Опубликовать Опубликовать
Опубликовать ОпубликоватьРеферат на тему:
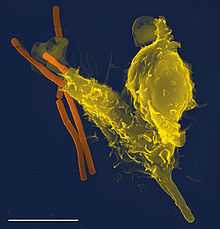
Микрофотография нейтрофила (из электронного микроскопа), фагоцитирующего Bacillus anthracis (оранжевая).
Фагоци́ты — клетки иммунной системы, которые защищают организм путём поглощения (фагоцитоза) вредных чужеродных частиц, бактерий, а также мёртвых или погибающих клеток[1]. Их название произошло от греческого phagein, «есть» или «поедать», и «-cyte», суффикс, в биологии означающий «клетка»[2]. Они важны для борьбы с инфекцией и постинфекционного иммунитета[3]. Фагоцитоз важен для всего животного мира[4] и высоко развит у позвоночных[5]. Фагоциты и фагоцитоз как способ пищеварения у животных были открыты И.И. Мечниковым при изучении губок и плоских червей. Роль фагоцитов в защите от бактерий была впервые открыта И.И.Мечниковым в 1882 году, когда он изучал личинок морских звёзд[6]. Мечников был удостоен в 1908 году Нобелевской премии по физиологии за создание клеточной теории иммунитета[7]. Фагоциты присутствуют в организмах многих видов; некоторые амёбы по многим деталям поведения похожи на макрофаги, что указывает на то, что фагоциты появились на ранних этапах эволюции.[8]
Фагоциты человека и других животных называют «профессиональными» или «непрофессиональными» в зависимости от того, насколько эффективно они фагоцитируют[9]. К профессиональным фагоцитам относятся нейтрофилы, моноциты, макрофагии, дендритные клетки и тучные клетки.[10] Основное отличие профессиональных фагоцитов от непрофессиональных в том, что профессиональные имеют молекулы, называемые рецепторы, на своей поверхности, которые обнаруживают чужеродные объекты, например бактерии.[11] Один литр крови взрослого человека в норме содержит около 2,5—7,5 млрд нейтрофилов, 200—900 млн моноцитов[12].
При инфекции химические сигналы привлекают фагоциты к месту, где патоген проник в организм. Эти сигналы могут исходить от бактерий или от других фагоцитов, уже присутствующих там. Фагоциты перемещаются путём хемотаксиса. Когда фагоциты контактируют с бактериями, рецепторы на их поверхности связываются с ними. Эта связь приводит к поглощению бактерий фагоцитами.[13] Некоторые фагоциты убивают проникших патогенов с помощью оксидантов и оксида азота.[14] После фагоцитоза, макрофаги и дендритные клетки могут также участвовать в презентации антигена — процессе, при котором фагоциты перемещают патогенный материал обратно на свою поверхность. Этот материал затем отображается (презентируется) для других клеток иммунной системы. Некоторые фагоциты поступают в лимфатические узлы и презентируют материал лимфоцитам. Этот процесс важен в формировании иммунитета.[15] Тем не менее, многие болезнетворные микроорганизмы устойчивы к атакам фагоцитов.[3]
Три стадии фагоцитоза:1. Несвязанные рецепторы на поверхности фагоцита не запускают фагоцитоз.2. Связывание рецепторов вызывает их группирование.3. Запускается фагоцитоз и частица поглощается фагоцитом.
Фагоцитоз — процесс поглощения чужеродных частиц клетками.[16] Он включает последовательность молекулярных процессов.[17] Фагоцитоз возникает после связывания рецепторами чужеродного агента (например, бактерии). Затем фагоцит окружает бактерию и поглощает её. Фагоцитоз бактерии человеческим нейтрофилом происходит примерно за 9 минут.[18] Внутри фагоцита бактерия оказывается в составе фагосомы. В течение минуты фагосома сливается с лизосомой или гранулой, содержащими ферменты, с образованием фаголизосомы. Заключённая бактерия подвергается агрессивному воздействию[19] и погибает через несколько минут.[18] Дендритные клетки и макрофаги действуют не так быстро и фагоцитоз в этих клетках может протекать в течение многих часов. Макрофаги поглощают большое количество чужеродного материала и часто выделяют некоторые непереваренные частицы обратно. Этот материал является сигналом для миграции макрофагов из крови.[20] Фагоциты способны поглощать почти любое вещество.
Макрофаги имеют специальные рецепторы, которые способствуют фагоцитозу
Фагоциты имеют большое множество типов рецепторов на своей поверхности, с помощью которых они связывают чужеродный материал.[3] К ним относятся опсониновые рецепторы, скевенджер-рецепторы и Толл-подобные рецепторы. Опсониновые рецепторы усиливают фагоцитоз бактерий которые покрыты иммуноглобулином G (IgG) или комплементом. «Комплемент» — комплекс белковых молекул в крови, которые разрушают клетки или помечают их для уничтожения.[21] Скевенджер-рецепторы крупными молекулами поверхности бактериальной клетки и Толл-подобные рецепторы связываются с более специфичными молекулами. Связывание Толл-подобных рецепторов усиливает фагоцитоз и вызывают выброс фагоцитами группы гормонов, которые вызывают воспаление.[3]
Упрощённая схема фагоцитоза и разрушения бактериальной клетки
Уничтожение микроорганизмов — важная функция фагоцитоза,[22] которая происходит либо при фагоцитозе (внутриклеточное уничтожение), либо вне фагоцита (внеклеточное уничтожение).
Когда фагоцит поглощает бактерию (или любой другой чужеродный материал), увеличивается потребление кислорода, что называют респираторным взрывом. При этом образуются реактивные кислород-содержащие молекулы, которые обладают противомикробным действием.[23] Соединения кислорода токсичны как для патогена, так и для самой клетки, поэтому они хранятся в ячейках внутри самой клетки. Такой метод уничтожения проникающих микроорганизмов называют кислород-зависимое внутриклеточное уничтожение, который делится на 2 типа.[14]
Первый тип — кислород-зависимое образование супероксидного радикала[3], уничтожающего бактерии.[24] Супероксид превращается в пероксид водорода и синглетный кислород под действием фермента супероксиддисмутаза. Супероксиды также взаимодействуют с пероксидом водорода с образованием гидроксильной группы, которая помогает в уничтожении патогенных микробов.[3]
Ко второму типу относят использование фермента миелопероксидаза из нейтрофильных гранул.[25] Когда гранулы сливаются с фагосомой, миелопероксидаза освобождается в фаголизосому, и этот фермент использует пероксид водорода и хлор для создания гипохлорита. Гипохлорит крайне токсичен для бактерий.[3] Миелопероксидаза содержит пигмент гем, за счёт которого образуется зелёный цвет секретов, богатых нейтрофилами (например гной, инфицированная мокрота).[26]
Микропрепарат гноя. Видны бактерии Neisseria gonorrhoea, окрашенные по Граму, внутри фагоцитов
Фагоциты также могут уничтожать микроорганизмы кислород-независимым методом, но он менее эффективен, чем кислород-зависимый. Различают 4 основных типа. При первом типе используются электрически заряженные белки, которые повреждают клеточную мембрану бактерий. При втором типе используются лизозимы; эти ферменты разрушают клеточную стенку бактерий. При третьем типе используются лактоферрины, которые присутствуют в гранулах нейтрофилов и удаляют необходимое железо из бактерий.[27] При четвёртом типе используются протеазы и гидролазы для переваривания белков разрушенных бактерий.[28]
Интерферон-гамма (который также называют фактор, активирующий макрофаги) активирует синтез макрофагами оксида азота. Источником интерферона-гамма могут быть CD4+ T лимфоциты, CD8+ T лимфоциты, естественные киллеры, B-лимфоциты, T-киллеры, моноциты, макрофаги или дендритные клетки.[29] Оксид азота затем высвобождается из макрофагов и, в силу своей токсичности, уничтожает микробы вблизи макрофага.[3] Активированные макрофаги образуют и секретируют фактор некроза опухоли. Этот цитокин (класс сигнальной молекулы)[30] уничтожает раковые клетки и клетки, инфицированные вирусом, помогают активировать другие клетки иммунной системы.[31]
При некоторых заболеваниях, например, при редких хронических гранулёматозных заболеваниях, эффективность фагоцитоза нарушается, что может привести к возникновению бактериальных инфекций.[32] При таких заболеваниях существует аномалия в работе различных элементов кислород-зависимого уничтожения микробов. Другие редкие врождённые аномалии, например синдром Шедьяка–Штайнбринка–Хигаси, также связаны с дефектным уничтожением проникающих в организм микробов.[33]
Вирусы могут воспроизводится только внутри клетки, и они проникают в неё используя множество рецепторов, участвующих в иммунной защите. Попав внутрь клетки, вирусы используют её биологические процессы в свою пользу, заставляя клетку создавать тысячи вирусных частиц, подобных материнской. Хотя фагоциты и другие компоненты иммунной системы могут в ограниченной степени контролировать вирусы, когда вирус внутри клетки, приобретённый иммунитет (в частности лимфоциты) более важен для защиты.[34] В области вирусной инфекции лимфоцитов скапливается гораздо больше, чем остальных клеток иммунной системы; что наиболее типично для вирусного менингита.[35]Клетки, инфицированные вирусами, уничтожаются лимфоцитами и выводятся из организма фагоцитами.[36]
Апоптоз—фагоциты очищают организм от остатков мёртвых клеток
У животных, растений и грибов клетки постоянно погибают. Баланс между делением клетки и их гибелью сохраняет относительно постоянное число клеток у взрослых.[1] Существует 2 механизма гибели клетки: некроз или апоптоз. В отличии от некроза, который чаще возникает как результат заболевания или травмы, апоптоз (или запрограммированная гибель клеток) — нормальный процесс, протекающий в организме. Организм избавляет себя от миллионов мёртвых или погибающих клеток каждый день и фагоциты играют важную роль в этом процессе.[37]
Погибающая клетка, которая подвергается финальной стадии апоптоза,[38] обнажает молекулы, например фосфатидилсерин, на своей поверхности для соединения с фагоцитом.[39] Фосфатидилсерин обычно находится на цитозольной поверхности плазматической мембраны, но перемещается при апоптозе на внешнюю поверхность предположительно с помощью белка под названием скрамблаза.[40] Эти молекулы помечают клетку для фагоцитоза клетками, которые обладают соответствующими рецепторами, например макрофаги.[41] Удаление погибающих клеток фагоцитами происходит упорядоченным образом, не вызывая воспаления.[42]
Фагоциты движутся в организме взаимодействуя с фагоцитарными и нефагоцитарными клетками иммунной системы. Они обмениваются информацией с другими клетками с помощью образования химических веществ, называемых цитокинами, которые вызывают другие фагоциты к области инфекции или активируют «спящие» лимфоциты.[43] Фагоциты составляют часть врождённого иммунитета, который появляется у животных, включая человека, при рождении. Врождённый иммунитет очень эффективен, но не специфичен в отношении определения разницы между видами патогенов. С другой стороны, приобретённый иммунитет более специализированный и может защищать почти от любого вида патогена.[44] Приобретённый иммунитет зависит от лимфоцитов, которые не фагоцитируют, но образуют защитные белки (антитела), которые помечают патогены для уничтожения и предупреждают инфицирование клеток вирусами.[45] Фагоциты, в частности дендритные клетки и макрофаги, стимулируют лимфоциты для образования антител при важном процессе, называемом презентация антигена.[46]
Схема презентации чужеродных пептидов MHC 1 молекулами
Презентация антигена — процесс, при котором некоторые фагоциты перемещают части поглощённого материала назад на свою поверхность и «предоставляют» их для других клеток иммунной системы.[47] Существуют 2 вида «профессиональных» антиген-презентирующих клеток: макрофаги и дендритные клетки.[48] После поглощения, чужеродные белки (антигены) разрушаются до пептидов внутри дендритной клетки или макрофага. Эти пептиды затем связываются с гликопротеинами главного комплекса гистосовместимости (MHC) клетки, которые осуществляют возвращение назад на поверхность фагоциты, где они могут быть «представлены» лимфоцитам.[15] Старые макрофаги не способны быстро перемещаться из области инфицирования, но дендритные клетки могут достигать лимфатических узлов организма, где находятся миллионы лимфоцитов.[49] Это улучшает иммунитет, потому что лимфоциты реагируют на антигены, презентированные дендритными клетками, также, как если бы они находились в первичной области инфекции.[50] Но дендритные клетки также способны разрушать или подавлять активность лимфоцитов, если они распознают компоненты тела хозяина; это важно для предупреждения аутоиммунных реакций. Этот процесс называют толерантностью.[51]
Дендритные клетки также способствуют иммунологической толерантности,[52], при которой предупреждается иммунологическая атака организма самого себя. Первый тип толерантности — центральная толерантность: когда созревшие T-лимфоциты впервые выходят из тимуса, дендритные клетки разрушают дефектные Т-лимфоциты, которые несут антигены, что может вызывать аутоиммунную реакцию. Второй тип иммунологической толерантности — периферическая толерантность. Некоторые Т-лимфоциты, которые обладают антигенным свойством, способны пройти первый тип толерантности, некоторые Т-лимфоциты развивают аутоиммунные антигены позднее в жизни, и некоторые аутоиммунные антигены не обнаруживаются в тимусе; поэтому дендритные клетки будут работать, опять же, для сдерживания активности аутоиммунных Т-лимфоцитов вне тимуса. Дендритные клетки могут сделать это путём разрушения их или с помощью привлечения Т-супрессоров для дезактивации вредоносных Т-лимфоцитов.[53] Когда иммунологическая толерантность не срабатывает, могут возникнуть аутоиммунные заболевания.[54] С другой стороны, повышенная толерантность может привести к возникновению инфекций, например ВИЧ-инфекции.[53]
Фагоциты образуются из стволовых клеток костного мозга
Фагоциты человека и других позвоночных разделяют на «профессиональные» и «непрофессиональные» группы на основе эффективности, при которой они участвуют фагоцитозе.[9] К профессиональным фагоцитам относят моноциты, макрофаги, нейтрофилы, тканевые дендритические клетки и тучные клетки.[10]
Все фагоциты, особенно макрофаги, находятся в состоянии готовности. Макрофаги, как правило, относительно пассивны в тканях и размножаются медленно. В таком состоянии полу-покоя они очищают организм от мёртвых клеток и другого неинфекционного мусора и редко принимают участие в презентации антигена. Но при возникновении инфекции они получают химические сигналы (обычно интерферон гамма), которые увеличивают продукцию ими MHC II молекул и подготавливают их к презентации антигена. В таком состоянии, макрофаги — хорошие антиген-презентаторы и киллеры. Однако, если они получают сигнал прямо от патогена, они становятся «гиперактивными», прекращают размножение и концентрируются на уничтожении. Их размер и скорость фагоцитоза увеличивается; некоторые становятся достаточно крупными, чтобы поглотить проникающих в организм простейших.[55]
В крови нейтрофилы неактивны, но движутся по ней с большой скоростью. Когда они получают сигналы от макрофагов из зоны воспаления, они замедляются и выходят из крови. В тканях они активируются цитокинами и поступают в зону действия готовыми уничтожать.[56]
Нейтрофилы выходят из крови в зону инфекции
Когда происходит инфекционное заражение, химический «SOS» сигнал выделяется для привлечения фагоцитов в зону инфекции.[57] Эти химические сигналы могут включать белки от поступающих бактерий, системы свёртывания пептидов, продукты системы комплемента, а также цитокины, которые выделяются макрофагами, расположенными в ткани в области инфекции.[3] Другая группа химических аттрактантов — цитокины, которые вызывают нейтрофилы и моноциты из кровеносного русла.[13]
Для достижения зоны инфекции, фагоциты выходят из кровеносного русла и проникают в поражённую ткань. Сигналы от инфекции вызывают синтез эндотелиальными клетками, выстилающие кровеносный сосуд, белка, называемого селектин, который сцепляется с проходящими нейтрофилами. Вазодилататоры ослабляют соединительные связи эндотелиальных клеток, что позволяет фагоцитами проходить через стенку сосуда. Хемотаксис — процесс, при котором фагоциты следуют на «запах» цитокинов к области инфекции.[3] Нейтрофилы проникают через органы, покрытые эпителиальной тканью, в зону инфекции, и хотя это важный компонент борьбы с инфекцией, миграция сама по себе может привести к возникновению симптомов заболевания.[58] При инфекции миллионы нейтрофилов вызываются из крови, но они погибают затем в течение нескольких дней.[59]
Моноциты с дольчатым ядром в окружении красных кровяных клеток (малое увеличение)
Моноциты развиваются в костном мозге и достигают зрелости в крови. Зрелые моноциты имеют крупное, гладкое, дольчатое ядро и цитоплазму, которая содержит гранулы. Моноциты поглощают чужеродные или опасные вещества и презентируют антигены другим клеткам иммунной системы. Моноциты образуют 2 группы: циркулирующая и краевая, которые остаются в других тканях (около 70 % находятся в краевой группе). Большинство моноцитов покидают кровеносное русло через 20-40 часов, попадая в ткани и органы, где они превращаются в макрофаги[60] или дендритические клетки в зависимости от получаемого сигнала.[61] В 1 литре крови человека находится около 500 миллионов моноцитов.[12]
Зрелые макрофаги не перемещаются быстро, но стоят на страже в тех областях организма, которые подвержены воздействию внешней среды. Там они действуют как сборщики мусора, антиген представляющие клетки или агрессивные киллеры в зависимости от получаемого сигнала.[62] Они образуются из моноцитов, гранулоцитов стволовых клеток или при клеточном делении уже существующих макрофагов.[63] Макрофаги человека диаметром около 21 микрометра.[64]
Гной выделяется из абсцесса, вызванного бактериями (гной содержит миллионы макрофагов)
Этот вид фагоцитов не имеет гранул, но содержит много лизосом. Макрофаги находятся по всему телу почти во всех тканях и органах (например, клетки микроглии в головном мозге и альвеолярные макрофаги в лёгких). Расположение макрофага можно определить по его размеру и внешнему виду. Макрофаги вызывают воспаление путём образования интерлейкина 1, интерлейкина 6 и фактора некроза опухоли.[65] Макрофаги обычно находятся только в тканях и редко попадают в кровоток. Продолжительность жизни тканевых макрофагов, по разным оценкам, от 4 до 5 дней.[66]
Макрофаги могут быть активированы для выполнения функций, которые покоящийся моноцит не может.[65] Т-хелперы подгруппа лимфоцитов, отвечающих за активацию макрофагов. Они активируют макрофаги, посылая сигнал в виде интерферона гамма и экспрессируя белок CD154.[67] Другие сигналы поступают от бактерий в виде фактора некроза опухоли альфа и липополисахаридов.[65] Т-хелперы способны привлекать другие фагоциты в зону инфекции несколькими путями. Они выделяют цитокины, которые действуют на костный мозг, стимулируя образование моноцитов и нейтрофилов и они выделяют некоторые цитокины, которые отвечают за миграцию моноцитов и нейтрофилов в кровеносное русло.[68] Т-хелперы появляются при дифференцировке CD4+ Т лимфоцитов, когда они реагируют на действие антигена в периферических лимфатических тканях.[65] Активированные макрофаги играют важную роль в разрушении опухолей путём образования фактора некроза опухоли альфа, гамма-интерферона, оксида азота, реактивных форм кислорода, катионных белков и гидролитических ферментов.[65]
Сегментоядерный нейтрофил (в центре) в окружении эритроцитов, в цитоплазме видны внутриклеточные гранулы (микрофотография, окраска по Романовскому — Гимзе)
Нейтрофилы обычно находятся в кровеносном русле и являются наиболее распространённым типом фагоцитов, составляя 50-60 % от всех циркулирующих в крови белых кровяных клеток.[69] Один литр крови взрослого человека в норме содержит около 2,5—7,5 миллиардов нейтрофилов.[12] Их диаметр около 10 микрометров[70] и живут только в течение 5 дней.[31] Как только поступает соответствующий сигнал, они в течение примерно 30 минут выходят из крови и достигают зоны инфекции.[71] Они способны быстро поглощать чужеродный материал. Нейтрофилы не возвращаются в кровь; они превращаются в клетки гноя и погибают.[71] Зрелые нейтрофилы меньше, чем моноциты и имеют сегментированные ядра с несколькими секциями; каждая секция соединяется с хроматиновыми нитями (нейтрофил может иметь 2-5 сегмента). Обычно нейтрофилы не выходят из костного мозга до наступления зрелости, но при инфекции высвобождаются в кровь предшественники нейтрофилов — миелоциты и промиелоциты.[72]
Внутриклеточные гранулы нейтрофилов человека разрушают белки и обладают бактерицидными свойствами.[73] Нейтрофилы способны выделять продукты, которые стимулируют моноциты и макрофаги. Нейтрофильные выделения усиливают фагоцитоз и образование реактивных форм кислорода, участвуя таким образом во внутриклеточном уничтожении.[74] Выделения от первичных гранул нейтрофилов стимулируют фагоциоз бактерий, покрытых IgG.[75]
Дендритная клетка
Дендритные клетки — специализированные антиген-презентирующие клетки, у которых есть длинные отростки, называемые дендритами,[76] которые помогают поглощать микробы и другие патогены.[77][78] Дендритные клетки находятся в тканях, которые контактируют с окружающей средой, в основном в коже, внутренней оболочке носа, лёгких, желудка и кишечника.[79] После активации, они созревают и мигрируют в лимфатические ткани, где взаимодействуют с Т- и B-лимфоцитами для возникновения и организации приобретённого иммунного ответа.[80] Зрелые дендритные клетки активируют Т-хелперы и Т-киллеры.[81] Активированные Т-хелперы взаимодействуют с макрофагами и B-лимфоцитами чтобы активировать их, в свою очередь. Кроме того, дендритные клетки способны влиять на возникновение того или иного типа иммунного ответа; когда они перемещаются в лимфатические зоны, они способны активировать находящиеся там Т-лимфоциты, которые затем дифференцируют в Т-киллеры и Т-хелперы.[82]
Тучные клетки имеют Толл-подобные рецепторы и взаимодействуют с дендритными клетками, Т- и B-лимфоцитами. Тучные клетки выделяют MHC молекулы класса II и могут принимать участие в презентации антигена; однако, роль тучных клеток в презентации антигена ещё не достаточно изучена.[83] Тучные клетки способны поглощать, убивать грамотрицательные бактерии (например, сальмонелла) и обрабатывать их антигены.[84] Они специализируются на обработке фимбриальных белков на поверхности бактерий, которые участвуют в прикреплении к тканям.[85][86] Кроме этих функций, тучные клетки образуют цитокины, которые запускают реакцию воспаления.[87] Это важная часть уничтожения микробов, потому что цитокины привлекают больше фагоцитов к зоне инфекции.[84]
| Кровь | нейтрофилы, моноциты |
| Костный мозг | макрофаги, моноциты, синусоидные клетки, обкладочные клетки |
| Костная ткань | Остеокласты |
| Кишечник и кишечные бляшки Пейера | макрофаги |
| Соединительная ткань | Гистиоциты, макрофаги, моноциты, дендритные клетки |
| Печень | клетки Купфера, моноциты |
| Лёгкое | самовоспроизводящиеся макрофаги, моноциты, тучные клетки, дендритные клетки |
| Лимфатическая ткань | свободные и фиксированные и моноциты, дендритные клетки |
| Нервная ткань | Клетки микроглии (CD4+) |
| Селезёнка | свободные и фиксированные макрофаги, моноциты, синусоидные клетки |
| Тимус | свободные и фиксированные макрофаги и моноциты |
| Кожа | постоянные клетки Лангерханса, другие дендритные клетки, макрофаги, тучные клетки |
Умирающие клетки и чужеродные организмы поглощаются клетками, отличными от «профессиональных» фагоцитов.[89] К таким клеткам относят эпителиальные, эндотелиальные, паренхиматозные клетки и фибробласты. Их называют непрофессиональными фагоцитами, чтобы подчеркнуть, что в отличии от профессиональных фагоцитов, фагоцитоз для них не основная функция.[90] Фибробласты, например, которые могут фагоцитировать коллаген в процессе ремоделирования шрамов, также способны частично поглощать чужеродные частицы.[91]
Непрофессиональные фагоциты более ограничены, чем профессиональные, в отношении частиц, которые они могут поглотить. Это связано с отсутствием у них эффективных фагоцитарных рецепторов, в частности опсонинов.[11] Кроме того, большинство непрофессиональных фагоцитов не образуют реактивные кислород-содержащие молекулы для фагоцитоза.[92]
| Кровь, лимфа и лимфатические узлы | Лимфоциты |
| Кровь, лимфа и лимфатические узлы | Естественные киллеры и крупные гранулярные лимфоциты |
| Кожа | Эпителиоциты |
| Кровеносные сосуды | Эндотелиоциты |
| Соединительная ткань | Фибробласты |
| Кровь | Эритроциты |
Клетки бактерии Staphylococcus aureus: крупные, волокнистые капсулы, защищающие от атаки фагоцитов.
Патоген вызывает инфекцию, если только он преодолел защиту макроорганизма. Патогенные бактерии и простейшие развивают различные механизмы устойчивости к атакам фагоцитов, и многие из них действительно выживают и размножаются внутри фагоцитирующих клеток.[93][94]
У бактерий есть несколько способов избежать контакта с фагоцитами. Во-первых, они могут расти в местах, куда фагоциты не способны попасть (например, повреждённый кожный покров). Во-вторых, бактерия может подавлять реакцию воспаления; без этой реакции фагоциты не способны реагировать на инфекцию адекватно. В-третьих, некоторые виды бактерий могут замедлять способность фагоцитов перемещаться в зону инфекции, препятствуя хемотаксису.[93] В-четвёртых, некоторые бактерии способны избегать контакта с фагоцитом путём обмана иммунной системы, которая начинает «думать», что бактерия — клетка самого макроорганизма. Бледная трепонема (бактерия, вызывающая сифилис) скрывается от фагоцитов, покрывая свою поверхность фибронектином,[95] который естественно образуется в макроорганизме и играет важную роль в заживлении раны.[96]
Бактерии часто образуют белки или сахара, которые покрывают их клетки и препятствуют фагоцитозу; они входят в состав бактериальной капсулы.[93] Например, K5 капсула и O75 O антиген присутствуют на поверхности Escherichia coli,[97] и экзополисахаридные капсулы Staphylococcus epidermidis.[98]Пневмококк образует несколько типов капсул, которые обеспечивают различные уровни защиты,[99] и стрептококки группы А образуют белки, например M белок и фимбриальные белки, которые блокируют процесс поглощения. Некоторые белки препятствуют опсонин-связанному поглощению; Staphylococcus aureus образует белок А для блокирования рецепторов антител, что снижает эффективность опсонинов.[100]
Риккетсия (красным цветом), которая живёт в цитоплазме непрофессионального фагоцита.
У бактерий развиты способы выживания внутри фагоцитов, где они продолжают уклонятся от иммунной системы.[101] Для безопасного проникновения внутрь фагоцита они выделяют белки, называемые «инвазинами». Оказавшись внутри клетки, они остаются в цитоплазме и избегают воздействия токсических веществ, содержащихся в фаголизосомах.[102] Некоторые бактерии препятствуют слиянию фагосомы и лизосомы.[93] Другие патогены, например лейшмании, образуют высокомодифицированные вакуоли внутри фагоцитов, что позволяет им сохранять устойчивость и размножаться.[103]Legionella pneumophila образует секрет, который заставляет фагоциты сливаться с везикулами, отличными от тех, что содержат токсичные вещества.[104] Другие бактерии способны жить внутри фаголизосом. Staphylococcus aureus, например, образует ферменты каталаза и супероксиддисмутаза, которые разрушают химические вещества (например, пероксид водорода), образуемые фагоцитами для уничтожения бактерий.[105] Бактерии могут выходить из фагосомы перед образованием фаголизосомы: Listeria monocytogenes способна образовывать отверстие в стенке фагосомы, используя ферменты, называемые листериолизин O и фосфолипаза C.[106]
У бактерий развиты некоторые способы уничтожения фагоцитов.[100] К ним относятся цитолизины, которые образуют поры в клеточном мембране фагоцитов, стрептолизины и лейкоцидины, которые вызывают разрыв гранул нейтрофилов освобождение токсических веществ,[107][108] и экзотоксины, которые снижают поступления АТФ для фагоцитов, необходимых для фагоцитоза. Будучи поглощёнными, бактерии могут убивать фагоциты высвобождая токсины, которые перемещаются из мембран фагосом или фаголизосом к другим частям клетки.[93]
Множество мелких клеток лейшмании внутри более курпных клеток
Некоторые стратегии выживания часто связаны с нарушением цитокиновой и других методов передачи сигнала в клетке для предотвращения реакции фагоцитов на инвазию.[109] Такие паразитические простейшие как например Toxoplasma gondii, Trypanosoma cruzi и Leishmania инфицируют макрофаги; каждый из них имеет уникальный способ подавления активности макрофагов. Некоторые виды лейшманий изменяют систему сигналов инфицированных макрофагов, подавляют продукцию цитокинов и микробоцидных молекул (оксида азота, реактивных форм кислорода) и нарушают презентацию антигена.[110]
Макрофаги и нейтрофилы, в частности, играют центральную роль в воспалительном процессе путём высвобождения белков и низкомолекулярных воспалительных медиаторов, которые контролируют инфекцию, но могут повреждать ткани макроорганизма. В общем, цель фагоцитов — разрушение патогенов путём поглощения их и последующего воздействия на них токсичными веществами внутри фаголизосом. Если фагоцит не в состоянии поглотить свою цель, эти токсические агенты могут быть выделены в окружающую среду (действие называется «фрустрированный фагоцитоз»). Так как эти агенты токсичны и для клеток макроорганизма, они могут вызвать значительное повреждение здоровых клеток и тканей.[111]
Если нейтрофилы освобождают содержимое гранул (реактивные формы кислорода и протеазы) в почках, то происходит деградация внеклеточного матрикса макроорганизма, что может привести к повреждению клубочковых клеток, влияя на их способность фильтровать кровь и вызывая изменения в их форме. Кроме этого, продукты фосфолипазы (например, лейкотриены) усиливают повреждение. Это освобождение веществ способствует хемотаксису большего количества нейтрофилов в зону инфекции и клубочковые клетки могут быть повреждены ещё сильнее путём прикрепления молекул при миграции нейтрофилов. Повреждения клубочковых клеток может привести к почечной недостаточности.[112]
Нейтрофилы также играют ключевую роль в развитии большинства форм острого повреждения лёгких.[113] При этом активированные нейтрофилы высвобождают содержимое своих токсичных гранул в окружающую среду лёгких.[114] Эксперименты показали, что уменьшение числа нейтрофилов снижают эффекты острого повреждения лёгких,[115] но лечение подавлением нейтрофилов клинически не реалистично, так как при этом организм становится уязвим для инфекций.[114] В печени, повреждение нейтрофилами может способствовать дисфункции и повреждению в ответ на освобождение эндотоксина, выделяемого бактерией, сепсис, травму, алкогольный гепатит, ишимию и гиповолемический шок в результате острой кровотечения.[116]
Химические вещества, высвобождаемые макрофагами, могут также повреждать ткани макроорганизма. Фактор некроза опухоли-α (ФНО) — важное химические вещество, выделяемое макрофагами, которое приводит к свёртыванию крови в мелких сосудах, что предотвращает распространение инфекции.[117] Однако, если бактериальная инфекция проникает в кровь, ФНО-α выделяется в жизненно-важных органах, где может вызвать вазодилатацию и уменьшить объём плазмы; это в свою очередь может привести к септическому шоку. При септическом шоке, высвобождение ФНО-α вызывает блокаду мелких сосудов, которые кровоснабжают жизненно-важные органы и может возникнуть их недостаточность. Септический шок может привести к смерти.[13]
Фагоцитоз, вероятно, появился на ранних этапах эволюции ,[118] впервые возникнув у одноклеточных эукариот.[119] Амёбы — одноклеточные простейшие, которые отделились от дерева, ведущего к многоклеточным, но они передали множество специфических функций фагоцитарным клеткам млекопитающих.[119] Амёба Dictyostelium discoideum, например, живёт в почве и питается бактериями. Как и фагоциты животных, они поглощают бактерии путём фагоцитоза в основном с помощью Толл-подобных рецепторов и имеют другие биологические функции, общие с макрофагами.[120] Амёбы Dictyostelium discoideum социальны; они слипаются вместе, когда голодают, превращаясь в мигрирующую псевдоплазмодию или личинку. Такой многоклеточный организм в итоге образует плодовое тело со спорами, которое устойчиво к агрессии окружающей среды. Перед образованием плодовых тел, клетки могут перемещаться как личинкоподобные организмы в течение нескольких дней. В течение этого времени воздействие токсинов или бактериальных патогенов может представлять опасность для выживания амёб, ограничивая образование спор. Некоторые амёбы поглощают бактерии и абсорбируют их токсины. В конечном итоге эти амёбы погибают. Они генетически схожи с другими амёбами в личинке и жертвуют собой для защиты других амёб от бактерий, что схоже с самопожертвованием фагоцитов иммунной системы человека. Такая врождённая иммунная функция у социальных амёб предполагает, что древний механизм поглощения клеток мог быть приспособлен для защитной функция задолго до появления разнообразных животных.[121] Но общее происхождение с фагоцитами млекопитающих не доказано. Фагоциты присутствуют во всё животном царстве,[4] начиная от морской губки и заканчивая насекомыми, низшими и высшими позвоночными.[122][123] Способность амёб различать своих и чужих стала основой иммунной системы многих видов.[8]
Русский зоолог Илья Ильич Мечников (1845—1916) впервые определил, что специализированные клетки участвуют в защите от микробной инфекции. В 1882 году, он изучал подвижные клетки в личинках морских звёзд, полагая, что они важны для иммунной защиты животных. Чтобы проверить эту мысль, он воткнул небольшие иголки от мандаринового дерева в личинку. Через несколько часов он обнаружил, что подвижные клетки окружили иголки.[124] Мечников отправился в Вену и поделился своей идеей с Карлом Фридрихом Вильгельмом, который предложил название «фагоцит» для клеток, которые наблюдал Мечников.[125]
Год спустя, Мечников изучал пресноводное ракообразное — Дафнию, маленькое прозрачное животное, которое можно изучать непосредственно под микроскопом. Он обнаружил, что грибные споры, которые нападали на дафний, были уничтожены фагоцитами. Он перенёс свои наблюдения на белые кровяные клетки млекопитающих и обнаружил, что Bacillus anthracis может быть уничтожена фагоцитами. Этот процесс он назвал фагоцитоз.[126] Мечников предположил, что фагоциты — первичная защита от проникающих микроорганизмов.
В 1903 году, Алмрот Райт обнаружил, что фагоцитоз поддерживается специфическими антителами, которые он назвал опсонинами.[127] Мечников был награждён (вместе с Паулем Эрлихом) в 1908 году Нобелевская премия по физиологии или медицине за свою работу по фагоцитам и фагоцитозу.[7]
Хотя значение этих открытий постепенно получило признание в начале двадцатого века, сложные взаимоотношения между фагоцитами и другими компонентами иммунной системы, не были известны вплоть до 1980-х годов.[128]
При написании этой статьи использовался материал из Энциклопедического словаря Брокгауза и Ефрона (1890—1907).
скачатьДанный реферат составлен на основе статьи из русской Википедии. Синхронизация выполнена 11.07.11 23:32:16Похожие рефераты: Мононуклеарный фагоцит.Категории: Физиология, Кровь, Клетки.
Текст доступен по лицензии Creative Commons Attribution-ShareAlike.wreferat.baza-referat.ru
Содержание
Краткий экскурс в историю……………………………………………………………………………………………… 2
Современное состояние учения офагоцитозе……………………………………………………………..5
Макрофагиперитонеального экссудата как модель
фагоцитоза и нарушений фагоцитарной активности…………………………………………….13
Получениемодели………………………………………………………………………………………………………….14
Методы регистрации результатов…………………………………………....................................................14
Некоторые моделируемые процессы
СНИЖЕНИЕ БАКТЕРИАЛЬНОЙ АКТИВНОСТИПЕРИТОНЕАЛЬНЫХ
МАКРОФАГОВ МЫШЕИ В УСЛОВИЯХ СОЧЕТАННОГО
ПРИМЕНЕНИЯ СТАФИЛОКОККОВОГО ЭНТЕРОТОКСИНА ТИПА А И ЭНДОТОКСИНА………………………………………………17
ОТМЕНАУСИЛИВАЮЩЕГО ФАГОЦИТОЗ ДЕЙСТВИЯ ОПСОНИНОВ
С ПОМОЩЬЮ ФРАГМЕНТОВ АНТИТЕЛ ПРОТИВ Fc-РЕЦЕПТОРОВ МАКРОФАГОВ……………………………...................................18
УСИЛЕНИЕ С ПОМОЩЬЮ ХИТОЗАНАРЕАКЦИИ
КОНТАКТНОГО ВЗАИМОДЕЙСТВИЯ МАКРОФАГА С ТИМОЦИТАМИ invitro…………………………………………………………..19
АКТИВАЦИЯ ФАГОЦИТАРНЫХ КЛЕТОК И КЛЕТОЧНОГО
ИММУНИТЕТА СИНТЕТИЧЕСКИМИ ПОЛИЭЛЕКТРОЛИТАМИ………………………………………………………………………………20
АКТИВАЦИЯМАКРОФАГОВ ПОД ВЛИЯНИЕМ СИНТЕТИЧЕСКОГОАНТИОКСИДАНТА……………………………………………… 22
ФАГОЦИТАРНАЯ АКТИВНОСТЬ МАКРОФАГОВ
ПЕРИТОНЕАЛЬНОГО ЭКССУДАТА МЫШЕЙ ДЕЙСТВИИ ПРЕПАРАТОВ ПЛАТИНЫ………………………………………………………23
ИЗУЧЕНИЕФАГОЦИТАРНОЙ АКТИВНОСТИ ПЕРИТОНЕАЛЬНЫХ МАКРОФАГОВ В
ОТНОШЕНИИYERSINIAPESTISС ДЕФЕКТНЫМИ И ПОЛНОЦЕННЫМИ FRA-ГЕНАМИ…………………………………………………25
ВЛИЯНИЕМОДИФИКАТОРОВ БИОЛОГИЧЕСКОГО ОТВЕТА ПРИРОДНОГО
ПРОИСХОЖДЕНИЯНА ФУНКЦИОНАЛЬНУЮ АКТИВНОСТЬ МАКРОФАГОВ……………………………………………………………….26
ПЕРИТОНЕАЛЬНЫЕМАКРОФАГИ КАК МОДЕЛЬ
ДЛЯИЗУЧЕНИЯ АТЕРОГЕННОГО ПОТЕНЦИАЛАСЫВОРОТКИ КРОВИ……………………………………………………………………...29
ВЛИЯНИЕГАМК, ГОМК И ГЛУТАМИНОВОИ
КИСЛОТЫНА ФУНКЦИОНАЛЬНУЮ АКТИВНОСТЬ ФАГОЦИТОВ……………………………………………………………………………32
Заключение………………………………………………………………………. ……………………………………………………………33
Некоторые другие моделиизучения фагоцитоза……………………………………………………………… 34
Литература…………………………………………………………………………………………………………………………………………36
Краткий экскурс в историю
Более 100 лет прошло с моментаоткрытия фагоцитарной теории, созданнойнашим великим натуралистом,лауреатом Нобелевской премии И. И. Мечниковым.Открытие, осмысление явления фагоцитоза и формулирование в общих чертахоснов фагоцитарной теории было сделано им в декабре 1882 г. В 1883 г. он изложил основы новой фагоцитарной теории вдокладе «О целебных силах организма» вОдессе на VIIсъезде естествоиспытателей и врачейи опубликовал их в печати. Были впервые высказаны основные положения фагоцитарной теории, которые И. И. Мечников развивал в последующем на протяжении всей своей жизни. Хотя сам фактпоглощения живыми клетками других частиц был описан многими натуралистамизадолго до ученого, однако только он дал блестящее толкование огромной ролифагоцитов в защите организма от болезнетворных микробов.
Много позже к70-летнему юбилею ученого коллега и друг И. И.Мечникова Эмиль Ру напишет: «Сегодня, мойдруг, Вы наблюдаете доктринуфагоцитоза со спокойным удовлетворением отца, дитя которого сделалохорошую карьеру в мире, но сколько беспокойств оно Вам доставило! Его появление вызвало протесты и сопротивление и в течение двадцати лет Вам пришлось сражаться за него». Доктрина фагоцитоза «… однаиз наиболее плодотворных в биологии: онасвязала явление иммунитета с внутриклеточным пищеварением, она объяснила наммеханизм воспаления и атрофии; она оживила патологическую анатомию,которая, не будучи в состоянии датьприемлемое объяснение, оставалась чисто описательной… Ваша эрудициятакая обширная и верная, что она служит всему миру».
И. И. Мечников утверждал, что«… иммунитет в инфекционных болезняхдолжен быть приписан активной целлюлярной деятельности. Среди клеточныхэлементов фагоциты должны занять первоеместо. Чувствительность и подвижность, способность поглощать твердыетела и вырабатывать вещества, могущие разрушать и переваривать микробов — вот главные факторы деятельностифагоцитов. Если эти свойства в достаточноймере развиты и парализуют патогенное действие микробов, тогда животноеот природы иммунно… когда фагоциты необнаруживают наличия всех или одного из этих свойств в достаточной степени, тоживотное восприимчиво к инфекции...».Вместе с тем, если бактериальные продукты вызывают у фагоцитовотрицательный хемотаксис или, если при положительном хемотаксисе фагоциты непоглощают бактерий или поглощают, но не убивают их, — также развиваетсясмертельная инфекция. Решение фундаментальных проблем сравнительнойэмбриологии и биологии, приведшее ккрупнейшим открытиям ученого, позволило И. И. Мечникову установить, что «фагоцитоз чрезвычайно распространен в животноммире… как на самой низшей ступени животнойлестницы, например, у простейших, так и… у млекопитающих животных ичеловека… фагоциты представляютсобой мезенхимальные клетки».
И. И. Мечников был в то же время первым, кто занялсясравнительным изучением явления фагоцитоза. Внимание ученого было обращено нетолько на традиционные лабораторные объекты, но и на таких представителей мираживотных, как дафнии, морские звезды, крокодилы, обезьяны. Сравнительноеизучение фагоцитоза было необходимо И. И. Мечникову для доказательствавсеобщности явлений поглощения и разрушения чужеродного материалафагоцитирующими мононуклеарами, широкого распространения в природе изучаемойим формы иммунологической защиты.
Клеточная теория Мечникова сразу наткнулась насопротивление. Прежде всего она была предложена в то время, когдабольшинство патологов видели в воспалительной реакции, а также в связанных сней микрофагах и макрофагах не защитную, а вредоносную реакцию. В то времясчитали даже, что, хотя фагоцитирующие клетки действительно способны поглощатьболезнетворные микроорганизмы, это приводит не к разрушению возбудителя, а кпереносу его в другие части тела и распространению болезни. Также в тот периодвремени интенсивно развивалась гуморальная теория иммунитета, основы которойбыли заложены П.Эрлихом. Были открыты антитела и антигены, были выявленымеханизмы гуморальной устойчивости организма против некоторых патогенныхмикроорганизмов и их токсинов (дифтерия, столбняк и др.). Как это ни странно, но два таких открытия немогли некоторое время ужиться вместе. Позднее в 1888 г. Наттолл нашел всыворотке нормальных животных вещества, токсичные для некоторыхмикроорганизмов, и показал, что такие антибактериальные свойства значительноповышаются в результате иммунизации животного. В дальнейшем было обнаружено,что в сыворотке имеются два разных вещества, совместное действие которыхприводит к лизису бактерий: термостабильный фактор, затем идентифицированныйкак сывороточные антитела, и термолабильный фактор, названный комплементом, илиалексином (от греч. aleksein — защищать).Учениксамого Мечникова Борде описал лизис эритроцитов гуморальными антителами икомплементом, и большинство исследователей стали соглашаться с Кохом, чтопобеду одержали гуморалисты.Мечников и егоученики отнюдь не собирались сдаваться. Были поставлены простые опыты, вкоторых микробы, помещенные в маленький мешочек из фильтровальнойбумаги, защищающий их от фагоцитов, сохраняли свою вирулентность, хотябуквально купались в тканевой жидкости, богатой антителами. ВАнглии сэр Элмрот Райт и С. Р. Дуглас попыталисьпримирить различия между этими двумя школами в своих капитальныхисследованиях процесса опсонизации (от греч. opsonein — делатьсъедобным). Эти ученые утверждали, что клеточный и гуморальный факторы являютсяодинаково важными и взаимозависимыми в том отношении, что гуморальные антитела,специфически реагируя со своей мишенью — микроорганизмом, подготавливаютего к фагоцитозу макрофагами.
В 1908 г. Шведская академия удостоила Нобелевской премии по медицинесовместно Мечникова — основателя клеточного направления и Эрлиха — олицетворявшего гуморалистские идеи того времени.Они были удостоены премии в качестве «признания их работ поиммунитету».
Заслуга Мечниковасостоит не только в создании им гениальной теории.<span Arial CYR",«sans-serif»">
Еще ранее он начал изучать заразные болезничеловека и домашних животных: вместе со своим учеником Н. Ф. Гамалеей он изучалтуберкулез, чуму рогатого скота, искал способы борьбы с вредителями сельскогохозяйства. К 1886 г. относится одно из важнейших событий в истории русскоймедицины. Летом этого года в Одессе начала работать созданная Мечниковым и еготалантливым учеником Н. Ф. Гамалеей первая русская бактериологическая станция.Он создал в России крупнейшую научную школу микробиологов. Выдающиеся ученые Н.Ф. Гамалея, Д. К. Заболотный, Л. А. Тарасевич и многие другие были учениками И.И. Мечникова. Илья Ильич Мечников умер в 1916 году, до конца жизни занимаясьвопросами иммунологии и клеточного иммунитета. А наука об иммунитете быстро истремительно развивалась. В этот период было необычайно много работ и ученых,изучавших факторы внутренней защиты организма.Период с 1910 по1940гг. был периодом серологии. В это время было сформулированоположение о специфичности и о том, чтоАТ являются естественными,высоковариабельными глобулинами. Большую роль здесь сыграли работы Ландштейнера, который пришел в выводу, что специфичность антител не являетсяабсолютной.
С 1905 появилисьработы (Сarrel, Guthrie) потрансплантации органов. В 1930г. К.Ландштейнер открывает группы крови. Работамипо фагоцитозу, бактериофагии, вирусам,патогенезу чумы занимается Амадей Боррель. Премия присуждена Ф. МакфарлейнуБернету (1899 — 1985) и Питеру Медавару (1915 — Англия) «за открытиеприобретенной иммунолотической толерантности». Медавар показал, что отторжение чужеродногокожного трансплантата подчиняется всем правилам иммунологической специфичности,и в основе его лежат такие же механизмы, как и при защите от бактериальных ивирусных инфекций. Последующая работа, которую он провел вместе с рядомучеников, заложила прочную основу для развития трансплантационнойиммунобиологии, которая стала важной научной дисциплиной и в дальнейшемобеспечила многие достижения в области клинической трансплантации органов. Бернетопубликовал книгу «Образование антител» (1941 г.). Со своим коллегой, ФранкомФеннером, Бернет утверждал, что способность к иммунологическим реакциямвозникает на сравнительно поздних стадиям эмбрионального развития и при этомпроисходит запоминание существующих маркеров «своего» у антигенов,присутствующих в данный момент. Организм в последующем приобретает к нимтолерантность и не способен отвечать на них иммунологической реакцией. Всеантигены, которые не запомнились, будут восприниматься как «не свои» и смогут вдальнейшем вызывать иммунологический ответ. Было высказано предположение, чтолюбой антиген, введенный в течение этого критического периода развития, будетзатем восприниматься как свой и вызывать толерантность, в результате чего несможет в дальнейшем активировать иммунную систему. Эти идеи были далее развитыБернетом в его клонально-селекционной теории образования антител. ПредположенияБернета и Феннера были подвергнуты экспериментальной проверке в исследованияхМедавара, который в 1953 г. на мышах чистых линий получили четкое подтверждениегипотезы Бернета — Феннера, описав феномен, которому Медавар дал названиеприобретенной иммунологической толерантности.
В 1969г. одновременно несколькими авторами (Р.Петров, М.Беренбаум, И.Ройт) былапредложена трехклеточнаясхема кооперации иммуноцитов виммунном ответе (Т-, В-лимфоцитов имакрофагов), определившая на многие годы изучение механизмов иммунного ответа, субпопуляционной организации клеток системы иммунитета.
Существенную роль в этих исследованиях сыграликинематографические методы. Возможность непрерывного динамического изучениямикробиологических объектов invivoи invitro в условиях, совместимыхс их жизнедеятельностью, визуализация невидимых для человеческого глаза электромагнитныхизлучений, регистрация как быстрых, так и медленных процессов, управлениемасштабом времени и некоторые другие характерные особенности исследовательскойкинематографии открыли большие и во многих отношениях уникальные возможностидля изучения взаимодействия клеток.
Представлениео фагоцитах за истекшее время подверглось существенной эволюции. В 1970 г. VanFurthи соавт.предложили новую классификацию, выделяющуюМФ из РЭС в отдельную системумононуклеарных фагоцитов. Исследователи отдали дань уважения И. И. Мечникову,пользовавшемуся термином «мононуклеарный фагоцит» еще в начале XXвека. Фагоцитарная теория не стала, однако, неизменяемойдогмой. Непрерывно накопляемые наукой факты изменили и усложнили понимание техявлений, в которых фагоцитоз казался решающим или единственным фактором.
Можно утверждать, что в наши днисозданное И. И. Мечниковым учение о фагоцитах переживает свое второе рождение,новые факты значительно обогатили его, показав, как это и предсказывал ИльяИльич, огромное общебиологическое значение. Теория И. И. Мечникова явилась мощныминдуктором прогресса иммунологии во всем мире, большой вклад в него внеслисоветские ученые. Однако и сегодня основные положения теории остаютсянезыблемыми.
Первостепенное значение фагоцитарнойсистемы подтверждается созданием в США общества ученых, занимающихся изучениемретикулоэндотелиальной системы (РЭС), издается специальный «JournalofReticulo-EndothelialSociety».
В последующие годы развитие фагоцитарнойтеории связано с открытием цитокиновой регуляции иммунного ответа и, конечно,изучения влияния цитокинов на клеточный ответ в том числе и макрофагов. На зареэтих открытий стоя ли работы таких ученых, как Н.Ерне,
Г. Келер, Ц. Милштейн.
В СССР бурный интерес к фагоцитам исвязанным с ними процессами наблюдался в 80-е годы. Здесь необходимо отметить работы А.Н.Маянского, изучавшеговлияния макрофагов не только в свете их иммунной функции. Он показал значениеклеток РЭС на функционирование таких органов как печень, легкие, желудочно-кишечныйтракт. Работы также проводили А.Д. Адо, В.М.Земсков, В.Г.Галактионов,эксперименты по изучению работы МФ в очаге хронического воспаления ставилСеров.
Следует сказать, что в 90-е годыинтерес к неспецифическому звену иммунитета упал. Отчасти это можно объяснитьтем, что все усилия ученых были в основном устремлены к лимфоцитам, но особенно– к цитокинам. Можно сказать, что сейчас продолжается «цитокиновый бум».
Однако это ни в коем случае неозначает, что актуальность проблемы упала. Фагоцитоз составляет пример тогопроцесса, интерес к которому не может пропасть. Будет открытие новых факторовстимулирующих его активность, будут обнаружены вещества угнетающие РЭС. Будутоткрытия, уточняющие тонкие механизмы взаимодействия МФ с лимфоцитами, с клеткамиинтерстиция, с антигенными структурами. Особенно это может быть актуальносейчас в связи с проблемой опухолевого роста и СПИД’а. Остается надеяться, чтов ряду открытий, начатых великим Мечниковым, будут стоять имена русских ученых.
СОВРЕМЕННОЕСОСТОЯНИЕ УЧЕНИЯ О ФАГОЦИТОЗЕ
Основныеположения о фагоцитах и системе фагоцитоза, блестяще сформулированные И. И.Мечниковым и разработанные его учениками и последователями, надолго определилиразвитие этого важнейшего направления биологии и медицины. Идея опротивоинфекционном иммунитете, которая так увлекла современников И. И.Мечникова, сыграла решающую роль в становлении клеточной иммунологии, эволюциивзглядов на воспаление, физиологию и патологию реактивности и резистентностиорганизма. Парадоксально и вместе с тем закономерно, что учение о фагоцитозеначалось с крупных обобщений и концепций, которые на протяжении многих летдополнялись фактами частного характера, мало повлиявшими на развитие проблемы вцелом. Волна современной иммунологической информации, изобилие изящных методови гипотез направили интересы многих исследователей в сторону изучениялимфоцитарных механизмов клеточного и гуморального иммунитета. И еслииммунологи быстро поняли, что без макрофага им не обойтись, то судьба другогокласса фагоцитирующих клеток — полинуклеарных (сегментоядерных) лейкоцитов — донедавнего времени оставалась неясной. Только теперь можно с уверенностьюсказать, что и эта проблема, сделав за последние 5—10 лет качественный скачок,прочно утвердилась и успешно развивается не только иммунологами, но ипредставителями смежных профессий — физиологами, патологами, биохимиками,клиницистами. Изучение полинуклеарных фагоцитов (нейтрофилов) — один изнемногих примеров в цитофизиологии, а тем более в иммунологии, когда числоисследований на объекте «человеческого происхождения» превосходит количестворабот, выполненных в эксперименте на животных.
Сегодняучение о фагоцитозе — это совокупность представлений о свободных и фиксированныхклетках костномозгового происхождения, которые, обладая мощным цитотоксическимпотенциалом, исключительной реактивностью и высокой мобилизационнойготовностью, выступают в первой линии эффекторных механизмов иммунологическогогомеостаза. Противомикробная функция воспринимается как частный, хотя и важный,эпизод этой общей стратегии. Доказаны мощные цитотоксические потенции моно- иполинуклеарных фагоцитов, которые, кроме бактерицидности, находят выражение вуничтожении малигнизированных и иных форм патологически измененных клеток,альтерации тканей при неспецифическом воспалении в иммунопатологическихпроцессах. Если нейтрофилы (доминирующий тип полинуклеаров) почти всегда нацеленына деструкцию, то функции мононуклеарных фагоцитов сложнее и глубже. Ониучаствуют не только в разрушении, но и в созидании, запуская фибробластическиепроцессы и репаративные реакции, синтезируя комплекс биологически активныхсубстанций (факторы комплемента, индукторы миелопоэза, иммунорегуляторныебелки, фибронектини пр.). Сбывается стратегический прогноз И. И. Мечникова,который всегда смотрел на фагоцитарные реакции с общефизиологических позиций,утверждая значение фагоцитов не только в защите от «вредных деятелей», но и вобщей борьбе за гомеостаз, которая сводится к поддержанию относительногопостоянства внутренней среды организма. «При иммунитете, атрофии, воспалении иизлечении, во всех явлениях, имеющих величайшее значение в патологии, участвуютфагоциты».
Мононуклеарные фагоциты, которые ранее относилик ретикулоэндотелиальной системе, выделены в самостоятельное семейство клеток— систему мононуклеарных фагоцитов, которая объединяет моноциты костногомозга и крови, свободные, и фиксированные тканевые макрофаги. Доказано, что,выходя из крови, моноцит меняется, адаптируясь к условиям той среды, в которуюпопадает. Это обеспечивает специализацию клетки, т. е. максимальноесоответствие тем условиям, в которых ей предстоит «работать». Не исключена идругая альтернатива. Подобие моноцитов может быть чисто внешним (как это случилосьс лимфоцитами), и часть из них предетерминирована к трансформации в различнееварианты макрофагов. Гетерогенность зрелых нейтрофилов хотя и существует, новыражена гораздо слабее. Они почти не меняются морфологически, попадая в ткани,в отличие от макрофагов живут там недолго (не более 2—5 сут) и явно необладают пластичностью, присущей моноцитам. Это высокодифференцированныеклетки, которые практически заканчивают свое развитие в костном мозге. Неслучайно, известные в прошлом попытки отыскать корреляцию между сегментациейядра и способностью лейкоцитов к фагоцитозу не увенчались успехом. Тем не менееидея о функциональной неоднородности морфологически зрелых нейтрофилов продолжаетполучать подтверждения. Известны различия между нейтрофилами костного мозга ипериферической крови, нейтрофилами крови, тканей и экссудатов. Причины ифизиологический смысл этих особенностей неизвестны. По-видимому, изменчивостьполинуклеаров в отличие от моноцитов-макрофагов носит тактический характер.
Изучение фагоцитоза ведется согласноклассическим постулатам И. И. Мечникова о фазах фагоцитарной реакции —хемотаксису, аттракции (связывании)и поглощении,уничтожении (переваривании). К характеристике каждого из этих процессов внастоящее время приковано внимание, им посвящают монографии, обзоры. Результатымногочисленных исследований позволили углубиться в суть этих реакций,конкретизировать молекулярные факторы, лежащие в их основе, нащупать общиеузлы и вскрыть частные механизмы клеточной реактивности. Фагоцитоз служитпрекрасной моделью для изучения миграционной функции, пространственнойориентации клеток и их органелл, слияния и новообразования мембран, регуляцииклеточного гомеостаза и других процессов. Иногда фагоцитоз нередко отождествляютс поглощением. Это явно неудачно, ибо нарушает исторически сложившееся представлениео фагоцитозе как об интегральном процессе, который объединяет сумму клеточныхреакций, начиная с распознавания объекта и кончая его разрушением илистремлением к разрушению. С функциональной точки зрения фагоциты могутпребывать в двух состояниях — покоящемся и активированном. В наиболее общемвиде активация — есть результат преобразования внешнего стимула в реакциюэффекторных органелл. Больше пишут об активированном макрофаге, хотя в принципето же самое можно сделать и для полинуклеаров. Надо выбрать лишь точку отсчета— к примеру, функциональный статус в сосудистом русле нормального организма.Активация различается не только степенью возбуждения индивидуальных клеток,но и масштабом охвата клеточной популяции в целом. В норме активированонебольшое количество фагоцитов. Появление раздражителя резко меняет этотпоказатель, отражая подключение фагоцитов к реакциям, корригирующимвнутреннюю среду организма. Стремление проактивировать фагоцитарную систему,усилив тем самым ее эффекторные возможности, неоднократно звучало в работах И.И. Мечникова. Современные исследования по адъювантам, биологическим ифармакологическим модуляторам мононуклеарных и полинуклеарных фагоцитов посуществу развивают эту мысль с позиций межклеточной кооперации, общей и частнойпатологии. В этом видится перспектива рационального воздействия навоспаление, репаративные и регенеративные процессы, иммунопатологию, резистентностьк острому и хроническому стрессу, устойчивость к инфекциям, опухолям и пр.
Многие признакиактивации стереотипны, повторяясь у всех фагоцитирующих клеток. К ним относятся изменение активности лизосомальных и мембранных ферментов,усиление энергетического и окислительного метаболизма, синтетических исекреторных процессов, изменение адгезивных свойств и рецепторной функцииплазматической мембраны, способности к случайной миграции и хемотаксису,поглощению и цитотоксичности. Если учесть, что каждая из этих реакций по своейприроде интегративна, то количество частных признаков, по которым можно судить о возбуждении клеток,будет огромным.
Один итот же раздражитель способен индуцировать все или большинство признаков активации.Однако это, скорее, исключение, чем правило. Сегодня многое известно оконкретных механизмах, реализующих эффекторные свойства моно и полинуклеарныхфагоцитов. Расшифрована структурная основа двигательных реакций, открытыорганеллы, обеспечивающие векторную ориентацию в пространстве, изученызакономерности и кинетика образования фаголизосом, установлена природа цитотоксичностии бактерицидности, определены синтетические и секреторные потенции, обнаруженырецепторные и каталитические процессы в плазматической мембране и пр.Становится все более очевидным, что дискретные проявления клеточнойреактивности обеспечиваются или по крайней мере инициируются обособленнымимеханизмами и могут возникать независимо друг от друга. Удается подавить или усилитьхемотаксис, не изменив способности к поглощению и цитотоксичности, секреция несвязана с поглощением, повышение адгезивности не зависит от потреблениякислорода и т. д. Известны генетические дефекты, когда выпадает одна илинесколько из перечисленных функций, причем многие из них стереотипны поклинической симптоматике. Если к этому присовокупить патологию медиаторныхсистем, генерирующих хемоаттрактанты и опсонины, станет понятно, насколькосложным и одновременно конкретным должен быть сегодня диагноз, констатирующийнарушение фагоцитоза.
Крупнымсобытием явилось утверждение молекулярных основ цитотоксичности (в том числе бактерицидности) и ее отношения к реактивности клеток.Стремление понять сущность реакций, приводящих к гибели поглощенных бактерий, проглядывает в большинстве работ И. И. Мечникова. Многиегоды эта проблема традиционно сводилась к«перевариванию», в котором участвуют гидролитические ферменты (цитазы, по И. И. Мечникову), определяющие,как полагали, антимикробные свойства фагоцитов. Эта позиция была сильнопоколеблена, после того как оказалось, что лизосомальные гидролазы обладают слабой бактерицидностью invitro или лишены ее. В основу современных взглядов положены наблюдения, свидетельствующие об усилении окислительногометаболизма активированныхфагоцитов и разобщении двух главных событий —киллинг-эффекта и деградации убитых, нежизнеспособных объектов. Прежняя терминология, в которой закреплена причиннаясвязь между уничтожением живой мишени и переваривающей функцией клетки, оставлена. Переваривание, которое обусловленокислыми и нейтральными гидролазами, преформированными в гранулах, являетсявторичным и нацелено на объекты, убитые зависимыми и независимыми от кислородамеханизмами — биооксидантами, системой миелопероксидазы, катионными белками, лактоферрином, лизоцимом. Реализация цитотоксичности отражает реактивноевозбуждение фагоцитирующих клеток, которые секретируют эффекторныемолекулы внутрь фагосом (собразованием фаголизосом) либо наружу, атакуя внеклеточные (непоглощенные) объекты. То, что количество кислорода, поглощаемоголейкоцитами, значительно увеличивается при фагоцитозе, известно давно. Однакоистинное значение этого феномена, который в современной литературе частоназывают респираторным, или метаболическим, взрывом, осмыслено лишь в последниегоды. В отличие от многих клеток кислородное дыхание не является системойжизнеобеспечения фагоцитов — они хорошо переносят гипоксию и выполняют рядфункций в условиях анаэробиоза. При помощи респираторного взрывамоноциты-макрофаги и нейтрофилы решают чисто эффекторные задачи, «вооружаясь»против микробов и других объектов, которые воспринимаются ими какантигомеостатические факторы. В анаэробной среде фагоциты сохраняют способностьк поглощению, но резко снижают токсичность в отношении многих патогенных и условно-патогенныхбактерий. Ключевой механизм сводится к активации мембранных оксидаз, катализирующихперенос электронов с НАДФН на молекулярный кислород. Это стимулирует окислениеглюкозы в гексозомонофосфатном шунте, приводя к гиперпродукции перекиси водородаи свободных радикалов кислорода — биологических оксидантов с мощнымицитотоксическими потенциями. Клиническое значение подобных реакций сталоочевидным после того, как было обнаружено фатальное снижениепротивоинфекционного иммунитета у детей с врожденной патологией системы респираторноговзрыва нейтрофилов. Впрочем, было бы неверно противопоставлять различныеантимикробные факторы. Их эффективность во многом зависит от взаимнойсбалансированности условий, в которых происходит фагоцитоз, вида микроба. Нейтрофилы,лишенные возможности использовать бактерицидные свойства активированногокислорода, тем не менее нормально убивают эпидермальный стафилококк, синегнойнуюпалочку, зеленящий стрептококк, облигатные анаэробы. Относительная устойчивостьк фагоцитозу определяется суммой признаков — поверхностными свойствамимикробной клетки, наличием факторов типа лейкотоксинов и антифагинов, инактивациейбиооксидантов и пр. Давно обнаружена способность некоторых бактерий тормозитьобразование фаголизосом. Этот механизм,который исключает контакт с цитотоксическими компонентами фагоцитов, обеспечиваетдлительное персистирование в макрофагах туберкулезной палочки, aвнейтрофилах — бруцелл, а также других микроорганизмов. Одну из причин видят вповышении внутриклеточного уровня циклического аденозинмонофосфата, блокирующегомобилизацию гранул и их слияние с фагосомами. Этот пример показывает, насколькоглубокими могут быть взаимоотношения, которые складываются в процессе фагоцитоза.
Становлениевзглядов на механизмы цитотоксичности фагоцитов не подорвало мечниковской концепции о цитазах как о медиаторах, опосредующих деструктивные функции клеток. И. И. Мечников не разподчеркивал, что, кроме разрушения микробов, фагоциты способны повреждать собственные ткани. Эти идеиполучили блестящее развитие в современных работах по ферментологиилизосомальных гранул и способам их подключения к фагоцитарным реакциям. В гранулах моно- иполинуклеарных фагоцитовидентифицированбольшой арсенал гидролитических ферментов (нейтральных и кислых гидролаз), для каждого из которыхможно подыскать мишень во внеклеточном пространстве. Под их прицелом находятсяколлагеновые и эластиновые волокна, пептидогликановый матрикс хряща, фибронектин, факторыкомплемента, системы калликреин-кининов, свертывания, фибрино-лиза,иммуноглобулины, клеточные мембраны. В противовес старым представлениям сегоднясделан акцент на активном, секреторном высвобожденииэффекторных молекул, которое значительноповышает эффекторную пластичность клетки, позволяя в кратчайший срокмобилизовать и рационально использовать свои возможности в физиологических ситуациях и в ходе разнообразных патологических процессов. Секретируя, фагоцитывоздействуют на другие медиаторные системы и разрушают внеклеточные объекты, размеркоторых исключает возможность поглощения. Так, по-видимому, обстоит дело приэмфиземе легких, в реакциях на иммунныекомплексы, при остром и хроническом воспалении. Кроме гидролаз и другихкомпонентов лизосомальных гранул, активированные фагоциты выделяют пирогены,интерфероны и интерфероноподобные вещества, простагландины, тромбоксаны,биооксиданты, монокины, факторы,стимулирующие предшественники миелопоэза и пр. Лейкотриены вызывают сокращение гладких мышц и повышениесосудистой проницаемости, действуя в 100— 10000 раз сильнее, чем гистамин.
Прав былИ. И. Мечников, когда говорил о широком спектре задач, решаемых фагоцитами, имногообразии их функциональных контактов («живой цепи», по И. И. Мечникову) сдругими клетками и тканями. Активированные макрофаги и нейтрофилы служат однимиз самых ярких примеров экстренной мобилизации эффекторных механизмов с обширнойсферой приложения в масштабе не только соединительной ткани, но и всегоорганизма.
Активаторымоноцитов-макрофагов и полинуклеаров образуются в системах комплемента, свертывания,фибринолиза, калликреин-кининов, иммуноглобулинов, выделяются лимфоцитами, фибробластами, тромбоцитами, эндотелием. Сложные взаимоотношенияскладываются и внутри самой фагоцитарной системы. Моноцитарный инфильтрат при воспалении формируется подвлиянием хемоаттрактантов, продуцируемых нейтрофилами, которые первыми мигрируют в зону альтерации. В своюочередь моноциты и макрофаги выделяют факторы, избирательно активирующие нейтрофилы. Существенное значение имеетфункциональная кооперация между однотипными фагоцитами, которая предполагает участие «аутокаталитических» механизмов, контролирующих миграционную, секреторную и другие функцииактивированных клеток. Липоксигеназные метаболиты арахидоновой кислоты — различные варианты гидроксиэйкозантетраеновые кислоты хемотаксичны в ничтожных дозах (особенно для нейтрофилов) и, являясьпотенциальными «осколками» клеточного метаболизма унифицируют сигналы, которыеобеспечивают направленную миграцию фагоцитов в очаги тканевого повреждения. Любая травма любой ткани может стать инициатором подобны
www.ronl.ru
Применительно к процессу фагоцитоза применяют следующие уточняющие определения:
1. Собственно фагоцитоз: поглощение клеток, их фрагментов и их внутриклеточное переваривание.
2. Незавершённый фагоцитоз.
3. Иммунный (специфический) фагоцитоз и опсонизация.
4. Неспецифический фагоцитоз характерен, например, для альвеолярных макрофагов, захватывающих пылевые частицы различной природы, сажу и т.п.
5. Ультрафагоцитоз — захватывание фагоцитом мелких корпускулярных частиц (пыли, попадающей с воздухом в лёгкие, или инородных частиц в тканях).
Стадии фагоцитоза.В процессе фагоцитоза условно выделяют несколько основных стадий:
1. Сближение фагоцита с объектом фагоцитоза.
2. Распознавание фагоцитом объекта поглощения и адгезия к нему.
3. Поглощение объекта фагоцитом с образованием фаголизосомы.
4. Разрушение объекта фагоцитоза.
Сближение фагоцита с объектом фагоцитоза.Первая стадия фагоцитоза — сближение фагоцита с объектом фагоцитоза — рассмотрена выше в разделе главы 5 «Направленная миграция лейкоцитов».
Распознавание объекта фагоцитоза. К наиболее существенным этапам относятся: распознавание поверхностных детерминант объекта фагоцитоза, опсонизация, адгезия фагоцита к объекту фагоцитоза, экспрессия на поверхности фагоцита гликопротеинов HLA I и II.
· Распознавание поверхностных детерминант объекта фагоцитоза. Большинство объектов идентифицируется с помощью рецепторов на поверхности лейкоцитов. К таким объектам относятся микроорганизмы, грибы, паразиты, собственные повреждённые или опухолевые, или ви-руссодержащие клетки, а также фрагменты клеток.
· Опсонизация (иммунный фагоцитоз) — связывание AT с клеточной стенкой микроорганизма с последующим эффективным поглощением образовавшегося комплекса фагоцитом при взаимодействии Fc-фрагмента AT с соответствующим Fc-рецептором (FcR) на мембране фагоцита. Наиболее активные опсонины: Fc-фрагмент IgG, IgM, факторы комплемента C3bi, лектины.
IgG. Бактерия, покрытая молекулами IgG, эффективно фагоцитируется макрофагом или нейтрофилом. Fab-фрагменты IgG связываются с антигенными детерминантами на поверхности бактерии, после чего те же молекулы IgG своими Fc-фрагментами взаимодействуют с рецепторами Fc-фрагментов, расположенными в плазматической мембране фагоцита, и активируют фагоцитоз.
IgM. Большая молекула IgM легко активирует комплемент и служит опсонином при фагоцитозе. Многие AT к грамотрицательным бактериям являются IgM.
Адгезия фагоцита к объекту фагоцитоза реализуется с участием рецепторов лейкоцита FcyR (при наличии у объекта соответствующего лиганда) и молекул адгезии (при отсутствии лиганда, например, у неклеточных частиц). При фагоцитозе в зернистых лейкоцитах происходит активация реакций метаболизма («метаболический взрыв»), что обеспечивает ряд важных событий: экспрессию гликопротеинов HLA I и II и молекул адгезии, респираторный взрыв, а также дегрануляцию лейкоцитов.
Метаболический взрыв. К наиболее значимым метаболическим изменениям относятся активация реакций пентозофосфатного шунта, усиление гликолиза, потенцирование глико-генолиза, накопление восстановленного НАДФ.
Дегрануляция лейкоцитов. Дегрануляция нейтрофилов, эозинофилов и базофилов сопровождается высвобождением в интерстициальную жидкость медиаторов воспаления (например, ИЛ-1 и ИЛ-6, ФНО, лейкотриенов) и активных форм кислорода, образовавшихся при респираторном взрыве.
Поглощение объекта и образование фаголизосомы.Фагоцитируемый материал погружается в клетку в составе фагосомы — пузырька, образованного плазматической мембраной. К фагосоме устремляются лизосомы и выстраиваются по её периметру. Затем мембраны фагосомы и ли-зосом сливаются и образуется фаголизосома. В образовании фаголизосомы принимают участие и специфические гранулы нейтрофильного лейкоцита — видоизменённые лизосомы, а для самого процесса слияния необходимы мик-рофиламенты цитоскелета, Са2+, протеинкиназа С.
Погружение объекта фагоцитоза в лейкоцит сопровождается секрецией медиаторов воспаления и других компонентов специфических гранул лейкоцита. При дегрануляции все эти факторы поступают в воспалительный экссудат, где оказывают бактериолитическое и цитолитическое действие.
Внутриклеточное «переваривание».Разрушение объекта фагоцитоза — внутриклеточное «переваривание» — реализуется в результате активации двух сложных механизмов: кислородзависимой (респираторный взрыв) и кислороднезависимой цитотоксичности фагоцитов.
Кислороднезависимые механизмы активируются в результате контакта опсонизированного объекта с мембраной фагоцита. В процессе фагосомо-лизосомального слияния первыми с мембраной фагосомы сливаются гранулы, содержащие лактоферрин и лизоцим, затем к ним присоединяются азурофильные гранулы, содержащие катионные белки (например, САР57, САР37), протеиназы (например, эластаза и коллагеназа), катепсин G, дефен-зины и др. Эти химические соединения вызывают повреждение клеточной стенки и нарушение некоторых метаболических процессов; в большей степени их активность направлена против грамположительных бактерий.
Кислородзависимая цитотоксичность фагоцитов играет ведущую роль в деструкции объекта фагоцитоза. Цитотоксичность сопряжена со значительным повышением интенсивности метаболизма с участием кислорода. Этот процесс получил название метаболического (дыхательного, респираторного, кислородного) взрыва. При этом потребление кислорода фагоцитом может увеличиться в течение нескольких секунд во много раз.
В результате дыхательного взрыва образуются цитотоксичные метаболиты кислорода (так называемые активные формы кислорода), свободные радикалы и перекисные продукты органических и неорганических соединений.
К этому времени в цитоплазме фагоцита накапливается большое количество восстановленного НАДФ. НАДФ-оксидаза (флавопротеин цитох-ромредуктаза) плазматической мембраны и цитохром b в присутствии хинонов трансформируют О2 в анион супероксида (О2~), проявляющий выраженное повреждающее действие.
В последующих реакциях О2~ может трансформироваться в другие активные формы: синглетный кислород, гидроксильный радикал (ОН-), пероксид водорода. Последний процесс катализирует СОД.
Пероксид водорода (Н2О2) проявляет меньший, чем О2~~ повреждающий эффект, но в присутствии миелопероксидазы конвертирует ионы С1~ в ионы НСlO-, обладающие бактерицидным свойством, во многом аналогичным эффекту хлорной извести (NaCIO).
Образующиеся активные радикалы обусловливают повреждение и деструкцию белков и липидов мембран, нуклеиновых кислот и других химических соединений объекта фагоцитоза. При этом сам фагоцит защищен от действия указанных выше агентов, поскольку в его цитоплазме имеются комплексы защитных неферментных факторов (глутатион, витамины Е, С, жирные кислоты) и ферментов (СОД, устраняющая супероксидный анион, глутатионпероксидаза и каталаза, инактивирую-щие Н2О2).
Повреждённый кислородзависимыми и независимыми механизмами объект фагоцитоза подвергается деструкции с участием лизосомальных ферментов. Образовавшиеся продукты какое-то время хранятся в остаточных тельцах и могут утилизироваться клеткой или выводиться из неё путём экзоцитоза.
www.ronl.ru
Содержание (план)
см. Миграция лейкоцитов
Механизм адгезии фагоцитов типоспецифичен и основывается на взаимодействии специальных мембранных рецепторов фагоцитов с патогенсвязанными молекулярными шаблонами, входящими в состав поверхностных структур бактериальных клеток. Чаще всего в качестве молекулярных шаблонов выступают липополисахариды, остатки маннозы и липотейхоевые кислоты. При этом важен не сам факт наличия молекул того или другого шаблона на поверхности клетки, а плотность их распределения на том участке поверхности бактерии, с которым непосредственно взаимодействует фагоцит. Так, единичные остатки маннозы могут встречаться и на собственных клетках организма, особенно при частичной потере поверхностных экранирующих молекул. В то же время на бактериальных клетках содержится огромное количество остатков маннозы, что позволяет фагоциту плотно фиксироваться на бактерии и создает достаточный по силе активационный сигнал для перестройки цитоскелета в месте фиксации объекта и последующей его интернализации (поглощения). Следует отметить, что даже при высокой плотности экспрессии молекулярных шаблонов захват объектов, опосредованный их распознаванием, во многих случаях является недостаточно эффективным.
Для решения этой проблемы в процессе адгезии для фагоцитов нарабатывается новый способ распознавания объектов — с помощью опсонинов-антител. Он состоит в синтезе специфических антител к многочисленным и разнообразным поверхностным молекулам бактериальных клеток (антигенам). Синтезированные иммуноглобулины, фиксируясь своими антигенсвязывающими центрами на поверхностных молекулах (т. е. осуществляя специфическое распознавание), густо покрывают поверхность бактериальной клетки, «визуализируя» ее для фагоцитов,
Способность к опсонизации обусловлена уникальной структурой антител. Молекула иммуноглобулинов имеет два полюса: на одном из них располагается антигенсвязывающий сайт (Fab-фрагмент), специфический по своей пространственной структуре, а на другом — так называемый Fc-участок, идентичный у всех антител. При этом Fab-фрагмент взаимодействует с антигеном бактериальной клетки, a Fc-участок молекулы оказывается ориентированным наружу и является вполне удобной мишенью для распознавания фагоцитом, тем более что последний имеет в своем распоряжении соответствующие рецепторы (так называемые Fc-рецепторы). Таким образом, антитела выполняют роль своеобразных адаптеров, трансформируя уникальный антигенный спектр патогена в однообразный набор собственных Fc-участков, доступный для распознавания фагоцитами. Исходя из физико-химических представлений, взаимодействие рецепторов фагоцитов с опсонинами является более эффективным, нежели с молекулярными шаблонами, так как первые — собственные молекулы, а вторые — молекулы патогена.
Поскольку между опсонинами и рецепторами макрофагов существует определенная комплементарность (пространственное и физико-химическое взаимосоответствие), механизм фиксации фагоцита на объекте фагоцитоза получил название зипперного и напоминает действие замка-молнии.
Второй этап фагоцитоза заключается в поглощение (интернализация) объекта фагоцитоза, инициирующееся активационными сигналами, которые поступают внутрь фагоцитирующей клетки при взаимодействии ее рецепторов с опсонинами или молекулярными шаблонами. В основе процесса поглощения лежит перестройка цитоскелета фагоцита, который, как известно, определяет форму клетки. В результате подобной перестройки происходит инвагинация (втягивание) того участка мембраны, на котором фиксирован патоген. Последний поглощается все глубже внутрь цитоплазмы, пока не настанет момент, когда мембрана над ним сомкнется и он окажется окруженным своеобразным мембранным мешком, получившим название фагоцитарной вакуоли, или фагосомы. Важно отметить, что в фагосомах бактериальные клетки остаются вполне жизнеспособными. Снижение pH среды за счет усиленной работы протонных насосов, содержащихся в мембране фагосомы, чаще всего приводит лишь к инактивации микроорганизмов. Для их полного уничтожения необходимо поступление факторов агрессии, сконцентрированных в других органеллах — лизосомах. Подобные механизмы составляют субстрат последующего этапа фагоцитоза.
Первичными называют лизосомы, содержащие факторы агрессии, но лишенные объекта фагоцитоза. При слиянии с фагосомами первичные лизосомы трансформируются в фаголизосомы, или вторичные лизосомы, что и составляет третий этап фагоцитоза. Именно здесь с помощью как кислород-зависимых, так и кислород-независимых механизмов осуществляется деструкция захваченного патогена.
см. Активные формы кислорода
К кислород-зависимым механизмам микробицидности фагоцитов относят повреждения патогенов с помощью активных форм кислорода, а именно супероксид-аниона, синглетного кислорода, гидроксильного аниона, перекиси водорода, иона хлора и гипохлорной кислоты (НПО). Следует отметить, что действие двух последних компонентов во многом аналогично действию хлорной извести (NaCIO).
см. Гуморальные факторы врожденного иммунитета
К кислород-независимым механизмам относят эффекты гуморальных факторов, таких как лактоферрин, лизоцим, катионные белки, катепсины, дефензины и протеиназы, в основном направленных на грамположительные бактерии и основывающихся на повреждении клеточной стенки и нарушении некоторых важных метаболических процессов. Материал с сайта http://wiki-med.com
Когда захваченная бактериальная клетка окажется полностью разрушенной на мелкие фрагменты, ее остатки удаляются из фагоцита путем экзоцитоза. При этом пептиды некоторых антигенов транспортируются на поверхность клетки, где связываются с молекулами гистосовместимости II класса, т.е. происходит подготовка к осуществлению антигенной презентации, которая обеспечивает как инициацию иммунного ответа, так и его дальнейшее поддержание. В этом заключается преемственность иммунных реакций, поскольку реализация механизмов, первично повреждающих патоген, одновременно является предпосылкой для включения дальнейших процессов, направленных на повышение качества механизмов противомикробной защиты.
см. Патогенность бактерий
Поскольку фагоцитоз является наиболее эффективным компонентом противобактериального ответа, защитные механизмы, осуществляемые бактериальными клетками, в основном направлены на данное звено. Речь идет о разных патоген-опосредованных реакциях, нарушающих третий этап фагоцитоза, поскольку это — одновременно и важнейший, и наиболее чувствительный его механизм:
 На этой странице материал по темам:
На этой странице материал по темам: wiki-med.com
Фагоцитоз (от греч. phago – пожираю и cytos – клетка) представляет собой процесс поглощения и переваривания антигенных веществ, в том числе микроорганизмов, клетками мезодермального происхождения, названными фагоцитами.
И. И. Мечников разделил фагоциты на макрофаги и микрофаги. В настоящее время макро- и микрофаги объединены в единую систему макрофагов (СМФ). К этой системе относят тканевые макрофаги – эпителиоидные клетки, звездчатые ретикулоэндотелиоциты (клетки Купфера), альвеолярные и перитонеальные макрофаги, находящиеся в альвеолах и полости брюшины, белые отросчатые эпидермоциты кожи (клетки Лангерганса) и др. к микрофагам относятся: нейтрофилы, эозинофилы и базофилы.
Функции макрофагов чрезвычайно разнообразны. Они первые реагируют на чужеродное вещество, являясь специализированными клетками, поглощающими и уничтожающими в организме чужеродные субстанции (отмирающие клетки, раковые клетки, бактерии, вирусы и другие микроорганизмы, антигены, неметаболизируемые неорганические вещества).
Кроме того, макрофаги вырабатывают многие биологически активные вещества – ферменты (в том числе лизоцим, пероксидазу, эстеразу), белки комплемента, иммуномодуляторы типа интерлейкинов. Наличие на поверхности макрофагов рецепторов к иммуноглобулинам (Am) и комплементу, а также система медиаторов обеспечивает их взаимодействие6 с Т- и В- лимфоцитами. При этом макрофаги активируют защитные функции Т-лимфоцитов. Благодаря наличию рецепторов к комплементу и Am, а также Аг системы гистосовместимости (HLA) макрофаги принимают участие в связывании и распознавании антигенов.
Т. О. фагоцитам присущи три функции:
1 – защитная, связанная с очисткой организма от инфекционных агентов, продуктов распада тканей и т. д.;
2 – представляющая, заключающаяся в презентации лимфоцитам антигенных эпитолов на мембране фагоцита;
3 – секреторная, связанная с секрецией лизосомных ферментов и других биологически активных веществ – цитокинов, играющих важную роль в иммуногенезе.
Различают следующие последовательно протекающие стадии фагоцитоза.
1. Хемотаксис – целенаправленное передвижение фагоцитов в направлении химического градиента хемоаттрактантов в окружающей среде. Способность к хемотаксису связана с наличием на мембране специфических рецепторов для хемоаттрактантов (объектов фагоцитоза), в качестве которых могут выступать бактерии, продукты деградации тканей организма и др.
2. Адгезия (прикрепление)также опосредована соответствующими рецепторами, но может протекать в соответствии с законами неспецифического физико-химического взаимодействия. Происходит адсорбция частиц на поверхности макрофага.
3. Эндоцитоз (захват) – происходит инвагинация клеточной мембраны, захват чужеродной частицы и погружение ее в протоплазму. В результате эндоцитоза образуется фагоцитарная вакуоль – фагосома (т. е. пузырек в протоплазме вокруг поглощенной частицы).
4. Внутриклеточное переваривание – начинается по мере поглощения фагоцитируемых объектов. Происходит слияние фагосомы с лизосомой фагоцита, содержащей десятки ферментов, и образование фаголизосомы (деструкция) захваченной частицы ферментами.
При поглощении частицы, принадлежащей самому организму (например, погибшая клетка или ее части, собственные белки), происходит расщепление ее ферментами фаголизосомы до неантигенных веществ (аминокислоты, жирные кислоты, нуклеотиды, моносахара).
Если поглощается чужеродная частица, то ферменты фаголизосомы не в состоянии расщепить вещество до неантигенных компонентов. В таких случаях фаголизосома с оставшейся и сохранившей чужеродность частью антигена передается макрофагом Т- и В-лимфоцитам, т. е. включается специфическое звено иммунитета. Описанный механизм лежит в основе распознавания «своего» и «чужого» на уровне макрофага и явления фагоцитоза. В этом состоит презентативная, или представляющая, функция макрофагов. Специфические антигенные рецепторы, которые фиксируются на наружной мембране макрофагов, распознаются Т-лимфоцитами, последние передают информацию об Аг другим популяциям Т- и В-лимфоцитов и развивается специфический иммунный ответ.
Внутриклеточная участь захваченных фагоцитами микроорганизмов может быть различной в зависимости от их вирулентности и способности к внутриклеточному паразитизму.
Авирулентные и низковирулентные бактерии погибают и перевариваются в фаголизосомах лизосомами гидролазами. Такой фагоцитоз называют завершенным.
Многие вирулентные бактерии часто не погибают и могут длительно персистировать внутри фагоцитов.
Факультативно или облигатно внутриклеточные паразиты после эндоцитоза сохраняют жизнеспособность и размножаются внутри фагоцитов, вызывая их гибель и разрушение.
Выживание фагоцитированных микроорганизмов могут обеспечивать различные механизмы. Одни патогенные агенты способны препятствовать слиянию лизосом с фагосомами (токсоплазмы, микобактерии туберкулеза). Другие обладают устойчивасью к действию лизосомных ферментов (гонококки, стафилококки). Третьи после эндоцитоза покидают фагосому, избегая действия микробоцидных факторов, и могут длительно персистировать в цитоплазме фагоцитов (риккетсии, хламидии). В этих случаях фагоцитоз называют незавершенным.
Секреторная функция заключается в секреции фагоцитами биологически активных веществ – цитокинов – это интерлейкин-1 и интерлейкин-2, которые являются клеточными медиаторами, оказывающими регулирующее действие на пролиферацию, дифференциацию и функции фагоцитов, лимфоцитов, лимфобластов и других клеток. Макрофаги продуцируют и секретируют такие важные регуляторные факторы, как простагландины, лейкотриены, циклические нуклеотиды с широким спектром биологической активности.
Кроме того макрофаги синтезируют и секретируют ряд продуктов, обладающих антибактериальной, антивирусной и цитотоксической активностью (кислородные радикалы О2— Н2О2, лизоцим, интерферон и др.).
Фагоцитоз усиливается антителами-опсонинами, так как связанный или антиген легче адсорбируется на поверхности фагоцита, вследствие наличия у последнего рецепторов к этим антителам. Такое усиление фагоцитоза антителами названо опсонизацией, т.е. подготовкой микроорганизмов к захвату фагоцитами. Фагоцитоз опсонизированых антигенов называют иммунным.
Для характеристики активности фагоцитоза введен фагоцитарный показатель. Для определения его подсчитывают под микроскопом число бактерий, поглощенных одним фагоцитом. Пользуются также опсонофагоцитарным индексом, представляющим отношение фагоцитарных показателей, полученных с иммунной и не иммунной сывороткой. Фагоцитарный показатель и опсонофагоцитарный индекс используют в клинической иммунологии для оценки состояния иммунитета и иммунного статуса.
Фагоцитоз играет большую роль в противобактериальной, противогрибковой и противовирусной защите, поддержании резистентности организма к чужеродным веществам. Фагоциты также оказывают активирующее и супрессивное действие на лимфоциты, принимают участие в реанимации иммунологической толерантности, антиинфекционного, трансплантационного и противоопухолевого иммунитета, некоторых форм аллергии (ГЗТ).
necontactor.com