жохов, 9,5 тыс. Лет назад). Затем мы определили набор моделей, чтобы охарактеризовать происхождение каждой сибирской собаки как происходящее от одного из следующих: 1) исключительно арктического собачьего происхождения, 2) как арктического, так и европейского собачьего происхождения, 3) как арктического, так и ближневосточного происхождения, и 4) исключительно европейские и ближневосточные собаки (рис. 2). Во всех моделях европейское происхождение моделировалось как смесь Ближнего Востока и Арктики (8).Затем мы применили все четыре возможные комбинации репрезентативных геномов и проверили степень соответствия каждой сибирской / степной собаке.
Древние собаки из озера Байкал (∼7 тыс. Лет назад) обладают ограниченным западноевразийским происхождением.
Мы впервые применили этот метод, чтобы оценить, имели ли три собаки из озера Байкал (7,4, 7,0 и 6,9 тыс. Лет назад) какое-либо неарктическое происхождение. Наш анализ показывает, что три из наших четырех моделей одинаково хорошо соответствуют данным для двух самых старых с более низким охватом (Байкал2 и Байкал3; ∼0.3 ×) собаки (рис.2). Однако в случае самой молодой и более высокой выборки (Байкал1; 2,2 ×) озера Байкал мы обнаружили, что модели, включающие примесь из Европы, подошли немного лучше (рис. 2). Хотя соблазнительно интерпретировать эти результаты как свидетельство появления собак, имеющих неарктическую родословную между 7,4 и 6,9 тыс. Лет назад, это открытие также может быть результатом более низкого охвата двух более старых геномов. Чтобы дополнительно спросить, обладали ли эти собаки историей потока генов вообще, мы выполнили исчерпывающий поиск модели с помощью qpBrute (17), в котором все три собаки озера Байкал были проанализированы как единая популяция ( SI Приложение ).Результаты показывают, что только ~ 3% из более чем 20 000 протестированных сценариев соответствуют данным, и большинство из них (~ 60%) не связано с примесью из западно-евразийского источника ни в одной из собак озера Байкал с момента их разделения с Жоховым.
Хотя более сложные модели с примесью из западного источника, датирующего общего предка собак из озера Байкал и острова Жохов, соответствовали данным (также очевидно, если рассматривать только самую молодую собаку [6,9 тыс. Лет назад; рис. 2), тесты на соотношение F4 предполагали что этот вклад был незначительным (~ 9% предков собак озера Байкал).Кроме того, анализ D-статистики, основанный на сопоставлении с эталонным геномом рыжей лисы ( Vulpes vulpes ) (VulVul2.2), показал небольшое смещение эталона по сравнению со сборкой canFam3.1 (европейский боксер) ( SI Приложение ). , Рис. S10 и набор данных S4). Однако, независимо от эталонного генома, D-статистика для Baikal1 показала статистически значимые сигналы для потока генов от собак из Западной Евразии, но этот сигнал не был обнаружен у более старых байкальских собак (наборы данных S3 и S4).В совокупности наши результаты предполагают, что собаки озера Байкал потенциально получили ограниченный приток предков, около 6,9 тыс. Лет назад от собак Западной Евразии. Для более уверенной оценки этого сценария требуется большая глубина охвата этими выборками. Однако, если это правильно, это может означать, что распространение собак из западных и южных регионов в сибирскую Арктику началось между 9,5 тыс. Лет назад (возраст жоховской собаки) и 6,9 тыс. Лет назад.
Повышение уровня западноевразийского происхождения у сибирских собак за последние 2 тыс. Лет назад.
Затем мы применили наш метод, основанный на диаграмме примесей, для оценки примеси из западноевразийских источников в более поздних сибирских собаках из контекстов, датируемых периодом от 2,0 тыс. Лет назад до наших дней, после периода очевидного отсутствия прямо датированных останков собак в Сибири между ∼6,0 до 4,3 тыс. лет (24). В отличие от более ранних собак озера Байкал, все наиболее подходящие модели для этих собак включали примесь из западно-евразийского источника (рис. 2), что предполагает дополнительный приток европейских или ближневосточных предков в Сибирь после ∼6.9 тыс. Лет назад (самый молодой образец озера Байкал). Хотя в большинстве случаев лучше всего подходят модели с примесью из европейского источника, некоторые собаки железного века (∼2,0 тыс. Лет назад) и средневековья (1,1–0,85 тыс. Лет назад), полученные из Ямало-Ненецкого региона в Северо-Западной Сибири (рис. 1 A ) потенциально потенциально также имели дополнительное происхождение, родственное ближневосточным собакам (рис. 2 и SI, приложение , рис. S8). Сигнал о дополнительном вкладе Ближнего Востока в развитие средневековых ямальских собак был немного слабее при использовании выравнивания генома рыжей лисы вместо эталона собаки, но все еще присутствует (сравните SI Приложение , рис.S6 B и C ).
Далее мы исследовали этот сценарий с помощью исчерпывающего поиска модели с использованием qpBrute (17), в котором собаки железного века и средневековые ямальские собаки рассматривались как две популяции ( SI, приложение , рис. S11). Хотя количество и топология подгоночных моделей незначительно варьировались в разных прогонах ( SI Приложение , Таблица S3 и Рис. S11), все модели показали различную независимую примесь европейских и / или ближневосточных собак в средневековый и железный век Ямал. собаки.В соответствии с нелокальными материалами, включая стеклянные бусины и различные металлические предметы (13, 15), извлеченные из одних и тех же археологических раскопок, наши результаты показывают, что сообщества, жившие на Ямале в период железного века и средневековья, были связаны с более крупной торговой сетью, через которую они приобрели собак неарктического происхождения ( SI Приложение , рис. S8).
Возникновение современных линий сибирских собак.
Затем мы оценили, был ли геном двух собак, взятых около 100 лет назад (1927 г. н.э.) из ненецкой общины на полуострове Ямал, связан с ямальскими собаками периодов средневековья и железного века.Парные расстояния идентичности по происхождению показали, что ближайшим родственником этих двух исторических ненецких собак была древняя арктическая собака — средневековая ямальская собака из Усть-Войкара ( SI Приложение , рис. S4 B и D ), что свидетельствует о наличии степень преемственности населения от средневековья до начала 20 века. Интересно, что этот анализ показал, что эти две 100-летние ненецкие собаки были также тесно связаны с самоедскими собаками, современной породой шпицев, завезенной в Соединенное Королевство в девятнадцатом веке из Сибири и ставшей популярной среди полярных исследователей, таких как Скотт и Шеклтон. (25).Этот результат показывает, что, хотя было несколько эпизодов смешивания собак Северо-Западной Сибири, компонент их арктического происхождения сохранился в современной породе самоедов. Точно так же мы обнаружили, что современные сибирские хаски имеют сходство с историческими восточно-сибирскими собаками и древними собаками озера Байкал (25) (Рис. 1 и SI Приложение , Рис. S4 C и F и S7). В совокупности эти результаты показывают, что несколько популярных современных арктических пород сохранили значительный уровень предков от линии, установленной до 9.5 тыс. Лет в Сибири. Важно отметить, что несибирское происхождение сибирских пород собак — это не современное явление, а непрерывный процесс, по крайней мере, 2000 лет назад.
Выводы
Наши результаты показывают, что арктические собаки, вероятно, развивались практически изолированно от других популяций собак, по крайней мере, до середины голоцена (~ 7 тыс. Лет назад). Эти древние арктические собаки, вероятно, населяли большой регион Сибири от Новосибирских островов до озера Байкал. Однако за последние 7000 лет на эволюционную историю сибирских собак повлияли многократные интродукции собак из степей Евразии и Западной Евразии и незначительный или нулевой поток генов от восточноазиатских собак.Некоторые из этих внедрений совпадают с периодами крупных преобразований в обществах Северо-Западной Сибири, включая появление металлургии в Арктике (15), появление использования северных оленей для транспортировки ∼2 тыс. Лет назад и рост оленеводства ∼800 лет назад. (26). В целом это говорит о том, что эти глубокие преобразования в Северо-Западной Сибири были связаны с импортом материальной культуры (в том числе собак) из соседних регионов через создание крупномасштабных торговых сетей.Этот приток генетической изменчивости, связанной с родословной собак, адаптированной к земледелию (Европа) и пастбищу (степь), потенциально привел к поведенческим и морфологическим изменениям у арктических собак, которые, возможно, облегчили переход от кормодобывания к скотоводству в сибирской Арктике. Создание, анализ и интерпретация дополнительных геномов древних собак в их археологическом контексте поможет ответить на вопросы, связанные с ролью, которую собаки сыграли в долгосрочном заселении Арктики человеком, включая появление новых стратегий выживания в более общем плане. и, более конкретно, одомашнивание оленей и переход к крупномасштабному оленеводству.
Материалы и методы
Образцы, использованные для этого исследования, были получены из археологических контекстов ( n = 19) или этнографических коллекций, хранящихся в музеях ( n = 10). Подробные описания образцов, использованных для этого исследования, включая информацию об объектах и контекстную информацию, представлены в Приложении SI . ДНК была извлечена из каждого образца в лабораториях, посвященных работе с древними или деградировавшими материалами ДНК в Копенгагенском университете, Шведском музее естественной истории и Тринити-колледже в Дублине.Все реакции ПЦР проводились в отдельных помещениях для подготовки библиотек секвенирования; Скрининг для определения содержания эндогенной ДНК для каждого образца был проведен с помощью технологии секвенирования Illumina, соответственно 20 из 29 образцов были отобраны для глубокого секвенирования на платформах Illumina и BGISeq. Каждый образец был сопоставлен с эталонным геномом собаки CanFam3.1 (27) с использованием алгоритма обратного отслеживания выравнивания Барроуза-Уиллера (BWA aln) (28, 29), впоследствии на образцах был проведен псевдогаплоидный вызов и панель общедоступных образцов собак с ANGSD (30) для последующего анализа.Полная информация о методах, используемых для отбора проб, лабораторных работ и компьютерных анализов в этом исследовании, представлена в Приложении SI .
Благодарности
Мы благодарим Б. Грённова, Ф. Расимо, Б. Сакса и Э. Острандера за вклад и комментарии при концептуализации и ранних черновиках этого исследования. В этом исследовании использовались как Advanced Research Computing Оксфордского университета, так и центр высокопроизводительных вычислений Apocrita королевы Марии. Мы хотели бы поблагодарить Лабораторию науки для жизни, Шведскую национальную инфраструктуру геномики, за помощь в секвенировании ДНК.Признательны за предоставление дополнительного доступа к коллекциям и логической поддержки следующие учреждения: музей Института экологии растений и животных УрО РАН (Екатеринбург) и Центр Арктических исследований (Салехард). T.R.F. был поддержан рамочной программой ЕС по исследованиям и инновациям Horizon 2020 в рамках Соглашения о гранте 676154. T.R.F. также получил финансирование для анализа в рамках проекта Qimmeq, который поступил от фондов Velux и Aage og Johanne Louis-Hansens Fond.M.-H.S.S. был поддержан Фондом независимых исследований Дании (8028-00005B). L.A.F. и GL были поддержаны грантами Европейского исследовательского совета (ERC-2013-StG-337574-UNDEAD и ERC-2019-StG-853272-PALAEOFARM) и грантами Совета по исследованиям окружающей среды (NE / K005243 / 1, NE / K003259 / 1, NE / S007067 / 1 и NE / S00078X / 1). L.A.F. и A.C. были поддержаны Wellcome Trust (210119 / Z / 18 / Z). R.J.L. и Т. были поддержаны грантом Совета социальных и гуманитарных исследований IG 435-2014-0075.ОПЕРАЦИОННЫЕ СИСТЕМЫ. был поддержан программой Европейского Союза Horizon 2020 (h3020-MSCA-IF-2015, проект «EpiCDomestic», грант 704254). Сбор образцов проводился по Государственному контракту ИФАЭ УрО РАН (№ ААА А-А19-11
-0).
Сноски
Вклад авторов: T.R.F., M.T.P.G., G.L., L.D., A.J.H., M.-H.S.S. и L.A.F. спланированное исследование; T.R.F., A.C., R.J.L., T.N., K.G.D., M.-H.S.S. и L.F. проводили исследования; Д.Г.Б., К.Г.Д., С.Г., К.Г., О.С., Г.Z., M.T.P.G., G.L., L.D. и A.J.H. внесены новые реагенты / аналитические инструменты; R.J.L., T.N., A.A., I.A., O.A., E.A., M.A., O.P.B., F.B., A.V.G., C.J., P.A.K., Y.V.K., V.M., A.V.P., A.L.S., D.S. и L.V.Y. предоставленные образцы и информация о сайте; К.М.Г. предоставленные образцы и контекстная информация; T.R.F., A.C., S.G., J.R.-M., O.S., M.-H.S.S. и L.F. проанализировали данные; и T.R.F., A.C., R.J.L., T.N., A.A., I.A., O.A., E.E.A., M.A., O.P.B., F.B., K.M.G., A.V.G., C.J., P.K., Y.V.K., V.M., A.R.P., A.V.P., J.R.-M., A.L.S., D.S., L.V.Y., E.W., M.M., M.T.P.G., G.L., L.D., A.J.H., M.-H.S.S. и L.F. написали статью.
Авторы заявляют об отсутствии конкурирующей заинтересованности.
Эта статья представляет собой прямое представление PNAS.
Эта статья содержит вспомогательную информацию в Интернете по адресу https://www.pnas.org/lookup/suppl/doi:10.1073/pnas.2100338118/-/DCSupplemental.
- Авторские права © 2021 Автор (ы). Опубликовано PNAS.
Древние люди в высоких широтах Аркти
фото: Изделия из обсидиана, найденные на Жоховской стоянке посмотреть еще
Кредит: Владимир В.Питулько и др.
Российские ученые изучили Жоховскую стоянку древних людей, которая находится в высоких широтах Арктики, и подробно описали быт проживавших здесь древних людей. Оказалось, что, несмотря на малонаселенность местности, древние люди общались с представителями других территорий и даже обменивались с ними различными предметами через какие-то ярмарки.
Арктика — самая холодная часть нашей планеты, где нет благоприятных условий для жизни.Арктические районы Земли практически не заселены людьми, хотя в некоторых странах, в том числе в России, США и Норвегии, есть промышленные зоны. В этих зонах средняя плотность населения увеличивается до 0,35 человек / км2, но в основном она не превышает 0,03-0,04 человек / км2.
Остров Жохов, расположенный на 76º с.ш. на Новосибирских островах, в 440 км к северу от современного побережья Восточно-Сибирского моря, относится к Высоким Арктикам. Здесь было обнаружено поселение древнего человека, Жоховская стоянка, считающаяся самым ранним свидетельством проживания человека в Высокой Арктике.В поселении, существовавшем 9300-8600 лет назад, постоянно проживало 25-50 человек. В далеком прошлом остров Жохов был частью обширной равнины, образовавшейся в результате резкого падения уровня моря во время последнего ледникового максимума. Из-за послеледникового подъема уровня моря эта равнина была затоплена и размыта. В настоящее время Жоховский участок расположен в юго-западной части современного острова Жохов, у подножия невысокого, около 120 м, холма, прикрывающего его от сильного северо-западного ветра. Это место было очень удобным для древних людей.Холм служил смотровой площадкой, и из-за близости к берегу у них всегда был запас дров, которые привозили по морю.
Ученые провели раскопки в вечной мерзлоте и уже исследовали значительную часть памятника — 571 квадратный метр. В ходе работ было найдено множество инструментов из камня, костей, рогов, бивней и дерева. Это разные инструменты, предметы охотничьего оружия, детали саней и разнообразная домашняя утварь. Среди каменных артефактов много микропризматических лезвий, которые служили боковыми лезвиями для композитных инструментов, служивших наконечниками для копий / дротиков / стрел и ножей.Большинство из них были изготовлены из различных кремнистых пород местного происхождения, а некоторые из них были сделаны из необычного для этой части мира экзотического сырья, такого как обсидиан или вулканическое стекло. Исследователи нашли 79 таких предметов. Древние люди ценили этот материал за лучшую расщепляемость среди других камней и ценили его чрезвычайно тонкую и острую режущую кромку. Исследователи высоко ценят этот материал, потому что каждое месторождение обсидиана имеет свою уникальную геохимическую подпись, которая помогает найти источник материала.Тогда можно увидеть, как он движется в пространстве, и, таким образом, мы сможем больше узнать о жизни древних людей.
Однако поблизости от острова Жохов источника обсидиана нет. Ближайший участок с таким источником находится у озера Красное в низовьях реки Анадырь на Чукотке, что составляет около 1500 км по прямой, а фактическое перемещение может составить более 2000 км. Казалось бы, древние люди физически не могли преодолеть такое расстояние.
Обнаруженные предметы обсидиана изучались авторами методом рентгеновской флуоресценции.Это метод неразрушающего контроля, который позволяет определить геохимические свойства образцов и, таким образом, найти способ определить местонахождение источника материала с помощью уникальных сигнатур.
«Мы обнаружили, что именно этот вид обсидиана попадает на остров Жохов из района озера Красное. Это сверхдальнее путешествие, и трудно представить, что древние люди могли совершать такие путешествия около 9000 лет назад. Скорее всего, в промежуточных точках они встречались с другими людьми и обменивались изделиями из обсидиана, либо занимались примитивной торговлей », — сказал один из авторов статьи, Владимир Питулко, главный исследователь проекта RSF, к.Доктор исторических наук, старший научный сотрудник отдела палеолита Института истории материальной культуры Российского научного фонда.
Исследователи также проанализировали более 54 000 охотничьих остатков фауны и реконструировали годовой экономический цикл жителей Жоховской стоянки. Жители стоянки были классическими наземными охотниками, занимавшимися охотой на оленей. Однако зимой охотились на белых медведей, спящих в медвежьих берлогах. Для них это был надежный и стабильный способ добыть пропитание, потому что в этом районе было довольно много медвежьих нор.
Ранее ученые исследовали кости средних псовых; они указывают на наличие хорошо сложенной собаки, которая жила вместе с жителями Жоховской стоянки. Кроме того, эти животные по размеру и массе тела близки к современной породе ездовых собак. Находки частей саней в виде полозьев доказывают, что у жителей этих территорий была хорошо развитая транспортная система. Благодаря этой системе жители Жоховской стоянки путешествовали на близлежащие территории, которые в то время еще были частью материка, и поддерживали отношения со своими жителями.
Владимир Питулько также отметил, что участки в устьях рек Колымы и Индигирки могут служить промежуточными пунктами в этой торговле. При этом расстояние между пунктами обмена составляло около 700 км, что вполне реально для проезда ранней весной на собачьих упряжках.
Позднее такие «собрания» превратились в ярмарки, так как жители севера Восточной Сибири устраивали их в XVIII-XIX веках, но не только для обмена товарами. Гораздо более важным считался обмен различной информацией.В любом случае вещественные доказательства на острове Жохов представлены минимумом находок. Вероятно, важным результатом таких встреч стали брачные контакты, необходимые для обеспечения биологической устойчивости небольших групп людей, ведущих довольно замкнутый образ жизни.
«Результаты нашего исследования свидетельствуют о высоком уровне социокультурных связей, существовавших среди древнего населения Арктики Восточной Сибири 9000 лет назад, и о вовлеченности жителей Жоховской стоянки.Эти люди, жившие на краю земли, не были изолированы. Напротив, контакты внутри этих обществ были намного более развитыми, чем считалось раньше », — заключил Владимир Питулько.
###
Работа выполнена при сотрудничестве с исследователями Дальневосточного геологического института ДВО РАН, Института геологии и минералогии им. Соболева СО РАН и Университета штата Миссури, США. Исследование финансируется Российским научным фондом (РНФ), его результаты опубликованы в журнале Antiquity .
Заявление об ограничении ответственности: AAAS и EurekAlert! не несут ответственности за точность выпусков новостей, размещенных на EurekAlert! участвующими учреждениями или для использования любой информации через систему EurekAlert.
Materials Research Express, том 7, номер 1, январь 2020 г., январь 2020 г.
С помощью стандартной твердофазной реакции синтезировали серию соединений CaSiO 3 : x % Tb 3+ , y % Sm 3+ .Для определения кристаллической структуры были проведены исследования дифракции рентгеновских лучей, сканирующей электронной микроскопии и энергодисперсионной рентгеновской спектроскопии. Спектр возбуждения фотолюминесценции (ФЛ) легированных соединений Tb 3+ и Sm 3+ состоит из серии пиков, соответствующих электронным переходам ионов Tb 3+ и Sm 3+ . Длина волны 375 нм была оптимальной для возбуждения люминесценции со-легированных соединений Tb 3+ / Sm 3+ .Спектр излучения ФЛ состоит из набора интенсивных линий на длинах волн 415 нм, 436 нм, 457 нм, 488 нм, 543 нм, 586 нм и 622 нм, соответствующих электронным переходам Tb 3+ 5 D 3 → 7 F 5 , 5 D 3 → 7 F 4 , 5 D 3 → 7 F 3 , 5 D → 7 F 6 , 5 D 4 → 7 F 5 , 5 D 4 → 7 F 4 , и 5 D → 7 F 3 соответственно.Спектр излучения ФЛ легированного соединения Sm 3+ имеет серию из трех пиков, расположенных на длинах волн 565 нм, 602 нм и 648 нм, соответствующих электронным переходам Sm 3+ из 4 G . 5/2 от до 6 H 5/2 , 6 H 7/2 и 6 H 9/2 , соответственно. Предложена диаграмма уровней энергии. Измерения срока службы были проведены для всех образцов, и таким же образом был исследован процесс передачи энергии.Были определены координаты цветности Международной комиссии по освещению, коррелированная цветовая температура и индекс цветопередачи люминофоров, и результаты показывают, что полученные материалы являются оптимальным кандидатом для применения в белых светодиодах.
Европа PMC
PLoS One. 2015; 10 (5): e0125759.
, 1 , 2 , ¤ * , 1 , 3 , 4 , 4 , 5 и 3Эстер Дж.Ли
1 Департамент антропологии, Бингемтонский университет, Бингемтон, штат Нью-Йорк, Соединенные Штаты Америки,
2 Департамент социологии, антропологии и социальной работы, Техасский технический университет, Лаббок, Техас, Соединенные Штаты Америки,
Д. Эндрю Мерриуэзер
1 Кафедра антропологии, Бингемтонский университет, Бингемтон, штат Нью-Йорк, Соединенные Штаты Америки,
Алексей К. Каспаров
3 Институт истории материальной культуры РАН , Г.Санкт-Петербург, Россия,
Павел Александрович Никольский
4 Геологический институт РАН, Москва, Россия,
Сотникова Марина Владимировна
4 Геологический институт РАН, Москва, Россия,
Елена Юрьевна Павлова
5 Арктический и антарктический научно-исследовательский институт, Санкт-Петербург, Россия,
Питулько Владимир Васильевич
3 Институт истории материальной культуры РАН, г.Санкт-Петербург, Россия,
Дэвид Карамелли, научный редактор
1 Департамент антропологии, Бингемтонский университет, Бингемтон, штат Нью-Йорк, Соединенные Штаты Америки,
2 Департамент социологии, антропологии и социальной работы, Техасский технологический институт Университет, Лаббок, Техас, Соединенные Штаты Америки,
3 Институт истории материальной культуры Российской академии наук, Санкт-Петербург, Россия,
4 Геологический институт Российской академии наук, Москва, Россия,
5 Арктический и антарктический научно-исследовательский институт, г.Санкт-Петербург, Россия,
Университет Флоренции, ИТАЛИЯ,
Конкурирующие интересы: Компания Ford Motor приняла участие в исследовании территории маяка Ахим. Нет никаких патентов, продуктов в разработке или продаваемых продуктов, которые можно было бы декларировать. Это не влияет на соблюдение авторами всех политик PLOS ONE в отношении обмена данными и материалами.
Задумал и спроектировал эксперименты: ДАМ ВВП. Проведены эксперименты: EJL DAM. Проанализированы данные: EJL DAM.Предоставленные реагенты / материалы / инструменты для анализа: EJL DAM AKK PAN MVS EYP VVP. Написал статью: EJL DAM PAN MVS VVP.
¤ Текущий адрес: Департамент социологии, антропологии и социальной работы Техасского технологического университета, Лаббок, Техас, Соединенные Штаты Америки
Поступила в редакцию 19 ноября 2014 г .; Принято 26 марта 2015 г.
Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии надлежащего указания автора и источника.
Эта статья цитируется в других статьях в PMC.Сопутствующие данные
- Дополнительные материалы
- S1 Рис. Частичная нижняя челюсть образца S809 из Улахана Суллара. Образец был получен из слоя 2 в Улахан-Сулларе и описан как Canis cf. variabilis .
(TIF)
S2 Рис. Частичная нижняя челюсть образца S504 из Дуванного Яра. Морфологически идентифицированный как Canis lupus , экземпляр был получен в нижнем течении реки Колыма.(TIF)
S1 Таблица: Описание останков псовых из Дуванного Яра. Информация о двух образцах из Дуванного Яра, включая код поля, описание останков, местонахождение и детали радиоуглеродного датирования.(DOCX)
S2 Таблица: Описание особей псовых из Яны РХС. Информация о четырех образцах из Яны РХС, включая код поля, описание останков, местонахождение и детали радиоуглеродного датирования.(DOCX)
S3 Таблица: Описание особей псовых из Жохова.Информация о четырех образцах из Жохова, включая код поля, описание останков, местонахождение и детали радиоуглеродного датирования.(DOCX)
S4 Table: Описание псовых особей с маяка Ахим. Информация о двух образцах из Ахима, включая код поля, описание останков, местонахождение и детали радиоуглеродного датирования.(DOCX)
S5 Таблица: последовательностей праймеров митохондриальной ДНК, использованных в этом исследовании. Для исследования были разработаны три пары праймеров, и одна пара праймеров была использована из предыдущего исследования [64].(DOCX)
S6 Table: Образцы, использованные в медианной сети древних образцов псовых. Идентификационные коды соответствуют указанным номерам доступа GenBank и справочнику.(DOCX)
- Заявление о доступности данных
Все файлы последовательностей доступны в GenBank (номера доступа KJ1-KJ4).
Abstract
Современная Арктическая Сибирь предоставляет богатые ресурсы для археологических, геологических и палеонтологических исследований, позволяющих изучить динамику популяций фаунистических сообществ плейстоцена, особенно потому, что фаунистический материал, поступающий из вечной мерзлоты, оказался подходящим для генетических исследований.Чтобы изучить историю видов Canid в Сибирской Арктике, мы провели генетический анализ четырнадцати останков псовых из различных памятников, включая хорошо задокументированные памятники верхнего палеолита Яна и остров Жохов раннего голоцена. Предполагаемый возраст образцов варьируется от 1700 лет до настоящего времени (YBP) до 360 000 YBP для останков вымершего волка, Canis cf. variabilis . Чтобы изучить генетическое родство видов древних сибирских псовых с домашней собакой и современными волками, мы получили последовательности контрольных участков митохондриальной ДНК и сравнили их с опубликованными последовательностями древних и современных псовых.Более старые образцы собак демонстрируют сходство с додомашними линиями собак / волков, в то время как другие встречаются в основных филогенетических кладах домашних собак. Наши результаты предполагают, что европейское происхождение домашних собак не может быть окончательным, и иллюстрируют растущую сложность генетического вклада региональных пород волков в современный генофонд Canis .
Введение
Широко признано, что домашняя собака ( Canis lupus knownis ) произошла от серого волка ( Canis lupus ), но процесс приручения, а также географическое происхождение и приблизительная дата первого приручения все еще остаются обсуждается [1,2,3].Генетические исследования современных популяций собак и волков показали различные взгляды, от единственного происхождения в Восточной / Южной Азии [4,5] или на Ближнем Востоке [6] до нескольких областей одомашнивания и / или гибридизации с региональными породами волков [6, 7]. Более того, возможность смешивания с другими видами псовых также предполагалась ранее [8,9]. С другой стороны, недавний анализ митохондриального генома древних собак показал европейское происхождение домашних собак [10]. Археологические данные не всегда просты для морфологической идентификации домашних собак, тем более, что самые ранние собаки были практически того же размера, что и волки [11,12,13], но продвинутый морфометрический анализ улучшил эти усилия [14,15,16].Древнейшие археологические находки домашних собак были обнаружены в Западной Европе и на Ближнем Востоке и датируются по крайней мере 14 000 кал. До н. Э. [17,18]. Некоторые утверждали, что домашние собаки присутствовали до последнего ледникового максимума, но в настоящее время это оспаривается [13,19,20,21,22].
Археологические и палеонтологические исследования, проведенные в Арктической Сибири в течение последних нескольких десятилетий, дали большое количество костного материала, пригодного для генетических исследований, поскольку они в основном происходят из отложений вечной мерзлоты, которые распространены в этом районе.Многие исследования древней ДНК были сосредоточены на вымерших плейстоценовых или диких видах, населявших Сибирь [23,24,25,26], но здесь мы сосредоточимся на старейшем одомашненном виде Canis . Различные виды Canidae, такие как песец и волк, были среди плейстоценовой арктической фауны, сохранившейся до наших дней [27,28]. Исследования в этом регионе подтвердили присутствие собак на Русской равнине и Камчатке к 13000 г. до н.э. [29,30]. Недавнее исследование показало, что домашняя собака на юге Сибири датируется ок.33 300 кал. До н. Э., Что предшествует самым старым свидетельствам из Западной Европы и Ближнего Востока [22]. Однако останки сибирских псовых были морфологически наиболее похожи на собак из Гренландии и в отличие от древних и современных волков и предполагаемых собак из центральной России [22]. Саблин и Хлопачев [29] утверждали, что присутствие плейстоценовых собак на равнине в центральной части России в Елисеевичи I, датируемое 13 000–17 000 кал. До н.э., было результатом приручения in situ у местных северных волков. Таким образом, возможность того, что волки позднего плейстоцена / раннего голоцена могли внести свой вклад в региональную породу собак, остается открытой.
Мы исследовали тринадцать останков доисторических псовых и один образец современного волка из сибирской Арктики: Улахан-Суллар, Дуванный Яр, Яна РХС, остров Жохов и маяк Ахим (и). Самые старые образцы происходят из обнажений четвертичных отложений (местонахождение № 1, Canis cf. variabilis ) и из обнажения Дуванный Яр (местонахождение № 2). Подробная информация о каждом участке представлена в следующем разделе. Анализ митохондриальной ДНК (мтДНК) образцов псовых был проведен, чтобы сделать вывод о филогенетических отношениях этих древних псовых с современными видами псовых, с особым вниманием к Canis cf. variabilis экземпляр из Улахан-Суллара, который может дать ключ к разгадке происхождения домашних собак.
Расположение изучаемых участков.Соответствующие номера и информация приведены в.
Таблица 1
Описание образцов псовых, проанализированных в данном исследовании.
| ID | Местоположение | Морфологическая классификация | Возраст (YBP) а № | № на карте б |
|---|---|---|---|---|
| S809 | Улахан-Суллар, р. Адыча | Canis ср. вариабилис | 360 000–400 000 | 5 |
| S503 | Дуванъяр, Нижняя Колыма | Canis lupus | > 47000 | 4 |
| S504 | Дуванъяр, Нижняя Колыма | Canis lupus | > 47,000 | 4 |
| S501 | Яна РХС, Нижняя р. Яна | Canis lupus | 28,520 ± 240 | 2 |
| S601 | Яна справа, Нижняя р. Яна | Canis lupus | 27,840 ± 220 | 2 |
| S805 | Яна справа, Нижняя р. Яна | Canis lupus | на | 2 |
| S806 | Яна РХС, Нижняя р. Яна | Canis lupus | на | 2 |
| S502 | Аахим, Восточно-Сибирское побережье | Canis sp . | 1,740 ± 40 | 3 |
| S603 | Аахим, Восточно-Сибирское побережье | Canis sp . | 1,760 ± 40 | 3 |
| S602 | Остров Жохов, Новосибирские острова | Canis sp . | 8,710 ± 50 | 1 |
| S902 | Остров Жохов, Новосибирские острова | Canis sp . | на | 1 |
| S903 | Остров Жохов, Новосибирские острова | Canis sp . | на | 1 |
| S904 | Остров Жохов, Новосибирские острова | Canis sp . | на | 1 |
| S905 | Острова Новой Сибири | Canis lupus | Современный | 6 |
Материалы и методы
Информация об образце
Наш набор данных состоит из пяти групп окаменелостей, собранных в нескольких районах Арктической Сибири, которые охватывают территорию от острова Жохов до средней части реки Яна в широтном направлении. в направлении (от 76 ° 06 ‘северной широты до 67 ° 50’ северной широты, или на расстоянии почти 1000 км) и от низкой реки Яна до полуострова Ахим, Западная Чукотка по долготе (от 135 ° 25 ‘восточной долготы до 173 ° 30’ E, или расстояние 1500 км).Современный образец волка был взят в качестве эталона на Новосибирских островах [31].
Образцы, проанализированные в данном исследовании, были собраны в ходе полевых раскопок (1999–2007 гг.), Которые проводились на основании разрешений, выданных государственным органом Владимиру Васильевичу Питулько, старшему научному сотруднику Института истории материальной культуры РАН ( Руководитель проекта «Жохов-2000»): 1) 1999 г .: Мерительная грамота №123 (форма 2 — обследование), выданная Полевой комиссией 7 мая 1999 г. на разведку и раскопки на Западной Чукотке, Певекский район; 2) 2000 г .: Мерительная грамота №307 (форма 2 — обследование), выданная Полевой комиссией 29 мая 2000 г. на обследование на Новосибирских островах и раскопки на Жоховском острове; 3) 2001 г .: Мерительная грамота №381 (форма 1 — раскопки), выданная Полевой комиссией 8 июня 2001 г. на обследование на Новосибирских островах, раскопки на острове Жохов и обследование в Северной Яно-Индигирской низменности; 4) 2002 г .: Мерительная грамота №638 (форма 1 — раскопки), выданная Полевой комиссией 28 июня 2002 г .; 5) 2003 г .: Мерительная грамота №525 (форма 1 — раскопки), выданная Полевой комиссией 16 июня 2003 г., на раскопки на острове Жохов, обследование в Северо-Янинской Индигирской низменности и раскопки на стоянке Яна; 6) 2004 г .: Мерительная грамота №87 (форма 4 — раскопки), выданная Полевой комиссией 16 апреля 2004 г. на раскопки на острове Жохов и раскопки на стоянке Яна; 7) 2005 г .: Мерительная грамота №506 (форма 4 — раскопки-утильсы), выданная Полевой комиссией 10 июня 2005 г. на раскопки на острове Жохов и раскопки на стоянке Яна; 8) 2006 г .: Мерительная грамота №308 (форма 4 — раскопки), выданная Полевой комиссией 26 мая 2006 г. на раскопки на стоянке Яна; 9) 2007 г .: Мерительные грамоты №99 и 100 (форма 1, раскопки и форма 2, обследование), выданные Полевой комиссией 13 апреля 2007 года, раскопки на стоянке Яна и съемка в Северо-Янинской Индигирской низменности.Всю информацию можно проверить через Полевой комитет Института археологии РАН по адресу ur.xednay@naraiipo.
Местоположение и информация о местонахождении
Улахан-Суллар
Нижняя челюсть Canis ср. variabilis (S809) был получен из слоя 2, датированного 360 000 ± 17 000 лет назад, на основе измерений ЭПР раковин двустворчатых моллюсков, обнаруженных в том же слое [32]. Участок представляет собой обрыв высотой 65 м и длиной 1,2 км, расположенный на правом берегу невысокой реки Адыча (бассейн реки Яна) на 67 ° 41’N, 135 ° 44’E.Вид Canis cf. variabilis был охарактеризован Тейяром де Шарденом и Пей по экземплярам мелких среднеплейстоценовых волков в Азии [33]. Часто сгруппированные вместе с Canis mosbachensis в Европе обозначаются как C . mosbachensis-variabilis , вид демонстрирует морфологические особенности, включая относительно небольшой размер тела, удлиненную и тонкую морду и относительно длинный метастиль P4.
Дуванный Яр
Дуванный Яр — ключевой разрез верхнеплейстоценовых отложений Западной Берингии, расположенный на правом берегу нижнего течения реки Колымы в Северо-Восточной Якутии (68 ° 38’N; 159 ° 03’E) [34,35 ].На этом участке были охарактеризованы четыре стратиграфических единицы, особенно группа III, датированная периодом от 45 000 до 13 500 14 C лет до настоящего времени (YBP) и обнаружила многочисленные останки млекопитающих и останки растений [36,37,38,39]. Два останка псовых были обнаружены в самой нижней части блока III, для которых радиоуглеродные данные предполагают дату не менее 47 000 14 C YBP (таблица S1).
Яна RHS
Четыре экземпляра Canis lupus были получены с участка 28 500–27 000 14 C YBP [40,41,42].Расположенный в низовьях реки Яна, на стоянке РГО Яна обнаружены богатые слои культурных артефактов и остатков фауны, которые были подробно описаны в предыдущих исследованиях [41,42,43]. Три из четырех образцов имеют индивидуальную дату AMS 14 C (таблица S2), и все вместе они имеют твердую оценку возраста около 28 000 радиоуглеродных лет назад. При раскопках Яны РХС был обнаружен ряд возможных останков волка с сильно изношенными зубами.
Жохов
Участок Жохов относится к острову Жохов, расположенному ниже 76 ° северной широты, и относится к Новосибирской цепи островов, которая представляет собой естественную границу между морями Лаптевых и Восточно-Сибирским морем.Датируемый 7 800–8 000 YBP, это один из самых северных археологических памятников в Арктике, в котором хранятся многочисленные артефакты и остатки фауны, в том числе многочисленные микропризматические ядра и микролезвия, охотничье снаряжение и деревянные артефакты [44,45,46]. Останки псовых были найдены среди большого скопления артефактов, включающего останки фауны белого медведя и северного оленя, что составляет около 150 идентифицируемых костей как минимум девяти животных. В это исследование были включены четыре образца собак (S602, S902, S904, S903) (таблица S3).
Маяк Ахим
Участок расположен в северной части полуострова Ахим на побережье Восточно-Сибирского моря [47]. Две нижние челюсти псовых с этого участка (таблица S4) датированы 1760 ± 40 л.н. (Beta-231449) и 1740 ± 40 (Beta-231444), и, исходя из потенциального эффекта морского резервуара, приемлемый возраст будет около 1700 лет назад или немного моложе [46]. Несмотря на возраст, эти экземпляры появились еще до недавнего контакта, поэтому представляют собой местные породы собак. Помимо двух обнаруженных останков псовых, которые мы анализируем в этом исследовании, здесь есть многочисленные останки морских млекопитающих, а также каменные отщепы и артефакты [47].
Подготовка образцов
Все останки, за исключением образцов, собранных в Аахиме, оставались в условиях вечной мерзлоты вскоре после осаждения до раскопок. Образцы были отобраны, чтобы гарантировать наличие отдельных животных (черепа или фрагменты черепа), чтобы избежать дублирования выборки. Зубы получены из останков черепа и нижней челюсти. Остатки для справочного материала были получены от волка, умершего естественной смертью на острове Новая Сибирь в 2003 году.
Экспериментальные методы
Все экспериментальные процедуры анализа ДНК проводились в специализированном стерильном помещении для исследования древней ДНК, включая отдельные помещения, предназначенные для бурения, извлечения и подготовки образцов для ПЦР.Помещение оборудовано воздухом, отфильтрованным с помощью высокоэффективных твердых частиц (HEPA), имеет ультрафиолетовые лучи на всех поверхностях, а воздушный поток между комнатами ограничен магнитными блокировками на дверях и поддержанием постоянного положительного давления. Общие методы включают использование одноразовой одежды и перчаток, а также стерилизацию открытых поверхностей отбеливателем перед каждой процедурой.
Когти и зубы получены для выделения ДНК. Сначала образцы обеззараживали отбеливателем перед сверлением для получения мелкого костного порошка от 0 до 0.От 45 г до 5,45 г. Образцы просверливали и извлекали группами по пять человек плюс один отрицательный контроль. Подробные процедуры экстракции были описаны ранее [48]. Вкратце, сначала образцы декальцинировали в EDTA, затем инкубировали с протеиназой K, затем очищали с использованием модифицированного протокола спин-колонки на основе диоксида кремния с последующим центрифугированием.
ПЦР проводили для контрольной области митохондриальной ДНК с использованием трех перекрывающихся праймеров, охватывающих нуклеотидные положения (np) от 15424 до 15837 (413 пар оснований).Мы использовали два разных праймера для первого сегмента между np 15424 и 15580, как указано в таблице S5. Амплификация проводилась несколько раз для проверки подлинности. При наличии использовали несколько экстрактов, и согласованные последовательности из нескольких амплификаций были определены надежно. Успешно амплифицированные продукты ПЦР проверяли электрофорезом в агарозном геле, и ампликоны очищали на 96-луночном планшете Multiscreen HTS (Millipore, Billerica, MA) для удаления однонитевой ДНК и праймеров.Ампликоны были подготовлены для секвенирования с использованием BigDye Terminator v.3.1 (Applied Biosystems, Foster City, CA). После стандартного осаждения спиртом образцы напрямую секвенировали на ДНК-секвенаторе ABI PRISM 377XL (Applied Biosystems, Foster City, CA).
Последовательности были собраны для древних данных ДНК собак, а также для репрезентативного спектра образцов современных собак и волков из литературы и GenBank [4,10,49,50,51,52,53,54,55,56,57, 58,59,60,61]. Последовательности выравнивали с помощью ClustalX [62] и MEGA 5.2.2 [63], и мы включили наши данные в три отдельные медианные сети, которые включали: 1) последовательности древней ДНК псовых (N = 33), 2) последовательности современных собак (N = 290) и 3) последовательности современных волков ( N = 101). Сети с медианным соединением были рассчитаны в сети 4.6 [64].
Результаты и обсуждение
Мы успешно получили эндогенные последовательности контрольной области мтДНК для всех четырнадцати образцов собак и идентифицировали девять гаплотипов. У двух экземпляров с маяка Ахим был один гаплотип, а у четырех останков с острова Жохов — другой гаплотип.Из четырех экземпляров псовых Yana RHS два имели один и тот же гаплотип. Только гаплотипы Аахима и Жохова соответствовали другим опубликованным последовательностям псовых, тогда как последовательности из Яны RHS, Улахан-Суллара и Дуванного Яра были уникальными. Последовательности можно найти под номерами доступа GenBank KJ1-KJ4.
Прямой анализ целых митогеномов древних образцов псовых недавно позволил предположить европейское происхождение домашних собак на основании филогенетического расположения трех псовых из Бельгии, датированных 26 000–36 000 YBP [10], что в отличие от азиатских или ближневосточных. происхождение по современным образцам [5,6].Основываясь на последовательностях контрольной области мтДНК, образцы псовых из Яны РХС и Улахан-Суллар из нашего исследования выглядят столь же расходящимися, как и древние европейские образцы, которые использовались в качестве доказательства европейского происхождения домашних собак. Поэтому в свете наших результатов европейское происхождение домашних собак не может быть окончательным. Медианная сеть соединений для последовательностей древних псовых () показывает гаплотип Яны (S805), который находится в одном шаге от гаплотипа Жохова (S902), который представляет одну из основных филогенетических клад среди псовых (Clade A).Несколько древних гаплотипов псовых ориентированы вокруг Yana S805, в том числе некоторые из самых старых гаплотипов псовых, о которых сообщалось на сегодняшний день, в том числе образец Улахана-Суллара ( Canis см. variabilis , S809) из нашего исследования, в дополнение к древним гаплотипам псовых из Бельгия датировала 30 000 YBP и 36 000 YBP, а гаплотип псовых из Костенков, Россия, датировал 22 000 YBP [10]. Древние экземпляры псовых в этом кластере могут представлять возможных предков домашней собаки, поскольку время слияния расхождения собаки и волка, как полагают, произошло около 10 000 лет назад [4, 65], хотя некоторые предполагали, что это было уже 32 000 лет назад. [66].Основываясь на положении гаплотипа Yana S805, он потенциально может представлять прямую связь от предполагаемого предка (включая Canis cf. variabilis ) с линиями домашних собак и современных волков.
Медианная сеть образцов древних псовых для контрольной области митохондриальной ДНК.Черные кружки обозначают образцы из нашего исследования (см. Таблицу S6 для идентификации образца). Последовательности, проанализированные в сети, охватывают положения нуклеотидов 15561–15789.
В целом, большинство сибирских псовых из нашего исследования филогенетически сгруппированы с другими волками из Азии и России (). Одна ветвь от гаплотипа Яны (S805) состоит из современного гаплотипа волка. Интересно, что этот гаплотип взят из ныне вымершего японского волка ( Canis lupus hodophilax ), датируемого 14-18 веками [54]. Сообщается, что японский волк был довольно широко распространен на основных островах Японии с периода Дзёмон (10 000–250 гг. До н. Э.).C.) вплоть до начала 1900-х годов, но неясно, откуда они происходят [54,67]. Хотя широко распространено мнение, что серый волк ( Canis lupus ) является прямым предком домашней собаки, филогенетическая связь между серым волком и Canis cf. variabilis до сих пор остается предметом споров. Canis ср. variabilis , как полагают, был широко распространен в Евразии примерно до 300 000 YBP и, по-видимому, не пересекается с самым ранним появлением морфологически отличительного серого волка [68,69].Наше исследование является первым, в котором представлены результаты анализа ДНК Canis cf. variabilis , и наши результаты предполагают, что variabilis является еще одним возможным источником генетического вклада в развитие домашней собаки.
Медианная сеть образцов волков для контрольной области митохондриальной ДНК.Черными кружками отмечены образцы из нашего исследования. Последовательности, проанализированные в сети, охватывают позиции нуклеотидов 15547–15792. См. Образцы идентификаторов из этого исследования. Известные гаплотипы включают GQ509 (номер доступа в GenBank).GQ376509; Уральские горы в России [59]), JW237, 240, 255, 257 (регистрационный номер GenBank AB480736-AB480742, AB500700; Canis lupus hodophilax , Япония [54]) и LU51 (регистрационный номер GenBank AY812735; Нью-Мексико. , США [57]).
Одна интересная особенность филогенетической группы древних псовых, окружающих гаплотип Яна (S805), — это псовый Дуванный Яр (S504), который датируется более чем 47 000 14 C YBP, но отделен от других древних гаплотипов псовых и похоже, связан с домашними собаками, но не с волками.Как в сетях медианного соединения древних собак, так и домашних собак (рис. И), S504 связан с собачьими гаплотипами, которые находятся на пару мутаций от Clade A, но сеть волков показывает, что S504 генетически отличается от любых других гаплотипов (). Это может указывать на то, что линия собачьих пород Дуванни Яр имеет отличную филогению от волков, но, возможно, генетически унаследована от домашних собак. Хотя возможно, что линия Дуванни Яр просто не сыграла важную роль в одомашнивании современных линий собак, она может представлять собой локализованный генетический источник современных линий собак.
Медианная сеть образцов собак для контрольной области митохондриальной ДНК.Черными кружками отмечены образцы из нашего исследования. Последовательности, проанализированные в сети, охватывают позиции нуклеотидов 15547–15705. Clade A выделен светло-серым цветом, а образец S504 из Дуванного Яра отмечен в сети. Нижний правый кластер, выделенный пунктирной линией круга, включает образцы S805, S806, S809 и S501.
Филогенетическая сеть с последовательностями домашних собак показывает, что кластер, ориентированный вокруг гаплотипа Яны (S805), сохраняется из сети волков ().И гаплотипы Жохова, и Аахима, расположенные в кладе А, являются общими для нескольких домашних собак из широкого географического распространения [4,53,55]. Эта основная филогенетическая группа, известная как Clade A, наряду с Clades B, C и D, была охарактеризована на основании ранее проведенных исследований на собаках, которые включают большинство линий домашних собак [53,70]. Гаплотипы Жохова и Аахима генетически неотличимы от домашних собак, что может свидетельствовать о том, что домашние собаки обитали в Арктической Сибири не менее 8000 14 C YBP.С другой стороны, жоховские и аахимские псовые демонстрируют генетическое родство с азиатскими волками из. В частности, C . lupus chanco и C . lupus hodophilax либо имеют общие гаплотипы, либо отделены одной мутацией от сибирских псовых, включая одного янского псового. В то время как недавний полногеномный анализ местных собак Китая показал более близкое генетическое родство с домашними собаками [66], другое объяснение состоит в том, что сибирские псовые сохранили генетическую подпись от смешения с местными породами благодаря географической изоляции, что предполагалось в других древних породы собак [71].
Выводы
В целом, наши данные предполагают генетический вклад из региональных источников волков, включая, возможно, Canis cf. variabilis , в современную линию собак и подчеркивает важность дальнейшего изучения генетического вклада региональных пород волков в генофонд домашних собак. Кроме того, наши результаты согласуются с недавними исследованиями, предполагающими, что одомашнивание собак происходило в результате сложного процесса смешения различных пород, интегрированного с изоляцией региональных пород на протяжении всей истории.Будущие исследования могут дополнительно пролить свет на филогенетические отношения между различными видами собак.
Дополнительная информация
S1 Рис.
Частичная нижняя челюсть образца S809 из Улахана Суллара.Образец был получен из слоя 2 в Улахан-Сулларе и описан как Canis cf. variabilis .
(TIF)
S2 Рис.
Частичная нижняя челюсть образца S504 из Дуванного Яра.Морфологически идентифицированный как Canis lupus , экземпляр был получен в нижнем течении реки Колыма.
(TIF)
S1 Таблица
Описание останков псовых из Дуванного Яра.Информация о двух образцах из Дуванного Яра, включая код поля, описание останков, местонахождение и детали радиоуглеродного датирования.
(DOCX)
S2 Таблица
Описание образцов псовых из Яны РХС.Информация о четырех образцах из Яны РХС, включая код поля, описание останков, местонахождение и детали радиоуглеродного датирования.
(DOCX)
S3 Таблица
Описание особей псовых из Жохова.Информация о четырех образцах из Жохова, включая код поля, описание останков, местонахождение и детали радиоуглеродного датирования.
(DOCX)
S4 Таблица
Описание образцов псовых с маяка Ахим.Информация о двух образцах из Ахима, включая код поля, описание останков, местонахождение и детали радиоуглеродного датирования.
(DOCX)
S5 Таблица
Праймерные последовательности митохондриальной ДНК, использованные в этом исследовании.Для исследования были разработаны три пары праймеров, и одна пара праймеров была использована из предыдущего исследования [64].
(DOCX)
S6 Таблица
Образцы, использованные в медианной сети древних образцов псовых.Идентификационные коды соответствуют указанным номерам доступа GenBank и справочнику.
(DOCX)
Благодарности
Мы благодарим Федора Шидловского за его поддержку в расследовании дела в Дуванном Яре.Мы также благодарим Стейси МакГрат, Дженнифер Люэдтке Кеннеди, Елену Кунески Рапу, Лизу Мандарино и Джулию Фресессу за их помощь с молекулярными процедурами.
Отчет о финансировании
Эти экспериментальные процедуры и раскопки на островах Жохов, Яна РХС и Новосибирские острова были поддержаны Rock Foundation. Компания Ford Motor приняла участие в исследовании района маяка Ахим. Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Доступность данных
Все файлы последовательностей доступны в GenBank (номера доступа KJ1-KJ4).
Список литературы
1. Clutton-Brock J (1995) Происхождение собаки: приручение и ранняя история В: Serpell J, редактор. Домашняя собака: эволюция, поведение и взаимодействие с людьми. Кембридж: Издательство Кембриджского университета; С. 7–20. [Google Scholar] 2. Crockford SJ (2000) Комментарий к эволюции собак: региональные вариации, развитие породы и гибридизация с волками В: Crockford SJ, редактор.Собаки сквозь время: археологическая перспектива. Оксфорд: Археопресс; С. 295–312. [Google Scholar] 3. Линдблад-То К., Уэйд С.М., Миккельсен Т.С., Карлссон Е.К., Яффе Д.Б., Камаль М. и др. (2005) Последовательность генома, сравнительный анализ и структура гаплотипов домашней собаки. Природа 438: 803–819. [PubMed] [Google Scholar] 4. Панг Дж. Ф., Клютч С., Цзоу XJ, Чжан А. Б., Ло Л. Ю., Энглби Х. и др. (2009) данные мтДНК указывают на единственное происхождение собак к югу от реки Янцзы менее 16 300 лет назад от многочисленных волков.Молекулярная биология и эволюция 26: 2849–2864. 10.1093 / молбев / msp195 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 5. Динг З.Л., Оскарссон М., Ардалан А., Энглби Х., Дальгрен Л.Г., Тепели С. и др. (2012) Происхождение домашних собак в южной части Восточной Азии подтверждается анализом ДНК Y-хромосомы. Наследственность 108: 507–514. 10.1038 / hdy.2011.114 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 6. vonHoldt BM, Pollinger JP, Lohmueller KE, Han E, Parker HG, Quignon P, et al. (2010) Полногеномный анализ SNP и гаплотипов показывает богатую историю, лежащую в основе одомашнивания собак.Природа 464: 898–902. 10.1038 / природа08837 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 8. Адамс Дж. Р., Леонард Дж. А., Уэйтс Л. П. (2003) Широкое распространение гаплотипа митохондриальной ДНК домашней собаки у койотов юго-востока США. Мол Экол 12: 541–546. [PubMed] [Google Scholar] 9. Уэйн Р.К., Рой М.С., Гиттлман Дж. Л. (1998) Происхождение красного волка: ответ Новаку, Федероффу и Гарденеру. Биология сохранения 12: 726–729. [Google Scholar] 10. Thalmann O, Shapiro B, Cui P, Schuenemann VJ, Sawyer SK, Greenfield DL, et al.(2013) Полные митохондриальные геномы древних собак предполагают европейское происхождение домашних собак. Наука 342: 871–874. 10.1126 / science.1243650 [PubMed] [CrossRef] [Google Scholar] 11. Musil R (1984) Первое известное приручение волков в Центральной Европе В: Grigson C, Clutton-Brock J, редакторы. Животные и археология. Оксфорд: Международная серия британских археологических отчетов; 227 с. 23–25. [Google Scholar]12. Nobis G (1986) Die Wildaugetiere in der Umwelt des Menschen von Oberkassel bei Bonn und das Domestikationsproblem von Wolfen in Jungpaläolithikum.Боннер Ярбухер 186.
13. Жермонпре М., Саблин М.В., Стивенс Р.Э., Хеджес РЭМ, Хофрайтер М., Стиллер М. и др. (2009) Ископаемые собаки и волки из палеолитических стоянок в Бельгии, Украине и России: остеометрия, древняя ДНК и стабильные изотопы. Журнал археологической науки 36: 473–490. [Google Scholar] 14. Макдональд Д.В., Силлеро-Зубири С., редакторы (2004) Биология и сохранение диких собак. Оксфорд: Издательство Оксфордского университета. [Google Scholar] 15. Пионье-Капитан М., Бемилли С., Боду П., Селерье Дж., Ферри Дж. Г., Фоссе П. и др.(2011) Новые свидетельства существования мелких домашних собак верхнего палеолита в Юго-Западной Европе. Журнал археологической науки 38: 2123–2140. [Google Scholar] 16. Drake AG, Coquerelle M, Colombeau G (2015) Трехмерный морфометрический анализ ископаемых черепов собак противоречит предполагаемому одомашниванию собак во время позднего палеолита. Научный представитель 5: 8299 10.1038 / srep08299 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 17. Davis SJM, Valla FR (1978) Свидетельства одомашнивания собаки 12000 лет назад у натуфийцев Израиля.Природа 276: 608–610. [Google Scholar] 18. Napierala H, Uerpmann H-P (2012) «Новая» палеолитическая собака из Центральной Европы. Международный журнал остеоархеологии 22: 127–137. [Google Scholar] 19. Крокфорд С.Дж., Кузьмин Ю.В. (2012) Комментарии к записи Жермонпре и др., Journal of Archaeological Science 36, 2009 г. «Ископаемые собаки и волки из палеолитических памятников в Бельгии, Украине и России: остеометрия, древняя ДНК и стабильные изотопы» и Жермонпре, Лазкичкова-Галетова и Саблин, Журнал археологических наук 39, 2012 «Палеолитические собачьи черепа на могильнике Пршедмости, Чешская Республика».Журнал археологической науки 39: 2797–2801. [Google Scholar] 20. Жермонпре М., Саблин М.В., Деспрес В., Хофрейтер М., Лазничкова-Галетова М., Стивенс Р.Э. и др. (2013) Палеолитические собаки и раннее приручение волка: ответ на комментарии Крокфорда и Кузьмина (2012). Журнал археологической науки 40: 786–792. [Google Scholar] 21. Crockford SJ (2006) Ритмы жизни: гормон щитовидной железы и происхождение видов. Виктория, Британская Колумбия: Траффорд; 272 с. [Google Scholar] 22. Оводов Н.Д., Крокфорд С.Дж., Кузьмин Ю.В., Хайэм Т.Ф., Ходгинс Г.В., ван дер Плихт Дж. (2011) Зарождающаяся собака возрастом 33000 лет из Горного Алтая в Сибири: свидетельство самого раннего приручения, нарушенного Последним ледниковым максимумом.PLoS ONE 6: e22821 10.1371 / journal.pone.0022821 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 23. Шапиро Б., Драммонд А.Дж., Рамбаут А., Уилсон М.К., Матеус П.Е., Шер А.В. и др. (2004) Взлет и падение берингийского степного зубра. Наука 306: 1561–1565. [PubMed] [Google Scholar] 24. Барнс И., Шапиро Б., Листер А., Кузнецова Т., Шер А., Гатри Д. и др. (2007) Генетическая структура и исчезновение шерстистого мамонта, Mammuthus primigenius. Текущая биология 17: 1072–1075. [PubMed] [Google Scholar] 25.Барнетт Р., Шапиро Б., Барнс И., Хо С.Ю., Бургер Дж., Ямагути Н. и др. (2009) Филогеография львов (Panthera leo ssp.) Выявляет три различных таксона и сокращение генетического разнообразия в позднем плейстоцене. Мол Экол 18: 1668–1677. 10.1111 / j.1365-294X.2009.04134.x [PubMed] [CrossRef] [Google Scholar] 26. Кампос П.Ф., Виллерслев Э., Шер А., Орландо Л., Аксельссон Э., Тихонов А. и др. (2010) Анализ древней ДНК исключает людей как движущую силу позднеплейстоценовой динамики популяции овцебыков (Ovibos moschatus).Proc Natl Acad Sci U S A 107: 5675–5680. 10.1073 / pnas.09107 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 27. Куртен Б., Андерсон Э. (1980) Плейстоценовые млекопитающие Северной Америки. Нью-Йорк: издательство Колумбийского университета. [Google Scholar] 28. Кармайкл Л. Е., Кризан Дж., Надь Дж. А., Фуглей Е., Дюмон М., Джонсон Д. и др. (2007) Исторические и экологические детерминанты генетической структуры арктических псовых. Мол Экол 16: 3466–3483. [PubMed] [Google Scholar] 29. Саблин М.В., Хлопачев Г.А. (2002) Собаки раннего ледникового периода: свидетельства из Елисеевичей 1.Современная антропология 43: 795–799. [Google Scholar] 30. Диков Н.Н. (1996) Ушкинские стоянки, полуостров Камчатка В: Вест ФХ, ред. Американские истоки Предыстория и палеоэкология Берингии. Чикаго: Издательство Чикагского университета; С. 244–250. [Google Scholar] 31. Питулько В.В., Анисимов М.А., Басилян А.Е., Гирия Е.Ю., Никольский П.А., Одесский Д.П. и др. (2003) Делая новый шаг: проект Жохов 2000, Экспедиция 2001 года Материалы практического семинара Арктической системной научной программы 2002 г. Фэрбенкс, Аляска: Консорциум арктических исследований США; стр.189. [Google Scholar] 32. Никольский П.А. (2010) Систематика и стратиграфическое значение позднекайнозойских лосей / лосей (Alcini, Cervidae, Mammalia) из Евразии и Северной Америки. [Кандидатская диссертация]. Москва: Геологический институт РАН; 258 с. [Google Scholar] 33. Тейяр де Шарден П., Пей В.К. (1941) Ископаемые млекопитающие местонахождения 13 в Шукутьене. Palaeontologia Sinica 11: 1–105. [Google Scholar] 34. Каплина Т.Н., Гитерман Р.Е., Лахтина О.В., Абрашов Б.А., Киселев С.В., Шер А.В. (1978) Утес Дуванный — ключевой разрез позднеплейстоценовых отложений Колымской низменности.Бюллетень четвертичной комиссии 48: 49–65. [Google Scholar] 35. Шер А.В., Каплина Т.Н., Гитерман Р.Е., Ложкин А.В., Архангелов А.А., Киселев С.В. и др. (1979) Поздний кайнозой Колымской низменности. XIV Тихоокеанский научный конгресс, Экскурсовод XI, Хабаровск, август 1979 г. Москва, Академия наук СССР: XIV Тихоокеанский научный конгресс; С. 1–116. [Google Scholar] 36. Гитерман Р. Э., Шер А. В., Мэтьюз Дж. В. Младший (1982) Сравнение развития тундро-степных сред на западе и востоке Берингии: данные о пыльце и макрофоссилиях из ключевых разрезов.В: Hopkins DM, Matthews JV, Schweger CE, Young SB, редакторы. Палеоэкология Берингии: Academic Press; С. 43–73. [Google Scholar] 37. Занина О.Г., Губин С.В., Кузьмина С.А., Максимович С.В., Лопатина Д.А. (2011) Палеообстановки позднего плейстоцена (MIS 3–2) по данным отложений, палеопочв и гнезд сусликов в Дуванном Яре, Колымская низменность, северо-восток Сибири. Четвертичные научные обзоры 30: 2107–2123. [Google Scholar] 38. Васильчук Ю.К., Васильчук А.С., Ранг D, Кучера В., Ким Дж.С. (2001) Радиоуглеродное датирование участков δ 18 O-δD в позднеплейстоценовых клиньях льда Дуванного Яра (нижняя часть реки Колыма, северная Якутия) .Радиоуглерод 43: 541–553. [Google Scholar] 39. Яшина С., Губин С., Максимович С., Яшина А., Гахова Е., Гиличинский Д. (2012) Регенерация целых плодородных растений из плодовых тканей возрастом 30 000 лет, погребенных в сибирской вечной мерзлоте. Proc Natl Acad Sci U S A 109: 4008–4013. 10.1073 / pnas.1118386109 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 40. Питулько В.В., Никольский П.А., Гиря Е.Ю., Басилян А.Е., Тумской В.Е., Кулаков С.А. и др. (2004) Яна РИТ: люди в Арктике до последнего ледникового максимума.Наука 303: 52–56. [PubMed] [Google Scholar] 41. Питулько В.В., Павлова Е.Ю. (2010) Геоархеология и радиоуглеродная хронология каменного века Северо-Восточной Азии. Санкт-Петербург: Наука; 263 с. [Google Scholar] 42. Питулько В.В., Павлова Е.Ю., Кузьмина С.А., Никольский П.А., Басилян А.Е., Тумской В.Е. и др. (2007) Природно-климатические изменения Яно-Индигирской низменности в конце каргининского времени и местообитания позднеплейстоценового человека в северной части Восточной Сибири. Доклады наук о Земле 417: 1256–1260.[Google Scholar] 43. Питулько В.В. (2010) Позднеплейстоценовое обитание и адаптации древнего человека в Северо-Восточной Азии // Под ред. Деревянко А.Б., Куделин А.Б., Тишков В.А., ред. Адаптация народов и культур к естественным изменениям, социальным и техногенным преобразованиям. Москва: Российская академия наук; С. 38–46. [Google Scholar] 44. Питулько В.В., Каспаров А.К. (1996) Охотники древней Арктики: материальная культура и стратегия выживания. Арктическая антропология 33: 1–36. [Google Scholar] 45. Питулько В.В. (2003) Охотники на медведя острова Жохов, Восточная Российская Арктика В: Хабу Дж, Савелле С., Кояма С., Хонго Х, редакторы.Охотники-собиратели северной части Тихого океана. Осака: Национальный музей этнологии; С. 141–152. [Google Scholar] 46. Питулько В.В. (2006) Заметки по радиоуглеродной хронологии голоценового каменного века Северо-Восточной Азии // Лебединцев А.И., ред. Неолит и палеометалл Северного Дальнего Востока. Магадан: НИСНИИ ДВО РАН; С. 50–66. [Google Scholar] 47. Питулько В.В. (2001) Морские адаптации в Северо-Восточной Азии // Лебединцев А.И., Гоголева Т.Ю., ред. Магадан: Северо-Восточный научный центр; С. 64–70.[Google Scholar] 48. Ли Э.Дж., Андерсон Л.М., Дейл V, Мерриуэзер Д.А. (2009) Происхождение MtDNA порабощенной рабочей силы из могильника Шайлер Флэттс 18-го века в колониальном Олбани, штат Нью-Йорк: африканцы, коренные американцы и малагасийцы? Журнал археологической науки 36: 2805–2810. [Google Scholar] 49. Лоузи Р.Дж., Гарви-Лок С., Леонард Дж. А., Катценберг М.А., Жермонпре М., Номоконова Т. и др. (2013) Похоронение собак в древнем Предбайкалье, Сибирь: временные тенденции и взаимосвязь с рационом питания человека и практикой существования.PLoS ONE 8: e63740 10.1371 / journal.pone.0063740 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 50. Дружкова А.С., Тальманн О., Трифонов В.А., Леонард Я.А., Воробьева Н.В., Оводов Н.Д. и др. (2013) Анализ древней ДНК подтверждает, что алтайский псовый является примитивной собакой. PLoS ONE 8: e57754 10.1371 / journal.pone.0057754 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 51. Леонард Дж. А., Уэйн Р. К., Уиллер Дж., Валадез Р., Гиллен С., Вила С. (2002) Древние доказательства ДНК происхождения собак Нового Света из Старого Света.Наука 298: 1613–1616. [PubMed] [Google Scholar] 52. Лоузи Р.Дж., Базалийский В.И., Гарви-Лок С., Жермонпре М., Леонард Дж. А., Аллен А. Л. и др. (2011) Псовые как личности: ранненеолитические захоронения собак и волков, Прибайкалье, Сибирь. Журнал антропологической археологии 30: 174–189. [Google Scholar] 53. Вила С., Саволайнен П., Мальдонадо Дж. Э., Аморим И. Р., Райс Дж. Э., Ханикатт Р. Л. и др. (1997) Множественное и древнее происхождение домашней собаки. Наука 276: 1687–1689. [PubMed] [Google Scholar] 54. Исигуро Н., Иношима Ю., Шигехара Н. (2009) Анализ митохондриальной ДНК японского волка (Canis lupus hodophilax Temminck, 1839) и сравнение с репрезентативными гаплотипами волка и домашней собаки.Zoolog Sci 26: 765–770. 10.2108 / zsj.26.765 [PubMed] [CrossRef] [Google Scholar] 55. Tsuda K, Kikkawa Y, Yonekawa H, Tanabe Y (1997) Широкое скрещивание произошло между множественными матриархальными предками во время приручения собак: свидетельства меж- и внутривидового полиморфизма в области D-петли митохондриальной ДНК между собаками и волками. Гены Genet Syst 72: 229–238. [PubMed] [Google Scholar] 56. Исигуро Н., Иношима Ю., Шигехара Н., Итикава Х., Като М. (2010) Остеологический и генетический анализ вымершего волка Эзо (Canis lupus hattai) с острова Хоккайдо, Япония.Zoolog Sci 27: 320–324. 10.2108 / zsj.27.320 [PubMed] [CrossRef] [Google Scholar] 57. Леонард Дж. А., Вила С., Уэйн Р. К. (2005) Утраченное наследие: генетическая изменчивость и размер популяции искорененных серых волков США (Canis lupus). Мол Экол 14: 9–17. [PubMed] [Google Scholar] 58. Weckworth BV, Dawson NG, Talbot SL, Flamme MJ, Cook JA (2011) Переход к побережью: общая эволюционная история между прибрежными волками Британской Колумбии и юго-восточной Аляской волками (Canis lupus). PLoS ONE 6: e19582 10.1371 / journal.pone.0019582 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 59.Weckworth BV, Talbot SL, Cook JA (2010) Филогеография волков (Canis lupus) на северо-западе Тихого океана. J Млекопитающее 91: 363–375. [Google Scholar] 60. Вергинелли Ф., Капелли С., Койа В., Мусиани М., Фалчетти М., Оттини Л. и др. (2005) Митохондриальная ДНК доисторических собак подчеркивает отношения между собаками и волками Юго-Восточной Европы. Молекулярная биология и эволюция 22: 2541–2551. [PubMed] [Google Scholar] 61. Klutsch CF, Seppala EH, Fall T, Uhlen M, Hedhammar A, Lohi H, et al. (2010) Региональная встречаемость, высокая частота, но низкое разнообразие митохондриальной ДНК гаплогруппы d1 предполагает недавнюю гибридизацию собаки и волка в Скандинавии.Anim Genet. [Бесплатная статья PMC] [PubMed] [Google Scholar] 62. Jeanmougin F, Thompson JD, Gouy M, Higgins DG, Gibson TJ (1998) Множественное выравнивание последовательностей с Clustal X. Trends Biochem Sci 23: 403–405. [PubMed] [Google Scholar] 63. Тамура К., Петерсон Д., Петерсон Н., Стечер Г., Ней М., Кумар С. (2011) MEGA5: анализ молекулярной эволюционной генетики с использованием методов максимального правдоподобия, эволюционного расстояния и максимальной экономии. Молекулярная биология и эволюция 28: 2731–2739. 10.1093 / molbev / msr121 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] 64.Bandelt HJ, Forster P, Rohl A (1999) Медианные сети для определения внутривидовой филогении. Мол Биол Эвол 16: 37–48. [PubMed] [Google Scholar] 65. Скоглунд П., Готерстром А., Якобссон М. (2011) Оценка времени расхождения популяций по неперекрывающимся геномным последовательностям: примеры для собак и волков. Молекулярная биология и эволюция 28: 1505–1517. 10.1093 / molbev / msq342 [PubMed] [CrossRef] [Google Scholar] 66. Ван Г.Д., Чжай В., Ян Х.С., Фан RX, Цао Х, Чжун Л. и др. (2013) Геномика отбора у собак и параллельная эволюция между собаками и людьми.Nat Commun 4: 1860 10.1038 / ncomms2814 [PubMed] [CrossRef] [Google Scholar] 67. Мияо Т., Нисидзава Т., Судзуки С. (1980) Остатки млекопитающих самого раннего периода Дзёмон в Каменном убежище Точибара, префектура Нагано I. Фауна останков млекопитающих. Журнал Маммологического общества Японии 8: 181–188. [Google Scholar] 68. Tedford RH, Wang X, Taylor BE (2009) Филогенетическая систематика североамериканских ископаемых Caninae (Carnivora: Canidae) Бюллетень AMNH № 325. Нью-Йорк, Нью-Йорк: Американский музей естественной истории — научные публикации; стр.218. [Google Scholar] 69. Сотникова М., Ладья Л. (2010) Распространение Canini (Mammalia, Canidae: Caninae) по Евразии в период от позднего миоцена до раннего плейстоцена. Четвертичный международный 212: 86–97. [Google Scholar] 70. Savolainen P, Zhang YP, Luo J, Lundeberg J, Leitner T (2002) Генетические доказательства восточноазиатского происхождения домашних собак. Наука 298: 1610–1613. [PubMed] [Google Scholar] 71. Ларсон Г., Карлссон Е.К., Перри А., Вебстер М.Т., Хо С.Ю., Петерс Дж. И др. (2012) Переосмысление приручения собак путем интеграции генетики, археологии и биогеографии.Proc Natl Acad Sci U S A 109: 8878–8883. 10.1073 / пнас.1203005109 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] ТранскрипцияEctopic MicroRNA-150-5p повышает чувствительность глюкокортикоидной терапии в клетках множественной миеломы MM1S, но не может преодолеть резистентность к гормональной терапии в клетках MM1R
Abstract
Глюкокортикоиды (ГК) избирательно запускают гибель клеток в линии клеток множественной миеломы MM1S, которые экспрессируют белок NR3C1 / глюкокортикоидного рецептора (GR), но не способны убить клетки MM1R, в которых отсутствует белок GR.Учитывая недавние демонстрации измененных профилей микроРНК при различных гематологических злокачественных новообразованиях и устойчивости к лекарственным средствам, мы охарактеризовали индуцируемые GC профили транскрипции мРНК и микроРНК в GC-чувствительных MM1S по сравнению с GC-устойчивыми клетками MM1R. Анализ транскриптома показал, что GC регулируют экспрессию множества генов, участвующих в контроле клеточного цикла, организации клеток, гибели клеток и иммунологическом заболевании в клетках MM1S, которые остаются неизменными в клетках MM1R. Что касается микроРНК, mir-150-5p был идентифицирован как наиболее стойкая микроРНК, регулируемая GC, из 5 подтвержденных QPCR микроРНК (mir-26b, mir-125a-5p, mir-146-5p, mir-150-5p, и mir-184), которые индуцируются GC в MM1S, но не в клетках MM1R.Функциональные исследования также показали, что эктопическая трансфекция синтетического mir-150-5p имитирует GR-зависимые изменения экспрессии генов, участвующие в путях гибели и пролиферации клеток. Примечательно, что, несмотря на наблюдаемые изменения экспрессии генов, сверхэкспрессия mir-150-5p в отсутствие GC не вызывала значительной цитотоксичности в клетках MM1S или MM1R. Это предполагает необходимость дополнительных шагов в индуцированной GC гибели клеток, которую нельзя имитировать только сверхэкспрессией mir-150-5p. Интересно, что комбинация трансфекции mir-150-5p с низкими дозами GC в клетках MM1S, как было обнаружено, сенсибилизирует ответ на терапию, тогда как противоположные эффекты могут наблюдаться со специфическим антагомиром mir-150-5p.Хотя сверхэкспрессия mir-150-5p существенно не меняет уровни экспрессии GR, было обнаружено, что mir-150-5p вызывает специфические эффекты GR посредством непрямой регуляции мРНК взаимодействующих с GR факторов транскрипции и рецепторов гормонов, шаперонов GR, а также различных эффекторов развернутый белковый стресс и передача сигналов хемокинов. В целом, GC-индуцибельный mir-150-5p добавляет еще один уровень регуляции к GC-специфическим терапевтическим ответам при множественной миеломе.
Образец цитирования: Palagani A, Op de Beeck K, Naulaerts S, Diddens J, Sekhar Chirumamilla C, Van Camp G, et al.(2014) Эктопическая транскрипция MicroRNA-150-5p сенсибилизирует ответ на глюкокортикоидную терапию в клетках множественной миеломы MM1S, но не может преодолеть резистентность к гормональной терапии в клетках MM1R. PLoS ONE 9 (12): e113842. https://doi.org/10.1371/journal.pone.0113842
Редактор: Ян П. Такерманн, Ульмский университет, Германия
Поступила: 19 марта 2014 г .; Принята к печати: 1 ноября 2014 г .; Опубликовано: 4 декабря 2014 г.
Авторские права: © 2014 Palagani et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Авторы подтверждают, что все данные, лежащие в основе выводов, полностью доступны без ограничений. Все соответствующие данные находятся в документе и его файлах с вспомогательной информацией. Необработанные данные массива экспрессии генов были загружены в базу данных Gene Expression Omnibus (GEO) и имеют номер доступа GSE59805.
Финансирование: Исследования были поддержаны грантом стратегических фундаментальных исследований (SBO) Агентства по инновациям в науке и технологиях (IWT, Бельгия), Фонда исследований множественной миеломы (MMRF, США), FWO и NOI / DOCPRO (UA) исследовательские гранты. Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили, что никаких конкурирующих интересов не существует.
Введение
Множественная миелома (ММ) — это В-клеточное новообразование, характеризующееся накоплением клональных злокачественных плазматических клеток в костном мозге и часто коррелирующее с различными цитогенетическими аномалиями, такими как del (13), t (11-14), non -гипердиплоидия и del (17p) [1], [2]. Заболевание является причиной 10% злокачественных гематологических заболеваний и примерно 1% смертей, связанных с раком, в западных странах [3]. Терапия против множественной миеломы состоит из лекарств, которые могут уменьшить популяцию клональных плазматических клеток.Первоначальное лечение болезни зависит в основном от возраста пациента и сопутствующих заболеваний. Способность глюкокортикоидов (ГК) эффективно убивать лимфоидные клетки привела к их включению практически во все протоколы химиотерапии лимфоидных злокачественных новообразований. Для пациентов в возрасте до 65 лет высокие дозы химиотерапии с различными комбинациями, такими как схемы на основе талидомида-дексаметазона-бортезомиба и леналидомид-дексаметазон, с последующей трансплантацией аутологичных гемопоэтических стволовых клеток стали практикой в клинике в последние годы [4]. [5], [6], [7], [8].Несмотря на прогресс в терапии, ММ остается неизлечимой из-за низких показателей ремиссии традиционных методов лечения, что приводит к короткой продолжительности жизни (3–4 года) и развитию лекарственной устойчивости. В настоящее время проходят испытания многочисленные новые комбинации лекарственных препаратов для предотвращения резистентности и повышения эффективности ГК в терапии лимфоидных злокачественных новообразований [9].
Глюкокортикоиды (ГК) — это стероидные гормоны, которые проявляют свое про- или антиапоптотическое действие через рецептор глюкокортикоидов (GR, NR3C1) и изменяют экспрессию генов [10], [11].Изоформа GRα, которая является основным медиатором эффектов GC, включает три основных функциональных домена: N-концевой домен (NTD), ДНК-связывающий домен (DBD) и лиганд-связывающий домен (LBD) [12]. Неактивная форма GRα в основном находится в цитоплазме, образуя шаперонный комплекс со своими партнерами по связыванию, такими как белки теплового шока, FKBP51, FKBP52, CYP40 и другие белки [13]. После связывания с GC GR активируется и перемещается в ядро. Активированный GR имеет несколько различных способов действия на клеточном уровне.Наиболее заметной из них является трансактивация, при которой активированный GR связывается с элементом ответа на глюкокортикоиды (GRE) и индуцирует экспрессию генов. Кроме того, GR может связываться с негативными сайтами GRE (nGRE) или связанными факторами транскрипции (NFκB, AP1, P53 или STAT) и, следовательно, (транс) репрессирует экспрессию генов [14]. Хотя он известен как сильный индуктор апоптоза лимфоидных клеток в течение почти столетия, сигнальные пути, регулирующие восприимчивость или устойчивость раковых клеток к ГК, выявлены лишь частично.Помимо медленных геномных действий GC, зависящих от регуляции ядерных генов, также быстрые негеномные эффекты GC в цитоплазме модулируют апоптотический ответ [15], [16]. Что касается развития резистентности к терапии GC, были предложены различные механизмы, включая деградацию GR, Akt-зависимое фосфорилирование GR, снижение активности гистондеацетилазы-2 (HDAC2), чрезмерную активацию фактора транскрипции NFkB или AP1, повышенный фактор ингибирования миграции макрофагов, окислительный стресс и опосредованная Р-гликопротеином регуляция оттока лекарств [17], [18], [19], [20].Новая возникающая гипотеза состоит в том, что микроРНК модулируют ответ на терапию GC посредством (посттранскрипционной) регуляции множества мишеней мРНК, участвующих в передаче сигналов GR, связанных с пролиферацией и гибелью клеток [21], [22], [23], [24]. MiRNA представляют собой короткие некодирующие РНК длиной приблизительно 22 нуклеотида, которые регулируют экспрессию от десятков до сотен генов посредством деградации мРНК или репрессии трансляции на основе комплементарности последовательности к затравочной области miRNA (положения 2-8) в 3’UTR мРНК [25].Альтернативно miRNAs могут косвенно репрограммировать транскрипционные ответы, изменяя уровни мРНК уровней транскрипционных факторов [26]. Таким образом, микроРНК могут представлять собой возможный механизм обратной связи для точной настройки величины индуцированной GC гибели клеток посредством прямой регуляции уровней мРНК / белка GR или косвенной регуляции партнеров по взаимодействию GR или последующих мРНК-мишеней, отвечающих за GC / GR. Поскольку нарушение регуляции miRNA оказывает глубокое влияние на дифференцировку, развитие, апоптоз и прогрессирование клеточного цикла, профили их экспрессии могут использоваться в качестве биомаркеров для определения типа и степени злокачественности (прогноза) и / или индивидуального ответа на терапию [27].Чтобы охарактеризовать регуляторную роль GC-индуцируемых микроРНК в ответе на терапию множественной миеломы, мы сравнили профили транскрипции мРНК и микроРНК в GC-чувствительных клетках MM1S и GC-устойчивых клетках MM1R при воздействии GC. Последняя клеточная модель часто используется для выяснения регуляторных механизмов, участвующих в резистентности к терапии ГК в клетках ММ [28].
Материалы и методы
Культура клеток и жизнеспособность клеток
GC-чувствительные линии клеток MM1S (CRL-2974) и GC-устойчивые MM1R (CRL-2975) были описаны ранее [28] и были приобретены в ATCC.Вкратце, клетки MM1S представляют собой линию клеток миеломы В-лимфобластов, полученную от 42-летней темнокожей женщины. Клеточная линия MM1R представляет собой устойчивый к стероидам субклон, полученный из клеток MM1S, хронически подвергающихся действию дексаметазона (Dex). Клетки культивировали в среде RPMI-1640, содержащей 10% фетальной бычьей сыворотки (одобрено ЕС; происхождение из Южной Америки) и 0,5% раствора пенициллина / стрептомицина (Invitrogen). К клеточным линиям дополнительно добавляли 1% незаменимых аминокислот MEM и 1% пирувата натрия (Invitrogen).Каждую клеточную линию поддерживали при 37 ° C в атмосфере 5% CO 2 и 95% воздуха и влажности 95–98% до слияния. Жизнеспособность клеток оценивали с помощью колориметрического анализа с 3- (4,5-диметилтиозол-2-ил) -2,5-дифенилтетразолийбромидом (МТТ) (Sigma Aldrich, Сент-Луис, Миссури, Соединенные Штаты Америки), как описано ранее. [29].
Антитела и реагенты
Водорастворимый дексаметазон был приобретен в (Sigma Aldrich, Сент-Луис, Миссури, Соединенные Штаты Америки), и 10 мМ исходные растворы были приготовлены в стерильной воде.Антитело против GRα (sc-8992), антитело против PARP (sc-7150) были приобретены в Santa Cruz Biotechnology (Даллас, Соединенные Штаты Америки), а антитела против α-тубулина (T5168) были приобретены у Sigma Aldrich (St Луис, Миссури, США).
Экстракция РНК и обработка микрочипов
Суммарная РНК из клеток MM1S и MM1R, обработанных Dex (1 мкМ) или трансфицированных фиктивными миметиками или mir-150-5p, из трех независимых экспериментов была выделена с использованием набора RNeasy Mini Kit (Qiagen, Venlo, Нидерланды) в соответствии с протоколом производителя. .После экстракции и измерения концентрации (NanoDrop 1000, Thermo Scientific, Уолтем, Массачусетс, США) качество общей РНК контролировали на приборе Bio-Rad Experion (Bio-Rad, Hercules, Калифорния, США). 500 нг тотальной РНК амплифицировали с использованием набора Illumina Total Prep RNA Amplification (Life Technologies, Карлсбад, Калифорния, США). Вкратце, РНК подвергали обратной транскрипции с использованием праймеров олиго (dT) Т7, после чего биотинилированную кРНК синтезировали посредством реакции транскрипции in vitro. 750 нг амплифицированной кРНК гибридизовали с соответствующим массивом бус-чипа HumanHT12 (Illumina, Сан-Диего, Калифорния, США).Были выполнены умножения. Гранулированный чип инкубировали в течение 18 часов при 58 ° C в гибридизационной печи при непрерывном качании. После нескольких последовательных этапов промывки (см. Протокол производителя) интенсивность шариков считывали на Illumina Iscan.
Анализ данных микрочипов
Все сгенерированные значения интенсивности необработанных данных были считаны с использованием пакета «beadarray» (v2.8.1) в R [30]. Интенсивность была нормирована на квантиль, и дифференциальная экспрессия генов между образцами оценивалась с помощью «Limma» (v3.14.1). Полученные p-значения были скорректированы для проверки множественных гипотез с использованием метода Бенджамини-Хохберга. После оценки экспрессии генов были рассчитаны евклидовы расстояния между образцами и генами, которые использовались в качестве метрики расстояния в иерархическом кластерном анализе с МэВ [31]. Анализ пути был выполнен в базе знаний Ingenuity Pathway (Ingenuity Systems, Редвуд-Сити, Калифорния, США) в соответствии с предоставленными инструкциями. Отсечка кратности изменения 2, а также коэффициент ложного обнаружения 0.1% были установлены для идентификации генов, экспрессия которых значительно дифференцировалась. Необработанные данные массива были загружены в базу данных Gene Expression Omnibus (GEO) и имеют номер доступа GSE59805.
Конверсия кДНК и анализ QPCR
Вкратце, 1 мкг РНК превращали в кДНК с использованием системы обратной транскрипции Go Script в соответствии с инструкциями производителя (Promega, Madison, WI, Соединенные Штаты Америки). Sybr green был приобретен в (Bioline, Лондон, Великобритания). QPCR выполняли с использованием системы ABI7300 (Applied biosystems, Life Technologies Corporation).
анализ miRNA
ВыделениеmiRNA выполняли с помощью набора miRNeasy (Qiagen, Venlo, Нидерланды) в соответствии с инструкциями производителя. Профили экспрессии 760 микроРНК стволовой петли RT-qPCR miRNA проводили, как описано ранее [32], [33]. Необработанные значения экспрессии были нормализованы с использованием стратегии нормализации глобального среднего [32]. Подтверждение результатов массива QPCR было выполнено путем обратной транскрипции тотальной РНК, содержащей miRNA, и обнаружение miRNA с помощью ПЦР в реальном времени с использованием соответственно miScript II RT Kit и miScript SYBR Green PCR Kit (Qiagen, Венло, Нидерланды).Наборы специфичных для микроРНК праймеров QPCR против человека mir-146a (MS00003535), mir-491-5p (MS00031899), mir-146b-5p (MS00003542), mir-26b-5p (MS00003234), mir-150-5p (MS00003577), mir-330-3p (MS00031738), mir-195 (MS00003703), mir-184 (MS00003640), mir-125a-5p (MS00003423) были приобретены у Qiagen (Венло, Нидерланды). Для прогнозирования мишеней и путей микроРНК использовалось программное обеспечение miRWalk [34]. Корреляцию профилей транскрипции мРНК и микроРНК клеток MM1S и MM1R, необработанных или обработанных Dex, или после имитации трансфекции или трансфекции миметиком mir-150-5p, проводили с помощью программного обеспечения IPA.
Трансфекция клеток ММ in vitro синтетическим миметиком miR-150-5p и антагомиром
Миметик Мир-150-5р и антагомир были приобретены в компании Qiagen (Венло, Нидерланды). Всего 5 × 10 6 клеток подвергали электропорации с имитацией контроля или синтетическим mir-150-5p при конечной концентрации 150 нмоль / л с помощью Nucleofector Kit V, с использованием системы трансфекции AMAXA, программа O23 из (LONZA, Базель, Швейцария), как описано в другом месте [35].
Изоляция белка
5 × 10 6 клеток высевали в чашки диаметром 10 см и готовили лизаты белков для различных экспериментальных установок для вестерн-анализа с буфером для образцов 1XSDS, содержащим 62.5 мМ трис-HCL pH-6,8, 2% SDS, 10% глицерин, 0,5% бромфеноловый синий, DTT (5 мМ).
Статистический анализ
Статистические данные были выполнены с использованием однофакторного дисперсионного анализа, двухфакторного дисперсионного анализа с последующим пост-тестом Даннета и Бонферрони и односторонним парным t-критерием Стьюдента с использованием программного обеспечения с графической подушечкой prism5.
Результаты
Дексаметазон снижает жизнеспособность клеток в зависимости от дозы и времени в GC-чувствительных MM1S, но не в GC-устойчивых MM1R клетках
Перед исследованием регуляторной роли GC-индуцибельной микроРНК в экспрессии NR3C1 / GR-зависимых генов и ответах на гибель клеток мы сначала охарактеризовали ответ на терапию GC в клетках MM1S и MM1R.Вкратце, клетки MM1S и MM1R оставляли необработанными или обрабатывали 1 мкМ синтетического агониста GC дексаметазона (Dex) в течение соответственно 6 часов, 24 часов, 48 часов, 72 часов. мРНК и белки выделяли в каждый момент времени и анализировали на соответствующую экспрессию NR3C1 / GRα. Обработка клеток различными концентрациями Dex в течение 24 ч, 48 ч и 72 ч с последующим измерением жизнеспособности клеток с помощью колориметрического анализа MTT выявила зависимое от дозы и времени снижение выживаемости клеток MM1S после воздействия GC (рис.1A и 1B). Напротив, Dex не обладает способностью вызывать цитотоксичность в клетках MM1R и скорее вызывает слабый эффект стимуляции роста [28], [35], [36], [37]. В соответствии с различным ответом GC в обеих клеточных линиях, анализ QPCR демонстрирует значительную транскрипцию мРНК NR3C1 / GRα в MM1S, но близкие к фоновым уровням мРНК NR3C1 / GRα в клетках MM1R (рис. 1C). Вестерн-анализ, наконец, подтверждает значительные уровни экспрессии белка GRα в MM1S и неопределяемые уровни белка GRα в клетках MM1R (рис.1D – E). Как видно из рис. 1D, клетки MM1S экспрессируют множественные трансляционные изоформы GRα [38], [39], экспрессия которых снижается после обработки GC.
Рис. 1. Дексаметазон снижает жизнеспособность клеток в зависимости от дозы и времени в GC-чувствительных MM1S, но не в GC-устойчивых MM1R клетках.
( A – B ) кривые доза-ответ клеток MM1S и MM1R, обработанных дексаметазоном в различных концентрациях в течение 24, 48 и 72 часов, как определено колориметрическим анализом МТТ.Данные представляют собой (среднее ± SEM) значения трех независимых экспериментов. ( C ) Зависимые от времени изменения уровней мРНК GRα после обработки 1 мкМ дексаметазоном. Данные представляют собой (среднее ± SEM) значения трех независимых экспериментов, нормализованные для гена домашнего хозяйства 28sРНК и относительно необработанного состояния MM1S (S UT). Средние с ***, **, * значительно различаются (p <0,001, <0,01 или <0,05 и ns существенно не различаются, как определено односторонним дисперсионным анализом ANOVA (пост-тест Даннета) ( D ) Изменения, зависящие от времени в уровнях белка GRα и расщеплении PARP в клетках MM1S и MM1R после обработки 1 мкМ Dex.( E ) Денситометрический анализ сигналов вестерн-блоттинга (n = 2) уровней белка GR (92 кДа), β-актина (40 кДа), полноразмерного PARP (116 кДа) и расщепленного PARP (89 кДа) в MM1S и MM1R после обработки 1 мкМ Dex. Показаны существенные различия между западными сигналами в MM1R и MM1S.
https://doi.org/10.1371/journal.pone.0113842.g001
Несмотря на зависящее от времени снижение уровней мРНК и белка GR после 24–72 ч обработки GC (рис. 1C – 1D), реакция гибели клеток в MM1S не отменяется.Более того, вызванная GC гибель клеток запускает зависимое от времени расщепление апоптотического маркера PARP в MM1S, но не в клетках MM1R (Fig. 1D-E). В отличие от уровней GR, наблюдаемых в MM1S, уровни экспрессии большинства изоформ GR в MM1R ниже предела обнаружения, хотя фоновые уровни варианта 70 кДа все еще могут быть обнаружены при длительном воздействии (данные не показаны). Примечательно, что хотя Dex полностью лишен способности вызывать цитотоксичность в клетках MM1R, слабый эффект стимуляции роста может наблюдаться при длительном воздействии GC на клетки MM1R.Это может быть связано с остаточной активностью возможной антиапоптотической трансляционной изоформы GR [38].
Дексаметазон вызывает специфические для GC транскрипционные изменения в клеточном цикле, повреждение / репарацию ДНК, гибель клеток и иммунологические пути в MM1S, но не в MM1R
Для оценки изменений экспрессии специфических генов, связанных с ответом на терапию GC при множественной миеломе, мРНК была выделена в трех независимых экспериментах из клеток MM1S и MM1R, подвергнутых воздействию GC Dex (1 мкМ) в течение 72 часов.После выделения мРНК профили транскрипции по всему геному определяли гибридизацией массива Illumina. Результаты были проанализированы с помощью R-пакета «Limma» (v3.14.1) [40], [41], что привело к идентификации 268 гена с пониженной регуляцией и 151 гена с повышенной регуляцией в MM1S после фильтрации для минимального двукратного изменения транскрипции в Образцы, обработанные Dex, по сравнению с контрольной установкой (абсолютное кратное изменение> 2) и скорректированное значение P <0,05 (база данных Gene Expression Omnibus (GEO) GSE59805). В MM1R не удалось обнаружить никаких значительных изменений, связанных с Dex, в соответствии с фенотипом устойчивости к GC и отсутствием GRα в клетках MM1R (см.рис.1). Тепловая карта (рис. 2A) представляет собой графическое представление специфических для Dex транскрипционных изменений в MM1S и отсутствия ответа Dex в клетках MM1R в трех биологических повторах, демонстрируя воспроизводимые и устойчивые паттерны экспрессии генов. С помощью анализа QPCR мы подтвердили GC-зависимую трансактивацию целевого гена GILZ (TSC22D3), а также GC-специфическую (транс) репрессию CDK2 в MM1S, но не в MM1R, в соответствии с данными массива Illumina и предыдущими отчетами [37], [42] ], [43], [44] (рис.2Б). Примечательно, что длительное воздействие GC (72 часа) на клетки MM1R выявляет слабо повышенные уровни CDK2 согласно анализу QPCR, что не может быть обнаружено с помощью менее чувствительного анализа микроматрицы. Этот эффект может быть связан с остаточной активностью антиапоптотических изоформ GR в MM1R, что также может способствовать слабому стимулирующему эффекту роста (рис. 1А) при длительном воздействии [38]. Комплексный анализ путей (IPA) также показал, что GC запускают изменения экспрессии генов, которые способствуют гибели клеток, уменьшают пролиферацию клеток (клеточный цикл, репликацию ДНК) и регулируют иммунологические гематологические пути в MM1S, но не в MM1R (Таблица 1), в соответствии с уменьшенной клеткой. анализ жизнеспособности MM1S (см. рис.1). Затем мы дополнительно подтвердили результаты микроматрицы некоторых дополнительных генов-мишеней, участвующих в клеточном цикле, повреждении / репарации ДНК и гибели клеток. Как видно из (рис. 2C и 2D), результаты проверки QPCR коррелируют с полученными данными микрочипа. Кроме того, анализируя паттерны корегулируемых генов, программное обеспечение IPA позволяет идентифицировать активацию или ингибирование возможных вышестоящих регуляторов (киназ, рецепторов, факторов транскрипции), участвующих в изменениях экспрессии генов [45]. Последний анализ предсказывает весьма значительную активацию факторов транскрипции NR3C1 и TP53, связанных с Dex-лиганом, в регуляции остановки клеточного цикла MM1S и / или апоптоза [46] (Таблица 2).Более того, предсказанная активация IKK и циклин-зависимых киназ CDKN2A, CDKN1A может быть индикаторами повреждения ДНК / гибели клеток и событий клеточного цикла, запускаемых Dex [47], [48], [49], [50]. Наконец, возможное ингибирование антиапоптотических сигналов просуществования, стимулированных киназами TNF и AKT, также может вносить вклад в индуцированную Dex гибель клеток в MM1S (таблица 2).
Рис. 2. Дексаметазон запускает специфические для GC транскрипционные изменения в клеточном цикле, повреждение / репарацию ДНК, гибель клеток и иммунологические пути в MM1S, но не в MM1R.
( A ) Тепловая карта представляет относительные уровни экспрессии генов в клетках MM1S и MM1R, оставленных без обработки (растворитель) или обработанных 1 мкМ Dex в течение 72 часов. Экспрессия гена изменяется более чем в 2 раза, а значение p <0,05 отображается на тепловой карте. ( B ) Левая панель: Количественная ПЦР в реальном времени (QPCR) проверка Dex-зависимой активации транскрипции мРНК GILZ (TSC22D3) Правая панель: Количественная ПЦР в реальном времени (QPCR) проверка Dex-зависимой репрессии мРНК CDK2 в MM1S, но нет в MM1R.Средние значения с ***, **, * значительно отличаются (p <0,001, <0,01 или <0,05 соответственно) по сравнению с контрольными установками, определенными односторонним методом (пост-тест Даннета) ( C – D ) в реальном времени. количественная проверка ПЦР (QPCR) и наблюдение на микроматрице Illumina верхних регулируемых GC генов включали клеточный цикл, повреждение / репарацию ДНК и гибель клеток в S = MM1S, но не в R = MM1R. Все валидационные исследования QPCR были нормализованы для гена домашнего хозяйства РНК 28S и представлены относительно необработанного состояния MM1S (S UT).Гистограммы представляют относительные уровни мРНК (среднее ± стандартная ошибка среднего) трех независимых экспериментов. Средние значения с ***, **, * значительно отличаются (p <0,001, <0,01 или <0,05 соответственно) от контрольных установок, как определено с помощью двухфакторного дисперсионного анализа (пост-тесты Бонферрони).
https://doi.org/10.1371/journal.pone.0113842.g002
Предполагается, что микроРНК, реагирующие на GC в клетках MM1S, но не отвечающие на GC в клетках MM1R, модулируют клеточный цикл, пути роста и пролиферации клеток
Тотальную РНК выделяли из клеток MM1S и MM1R, обработанных 1 мкМ Dex в течение 72 часов, профилирование 760 микроРНК методом qPCR, как описано ранее [32].Это позволило идентифицировать 30 GC индуцибельных и 14 GC репрессированных miRNA (более чем в 2 раза) в MM1S, которые не изменяют транскрипцию в клетках MM1R (суммировано в Таблице 3). Затем был применен анализ IPA для перекрестного сравнения профилей микроРНК и мРНК в MM1S для прогнозирования генов-мишеней и основных путей, на которые влияет экспрессия микроРНК. Анализ диаграммы Венна 5 основных молекулярных и клеточных путей, обогащенных генами, дифференциально экспрессирующимися в MM1S при воздействии Dex, и прогнозируемые гены-мишени микроРНК, реагирующие на лечение Dex, показывает, что чувствительные к GC микроРНК могут нацеливаться на мРНК, участвующие в клеточном цикле и путях пролиферации клеток (рис.3А). Например, индуцируемые Dex микроРНК mir-150-5p, mir-152, mir-146b-5p, mir-1290, mir-125a-5p, mir-1206, как предполагается, модулируют несколько генов-мишеней, участвующих в клеточном цикле, клеточном росте и пути распространения (отмечены желтым цветом в таблице S1).
Рис. 3. Экспериментальная проверка GC-индуцируемых микроРНК, потенциально участвующих в регуляции пути NR3C1 (GR) в клетках MM1S
( A ) Диаграмма Венна, которая суммирует перекрытие между 5 основными предсказанными молекулярными и клеточными функциями, генерируемыми IPA из GC-регулируемых miRNA и GC-регулируемых мРНК ( B ) Venn-диаграмма, которая представляет собой перекрестное сравнение списка GC-чувствительных miRNA, идентифицированных в MM1S, со списком miRNA, специфичных для пути GR (NR3C1), в соответствии с бесплатной программой прогнозирования MiRWalk.( C ) Проверка уровней транскрипции 9 общих miRNA, выбранных на фиг. 3B, в клетках MM1S и MM1R, обработанных в течение 24, 48 или 72 ч 1 мкМ Dex, в режиме реального времени. ( D ) ГХ дозозависимая регуляция уровней mir-150-5p в клетках MM1S, обработанных ГХ в течение 72 часов. Все валидационные исследования miRNA QPCR были нормализованы для микроРНК SNORD 95 и SNORD 96A в соответствии с инструкциями производителя и относительно необработанного состояния MM1S (S UT). Гистограммы представляют относительные уровни miRNA (среднее ± стандартная ошибка среднего) трех независимых экспериментов по сравнению с контрольными установками.Средние значения с ***, **, * значительно отличаются от контрольных установок (p <0,001, <0,01 или <0,05), как определено односторонним дисперсионным анализом (тест Dunnetts Post).
https://doi.org/10.1371/journal.pone.0113842.g003
Чтобы уменьшить количество Dex-чувствительных микроРНК для дальнейшей проверки и функциональных исследований, мы затем сравнили список GC-чувствительных miRNA, идентифицированных в MM1S, со списком микроРНК, предположительно регулирующих путь GR (NR3C1) (Biocarta) в соответствии с базой данных MiRWalk [34] (рис.3Б). Это сгенерировало короткий список из 11 miRNA, которые были отобраны для дальнейшей проверки QPCR. В том же духе корреляция IPA репрессированных мРНК Dex и 30 индуцибельных miРНК Dex выявила аналогичную специфическую роль GR для mir-150-5p, mir-146b-5p и mir-125a-5p в клеточном цикле и путях клеточного роста и пролиферации. (Таблица S1). Зависимые от времени GC-ответные изменения уровней транскрипции 11 выбранных микроРНК оценивали с помощью QPCR в клетках MM1S и MM1R, подвергнутых в разные моменты времени (6 ч, 24 ч, 48 ч и 72 ч) 1 мкМ Dex.Анализ QPCR может подтвердить время и специфическую индукцию GC 5 hsa-микроРНК, т.е. mir-26b, mir-125a-5p, mir-146-5p, mir-150-5p и mir-184 (рис. 3C). Наконец, hsa-mir-150-5p был выбран для дальнейших функциональных исследований из-за его стойкой GC-чувствительной транскрипции между 6 и 72 часами и зависимой от дозы GC регуляции mir-150-5p после 72-часовой обработки (рис. 3D). При перекрестном сравнении 1203 Dex-зависимых генов-мишеней из клеток MM1S с обработкой 1 мкМ Dex в течение 72 часов (выбор гена производился на основе значения p <0.05 и абсолютное кратное изменение> 1,5) и 3626 предсказали цели hsa-mir-150-5p через базу данных miRWalk (предсказание по крайней мере 4 из 10 доступных алгоритмов, то есть DIANAmT, miRanda, miRDB, miRWalk, RNAhybrid, PICTAR4, PICTAR5, PITA, RNA22 и Targetscan), 196 общих мРНК / генных мишеней (рис. 4A) были идентифицированы в MM1S, которые остаются незатронутыми в клетках MM1R (подробный список генов можно найти в таблице S2). При использовании анализа IPA, 196 мРНК предсказывали GC-чувствительные гены-мишени mir-150-5p, обогащенные по гибели клеток, морфологии клеток, клеточному циклу, путям клеточного роста и пролиферации (рис.4А). Особо следует отметить, что восходящий анализ IPA предсказывает, что гены-мишени гена mir-150-5p чувствительны к регуляции с помощью передачи сигналов GC (Dex) / NR3C1, PPAR или STAT / IFNγ (фиг. 4B).
Рисунок 4. Прогнозируемая роль mir-150-5p в регуляции гибели и выживания клеток, клеточного цикла, клеточного роста и пролиферации в клетках MM1S.
( A ) Диаграмма Венна, которая представляет собой перекрестное сравнение 3626 предсказанных целей hsa-mir-150-5p в соответствии с программным обеспечением miRWalk и 1203 чувствительных к дексаметазону генов-мишеней в клетках MM1S, которые остаются неизменными в клетках MM1R ( B ) IPA предварительный анализ предсказанных мРНК-мишеней микроРНК в клетках MM1S.
https://doi.org/10.1371/journal.pone.0113842.g004
Трансфекция миметика mir-150-5p запускает экспрессию специфического гена GR в MM1S, но не в клетках MM1R, без модуляции уровней белка GR
Чтобы проверить возможные эффекты mir-150-5p на предсказанные мишени мРНК и / или клеточный цикл или пути клеточной пролиферации, мы провели эксперименты по трансфекции в клетках MM1S и MM1R с синтетическим миметиком mir-150-5p по сравнению с лечением Dex. Примечательно, что хотя MM1S и MM1R демонстрируют сходные уровни транскрипции mir-150-5p при миметической трансфекции (рис.5A), мы могли наблюдать значительные изменения в мРНК, отвечающих за mir-150-5p, в MM1S, но не в клетках MM1R, после анализа профилей мРНК Illumina. Интересно, что при перекрестном сравнении обработанных Dex и трансфицированных миметиком mir-150-5p клеток MM1S / MM1R анализ IPA выявил заметное перекрытие в затронутых мРНК, связанных с пролиферацией клеточного цикла, репарацией ДНК и путями гибели клеток в MM1S, а также отсутствие эффекты для обеих установок в клетках MM1R (рис. 5B) (таблица 4 и 5). Чтобы оценить, может ли mir-150-5p изменять уровни мРНК GR / NR3C1 или стабильность белка, мы выполнили QPCR и вестерн-блот-анализ после трансфекции mir-150-5p в клетках MM1S.Из фиг. 5C и D можно наблюдать, что трансфекция mir-150-5p не изменяет уровни мРНК или белка GR / NR3C1 соответственно. Эти результаты согласуются с данными in silico, показывающими, что ни один из проверенных GC-индуцибельных miR не нацелен на мРНК GR-3’UTR (фиг. 5E и таблица S3). В целом, эти результаты предполагают, что mir-150-5p может опосредовать косвенные эффекты, подобные GR.
Рисунок 5. Трансфекция миметиком Mir-150-5p запускает экспрессию специфического гена GR в MM1S, но не в клетках MM1R, без модуляции уровней белка GR.
( A ) Количественная оценка синтетических уровней mir-150-5p на основе QPCR через 72 часа после трансфекции клеток MM1S и MM1R. Уровни экспрессии miRNA были нормализованы для микроРНК SNORD 95 и SNORD 96A и представлены относительно контроля MOCK. Гистограммы представляют относительные уровни miRNA (среднее ± стандартная ошибка среднего) трех независимых экспериментов. Средние значения с ***, **, * значительно отличаются (p <0,001, <0,01 или <0,05) от контрольных установок в соответствии с двухфакторным дисперсионным анализом ANOVA (пост-тесты Бонферрони).( B ) Сравнение верхних обогащенных путей (IPA) изменений мРНК в клетках MM1S, обработанных в течение 72 часов до 1 мкМ Dex, с изменениями в клетках MM1S и MM1R после 72 часов ложной трансфекции трансфекции синтетическим mir-150-5p. ( C ) Уровни белка NR3C1 / GRα, обнаруженные после 72-часовой трансфекции синтетического миметика mir-150-5p в клетки MM1S. ( D ) Диаграмма Венна, которая представляет собой перекрытие экспериментально подтвержденных GC-индуцибельных микроРНК в клетках MM1S со списком микроРНК, предположительно нацеленных на 3’UTR NR3C1.( E ) QPCR-анализ уровней мРНК NR3C1 / GRα, присутствующих в клетках MM1S после 72-часовой трансфекции синтетическим миметиком mir-150-5p (левая панель) или после 72-часовой обработки Dex (1 мкМ) (правая панель).
https://doi.org/10.1371/journal.pone.0113842.g005
Гены, участвующие в путях пролиферации клеточного цикла, репарации ДНК и гибели клеток, которые регулируются в обработанном Dex и трансфицированном миметиком mir-150-5p MM1S / Ячейки MM1R показаны на фиг. 6A и 6B или обобщены в таблице S4.Чтобы дополнительно охарактеризовать перекрестные помехи между обработкой GC и трансфекцией mir-150-5p, мы также выполнили микроматричный анализ комбинаций mir-150-5p с 1 нМ Dex, поскольку комбинации обработки 1 мкМ Dex не давали РНК качества матрицы из-за чрезмерной цитотоксичности . Как можно заметить из фиг. 6A и 6B, обработка 1 нМ Dex вызывает более слабые изменения экспрессии генов по сравнению с обработкой 1 мкМ Dex. Кроме того, поскольку уровни mir-150-5p после трансфекции увеличиваются в 30 раз (фиг. 6A) по сравнению с 3-кратной индукцией после обработки Dex (фиг.3D), сверхэкспрессия миметика mir-150-5p вызывает более сильные изменения экспрессии генов, чем обработка 1 нМ Dex. Таким образом, комбинирование трансфекции mir-150-5p с обработкой 1 нМ Dex не приводило к дальнейшему увеличению изменений экспрессии генов, вызванных одной трансфекцией mir-150-p (фиг. 7A и B). Дальнейшее подробное сравнение активации GC и mir-150-5p (т.е. GILZ и FKBP5) (рис. 7A) или репрессии (IL23A, SKP2, BUB, SREBF1) (рис. 7B) типичных генов-мишеней GR выявило аналогичные эффекты в MM1S, но отсутствие эффектов в MM1R.Например, трансактивация генов FKBP5 и GILZ может наблюдаться после трансфекции mir-150-5p и обработки GC в клетках MM1S, хотя эффекты mir-150-5p на экспрессию FKBP5 сильнее. Аналогичным образом репрессия генов-мишеней GR IL23A, SKP2, BUB или SREBF1 может наблюдаться при обработке Dex, а также после трансфекции mir-150-5p. Наконец, помимо общих регулируемых генов, можно было идентифицировать дополнительные гены-мишени GR, на которые более избирательно влияла трансфекция mir-150-5p (SCNN1G) или обработка GC (GILZ, DUSP1, SGK1), что указывает на сложную тонкую настройку регуляции GR (рис. .7C).
Рисунок 6. Специфические для Mir-150-5p изменения мРНК в MM1S связаны с путями, связанными с гибелью и выживанием клеток, клеточным циклом, клеточным ростом и пролиферацией (A – B)
mir-150-5p-специфическая регуляция мРНК генов участвует в (i) клеточном цикле, (ii) сборке и организации клеток, (iii) репликации, рекомбинации и репарации ДНК, (iv) росте и пролиферации клеток, (v) гибели и выживании клеток. Результаты матрицы экспрессии генов Illumina BeadChip представлены в виде гистограмм, отражающих среднее кратное изменение экспрессии генов из трех независимых экспериментов с клетками MM1S, обработанными в течение 72 часов 1 мкМ Dex, по сравнению с контрольными клетками (верхние панели A – B) или MM1S. клетки, трансфицированные в течение 72 часов синтетическим mir-150-5p по сравнению с ложной трансфекцией, обработанные в течение 72 часов 1 нМ Dex или их комбинацией по сравнению с контрольными образцами (нижние панели A – B).
https://doi.org/10.1371/journal.pone.0113842.g006
Рис. 7. Трансфекция Mir-150-5p имитирует GR-подобный ответ на активацию и репрессию гена в MM1S и отсутствие ответа в клетках MM1R.
( A ) Проверка индукции мРНК GILZ (TSC22D3) и FKBP5 мРНК GILZ (TSC22D3) и FKBP5 в клетках MM1S и MM1R с помощью количественной ПЦР в реальном времени после 72-часовой обработки 1 нМ или 1 мкМ Dex (левая панель) или через 72 часа трансфекция синтетическим mir-150-5p, 72-часовая обработка 1 нМ Dex или их комбинацией (правая панель).Данные представляют собой значения (среднее ± стандартная ошибка среднего) трех независимых экспериментов, нормализованные для гена домашнего хозяйства РНК 28S и относительно соответствующих контрольных установок без обработки / растворителя (UT) или MOCK. Средние значения с ***, **, * значительно отличаются (p <0,001, <0,01 или <0,05) от контрольной установки, как определено с помощью двухфакторного дисперсионного анализа (посттест Бонферрони). ( B ) Проверка уровней репрессии мРНК MYB, IL23A, SKP2, BUB1, SREBF1 в клетках MM1S или MM1R с помощью количественной ПЦР в реальном времени (QPCR) после 72-часовой обработки 1 нМ или 1 мкМ Dex или после 72-часовой трансфекции синтетическим mir-150-5p, 72-часовая обработка 1 нМ Dex или их комбинацией.Данные представляют собой значения (среднее ± стандартная ошибка среднего) трех независимых экспериментов, нормализованные для гена домашнего хозяйства РНК 28S и относительно соответствующих контрольных установок без обработки / растворителя (UT) или MOCK. Средние значения с ***, **, * значительно отличаются (p <0,001, <0,01 или <0,05) от контрольной установки, как определено с помощью двухфакторного дисперсионного анализа (посттесты Бонферрони) ( C ), результаты матрицы экспрессии генов Illumina BeadChip выбранные гены представлены в виде гистограмм, отражающих среднее кратное изменение экспрессии генов из трех независимых экспериментов с клетками MM1S, обработанными в течение 72 часов 1 мкМ Dex, или же с клетками MM1S, трансфицированными в течение 72 часов синтетическим mir-150-5p по сравнению с ложной трансфекцией. , обработанные в течение 72 часов 1 нМ Dex или их комбинацией по сравнению с контрольными образцами.
https://doi.org/10.1371/journal.pone.0113842.g007
GR-подобный генный ответ, запускаемый трансфекцией mir150-5p, связан со сложной регуляцией факторов транскрипции, шаперонов hsp, киназ, белков клеточного цикла, стресса развернутого белка (UPR), AMPK / mTOR и хемокин-специфичных сигнальных сетей
Из-за отсутствия прямых эффектов mir-150-5p на уровни мРНК GR (NR3C1) и белка, мы предположили, что GR-подобные нисходящие эффекты могут быть вызваны регуляцией mir-150-5p возможных партнеров по взаимодействию с GR (факторы транскрипции, шапероны). или альтернативные ядерные рецепторы гормонов (рис.8A – B). Хотя установки mir-150-5p и Dex снижают уровни мРНК MYB и ATF3, для факторов транскрипции FOS наблюдались противоположные ответы. Особый интерес представляет MYB — ключевой регулятор GR-зависимой экспрессии генов при лейкемии [51], [52], [53], [54], [55], [56], [57] (рис. 8A).
Рисунок 8. Трансфекция Мир-150-5р связана со сложной регуляцией факторов транскрипции, рецепторов гормонов и шаперонов.
Illumina BeadChip Результаты экспрессии генов факторов транскрипции ( A ), рецепторов ядерных гормонов ( B ) и шаперонов ( C ) представлены в виде столбчатых диаграмм, отражающих среднее изменение кратности экспрессии генов из трех независимых экспериментов MM1S, обработанный в течение 72 часов 1 мкМ Dex по сравнению с контрольной установкой (UT), или клетки MM1S, трансфицированные в течение 72 часов синтетической трансфекцией mir-150-5p по сравнению с ложной трансфекцией.
https://doi.org/10.1371/journal.pone.0113842.g008
Что касается рецепторов гормонов, большинство рецепторов гормонов оставались постоянными после трансфекции mir-150-5p (рис. 8B), хотя уровни NR3C2 ( MR) может быть обнаружен в установке с помощью mir-150-5p, тогда как уровни NR3C2 остаются стабильными в присутствии Dex. Интересно, что как следствие реципрокного перекрестного взаимодействия действий NR3C2 (MR) и NR3C1 (GR), подавление функции MR действительно может приводить к независимой от лиганда стимуляции действий GR [58].Затем мы оценили изменения уровней транскрипции молекул шаперонов рецепторов гормонов в присутствии mir-150-5p. Хотя большинство мРНК HSP остаются незатронутыми, значительные изменения в уровнях HSPA8 и HSP90AB1 могут быть обнаружены после трансфекции mir-150-5p, но не с обработкой Dex, что может дополнительно способствовать тонкой настройке базальной активности рецепторов ядерных гормонов (рис. 8C) [59 ], [60], [61]. Кроме того, mir-150-5p избирательно снижает уровни транскриптов ATF4, JUN, NFE2L1 (NRF1), CEBP / B и IRF4, но не NFE2L2, TP53 и RelA (рис.8А).
Наконец, мы также исследовали возможные восходящие эффекты mir-150-5p на пути передачи сигналов рецепторной киназы. Активация (не) лиганд-независимого рецептора гормона через киназы, хемокины и цитокины хорошо документирована в настоящее время [62], [63], [64], [65], [66]. Что касается передачи сигналов хемокинами, было обнаружено, что трансфекция mir-150-5p и обработка GC увеличивают уровни мРНК CXCR4 и CX3CR1 и снижают уровни CXCR3, CCL3, CCL5, тогда как другие члены остаются неизменными (фиг. 9A). Интересно, что различные сообщения демонстрируют положительную роль CXCR4 / CXCL12 в гормонально-независимой активации рецепторов [65], [66], [67], [68], [69].Неожиданно мы наблюдали значительное увеличение уровней CXCR4 после трансфекции mir-150-5p, в противоположность более ранним сообщениям, демонстрирующим подавление мРНК CXCR4 его 3′-UTR через mir-150-5p [68], [70]. Хотя различные исследования иллюстрируют опухолевые и прометастатические эффекты CXCR4 [71], [72], его экспрессия также играет ключевую роль в гематопоэзе [73] и недавно была описана как хороший прогностический индикатор при множественной миеломе [74]. Наконец, мы также выявили заметные эффекты на эффекторы клеточного цикла (CDKN1A) и пути передачи сигналов UPR / mTOR (DDIT3, DDIT4, TXNIP) и псевдокиназу Ser / Thr семейства Триббла (TRIB3) [75], которые в целом могут пересекаться с передачей сигналов рецептора глюкокортикоидов. ответ на терапию ГК в клетках ММ [76], [77], [78] (рис.9Б). Особо следует отметить, что участие C / EBP и ATF4 (Fig. 8A) было продемонстрировано в регуляции транскрипции UPR стрессовых белков DDIT3, DDIT4, TXNIP [79], [80], что дополнительно подтверждает наши результаты.
Рисунок 9. Трансфекция Mir-150-5p связана со сложной регуляцией хемокинов, клеточного цикла, передачи сигналов UPR / mTOR.
Illumina BeadChip Результаты экспрессии гена хемокиновых рецепторов или лигандов ( A ) или регуляторов клеточного цикла, развернутого белкового стресса (передача сигналов UPR / mTOR) ( B ) представлены в виде столбиков, отражающих среднее кратность экспрессии гена. изменение от трех независимых экспериментов MM1S, обработанных в течение 72 часов 1 мкМ Dex по сравнению с контрольной установкой (UT), или еще клеток MM1S, трансфицированных в течение 72 часов синтетической трансфекцией mir-150-5p по сравнению с ложной трансфекцией.
https://doi.org/10.1371/journal.pone.0113842.g009
Слабая сенсибилизация ответа на терапию низкими дозами GC миметической трансфекцией mir-150-5p может быть отменена трансфекцией mir-150-5p-специфичного антагомира в клетки MM1S
Поскольку mir-150-5p модулирует различные гены, связанные с клеточным циклом, клеточной пролиферацией и путями гибели клеток, мы затем хотели измерить гибель клеток, вызванную эктопической трансфекцией mir-150-5p. Удивительно, но с помощью МТТ-анализа через 24, 48 и 72 часа после трансфекции клеток MM1S увеличением количества mir150-5p не наблюдалось существенной индукции гибели клеток (рис.10А). Последнее предполагает, что изменения экспрессии генов, вызванные mir-150-5p, недостаточны для запуска вызванной GC гибели клеток, и необходимы дополнительные действия GC для уничтожения клеток [15]. Затем мы оценили, может ли эктопическая трансфекция mir-150-5p сенсибилизировать к индуцированной GC гибели клеток. В этом отношении ложные и трансфицированные mir-150-5p клетки MM1S подвергали воздействию различных доз Dex в течение 72 часов, и выживаемость клеток снова оценивали с помощью МТТ-анализа. Из фиг. 10В можно наблюдать, что присутствие mir-150-5p слабо увеличивает индуцированную GC гибель клеток при наномолярных (нМ) концентрациях, хотя при микромолярных (мкМ) концентрациях обработки Dex не было выявлено никаких значительных эффектов.Более того, после обработки GC клеток MM1S, трансфицированных специфическим антагомиром mir-150-5p, чтобы обратить вспять эффекты mir-150-5p, гибель клеток может быть обращена вспять при низких дозах GC. Это побудило нас оценить соответствующие изменения в экспрессии генов после обработки GC клеток MM1S, трансфицированных антагомиром mir-150-5p. Хотя мы смогли подтвердить слабую GC-специфическую повышающую регуляцию mir150-5p и умеренное снижение уровней mir 150-5p при обработке GC клеток MM1S, трансфицированных mir-150-5p-специфическим антагомиром (рис.10C) подходы с использованием массивов казались недостаточно чувствительными, чтобы идентифицировать слабые изменения в репрессии или активации генов в обработанных GC клетках в присутствии или в отсутствие антагомира (фиг. 10D-E и данные не показаны).
Фигура 10. Слабая сенсибилизация ответа на терапию низкими дозами GC посредством трансфекции миметиком mir-150-5p может быть отменена трансфекцией mir-150-5p-специфичного антагомира в клетки MM1S.
( A ) Количественная оценка выживаемости клеток MM1S на основе МТТ после 24, 48 или 72 часов трансфекции увеличивающимися дозами синтетического миметика mir-150-5p по сравнению с имитационным контролем трансфекции ( B ) Количественная оценка на основе МТТ Выживаемость клеток MM1S после 72-часовой обработки увеличивающимися дозами Dex в сочетании либо с имитацией трансфекции, либо с трансфекцией синтетическим миметиком mir-150-5p или специфическим антагомиром mir-150-5p (как показано на рисунке).Представленные данные представляют собой относительные уровни выживаемости клеток (среднее значение ± стандартная ошибка среднего) трех независимых экспериментов. Средние значения с ***, **, * значительно отличаются (p <0,001, <0,01 или <0,05) от контрольной установки, как определено с помощью двухфакторного дисперсионного анализа (посттесты Бонферрони). ( C ) Количественная ПЦР в реальном времени (QPCR) количественная оценка уровней транскрипции mir-150-5p после 72-часовой обработки 1 нМ Dex в комбинации или без трансфекции антагомира, специфичного для mir-150-5p. ( D – E ) Результаты экспрессии гена Illumina BeadChip для подмножества генов представлены в виде гистограмм, отражающих среднее кратное изменение из трех независимых экспериментов с клетками MM1S, обработанными в течение 72 часов 1 нМ Dex vs.контрольная установка, либо в сочетании с трансфекцией специфическим антагомиром mir-150-5p, либо нет.
https://doi.org/10.1371/journal.pone.0113842.g010
Обсуждение
Снижение пролиферации и индукция гибели клеток в результате апоптоза — два основных клеточных события, направленных на уменьшение роста и уничтожение раковых клеток. Известно, что ГК вызывают остановку клеточного цикла, предшествующую апоптозу. Лечение раковых клеток более высокими дозами Dex может дестабилизировать глюкокортикоидный рецептор (GR), что в конечном итоге приводит к устойчивости к GC [38], [81], [82], [83].Характеристика miRNA и их целевых мРНК, участвующих в регуляции ответа на терапию GC и / или резистентности, является областью интенсивных исследований, и относительно мало известно о регуляторных сетях GC-специфической miRNA. Комбинируя анализ транскриптома по всему геному и анализ пути изобретательности, мы предполагали раскрыть биологические эффекты GC-чувствительных miRNA и их предполагаемых мРНК-мишеней в терапии MM GC. Мы идентифицировали 30 GC-индуцибельных и 14 GC-репрессированных микроРНК, которые могут модулировать несколько аспектов ответа на GC-терапию при ММ, включая клеточный цикл, пролиферацию клеток, репликацию ДНК, экспрессию генов и пути гибели клеток.Сходным образом, исследование GC-чувствительных микроРНК в биоптатах слизистой оболочки пациентов с эозинофильным эзофагитом охарактеризовало 32 GC-индуцибельные и 4 GC репрессированные микроРНК, из которых 7 GC-индуцибельных микроРНК являются общими с нашим списком, независимо от клеточного контекста [84]. Затем мы выбрали mir-150-5p как наиболее интересную GC-индуцибельную микроРНК для дальнейшего исследования из-за его постоянной активации GC в клетках MM1S и его возможной опухолевой супрессорной функции при (злокачественном) гематопоэзе [26], [54], [85] ], [86], [87].Согласно анализу IPA, предсказанные мишени мРНК mir-150-5p были связаны с клеточным циклом и ответами клеточной пролиферации, предшествующими апоптозу. В соответствии с анализом IPA, эктопическая экспрессия mir-150-5p в присутствии низких концентраций GC, как было обнаружено, слабо сенсибилизирует индуцированную GC гибель клеток, тогда как антагомир mir-150-5p может обратить этот эффект. Примечательно, что при эктопической трансфекции mir-150-5p или обработке GC в клетках MM1S были идентифицированы различные мРНК, которые аналогично реагируют на повышенные уровни mir-150-5p или обработку GC (т.е.е. MYB, IL23A, SKP2, BUB1, SREBP1, FKBP5). Это говорит о том, что эффекты mir-150-5p тесно связаны с функцией GR. Более того, GR-специфические эффекты mir-150-5p полностью теряются в клетках MM1R, и mir-150-5p не может восстанавливать чувствительность GC в клетках MM1R, лишенных экспрессии белка GR. Поскольку сверхэкспрессия mir-150-5p существенно не изменила уровни экспрессии GR, мы полагаем, что GR-подобные эффекты могут быть вызваны сложной непрямой регуляцией факторов транскрипции, взаимодействующих с GR (MYB, C / EBP, JUN, FOS) или рецепторов гормонов (NR3C2 / MR), шапероны GR (FKBP5, HSPA8, HSP90AB1), а также различные эффекторы стресса развернутого белка (ATF4, DDIT3, DDIT4, TXNIP) и хемокин-специфическая передача сигналов (CXCR3, CCL3, CCL5, CXCR4).Особо следует отметить, что избирательное снижение уровня NR3C2 (MR) mir-150-5p может приводить к реципрокной стимуляции действий GR [58]. GR-зависимые эффекты mir-150-5p также предполагают, что GR-белок может вносить вклад в специфическую посттранскрипционную регуляцию mir-150-5p. Напр., Зависимая от NR3C1 (GR) посттранскрипционная регуляция CCL2 и CCL7, но не мРНК CCL5, была продемонстрирована при связывании РНК GR с мотивом, богатым GU в 5’UTR [88]. Альтернативно, mir-150-5p может модулировать связывание GR / ДНК по аналогии с длинной некодирующей РНК Gas5 [24].
В целом мы идентифицировали различные новые и ранее опубликованные GC-индуцибельные микроРНК. Кроме того, множественные мРНК, отвечающие за mir-150-5p, связаны с GC-индуцированной пролиферацией клеток и ответом на терапию гибелью клеток при множественной миеломе. Исследования функциональной выживаемости клеток в клетках MM1S подтвердили слабые сенсибилизирующие эффекты терапии ГК сверхэкспрессией mir-150-5p при низкой дозе GC, тогда как противоположные эффекты могли наблюдаться в присутствии специфического антагомира mir-150-5p. Однако, хотя анализы QPCR выявили обратное изменение уровней mir-150-5p, индуцированных Dex, после трансфекции антагомиром и ослабление индуцированной GC гибели клеток, последующий анализ микроматрицы не смог выявить значительных специфичных для mir-150-5p изменений уровней мРНК, регулируемых GC (рис.10D – E и данные не показаны). Мы полагаем, что массивному исследованию может не хватать чувствительности и статистической мощности для выявления значительных изменений мРНК при обращении 2-кратной зависящей от времени GC-специфической индукции уровней mir-150p его антагомиром, тогда как 30-кратная избыточная экспрессия на уровнях mir150-5p после трансфекция может вызывать более устойчивые эффекты в массивах экспрессии генов. В аналогичном исследовании неспособность антагомиров обратить вспять изменения экспрессии специфического гена mir-92, несмотря на сниженные уровни mir-92 в присутствии его антагомира, была объяснена возможной избыточностью в сетях мРНК, регулируемых mir-92 [89].В том же направлении, поскольку мы идентифицировали различные индуцируемые GC микроРНК, связанные с путем NR3C1, мы также не можем исключить избыточные регулируемые mir-150-5p сети мРНК, участвующие в регуляции NR3C1. Например, ранние GC-индуцируемые микроРНК, т.е. mir-26b, mir125-5p и mir184, могут быть ответственны за последующее повышение уровней mir150-5p для точной настройки активности GR (рис. 3C). Недавние отчеты также показывают связь mir150-5p с mir124- и mir155-сетями [90], [91]. Альтернативно, избыточные изомиры МИР-150-5р могут ускользать от контроля антагомира [91].Будущие исследования потребуются для дальнейшего раскрытия специфичности или избыточности mir-150-5p-специфичных сетей NR3C1 с помощью дополнительных транскриптомных и протеомных подходов. Наконец, недавние сообщения, демонстрирующие секрецию mir-150-5p в микровезикулы, могут позволить разработать диагностический биомаркер для отслеживания ответа на терапию GC, иначе комбинация низких доз GC с синтетическими везикулами mir-150-5p может повысить чувствительность ответа на терапию GC при лимфоидном раке [ 92], [93].
Дополнительная информация
Таблица S1.
IPA-прогнозирование микроРНК-мишеней на основе корреляции транскрипционных изменений конкретных микроРНК (вверх / вниз) с изменениями уровней мРНК (противоположное направление вниз / вверх) и соответствующих путей в клетках MM1S, обработанных в течение 72 часов Dex.
https://doi.org/10.1371/journal.pone.0113842.s001
(XLSX)
Таблица S2.
Перекрестное сравнение 1203 Dex-зависимых генов-мишеней из клеток MM1S, подвергнутых воздействию в течение 72 часов, с 1 мкМ Dex (отбор генов производился на основе значения p <0.05 и абсолютное кратное изменение> 1,5) и 3626 предсказали цели hsa-mir-150-5p через базу данных miRWalk (предсказание по крайней мере 4 из 10 доступных алгоритмов, то есть DIANAmT, miRanda, miRDB, miRWalk, RNAhybrid, PICTAR4, PICTAR5, PITA, RNA22 и Targetscan) 196 общих мРНК / генных мишеней были идентифицированы в MM1S, которые остаются неизменными в клетках MM1R.
https://doi.org/10.1371/journal.pone.0113842.s002
(XLSX)
Таблица S4.
Подробный список генов, участвующих в путях пролиферации клеточного цикла, репарации ДНК и гибели клеток, которые аналогичным образом регулируются в обработанных Dex и трансфицированных миметиком mir-150-5p клетках MM1S, но которые остаются неизменными в клетках MM1R (значения представляют (кратное изменение по сравнению с контрольными клетками)).
https://doi.org/10.1371/journal.pone.0113842.s004
(XLSX)
Благодарности
Авторы благодарят всех сотрудников лаборатории за ценные научные обсуждения.
Вклад авторов
Задумал и спроектировал эксперименты: AP KH SG PM JV WVB. Проведены эксперименты: AP KODB JD CSC GVC KH PM JV WVB. Проанализированы данные: АП КОДБ СН КЛ WVB. Предоставленные реагенты / материалы / инструменты анализа: AP KODB GVC KH SG PM JV WVB. Участвовал в написании рукописи: AP KH SG WVB.
Ссылки
- 1. Фонсека Р., Барлоги Б., Батай Р., Ублюдок С., Бергсагель П. Л. и др. (2004) Генетика и цитогенетика множественной миеломы: отчет семинара. Cancer Res 64: 1546–1558.
- 2. Bommert K, Bargou RC, Stuhmer T (2006) Пути передачи сигналов и выживания при множественной миеломе. Eur J Cancer 42: 1574–1580.
- 3. Кастринакис Н.Г., Горгулис В.Г., Фукас П.Г., Димопулос М.А., Киттас С. (2000) Молекулярные аспекты множественной миеломы.Энн Онкол 11: 1217–1228.
- 4. Кайл Р.А., Раджкумар С.В. (2008) Множественная миелома. Кровь 111: 2962–2972.
- 5. Несвицкий Р., Джаябалан Д.С., Христос П.Дж., Фурст Дж. Р., Наиб Т. и др. (2008) Комбинированная терапия BiRD (биаксин [кларитромицин] / ревлимид [леналидомид] / дексаметазон) приводит к высоким показателям полного и общего ответа при не получавшей лечения симптоматической множественной миеломе. Кровь 111: 1101–1109.
- 6. Гей Ф., Раджкумар С.В., Коулман М., Кумар С., Марк Т. и др.(2010) Кларитромицин (Биаксин) -леналидомид в низких дозах дексаметазона (BiRd) по сравнению с леналидомидом в низких дозах дексаметазона (Rd) при недавно диагностированной миеломе. Am J Hematol 85: 664–669.
- 7. Росси А., Марк Т., Джаябалан Д., Христос П., Зафар Ф. и др. (2013) BiRd (кларитромицин, леналидомид, дексаметазон): обновленная информация о долгосрочной терапии леналидомидом у ранее нелеченных пациентов с множественной миеломой. Кровь 121: 1982–1985.
- 8. Korde N, Kristinsson SY, Landgren O (2011) Моноклональная гаммопатия неопределенного значения (MGUS) и тлеющая множественная миелома (SMM): новые биологические идеи и разработка стратегий раннего лечения.Кровь 117: 5573–5581.
- 9. Piovan E, Yu J, Tosello V, Herranz D, Ambesi-Impiombato A, et al. (2013) Прямая реверсия устойчивости к глюкокортикоидам путем ингибирования AKT при остром лимфобластном лейкозе. Cancer Cell 24: 766–776.
- 10. Тернер Дж. Д., Мюллер С. П. (2005) Структура 5 ‘нетранслируемой области гена глюкокортикоидного рецептора (NR3C1): идентификация и распределение в тканях множества новых экзонов человека 1. J Mol Endocrinol 35: 283–292.
- 11. Шмидт С., Райнер Дж., Плонер С., Пресул Е., Римл С. и др.(2004) Апоптоз, индуцированный глюкокортикоидами, и резистентность к глюкокортикоидам: молекулярные механизмы и клиническое значение. Cell Death Differ 11 Приложение 1: S45–55.
- 12. Де Босшер К. (2010) Селективные модуляторы рецепторов глюкокортикоидов. Дж. Стероид Биохим Мол Биол 120: 96–104.
- 13. Kumar R, Thompson EB (2005) Регулирование генов глюкокортикоидным рецептором: структура: взаимосвязь функций. Дж. Стероид Биохим Мол Биол 94: 383–394.
- 14. Де Босчер К., Бек И.М., Хегеман Г. (2010) Классические глюкокортикоиды против нестероидных модуляторов глюкокортикоидных рецепторов: выживаемость наиболее приспособленного регулятора иммунной системы? Иммунное поведение мозга 24: 1035–1042.
- 15. Kfir-Erenfeld S, Yefenof E (2014) Негеномные события, определяющие чувствительность гемопоэтических злокачественных новообразований к апоптозу, индуцированному глюкокортикоидами. Cancer Immunol Immunother 63: 37–43.
- 16. Kfir-Erenfeld S, Sionov RV, Spokoini R, Cohen O, Yefenof E (2010) Сети протеинкиназ, регулирующие индуцированный глюкокортикоидами апоптоз кроветворных раковых клеток: фундаментальные аспекты и практические соображения. Лимфома Лейка 51: 1968–2005.
- 17.De Iudicibus S, Franca R, Martelossi S, Ventura A, Decorti G (2011) Молекулярный механизм устойчивости к глюкокортикоидам при воспалительном заболевании кишечника. World J Gastroenterol 17: 1095–1108.
- 18. Барнс П.Дж., Адкок И.М. (2009) Устойчивость к глюкокортикоидам при воспалительных заболеваниях. Ланцет 373: 1905–1917.
- 19. Чжан Дж., Яо Й, Сяо Ф, Лань Х, Ю Ц и др. (2013) Введение дексаметазона защищает мышей от повреждения почек, вызванного ишемией / реперфузией, путем подавления передачи сигналов PI3K / AKT.Int J Clin Exp Pathol 6: 2366–2375.
- 20. Schlossmacher G, Stevens A, White A (2011) Апоптоз, опосредованный глюкокортикоидными рецепторами: механизмы устойчивости в раковых клетках. J Endocrinol 211: 17-25.
- 21. Ли XJ, Luo XQ, Han BW, Duan FT, Wei PP и др. (2013) MicroRNA-100 / 99a, дерегулированная при остром лимфобластном лейкозе, подавляет пролиферацию и способствует апоптозу, регулируя пути передачи сигналов FKBP51 и IGF1R / mTOR. Br J Рак.
- 22.Хун И, Ву Дж, Чжао Дж, Ван Х, Лю И и др. (2013) miR-29b и miR-29c участвуют в контроле толл-подобных рецепторов индуцированного глюкокортикоидами апоптоза в плазматических дендритных клетках человека. PLoS One 8: e69926.
- 23. Pore SK, Choudhary A, Rathore B, Ganguly A, Sujitha P и др. (2013) Липосомный препарат миРНК, нацеленный на Hsp90, для системного противоопухолевого действия. Биоматериалы 34: 6804–6817.
- 24. Kino T, Hurt DE, Ichijo T, Nader N, Chrousos GP (2010) Некодирующая РНК gas5 является репрессором глюкокортикоидного рецептора, связанным с остановкой роста и голоданием.Научный сигнал 3: ra8.
- 25. Ambros V (2003) Пути микроРНК у мух и червей: рост, смерть, жир, стресс и время. Cell 113: 673–676.
- 26. Моррис В.А., Чжан А., Ян Т., Стируолт Д.Л., Рамамурти Р. и др. (2013) Экспрессия MicroRNA-150 индуцирует миелоидную дифференцировку клеток острого лейкоза человека и нормальных гематопоэтических клеток-предшественников. PLoS One 8: e75815.
- 27. Kong YW, Ferland-McCollough D, Jackson TJ, Bushell M (2012) микроРНК в лечении рака.Ланцет Онкол 13: e249–258.
- 28. Гринштейн С., Кретт Н.Л., Куросава Ю., Ма С., Чаухан Д. и др. (2003) Характеристика линий клеток множественной миеломы человека (ММ) MM.1. Экспериментальная гематология 31: 271–282.
- 29. Суттана В., Манхеткорн С., Поомпимон В., Палагани А., Жохов С. и др. (2010) Дифференциальная хемосенсибилизация Р-гликопротеина, сверхэкспрессирующего клетки K562 / Adr, с помощью витаферина А и полифенолов Сиамуа. Молочный рак 9:99.
- 30. Dunning MJ, Smith ML, Ritchie ME, Tavare S (2007) beadarray: классы R и методы для данных на основе шариков Illumina.Биоинформатика 23: 2183–2184.
- 31. Саид А.И., Шаров В., Уайт Дж., Ли Дж., Лян В. и др. (2003) TM4: бесплатная система с открытым исходным кодом для управления и анализа данных микрочипов. Биотехники 34: 374–378.
- 32. Местдаг П., Ван Влиерберг П., Де Веер А., Мут Д., Вестерманн Ф. и др. (2009) Новый и универсальный метод нормализации данных микроРНК RT-qPCR. Геном Биол 10: R64.
- 33. Местдаг П., Фейс Т., Бернард Н., Гюнтер С., Чен С. и др.(2008) Профилирование экспрессии miRNA в режиме «ствол-петля» RT-qPCR с высокой пропускной способностью с использованием минимальных количеств входной РНК. Нуклеиновые кислоты Res 36: e143.
- 34. Dweep H, Sticht C, Pandey P, Gretz N (2011) miRWalk – база данных: предсказание возможных сайтов связывания miRNA путем «прогулок» по генам трех геномов. Дж. Биомед Информ, 44: 839–847.
- 35. Tessel MA, Benham AL, Krett NL, Rosen ST, Gunaratne PH (2011) Роль микроРНК в регулировании глюкокортикоидного ответа и устойчивости при множественной миеломе.Гормональный рак 2: 182–189.
- 36. Murray MY, Rushworth SA, Zaitseva L, Bowles KM, Macewan DJ (2013) Ослабление индуцированной дексаметазоном гибели клеток при множественной миеломе опосредуется экспрессией miR-125b. Клеточный цикл 12: 2144–2153.
- 37. Grugan KD, Ma C, Singhal S, Krett NL, Rosen ST (2008) Двойная регуляция индуцированной глюкокортикоидами лейциновой молнии (GILZ) рецептором глюкокортикоидов и путями PI3-киназы / AKT при множественной миеломе. Дж. Стероид Биохим. Мол Биол 110: 244–254.
- 38. Gross KL, Oakley RH, Scoltock AB, Jewell CM, Cidlowski JA (2011) Селективная регуляция альфа-изоформы глюкокортикоидного рецептора антиапоптотических генов в клетках остеосаркомы: новый механизм устойчивости к глюкокортикоидам. Мол эндокринол 25: 1087–1099.
- 39. Sanchez-Vega B, Krett N, Rosen ST, Gandhi V (2006) Изоформы транскрипции глюкокортикоидного рецептора и резистентность в клетках множественной миеломы. Mol Cancer Ther 5: 3062–3070.
- 40. Джентльмен Р. (2005) Решения в области биоинформатики и вычислительной биологии с использованием R и Bioconductor.Нью-Йорк: Springer Science + Business Media. XIX, 473 с.
- 41. Смит Г.К. (2004) Линейные модели и эмпирические байесовские методы для оценки дифференциальной экспрессии в экспериментах с микрочипами. Stat Appl Genet Mol Biol 3: Статья3.
- 42. Itani OA, Liu KZ, Cornish KL, Campbell JR, Thomas CP (2002) Глюкокортикоиды стимулируют экспрессию гена sgk1 человека путем активации GRE в его 5′-фланкирующей области. Am J Physiol Endocrinol Metab 283: E971–979.
- 43. Granes F, Roig MB, Brady HJ, Gil-Gomez G (2004) Активация Cdk2 действует выше митохондрии во время индуцированного глюкокортикоидами апоптоза тимоцитов.Eur J Immunol 34: 2781–2790.
- 44. Kullmann MK, Grubbauer C, Goetsch K, Jakel H, Podmirseg SR, et al. (2013) Ось p27-Skp2 опосредует вызванную глюкокортикоидами остановку клеточного цикла в клетках Т-лимфомы. Клеточный цикл 12: 2625–2635.
- 45. Крамер А., Грин Дж., Поллард Дж. Младший, Тугендрайх С. (2014) Подходы к причинно-следственному анализу в анализе пути изобретательности. Биоинформатика 30: 523–530.
- 46. Ли Х, Цянь В., Вен Х, Ву З., Чжуан К. и др. (2012) Глюкокортикоидный рецептор и последовательная активация P53 дексаметазоном опосредуют апоптоз и остановку клеточного цикла остеобластных клеток MC3T3-E1.PLoS One 7: e37030.
- 47. Обексер П., Хагенбухнер Дж., Рупп М., Сальвадор С., Хольцнер М. и др. (2009) p16INK4A сенсибилизирует клетки лейкемии человека к апоптозу, индуцированному FAS и глюкокортикоидами, посредством индукции BBC3 / Puma и репрессии MCL1 и BCL2. J Biol Chem 284: 30933–30940.
- 48. Багдассарян Н., Каталло Р., Махли М.А., Френч П., Чизат Ф. и др. (1998) Глюкокортикоиды индуцируют удлинение G1, а также S-фазы в нормальных стимулированных лимфоцитах человека: дифференциальные эффекты на регуляторные белки клеточного цикла.Exp Cell Res 240: 263–273.
- 49. Николсон Л., Холл А.Г., Редферн С.П., Ирвинг Дж. (2010) Модуляторы NFkappaB в модели устойчивого к глюкокортикоидам острого лимфобластного лейкоза у детей. Leuk Res 34: 1366–1373.
- 50. Блад Л.Г., Лиден Дж., Пазиранде А., Стропила I, Дальман-Райт К. и др. (2005) Идентификация генов-мишеней, участвующих в антипролиферативном эффекте глюкокортикоидов, раскрывает роль репрессии ядерного фактора (каппа) B. Мол эндокринол 19: 632–643.
- 51. Бянь З., Ли Л., Цуй Дж., Чжан Х., Лю Й. и др. (2011) Роль c-Myb, нацеленного на miR-150, в разрушении эпителия толстой кишки во время экспериментального колита мышей, индуцированного декстрансульфатом натрия, и язвенного колита человека. Дж. Патол 225: 544–553.
- 52. Chen S, Wang Z, Dai X, Pan J, Ge J и др. (2013) Повторная экспрессия микроРНК-150 индуцирует ВЭБ-положительную дифференцировку лимфомы Беркитта путем модуляции c-Myb in vitro. Cancer Sci 104: 826–834.
- 53.Цзян Х, Хуанг Х, Ли З, Ли И, Ван Х и др. (2012) Блокада созревания miR-150 за счет слияния MLL / MYC / LIN-28 необходима для MLL-ассоциированного лейкоза. Cancer Cell 22: 524–535.
- 54. Adams BD, Guo S, Bai H, Guo Y, Megyola CM и др. (2012) Функциональный скрининг in vivo раскрывает опосредованную miR-150 регуляцию реакции кроветворения на повреждение. Cell Rep 2: 1048–1060.
- 55. Zheng Q, Zhou L, Mi QS (2012) МикроРНК miR-150 участвует в развитии и функционировании Valpha14-инвариантных NKT-клеток.J Immunol 188: 2118–2126.
- 56. Sarvaiya PJ, Schwartz JR, Geng CD, Vedeckis WV (2012) c-Myb взаимодействует с рецептором глюкокортикоидов и регулирует его уровень в клетках пред-B-острой лимфобластной лейкемии. Эндокринол клеток Mol 361: 124–132.
- 57. Geng CD, Vedeckis WV (2011) Новый, специфичный для клонов, механизм ауторегуляции экспрессии гена рецептора глюкокортикоидов человека в 697 клетках пре-B-острого лимфобластного лейкоза. Мол эндокринол 25: 44–57.
- 58.Хаттори Т., Мурасе Т., Ивасе Э, Такахаши К., Отаке М. и др. (2013) Глюкокортикоид-индуцированная гипертензия и сердечное повреждение: эффекты антагонизма минералокортикоидных и глюкокортикоидных рецепторов. Nagoya J Med Sci 75: 81–92.
- 59. Jaaskelainen T, Makkonen H, Palvimo JJ (2011) Стероидная регуляция FKBP51 и его роль в передаче сигналов гормона. Curr Opin Pharmacol 11: 326–331.
- 60. Tatro ET, Eshops IP, Kaul M, Achim CL (2009) Модуляция ядерной транслокации глюкокортикоидных рецепторов в нейронах с помощью иммунофилинов FKBP51 и FKBP52: последствия для большого депрессивного расстройства.Brain Res 1286: 1–12.
- 61. Бек И.М., Дреберт З.Дж., Хойя-Ариас Р., Бахар А.А., Девос М. и др. (2013) Соединение А, селективный модулятор рецептора глюкокортикоидов, усиливает активацию промотора гена белка теплового шока Hsp70. PLoS One 8: e69115.
- 62. Robertson S, Hapgood JP, Louw A (2013) Концентрация глюкокортикоидных рецепторов и способность димеризоваться влияют на ядерную транслокацию и распределение. Стероиды 78: 182–194.
- 63. Ху А., Джозефсон МБ, Динер Б.Л., Нино Г., Сюй С. и др.(2013) Проастматические цитокины регулируют нелигандзависимую и зависимую от лиганда передачу сигналов глюкокортикоидных рецепторов в гладких мышцах дыхательных путей. PLoS One 8: e60452.
- 64. Эйкельберг О., Рот М., Лоркс Р., Брюс В., Рудигер Дж. И др. (1999) Лиганд-независимая активация глюкокортикоидного рецептора агонистами бета2-адренергического рецептора в первичных фибробластах легких человека и гладкомышечных клетках сосудов. J Biol Chem 274: 1005–1010.
- 65. Kasina S, Macoska JA (2012) Ось CXCL12 / CXCR4 способствует лиганд-независимой активации рецептора андрогена.Mol Cell Endocrinol 351: 249–263.
- 66. Sauve K, Lepage J, Sanchez M, Heveker N, Tremblay A (2009) Активация рецепторов эстрогена с положительной обратной связью по пути CXCL12-CXCR4. Cancer Res 69: 5793–5800.
- 67. Boudot A, Kerdivel G, Habauzit D, Eeckhoute J, Le Dily F и др. (2011) Дифференциальная эстрогеновая регуляция хемокиновых рецепторов CXCL12, CXCR4 и CXCR7, способствует росту эстрогенов в клетках рака груди. PLoS One 6: e20898.
- 68.Роллан-Тернер М., Горетти Э., Бускено М., Леонард Ф., Николя С. и др. (2013) Аденозин стимулирует миграцию эндотелиальных клеток-предшественников человека. Роль CXCR4 и микроРНК-150. PLoS One 8: e54135.
- 69. Родс Л.В., Шорт С.П., Нил Н.Ф., Сальво В.А., Чжу Ю.и др. (2011) Цитокиновый рецептор CXCR4 опосредует эстроген-независимый туморогенез, метастазирование и устойчивость к эндокринной терапии при раке груди человека. Cancer Res 71: 603–613.
- 70. Tano N, Kim HW, Ashraf M (2011) microRNA-150 регулирует мобилизацию и миграцию мононуклеарных клеток костного мозга путем нацеливания на Cxcr4.PLoS One 6: e23114.
- 71. Cojoc M, Peitzsch C, Trautmann F, Polishchuk L, Telegeev GD, et al. (2013) Новые цели в лечении рака: роль оси CXCL12 / CXCR4. Onco Targets Ther 6: 1347–1361.
- 72. Han TT, Fan L, Li JY, Xu W (2014) Роль хемокинов и их рецепторов в хроническом лимфоцитарном лейкозе: функция в микросреде и таргетная терапия. Рак Biol Ther 15: 3–9.
- 73. Zou YR, Kottmann AH, Kuroda M, Taniuchi I, Littman DR (1998) Функция хемокинового рецептора CXCR4 в гематопоэзе и развитии мозжечка.Nature 393: 595–599.
- 74. Бао Л, Лай И, Лю И, Цинь И, Чжао Х и др. (2013) CXCR4 является хорошим прогностическим показателем выживаемости у пациентов с множественной миеломой. Leuk Res 37: 1083–1088.
- 75. Лян К.Л., Риши Л., Кишан К. (2013) Tribbles при остром лейкозе. Кровь 121: 4265–4270.
- 76. Das I, Png CW, Oancea I, Hasnain SZ, Lourie R и др. (2013) Глюкокортикоиды снижают стресс ER кишечника за счет усиления сворачивания белков и деградации неправильно свернутых белков.J Exp Med 210: 1201–1216.
- 77. Гавами С., Эшраги М., Анде С.Р., Чазин В.Дж., Клониш Т. и др. (2010) S100A8 / A9 индуцирует аутофагию и апоптоз посредством АФК-опосредованного перекрестного взаимодействия между митохондриями и лизосомами, в котором участвует BNIP3. Cell Res 20: 314–331.
- 78. Ли С, Чен Х, Дин Ф, Чжан И, Ло А и др. (2009) Новый ген-мишень p53, S100A9, индуцирует p53-зависимый клеточный апоптоз и опосредует путь апоптоза p53. Biochem J 422: 363–372.
- 79.Полман Я.А., Хантер Р.Г., Спекснейдер Н., ван ден Овер Я.М., Коробко О.Б. и др. (2012) Глюкокортикоиды модулируют путь mTOR в гиппокампе: разные эффекты в зависимости от стресса в анамнезе. Эндокринология 153: 4317–4327.
- 80. Protiva P, Hopkins ME, Baggett S, Yang H, Lipkin M, et al. (2008) Подавление роста клеток рака толстой кишки полиизопренилированными бензофенонами связано с индукцией ответа эндоплазматического ретикулума. Int J Cancer 123: 687–694.
- 81.Ramamoorthy S, Cidlowski JA (2013) Изучение молекулярных механизмов действия рецепторов глюкокортикоидов от чувствительности к сопротивлению. Endocr Dev 24: 41–56.
- 82. Barnes PJ (2013) Устойчивость к кортикостероидам у пациентов с астмой и хронической обструктивной болезнью легких. J Allergy Clin Immunol 131: 636–645.
- 83. Gross KL, Lu NZ, Cidlowski JA (2009) Молекулярные механизмы, регулирующие чувствительность и устойчивость к глюкокортикоидам. Клеточный эндокринол 300: 7–16.
- 84. Лу С., Муккада В.А., Мангрей С., Кливленд К., Шиллингфорд Н. и др. (2012) Профилирование микроРНК в биоптатах слизистой оболочки пациентов с эозинофильным эзофагитом до и после лечения стероидами и взаимосвязь с мишенями мРНК. PLoS One 7: e40676.
- 85. He Y, Jiang X, Chen J (2013) Роль miR-150 в нормальном и злокачественном гематопоэзе. Онкоген.
- 86. Xiao C, Calado DP, Galler G, Thai TH, Patterson HC, et al. (2007) MiR-150 контролирует дифференцировку В-клеток, воздействуя на фактор транскрипции c-Myb.Ячейка 131: 146–159.
- 87. He Y, Jiang X, Chen J (2014) Роль miR-150 в нормальном и злокачественном гематопоэзе. Онкоген 33: 3887–3893.
- 88. Ishmael FT, Fang X, Houser KR, Pearce K, Abdelmohsen K и др. (2011) Глюкокортикоидный рецептор человека как РНК-связывающий белок: глобальный анализ транскриптов, связанных с глюкокортикоидным рецептором, и идентификация целевого РНК-мотива. J Immunol 186: 1189–1198.
- 89. Sengul A, Santisuk R, Xing W, Kesavan C (2013) Системное введение антагомира, предназначенного для ингибирования miR-92, регулятора ангиогенеза, не смогло модулировать анаболический ответ скелета на механическую нагрузку.Physiol Res 62: 221–226.
- 90. Manoharan P, Basford JE, Pilcher-Roberts R, Neumann J, Hui DY et al. (2014) Пониженные уровни miR-124a и miR-150 связаны с повышенной экспрессией провоспалительного медиатора в KLF2-дефицитных макрофагах. J Biol Chem.
- 91. Ван М., Тан Л.П., Дейкстра М.К., ван Лом К., Робертус Дж.Л. и др. (2008) анализ miRNA при B-клеточном хроническом лимфоцитарном лейкозе: центры пролиферации, характеризующиеся низкой экспрессией miR-150 и высокой экспрессией BIC / miR-155.Дж. Патол 215: 13–20.
- 92. Ли Дж, Чжан И, Лю И, Дай Х, Ли В. и др. (2013) Опосредованный микровезикулами перенос микроРНК-150 от моноцитов к эндотелиальным клеткам способствует ангиогенезу. J Biol Chem 288: 23586–23596.
- 93. Almanza G, Anufreichik V, Rodvold JJ, Chiu KT, DeLaney A, et al. (2013) Синтез и доставка короткой некодирующей РНК B-лимфоцитами. Proc Natl Acad Sci U S A 110: 20182–20187.
Безопасность и фармакокинетика витаферина-А при остеосаркоме высокой степени злокачественности: испытание фазы I
Журнал аюрведы и интегративной медицины xxx (xxxx) xxxСписки содержания доступны на ScienceDirect
Домашняя страница журнала аюрведы и интегративной медицины: http: // elsevier.com / locate / jaim
Оригинальная статья исследования
Безопасность и фармакокинетика витаферина-A при остеосаркоме высокой степени злокачественности: испытание фазы I Нишель Пирес a, Викрам Гота a, Ашиш Гулия b, Лал Хингорани c, Маниш Агарвал b, Ajay Puri b, * abc
Отдел клинической фармакологии, ACTREC, Мемориальный центр Тата, Харгар, Нави Мумбаи, 410210, Отделение костей и мягких тканей Индии, Мемориальный центр Тата, Парел, Мумбаи, 400012, Индия Pharmanza Herbal Pvt Ltd., Ананд, Гуджарат, Индия
articleinfo
аннотация
История статьи: Получено 31 августа 2018 г. Получено в пересмотренной форме 24 октября 2018 г. Принято 26 декабря 2018 г. Доступно онлайн xxx
Введение: Withaferin-A (WA), получено активное начало из традиционной индийской травы, известной как ашваганда или индийский женьшень, было показано, что она предотвращает и излечивает уретановые опухоли легких у мышей, а также подавляет рост пересаженной саркомы у мышей.Цели: в этом исследовании мы оценили безопасность и фармакокинетику WA у пациентов с запущенной стадией остеосаркомы высокой степени злокачественности. Методы. Исследование увеличения дозы I фазы было запланировано с использованием классического дизайна 3 × 3 (C33D). Когорты увеличения дозы включали 72, 108, 144 и 216 мг WA, вводимых в виде двух-четырех разделенных доз в день. В каждую когорту были включены по три пациента, и последний пациент наблюдался в течение не менее 30 дней на предмет любой ограничивающей дозу токсичности, прежде чем перейти в более высокую когорту.Фармакокинетические исследования проводили с использованием метода высокоэффективной жидкостной хроматографии (ВЭЖХ) с чувствительностью до 50 нг / мл. При каждом посещении проводилась оценка безопасности, включая клиническое обследование, подробную историю нежелательных явлений, тесты функции печени, тесты функции почек и полный анализ крови. WA вводили ежедневно до прогрессирования. Общие критерии терминологии для нежелательных явлений (CTCAE) версии 3.0 использовались для классификации нежелательных явлений. Результаты: Используемый состав обычно хорошо переносился.Наблюдалось одиннадцать нежелательных явлений 1 или 2 степени тяжести. Побочных эффектов 3 или 4 степени не наблюдалось. Наиболее частыми побочными эффектами были повышение ферментов печени (5/11) и кожная сыпь (2/11). Другие побочные эффекты включают усталость, жар, отек и диарею (по одному). Ни у одного из пациентов не было обнаруживаемых уровней WA в кровотоке. Заключение: препарат хорошо переносился. Однако, по-видимому, WA имеет низкую пероральную биодоступность. Необходимы дальнейшие исследования с улучшенными составами.© 2019 Трансдисциплинарный университет, Бангалор и Всемирный фонд аюрведы. Издательские услуги Elsevier B.V. Это статья в открытом доступе под лицензией CC BY-NC-ND (http://creativecommons.org/licenses / by-nc-nd / 4.0 /).
Ключевые слова: Витхаферин-А Ашваганда Остеосаркома Фармакокинетика безопасности
1. Введение Существует растущая потребность в новых методах лечения рака, особенно на поздних стадиях. Большой интерес вызывают соединения природного происхождения как мишени для потенциального лечения рака.Многие ингредиенты, полученные из растений, в прошлом показали отличную противораковую активность. Противораковые препараты, такие как этопозид, винкристин и таксаны, и многие другие были получены из растений.
* Автор, ответственный за переписку. Отделение хирургии, Мемориальная больница Тата, Парел, Мумбаи, 400012, Индия. Факс: þ91 22 27405061. Адрес электронной почты: [электронная почта защищена] (А. Пури). Рецензирование проводится Трансдисциплинарным университетом, Бангалор.
Withania somnifera (Ашваганда) или индийский женьшень веками использовался в Аюрведе для лечения ряда заболеваний.Он обладает многочисленными фармакологическими свойствами, включая нейрозащиту, противовоспалительное действие, афродизиак, ревматизм, бессонницу и зоб среди других [1]. Экстракт корней содержит группу биологически активных компонентов, известных как витанолиды. Химическая структура витанолидов изучена, и они широко распространены в семействе Solanaceae. Витаферин-A (WA) является терапевтически активным витанолидом, который, как сообщается, присутствует в корнях и листьях растения. В нескольких доклинических исследованиях он показал значительную противораковую активность.Структура WA показана на рис. 1. Было показано, что WA предотвращает и излечивает уретановые опухоли легких у мышей, а также ингибирует
https://doi.org/10.1016/j.jaim.2018.12.008 0975 -9476 / © 2019 Трансдисциплинарный университет, Бангалор и Всемирный фонд аюрведы. Издательские услуги Elsevier B.V. Это статья в открытом доступе под лицензией CC BY-NC-ND (http://creativecommons.org/licenses/by-nc-nd/4.0/).
Цитируйте эту статью как: Пирес Н. и др., Безопасность и фармакокинетика витаферина-А при поздней стадии остеосаркомы высокой степени: испытание фазы I, J Ayurveda Integr Med, https: // doi.org / 10.1016 / j.jaim.2018.12.008
2
N. Pires et al. / Journal of Ayurveda and Integrative Medicine xxx (xxxx) xxx
Рост пересаженной саркомы у мышей. Корни ашваганды обладают активностью, способствующей укреплению здоровья, некоторые из действий экстракта корня и его компонентов включают ингибирование факторов транскрипции NF-kappaB и AP-1 [2], индукцию апоптоза через опосредованное митохондриями высвобождение цитохрома с и активацию каспазы [3] , выявление гуморальных и клеточно-опосредованных иммунных ответов за счет усиления Th2-доминантной поляризации [4] и противоопухолевых эффектов за счет подавления экспрессии p34cdc2 [5].Экстракт листьев ашваганды продемонстрировал противоопухолевую активность, которая в основном была связана с его компонентом витаноном, который избирательно активировал путь p53 в опухолевых клетках [6,7]. WA испытывали на солидных и асцитных опухолевых клетках саркомы 180 (S-180) мышей, которые лечили in vivo и in vitro. При наблюдении под электронным микроскопом было обнаружено, что соединение влияет на эти веретенообразные микротрубочки клеток в метафазе [8]. WA продемонстрировал коэффициент усиления сенсибилизатора 1,5 для in vitro уничтожения клеток китайского хомячка V79 при нетоксичной концентрации примерно 2 мМ [9].WA может ингибировать активацию и пролиферацию Т-и В-клеток, не вызывая гибели клеток. Таким образом, демонстрируется потенциал в качестве противовоспалительного агента [10]. WA демонстрирует цитотоксичность для различных линий опухолевых клеток. В своем исследовании они показали, что он в первую очередь вызывает окислительный стресс в клетках HL-60 лейкемии человека и в некоторых других линиях раковых клеток [11]. Генетические исследования показывают, что он может быть привлекательным вариантом лечения тройного отрицательного рака груди (TNBC) с плохим прогнозом [12].В доклинических данных, WA продемонстрировал антипролиферативный потенциал и привел к апоптозу в клеточных линиях остеосаркомы человека MG-63 и U2OS, индуцируя ROS-опосредованное снижение митохондриального мембранного потенциала и ингибируя белки контрольной точки G2 / M, тем самым способствуя остановке клеточного цикла [13,14 ]. Возможности лечения запущенной остеосаркомы ограничены. Метастатическое заболевание представляет собой проблему лечения, и у пациентов часто плохой прогноз. Тем самым указывается на необходимость новых методов лечения и вариантов лечения запущенной стадии болезни.Некоторые из проблем, которые необходимо преодолеть на клиническом уровне перед использованием WA в качестве противоопухолевого средства, — это его эффективность, эффективность и побочные эффекты, т.е. создание безопасного и эффективного протокола дозирования [15]. Несмотря на то, что он является сильным кандидатом, клинических испытаний этого продукта при раке не проводилось. Чтобы определить установленную пользу в клинических условиях, мы провели фазу I исследования у пациентов с остеосаркомой. Мы выбрали стандартизованный экстракт корней W. somnifera, содержащий 4,5% WA (вес / вес) для исследования фазы I.W. somnifera принимается с топленым маслом в Аюрведе, традиционной индийской медицине. Мы использовали формулу, в которой экстракт был обработан в соответствии с текстом Аюрведы в топленом масле. Наши цели в этом исследовании состояли в том, чтобы определить наличие какой-либо ограничивающей дозу токсичности (DLT)
Рис. 1. Структура витхаферина А.
на 30-й день и оценить безопасность и фармакокинетику препарата, содержащего WA, у пациентов с продвинутой стадией. остеосаркома высокой степени злокачественности. 2. Материалы и методы. Исследование было одобрено нашим комитетом по институциональной этике, который в настоящее время зарегистрирован Генеральным контролером по лекарственным средствам Индии (регистрационный номер IEC: ECR / 170 / Inst / MH / 2013).Исследование было зарегистрировано на сайте clinictrials.gov (идентификатор ClinicalTrials.gov: NCT00689195). 2.1. Пациенты и условия Это было одноцентровое исследование фазы I повышения дозы, проведенное в онкологической больнице третичного уровня в западной Индии. Мы набрали 13 пациентов, которые были готовы участвовать и соответствовали критериям отбора, включенные пациенты предоставили письменное информированное согласие на участие в исследовании. Критерии отбора для исследования включали: 1) Дети и взрослые в возрасте от 8 до 65 лет 2) Гистологически подтвержденная остеосаркома конечности высокой степени злокачественности и 3) Рецидив заболевания после первичного лечения, которые не подходят или отказываются от вторичной химиотерапии.Критерии исключения включали: 1) беременных / кормящих женщин, 2) женщин, которым требуется сопутствующее лечение индукторами или ингибиторами CYP, и 3) женщин без хорошего венозного доступа. 2.2. Вмешательство в исследование Для этого исследования использовался стандартизированный экстракт корня W. somnifera, содержащий 4,5% WA (вес / вес) (AshwaMAX 400, Pharmanza Herbal Pvt Ltd., Гуджарат, Индия). Каждая 400 мг капсула AshwaMAX содержала 18 мг WA. Экстракт был обработан в топленом масле, как предписано в традиционных текстах Аюрведы.2.3. Дизайн исследования Дизайн нашего исследования представлял собой классический подход к увеличению дозы в 3 × 3, как показано на рис. 2. Целью испытаний фазы 1 является определение максимально переносимой дозы (МПД). Конструкции 3 3 просты в эксплуатации и идентифицируют МПД с приемлемым уровнем точности. Основным ограничением дизайна 3 3 является то, что он включает значительную часть пациентов при субтерапевтических дозах [16]. Однако его консервативная стратегия повышения дозы сводит к минимуму риск воздействия токсичных доз исследуемого продукта.Ранее мы провели 28-дневное исследование токсичности повторных доз на крысах, в котором использовали стандартизированный экстракт W. somnifera, содержащий 4,5% WA. Уровень отсутствия наблюдаемых побочных эффектов (NOAEL), обнаруженный в исследовании, составил 2000 мг / кг. Соответствующая эквивалентная доза для человека (HED) составляет приблизительно 325 мг / кг / день. Для получения максимальной рекомендованной начальной дозы (MRSD) 65 мг / кг / день формулы для человека был применен коэффициент безопасности пять [17]. Учитывая, что средний вес человека составляет 60 кг, мы пришли к MRSD 3900 мг / день.В настоящем исследовании начальная доза используемого экстракта W. somnifera составляет 1600 мг, которую постепенно увеличивают до 4800 мг / день. Наивысшая доза WA, при которой у 1 из 6 пациентов наблюдалась ограничивающая дозу токсичность (DLT), была определена как максимальная переносимая доза (MTD). Ограничивающая дозу токсичность определялась как любое негематологическое событие 3 степени или гематологическое событие 4 степени, связанное с Витаферином-А, на основании Общих критериев терминологии для нежелательных явлений Национального института рака (CTCAE) версии 3.0. Когорты увеличения дозы включали 2 капсулы BID, 3 капсулы BID, 4 капсулы BID и 4 капсулы TID, соответствующие суточным дозам 72, 108, 144 и 216 мг WA.По три пациента были включены в каждую группу
. Процитируйте эту статью как: Pires N et al., Безопасность и фармакокинетика витхаферина-A при остеосаркоме высокой степени злокачественности: испытание фазы I, J Ayurveda Integr Med, https: // doi. org / 10.1016 / j.jaim.2018.12.008
N. Pires et al. / Журнал аюрведы и интегративной медицины xxx (xxxx) xxx
3
Таблица 1 Исходные характеристики исследуемой популяции. ALP e Щелочная фосфатаза. Характеристики пациентов Число пациентов Мужчины / женщины Средний возраст (лет) Средний гемоглобин (± SD) a (г / дл) Среднее значение Sr.Креатинин (± SD) a (мг / дл) Среднее значение Sr. ALPa (МЕ / л)
13 10/3 16 (13e43) 10,62 (± 2,53) 0,9 (± 0,14) 597,5 (± 849,5)
a Значения гемоглобина в г / дл, значения креатинина сыворотки в мг / дл и значения ЩФ в МЕ / л — это значения, измеренные на исходном уровне.
3.1. Результаты лабораторных исследований по безопасности У одного пациента в когорте 1 развилось обострение анемии с понижением уровня гемоглобина до 6,9 г / дл в течение 3 месяцев наблюдения по сравнению с 7,7 г / дл на исходном уровне. У пяти субъектов (38,46%) отмечалось повышение уровня печеночных ферментов — аспартатаминотрансферазы (АСТ) и аланинаминотрансферазы (АЛТ).Все повышения уровня ферментов печени были 1 степени тяжести. Только АСТ был повышен у одного пациента, только АЛТ — у одного пациента, в то время как у 3 пациентов было совместное повышение АЛТ и АСТ (таблица 2). 3.2. Неблагоприятные события
Рис. 2. 3 þ 3 Дизайн исследования с увеличением дозы для фазы I клинических испытаний. DLT ¼ Токсичность, ограничивающая дозу; MTD ¼ максимально переносимая доза. Все DLT оценивали на 30-е сутки.
Когортаи последний пациент наблюдались в течение не менее 30 дней на предмет токсичности, ограничивающей дозу, перед переходом в более высокую когорту.
Одиннадцать побочных эффектов произошли у 8 субъектов в ходе исследования. По два события произошли в когорте 1 и когорте 3, в то время как четыре события наблюдались в когорте 2 и три в когорте 4. Нежелательные явления делятся на 6 основных категорий: усталость (1), лихорадка (1), сыпь (2), диарея. (1), отек (1) и аномальные LFT (5). События были либо 1-й, либо 2-й степени. Неблагоприятные события, которые произошли во время исследования в 4 когортах, показаны выше в (Таблица 2).
2.4. Отбор проб и анализ безопасности и фармакокинетики
3.3. Фармакокинетические данные
Фармакокинетические образцы были собраны в день 1 в следующие моменты времени: е 0,0 (до введения дозы), 0,5, 1,0, 1,5, 2,0, 2,5, 3,0, 3,5, 4,0, 5,0, 6,0, 7,0, 8,0, 10, 12,0 и Через 24,0 часа после введения уровни в плазме измеряли с использованием утвержденной методики высокоэффективной жидкостной хроматографии (ВЭЖХ), как описано Patial et al. имеющий предел количественного определения 50 нг / мл [18]. В хроматографической системе использовалась колонка C18 с обращенной фазой с УФ-детектированием при 225 нм.Подвижная фаза состояла из воды и ацетонитрила, нанесенных градиентным потоком. Витаферин-A экстрагировали простым методом осаждения белков. Градуировочная кривая была линейной в диапазоне концентраций 0,05–1,6 мг / мл. Этот метод имеет желаемую чувствительность для определения диапазона концентраций витаферина-А в плазме, который может проявлять биологическую активность на основании данных in vitro. Пациенты наблюдались каждые 3 месяца. При каждом посещении проводилась оценка безопасности, включая клиническое обследование, подробный анамнез нежелательных явлений, функциональные тесты печени, функциональные тесты почек и общий анализ крови.WA вводили ежедневно до прогрессирования. CTCAE версии 3.0 использовался для классификации нежелательных явлений.
Все пациенты (N 13), включенные в исследование, предоставили образцы для фармакокинетического анализа WA. Ни в одном из отобранных образцов не было обнаружено обнаруживаемых уровней витаферина-А.
3. Результаты. Тринадцать пациентов, удовлетворяющих критериям отбора, были включены в исследование с увеличением дозы. Обобщены исходные характеристики участников исследования (Таблица 1). Блок-схема, изображающая процедуру включения и распределения пациентов, которой следовали в этом исследовании, а также количество пациентов, начинающих терапию при каждой дозе, показана на рис.3.
4. Обсуждение W. somnifera (WS) изучалась для лечения нескольких состояний. В одном исследовании субъекты со снижением когнитивных способностей были рандомизированы для приема либо WS 300 мг перорально два раза в день, либо плацебо в течение 8 недель. Исследование показало, что WS был эффективен в улучшении
зарегистрированных N = 13 последующих наблюдений N = 1 потерянных для наблюдения в когорте №. 3 до 30 дня, следовательно, заменено
Когорта 1 (2 капсулы два раза в день или 72 мг WA / день) (N = 3) Когорта 2 (3 капсулы BID или 108 мг WA / день) (N = 3) Когорта 3 (4 капсулы BID или 144 мг WA / день) (N = 4) Когорта 4 (4 капсулы 3 раза в день или 216 мг WA / день) (N = 3)
Оценка
N = 13 для PK N = 12 для безопасности
Рис.3. Блок-схема, отображающая регистрацию, распределение, наблюдение и оценку пациентов. WA Withaferin-A, PK ¼ Фармакокинетика.
Цитируйте эту статью как: Пирес Н. и др., Безопасность и фармакокинетика витаферина-А при поздней стадии остеосаркомы высокой степени: испытание фазы I, J Ayurveda Integr Med, https://doi.org/10.1016/j. jaim.2018.12.008
4
N. Pires et al. / Journal of Ayurveda and Integrative Medicine xxx (xxxx) xxx
Таблица 2 Показана частота нежелательных явлений в разных когортах.Неблагоприятное событие
72 мг (n 3)
108 мг (n 3)
144 мг (n 4)
216 мг (n 3)
Всего (n 13)
Количество неблагоприятные аномальные LFT Усталость Лихорадка Сыпь Диарея Отек DLT
события 1 1 eeeee
2 e 1 1 eee
eee 1 1 ee
2 eeee 1 e
5 1 1 2 1 1 e
AE- Неблагоприятное событие DLT — ограничивающая дозу токсичность, LFT — тесты функции печени.
немедленная и общая память у пациентов с умеренными когнитивными нарушениями, а также улучшение управляющих функций, внимания и скорости обработки информации [19].Другое исследование показало, что лечение экстрактом корня Ашваганды у взрослых с хроническим стрессом помогло контролировать массу тела и привело к значительному снижению массы тела, уровня кортизола в сыворотке и индекса массы тела (ИМТ) [20]. WA, компонент Ашваганды, был изучен на предмет его противораковой активности на многих доклинических моделях in vitro, а также на доклинических моделях in vivo. Однако на сегодняшний день клинических исследований WA при раке не проводилось. При запущенной остеосаркоме варианты лечения ограничены, а фитохимические препараты продемонстрировали полезную противоопухолевую активность в прошлом.Фитохимический WA — многообещающий противораковый агент. На молекулярном уровне было показано, что он нацелен на множество киназных путей выживания клеток, включая NF-kB, киназу PI3, протеинкиназу, протеинкиназу, активированную митогеном, среди других. Сообщалось, что WA индуцирует апоптоз через внутренние и внешние пути в человеческих раковых клетках простаты, груди и лейкемии среди других. Он также продемонстрировал свою способность вносить свой вклад в противоопухолевый эффект как in vitro, так и in vivo [21]. Доклинические исследования с использованием ортотропных моделей на мышах продемонстрировали, что пероральное введение витаферина-A эффективно против мультиформной глиобластомы (GBM), агрессивного злокачественного новообразования [22].Он также показал способность индуцировать апоптоз в клеточных линиях остеосаркомы U2OS путем генерации ROS, что также вызывает остановку клеточного цикла в клеточных линиях остеосаркомы за счет ингибирования белков контрольных точек G2 / M [13,14]. В этом исследовании нашей целью было оценить безопасность и фармакокинетику WA на запущенной стадии остеосаркомы высокой степени. Результаты показали, что препарат хорошо переносился пациентами до дозы 4800 мг, что эквивалентно 216 мг WA в день, без какой-либо токсичности, ограничивающей дозу.Это была самая высокая доза, которую можно было вводить при разумной переносимости и соблюдении пациентом режима лечения, поскольку пациенты в группе с самой высокой дозой (216 мг) получали 4 капсулы WA 3 раза в сутки. Никаких DLT не наблюдалось даже при самой высокой дозе, что указывает на то, что пациентам можно попробовать более высокие дозы, но общая доза ограничена количеством капсул, которые пациент может разумно потреблять в день. Стандартизованный экстракт с более высоким содержанием WA может обойти эту проблему, что приведет к введению более высоких доз в день при приемлемом количестве капсул и частоте приема.Общие наблюдаемые побочные эффекты включают повышение уровня ферментов печени (степень 1) у 5 из 11 пациентов и кожную сыпь у 2 из 11 пациентов. Другие наблюдаемые побочные эффекты включают жар, усталость, отек и диарею (по одному событию). Побочных эффектов 3 или 4 степени не наблюдалось. WA не был обнаружен ни в одной из исследуемых выборок. Используемый метод представлял собой анализ ВЭЖХ с чувствительностью 50 нг / мл. На момент исследования была доступна только система ВЭЖХ, способная определять 50 нг / мл. Биодоступность витанолидов зависит от степени их абсорбции и транспортировки через эпителий кишечника.Исследование, проведенное для оценки биодоступности витанолидов с использованием модели абсорбции in vitro, то есть системы почек Mandin Darby Canine, обнаружило, что WA непроницаема или метаболизируется при прохождении через клеточный слой [23]. Однако другое исследование, в котором пытались проанализировать PK WA и витанолида A (WEA) в плазме мышей после перорального приема, показало, что пероральное всасывание витанолидов было быстрым. Он также продемонстрировал, что относительная биодоступность WA была в полтора раза больше, чем WEA.В исследовании использовался анализ HPLC-ESI-MS / MS, чувствительность которого составила 0,484 нг / мл [24], что позволяет предположить, что для характеристики PK WA у людей требуются более чувствительные аналитические методы. Мы не смогли установить биодоступность WA по указанным выше причинам. IC50 WA находится на субмикромолярном уровне, что соответствует концентрации приблизительно 470 нг / мл. Однако вряд ли WA будет использоваться как отдельный агент. Лучше всего использовать WA в сочетании со стандартной химиотерапией.Синергетические эффекты витаферина А были продемонстрированы в исследовании, в котором WA в сочетании с цисплатином действовал синергетически и, как было обнаружено, усиливал противоопухолевые эффекты на клеточные линии яичников, подавляя пролиферацию клеток, генерацию ROS и вызывая повреждение ДНК, ведущее к апоптозу [25]. . Другое исследование показывает потенциал WA в качестве радиосенсибилизатора. Исследование проводилось на опухоли меланомы мыши, которую предварительно лечили WA с последующим местным гамма-излучением. WA вызывает дозозависимое увеличение задержки роста и времени удвоения объема [26].Фактор транскрипции NFkB, который регулирует воспаление, выживаемость клеток и экспрессию P-гликопротеина, играет важную роль в индукции устойчивости к противораковым препаратам [27]. Наблюдается, что WA ингибирует NFkB в резистентных к доксорубицину клетках K562 / Adr и может ослаблять активацию ослабленных каспаз и вызывать апоптоз [27]. Это делает WA привлекательным соединением для хемосенсибилизации и для индукции гибели клеток в химиорезистентных типах клеток. Таким образом, WA может взаимодействовать с другими противоопухолевыми средствами, а его активность, модулирующая Р-гликопротеин, может быть использована для улучшения пероральной биодоступности и внутриопухолевых концентраций совместно вводимых противоопухолевых препаратов [27].Необходимы дальнейшие клинические исследования WA в качестве дополнения к химиотерапии при остеосаркоме. Есть также предварительные данные, касающиеся активности WA при раке желудка, щитовидной железы и яичников. Здесь снова можно оценить полезность WA в качестве дополнительного лечения к стандартным модальностям. Другие преимущества WA, включая иммуномодулирующие эффекты, предполагают, что он может смягчать / защищать от токсичности и побочных эффектов химиотерапии. Будучи экономичным, а также природным фитохимическим веществом, разработка улучшенной рецептуры витхаферина-А может оказаться привлекательным вариантом противоопухолевого лечения как в развитых, так и в развивающихся странах.4.1. Ограничение Одним из ограничений этого исследования является то, что максимальная переносимая доза (МПД) не была достигнута. Максимальная доза, использованная в этом исследовании, составляла 216 мг, что, вероятно, является заниженной оценкой возможной MTD. Могут быть введены более высокие дозы. WA имеет улучшенный состав более высокой силы, который позволяет упростить режим дозирования и уменьшить частоту введения. Биоаналитический метод, использованный в этом исследовании, имел чувствительность 50 нг / мл. Чувствительность и конкретность аналитического метода были ограничением нашего исследования.В последующих исследованиях рекомендуются биоаналитические методы с более высокой чувствительностью. 5. Заключение Наше исследование фазы I показало, что пероральный витаферин-А у пациентов с запущенной стадией остеосаркомы высокой степени имеет хороший профиль безопасности.
Цитируйте эту статью как: Пирес Н. и др., Безопасность и фармакокинетика витаферина-А при поздней стадии остеосаркомы высокой степени: испытание фазы I, J Ayurveda Integr Med, https://doi.org/10.1016/j. jaim.2018.12.008
N. Pires et al. / Журнал аюрведы и интегративной медицины xxx (xxxx) xxx
Дальнейшие исследования фазы II WA можно проводить в дозе 216 мг / день.
[11]
Конфликт интересов Нет. Финансирование Исследование финансировалось через внутренний грант Мемориального центра Тата, Индия (номер гранта: 381/07).
[12]
[13]
[14]
Благодарности [15]
Авторы выражают признательность за финансовую поддержку внутреннему гранту Мемориального центра Тата за проведение этого исследования. Ссылки [1] Дар Н.Дж., Хамид А., Ахмад М. Фармакологический обзор Withania somnifera, индийского женьшеня. Cell Mol Life Sci 2015; 72: 4445e60.https://doi.org/ 10.1007 / s00018-015-2012-1. [2] Сингх Д., Аггарвал А., Маурья Р., Наик С. Withania somnifera ингибирует факторы транскрипции NF-kB и AP-1 в периферической крови человека и мононуклеарных клетках синовиальной жидкости. Phyther Res 2007; 21: 905e13. https://doi.org/10.1002/ ptr.2180. [3] Сентил В., Рамадеви С., Венкатакришнан В., Гиридхаран П., Лакшми Б.С., Вишвакарма Р.А. и др. Витанолид вызывает апоптоз в лейкозных клетках HL-60 через митохондриально-опосредованное высвобождение цитохрома с и активацию каспазы. Chem Biol Interact 2007; 167: 19e30.https://doi.org/10.1016/ j.cbi.2007.01.004. [4] Малик Ф., Сингх Дж., Кхаджурия А., Сури К.А., Сатти Н.К., Сингх С. и др. Стандартизированный экстракт корня Withania somnifera и его основной компонент витанолид-A вызывают гуморальные и клеточно-опосредованные иммунные ответы за счет усиления регуляции Th2-доминантной поляризации у мышей BALB / c. Life Sci 2007; 80: 1525e38. https: // doi.org/10.1016/j.lfs.2007.01.029. [5] Сингх Д. Подавление экспрессии p34cdc2 водной фракцией из-за возможного молекулярного механизма противоопухолевого и других фармакологических эффектов.Фитомедицина 2001; 8: 492e4. https://doi.org/10.1078/S09447113(04)70072-0. [6] Каур К., Рани Дж., Видодо Н., Нагпал А., Тайра К., Каул С. К. и др. Оценка антипролиферативной и антиоксидантной активности экстракта листьев in vivo и in vitro, выращенных ашвагандхой. Food Chem Toxicol 2004; 42: 2015e20. https: // doi.org/10.1016/j.fct.2004.07.015. [7] Видодо Н., Каур К., Шреста Б.Г., Такаги Й., Исии Т., Вадхва Р. и др. Селективное уничтожение раковых клеток экстрактом листьев ашваганды: идентификация фактора, ингибирующего опухоль, и первые молекулярные исследования его эффекта.Clin Cancer Res 2007; 13: 2298e306. https://doi.org/10.1158/1078-0432.CCR-060948. [8] Шохат Б., Шалтиэль А., Бен-Бассат М., Джошуа Х. Влияние витаферина А, природного стероидного лактона, на тонкую структуру опухолевых клеток S-180. Cancer Lett 1976; 2: 71e7. https://doi.org/10.1016/S0304-3835(76)80014-6. [9] Деви ПУ. Withania somnifera Dunal (Ашваганда): потенциальный растительный источник многообещающего лекарства для химиотерапии рака и радиосенсибилизации. Индийский журнал J Exp Biol 1996; 34: 927e32. [10] Гамбхир Л., Чекер Р., Шарма Д., Тхох М., Патил А., Дегани М. и др.Тиол-зависимое подавление NF-kB и ингибирование адаптивных иммунных ответов, опосредованных Т-клетками, с помощью природного стероидного лактона витхаферина А.
[16]
[17]
[18]
[19]
[20]
[21]
[22]
[23]
[24]
[25]
[26] [27]
5
Toxicol Appl Pharmacol 2015; 289: 297e312. https://doi.org/10.1016/ j.taap.2015.09.014. Малик Ф., Кумар А., Бхушан С., Хан С., Бхатиа А., Сури К.А. и др.Генерация активных форм кислорода и митохондриальная дисфункция при апоптотической гибели клеток HL-60 миелоидной лейкемии человека диетическим соединением с гаферином А с сопутствующей защитой N-ацетилцистеином. Апоптоз 2007; 12: 2115e33. https://doi.org/10.1007/s10495-007-0129-x. Сарк Вел Шич К., Деклерк К., Кранс РАДЖ, Дидденс Дж., Шерф Д. Б., Герхаузер К. и др. Эпигенетическое подавление тройных отрицательных признаков рака молочной железы, Витхаферин А. Oncotarget, 2017; 8: 40434e53. https://doi.org/10.18632 / oncotarget.17107. Lv T-Z, Wang G-S. Потенциал антипролиферации витаферина А на клетках остеосаркомы человека посредством ингибирования белков контрольной точки G2 / M. Exp Ther Med 2015; 10: 323e9. https://doi.org/10.3892/etm.2015.2480. Li A-X, Sun M, Li X. Витаферин-A индуцирует апоптоз в клеточной линии остеосаркомы U2OS посредством генерации ROS и нарушения потенциала митохондриальной мембраны. Eur Rev Med Pharmacol Sci 2017; 21: 1368e74. http: //www.ncbi.nlm. nih.gov/pubmed/28387888. по состоянию на 11 июля 2017 г. Palliyaguru DL, Singh SV, Kensler TW.Withania somnifera: от профилактики к лечению рака. Mol Nutr Food Res 2016; 60: 1342e53. https://doi.org/ 10.1002 / mnfr.201500756. Хансен А., Грэм Д., Понд Дж., Сиу Л. Дизайн испытания фазы I: является ли 3þ3 лучшим? Cancer Contr 2014; 21: 200e8. https://doi.org/10.1177/107327481402 100304. Патель С., Рао Н., Хингорани Л. Оценка безопасности экстракта Withania somnifera, стандартизированного для Withaferin A: исследование острой и подострой токсичности. J Ayurveda Integr Med 2016: 30e7. https://doi.org/10.1016/j.jaim.2015.08.001. Патиал П., Гота В. Быстрый и чувствительный метод определения витаферина-А в плазме человека с помощью ВЭЖХ. Биоанализ 2011; 3: 285e9. https://doi.org/ 10.4155 / bio.10.207. Чоудхари Д., Бхаттачарья С., Бозе С. Эффективность и безопасность экстракта корня ашваганды (Withania somnifera (L.) Dunal) для улучшения памяти и когнитивных функций. J Diet Suppl 2017; 14: 599e612. https://doi.org/10.1080/ 193.2017.1284970. Чоудхари Д., Бхаттачарья С., Джоши К. Управление массой тела у взрослых в условиях хронического стресса с помощью лечения экстрактом корня Ашваганды.Дж. Эвид на основе дополнительной альтернативной медицины 2017; 22: 96e106. https://doi.org/ 10.1177/2156587216641830. rez-Novo C, Van Ostade X, Vanden Berghe W. Molecular Chirumamilla CS, Pe анализ терапевтических эффектов диетического лекарственного фитохимического вещества Withaferin A. Proc Nutr Soc 2017; 76: 96e105. https://doi.org/10.1017/ S0029665116002937. Чанг Э., Полинг С., Натараджан А., Уитни Т.Х., Каур Дж., Сюй Л. и др. AshwaMAX и Withaferin A подавляют глиомы на клеточных и мышиных ортотопических моделях. Журнал Neurooncol 2016; 126: 253e64.https://doi.org/10.1007/s11060-015-1972-1. Devkar S, Kandhare A, Sloley B, Jagtap S, Lin J, Tam Y и др. Оценка биодоступности основных витанолидов Withania somnifera с использованием модельной системы абсорбции in vitro. J Adv Pharm Educ Res 2015; 6: 159e64. https: // doi.org/10.4103/2231-4040.165023. Патил Д., Гаутам М., Мишра С., Карупотула С., Гайрола С., Джадхав С. и др. Определение витаферина А и витанолида А в плазме мышей с использованием высокоэффективной жидкостной хроматографии и тандемной масс-спектрометрии: применение к фармакокинетике после перорального введения водного экстракта Withania somnifera.Дж. Фармацевтика Биомед Анал 2013; 80: 203e12. https://doi.org/ 10.1016 / j.jpba.2013.03.001. Какар СС, Джала В.Р., Фонг М.Ю. Синергетическое цитотоксическое действие цисплатина и витаферина А на клеточные линии рака яичников. Biochem Biophys Res Commun 2012; 423: 819e25. https://doi.org/10.1016/j.bbrc.2012.06.047. Деви ПУ, Камат Р., Рао Б.С. Радиосенсибилизация меланомы мыши витаферином А: исследования in vivo. Индийский журнал J Exp Biol 2000; 38: 432e7. Суттана В., Манхеткорн С., Поопимон В., Палагани А., Жохов С., Герло С. и др.Дифференциальная хемосенсибилизация Р-гликопротеина, сверхэкспрессирующего клетки K562 / Adr, с помощью витаферина А и полифенолов сиамоза.