Потенциал действия. Если участок нервного или мышечного волокна подвергнуть действию достаточно сильного раздражителя (например, толчка электрического тока), в этом участке возникает возбуждение, одним из наиболее важных проявлений которого служит быстрое колебание МП, называемое потенциалом действия (ПД)
Причиной возникновения ПД является изменение ионной проницаемости мембраны. В состоянии покоя, как уже говорилось, проницаемость мембраны для К+превышает натриевую проницаемость. Вследствие этого поток положительно заряженных ионов из протоплазмы наружу превышает противоположный поток Na+. Поэтому мембрана в покое снаружи заряжена положительно.
При действии на клетку раздражителя проницаемость мембраны для ионов Na+ резко повышается, и в конечном итоге становится примерно в 20 раз больше проницаемости для К+. Поэтому поток ионов Na+в клетку начинает значительно превышать направленный наружу поток К+. Ток Na+достигает величины +150 мв. Одновременно несколько уменьшается выход К+из клетки. Все это приводит к извращению (реверсии) МП, и наружная поверхность мембраны становится заряженной электро отрицательно по отношению к внутренней поверхности. Указанный сдвиг и регистрируется в виде восходящей ветви пика ПД (фаза деполяризации).
При внутриклеточном отведении можно обнаружить, что поверхность возбужденного участка на очень короткий интервал, измеряемый тысячными долями секунды, становится заряженным электроотрицательно по отношению к соседнему, покоящемуся участку, т.е. при возбуждении происходит т.н. "перезарядка мембраны". Точные измерения показали, что амплитуда ПД на 30-50 мв превышает величину МП. Причина этого состоит в том, что при возбуждении происходит не просто исчезновение ПП, а возникает разность потенциалов обратного знака, в результате чего наружная поверхность мембраны становится заряженной отрицательно по отношению у ее внутренней стороне.
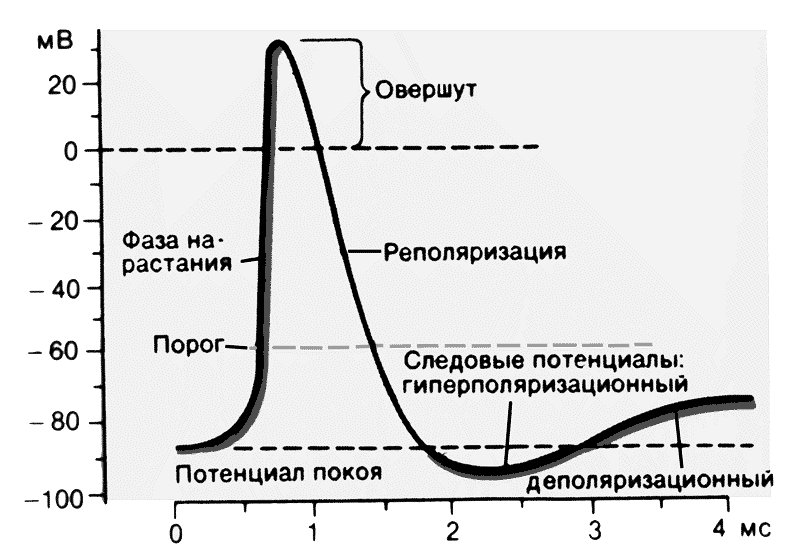
Потенциал действия протекает фазно. Временной ход потенциала действия включает четыре последовательных этапа: локальный ответ, деполяризацию, реполяризацию и следовые потенциалы (рис. 2). В ПД принято различать его пик (т.н. спайк - spike) и следовые потенциалы. Пик ПД имеет восходящую и нисходящую фазы. Перед восходящей фазой регистрируется более или менее выраженный т.н. местный потенциал , или локальный ответ. Поскольку во время восходящей фазы исчезает исходная поляризация мембраны, ее называют фазой деполяризации; соответственно нисходящую фазу, в течение которой поляризация мембраны возвращается к исходному уровню, называется фазой реполяризации. Продолжительность пика ПД в нервных и скелетных мышечных волокнах варьирует в пределах 0,4-5,0 мсек. При этом фаза реполяризации всегда продолжительнее.
Рис. 2. Фазы и временной ход потенциала действия.
Кроме пика, в ПД различают два следовых потенциала - следовую деполяризацию (следовой отрица-тельный потенциал) и следовую гиперполяризацию (следовой положи-тельный потенциал. Амплитуда этих потенциалов не превышает нескольких милливольт, а длительность варьирует от нескольких десятков до сотен миллисекунд. Следовые потенциалы связаны с восстановительными процессами, развивающимися в мышцах и нерве после окончания возбуждения.
Промежуток времени, в течение которого сохраняется активное состояние в виде ПД, неодинаков в разных возбудимых структурах. В нейронах он составляет около 1 мс, в волокнах скелетных мышц – 10 мс, в миокарде достигает 200–250 мс.
Левое крыло графической записи ПД, отражающее изменение потенциала в электроположительную сторону называется деполяризацией. Область электроположительности носит название овершута, правое крыло ПД, свидетельствующее о восстановлении исходного поляризованного состояния мембраны принято называть реполяризацией. Часто, но не всегда возвращение ПД к исходному уровню в состоянии покоя происходит с наличием фаз в форме так называемых следовых потенциалов. Следовые потенциалы неодинаковы в мышцах и нервах. В волокнах скелетных мышц фаза реполяризации очень замедлена. Примерно через 1 мс после начала ПД наблюдается отчетливый перегиб крыла реполяризации – это следовая деполяризация. В нейроне чаще всего кривая реполяризации быстро пересекает уровень МПП и на некоторое время потенциал мембраны становится более электроотрицательным, чем МП. Это явление называют следовой гиперполяризацией.
Повышение проницаемости мембраны для ионов Na+продолжается в нервных клетках очень короткое время. Связано оно с кратковременным открытием т.н. Na+-каналов (точнее, заслонок М в этих каналах), которое затем сменяется срочным закрытием Na+-пор с помощью т.н. Н-ворот. Этот процесс называется натриевой инактивацией. В результате поток Na в клетку прекращается.
Наличие специальных Na- и К- каналов и сложного механизма запирания и открытия ворот изучено биофизиками достаточно хорошо. Показано, что существуют избирательные механизмы, регулирующие те или иные каналы. Например, яд тетродотоксин блокирует только Na-поры, а тетраэтиламмоний - только К-поры. Показано, что у некоторых клеток возникновение возбуждения связано в изменением проницаемости мембраны для Са++, в других - для Mg+. Исследования механизмов изменения проницаемости мембран продолжаются.
В результате Na-инактивации и одновременного увеличения К- проницаемости происходит усиленный выход положительных ионов К+ из протоплазмы во внешний раствор. В итоге этих двух процессов происходит восстановление поляризованного состояния мембраны (реполяризация) , и наружная ее поверхность вновь приобретает положительный заряд. В дальнейшем происходят процессы восстановления нормального ионного состава клетки и необходимого градиента концентрации ионов за счет активизации деятельности Na-К-насоса. В результате повышения проводимости резко возрастает поток катионов Na+, поэтому отрицательный заряд в клетке вблизи внутренней стороны поверхности мембраны также резко уменьшается вплоть до преобладания положительных зарядов. В результате происходит изменение знака потенциала, достигающего +30 мВ. После этого проводимость мембраны дляNa+также резко снижается.
Для нормального протекания ПД играет существенную роль и изменение проводимости мембраны для K+, которая начинает возрастать позже возрастания проводимости дляNa+. Увеличение относительно медленного выходаK+из клетки в фазу снижения проводимости дляNa+вызывает реполяризацию мембраны.
Таким образом, в живой клетке существуют два различных типа движения ионов через мембрану. Один из них осуществляется по градиенту концентрации ионов и не требует затраты энергии, поэтому его называют пассивным транспортом. Он ответственен за возникновение МП и ПД и ведет в конечном итоге к выравниванию концентраций ионов по обе стороны клеточной мембраны. Второй тип движения ионов через мембрану, осуществляющийся против концентрационного градиента, состоит в "выкачивании" ионов Na+ из протоплазмы и "нагнетании" ионов К+ внутрь клетки. Этот тип ионного транспорта возможет лишь при условии затраты энергии - это активный транспорт. Он является результатом работы специальных ферментных систем (т.н. насосов), и благодаря ему восстанавливается исходная разность концентраций, необходимая для поддержания МП.
Условия возникновения возбуждения. Для возникновения ПД необходимо, чтобы под влиянием какого-либо раздражителя произошло повышение ионной проницаемости мембраны возбудимой клетки. Однако, возбуждение возможно лишь при условии, если действующий на мембрану агент имеет некоторую минимальную (пороговую) величину, способную изменить мембранный потенциал (МП, или Ео ) до некоторого критического уровня (Ек, критический уровень деполяризации). Стимулы, сила которых ниже пороговой величины, называются подпороговыми, выше - надпороговыми. Показано, что пороговая сила, необходимая для возникновения возбуждения при внутриклеточном микроэлектроде равна 10 -7 - 10-9 А.
Таким образом, главным условием для возникновения ПД является следующее: мембранный потенциал должен стать равным или меньше критического уровня деполяризации ( Ео <= Eк)
Инактивация Na+-системы.Na+-системой обозначают механизм, позволяющий в течение нескольких долей миллисекунды многократно (до 20 раз) увеличить проводимость клеточной мембраны для Na+. Достигнув пикового значения, примерно через 0,5 мс проводимость мембраны для Na+ начинает снижаться. Быстрое снижение проводимости для Na+ называют инактивацией Na+-системы. В основе инактивации Na+-системы лежит переход в инактивационное состояние потенциалзависимых Na+-каналов. Поэтому скорость и степень снижения проводимости потенциалзависимы. Это означает, что чем больше отличается потенциал мембраны от мембранного потенциала покоя в сторону электроположительности, тем сильнее инактивирована Na+-система. Поэтому деполяризация мембраны вызывает снижение тока Na+ внутрь клетки. С одной стороны, это свидетельствует о том, что усиление тока Na+ само себе служит причиной его быстрого последующего снижения и начала развития реполяризации. С другой стороны, это означает, что если исходный потенциал клетки выше потенциала покоя на 20–30 мВ, то Na+-система полностью инактивирована и никакая последующая деполяризация уже не может активировать ее, т.е. вызвать резкое увеличение проводимости для Na+ и генерацию ПД.
studfiles.net
Раздражимость-способность живых клеток реагировать изменением обмена веществ в ответ на действие раздражителей.
Раздражние-реакция живой клетки,ткани,органа на внешнее воздействие.
Возбудимость- свойство клеток отвечать на раздражение возбуждением.
Возбуждение-ответ ткани на ее раздражение, проявляющийся в специфической для нее функции(проведение возбуждения, сокращение мышцы, секреция железы) и неспецифических реакциях(генерация потенциала действия, метаболические изменения).
Адекватными называются раздражители, на действие которых определенный вид организмов, орган или живая ткань приспособились соответственно реагировать в естественных условиях на протяжении многих тысячелетий исторического развития. Например, для глаза адекватны световые раздражители, для уха — звуковые и т. д.
Неадекватными называются раздражители, не соответствующие строению и функции воспринимающего органа, например, механические и электрические раздражители для глаза. Они также вызывают изменение в нем обмена веществ, но отличающееся от вызванного действием адекватного раздражителя, и поэтому не могут вызвать точно такие же изменения функции, как адекватный раздражитель.
Изменение возбудимости при возбуждении. При возбуждении возбудимость изменяется пофазно. 1) фаза первичной экзальтации - возбудимость выше нормы, реакция на порошковый и подпороговый раздражитель (соответствует 1 фазе ПД - медленной деполяризации) 2) фаза абсолютной рефрактерности - ответная реакция на раздражитель отсутствует, что обусловлено инактивацией натриевых каналов (соответствует быстрой деполяризации ПД) 3) фаза относительной рефрактерности - возбудимость восстанавливается, и ответная реакция становится возможной только при действии раздражителя надпороговой силы, что обусловлено выходящим калиевым током (соответствует фазе реполяризации) 4) фаза вторичной экзальтации - ответная реакция на подпороговый раздражитель (соответствует следовой деполяризации) 5) фаза субнормальной возбудимости - возбудимость ниже нормы, ответ возможен на действие надпороговой силы (соответствует следовой гиперполяризации)
Прямым раздражением называется непосредственное действие раздражителя на орган, например, раздражение электрическим током мышцы, выпрепарованной из организма. Непрямое раздражение производится действием раздражителя на рецепторы—специальные органы, расположенные на внешней поверхности организма или внутри его и воспринимающие раздражение, например, глаза, уши, органы обоняния, вкуса, рецепторы кожи, мышц, суставов, сухожилий, внутренних органов. При непрямом раздражении органа волны, или импульсы возбуждения из рецептора сначала поступают по центростремительным нервам в центральную нервную систему, а затем уже направляются в орган по центробежным нервам и вызывают его функцию или вызывают образование в некоторых органах веществ, действующих через кровь.
3)Критерии оценки возбудимости: пороговая сила, пороговое время, критический уровень деполяризации.
Пороговая сила – это наименьшая сила раздражителя, способная вызвать ПД при неограниченном во времени действии раздражителя. При использовании в качестве раздражителя электрический ток, его пороговая сила равна 1 реобазе. Если возбудимость ткани высока, пороговая сила раздражителя мала.
Пороговое время – это минимальное время, в течение которого должен действовать на ткань раздражитель пороговой силы, чтобы вызвать ее возбуждение. Хронаксия – наименьшее время, в течение которого должен действовать ток в две реобазы, чтобы вызвать возбуждение
Критический уровень деполяризации - пороговый уровень деполяризации плазмалеммы возбудимой клетки. Уровень деполяризации, при достижении которого возникает потенциал действия. В пределах от потенциала покоя до критического уровня деполяризации между интенсивностью раздражителя и уровнем деполяризации существует приблизительно линейная зависимость. При деполяризации достигающей критического уровня такая зависимость отсутствует (закон «все или ничего»).
studfiles.net
Общие положения
Поляризация мембраны живой клетки обусловлена отличием ионного состава с её внутренней и наружной стороны. Когда клетка находится в спокойном (невозбуждённом) состоянии, ионы по разные стороны мембраны создают относительно стабильную разность потенциалов, называемую потенциалом покоя. Если ввести внутрь живой клетки электрод и измерить мембранный потенциал покоя, он будет иметь отрицательное значение (порядка −70 — −90 мВ). Это объясняется тем, что суммарный заряд на внутренней стороне мембраны существенно меньше, чем на внешней, хотя с обеих сторон содержатся и катионы, и анионы. Снаружи — на порядок больше ионов натрия, кальция и хлора, внутри — ионов калия и отрицательно заряженных белковыхмолекул, аминокислот, органических кислот, фосфатов, сульфатов. Надо понимать, что речь идёт именно о заряде поверхности мембраны — в целом среда и внутри, и снаружи клетки заряжена нейтрально.
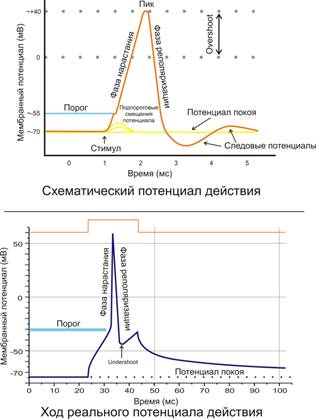
Потенциал мембраны может изменяться под действием различных стимулов. Искусственным стимулом может служить электрический ток, подаваемый на внешнюю или внутреннюю сторону мембраны через электрод. В естественных условиях стимулом часто служит химический сигнал от соседних клеток, поступающий черезсинапс или путём диффузной передачи через межклеточную среду. Смещение мембранного потенциала может происходить в отрицательную (гиперполяризация) или положительную (деполяризация) сторону.
В нервной ткани потенциал действия, как правило, возникает при деполяризации — если деполяризация мембраны нейрона достигает некоторого порогового уровня или превышает его, клетка возбуждается, и от её тела к аксонам и дендритам распространяется волна электрического сигнала. (В реальных условиях на теле нейрона обычно возникают постсинаптические потенциалы, которые сильно отличаются от потенциала действия по своей природе — например, они не подчиняются принципу «всё или ничего». Эти потенциалы преобразуются в потенциал действия на особом участке мембраны — аксонном холмике, так что потенциал действия не распространяется на дендриты).
Это обусловлено тем, что на мембране клетки находятся ионные каналы — белковые молекулы, образующие в мембране поры, через которые ионы могут проходить с внутренней стороны мембраны на наружную и наоборот. Большинство каналов ионоспецифичны — натриевый канал пропускает практически только ионы натрия и не пропускает другие (это явление называют селективностью). Мембрана клеток возбудимых тканей (нервной и мышечной) содержит большое количество потенциал-зависимых ионных каналов, способных быстрореагировать на смещение мембранного потенциала. Деполяризация мембраны в первую очередь вызывает открытие потенциал-зависимых натриевых каналов. Когда одновременно открывается достаточно много натриевых каналов, положительно заряженные ионы натрия устремляются через них на внутреннюю сторону мембраны. Движущая сила в данном случае обеспечивается градиентом концентрации (с внешней стороны мембраны находится намного больше положительно заряженных ионов натрия, чем внутри клетки) и отрицательным зарядом внутренней стороны мембраны (см. Рис. 2). Поток ионов натрия вызывает ещё бо́льшее и очень быстрое изменение мембранного потенциала, которое и называют потенциалом действия(в специальной литературе обозначается ПД).
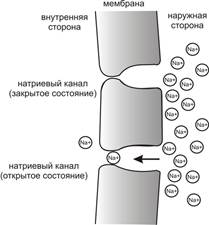
Рис. 3. Простейшая схема, демонстрирующая мембрану с двумя натриевыми каналами в открытом и закрытом состоянии, соответственно
Согласно закону «всё-или-ничего» мембрана клетки возбудимой ткани либо не отвечает на стимул совсем, либо отвечает с максимально возможной для неё на данный момент силой. То есть, если стимул слишком слаб и порог не достигнут, потенциал действия не возникает совсем; в то же время, пороговый стимул вызовет потенциал действия такой же амплитуды, как и стимул, превышающий пороговый. Это отнюдь не означает, что амплитуда потенциала действия всегда одинакова — один и тот же участок мембраны, находясь в разных состояниях, может генерировать потенциалы действия разной амплитуды.
После возбуждения нейрон на некоторое время оказывается в состоянии абсолютной рефрактерности, когда никакие сигналы не могут его возбудить снова, затем входит в фазу относительной рефрактерности, когда его могут возбудить исключительно сильные сигналы (при этом амплитуда ПД будет ниже, чем обычно). Рефрактерный период возникает из-за инактивации быстрого натриевого тока, то есть инактивации натриевых каналов.
megaobuchalka.ru
Литература: 1. Пенроуз Р. НОВЫЙ УМ КОРОЛЯ. О компьютерах, мышлении и законах физики. 2. Грегори Р. Л. Разумный глаз. 3. Леках В. А. Ключ к пониманию физиологии. 4. Гамов Г., Ичас М. Мистер Томпкинс внутри самого себя: Приключения в новой биологии.
bukvasha.ru
Возбуждение — ответ ткани на раздражение, проявляющийся помимо неспецифических реакций (генерация потенциала действия, метаболические изменения) в выполнении специфической для этой ткани функции. Возбудимыми являются нервная (проведение возбуждения), мышечная (сокращение) и железистая (секреция) ткани.Возбудимость — свойство клеток отвечать на раздражение возбуждением.
При возбуждении живая система переходит из состояния относительного физиологического покоя к состоянию физиологической активности. В основе возбуждения лежат сложные физико-химические процессы. Мерой возбуждения является сила раздражителя, которая вызывает возбуждение.
Возбудимые ткани обладают высокой чувствительностью к действию слабого электрического тока (электрическая возбудимость), что впервые продемонстрировал Л. Гальвани.
Возбуждение бывает местное (или локальное) и распространяющееся. Местное возбуждение представляет незначительные изменения в поверхностной мембране клеток, а распространяющееся возбуждение связано с передачей всего комплекса физиологических изменений (импульса возбуждения) вдоль нервной или мышечной ткани.
Торможение – активный нервный процесс, вызываемый возбуждением и проявляющийся в угнетении или предупреждении другой волны возбуждения.
Торможение может развиваться только в форме локального ответа.
Выделяют два типа торможения:
В ней выделяют фазу быстрой реполяризации и фазу медленной реполяризации, в свою очередь, фаза медленной реполяризации представлена следовыми процессами (потенциалами):
следовая негативность (следовая деполяризация) и следовая позитивность (следовая гиперполяризация). Амплитудно-временные характеристики потенциала действия нерва, скелетной мышцы таковы: амплитуда потенциала действия 140-150 мВ; длительность пика потенциала действия (фаза деполяризации + фаза реполяризации) составляет 1-2 мс, длительность следовых потенциалов - 10-50 мс.
Форма потенциала действия (при внутриклеточном отведении) зависит от вида возбудимой ткани: у аксона нейрона, скелетной мышцы - пикообразные потенциалы, у гладких мышц в одних случаях пикообразные, в других - платообразные (например, потенциал действия гладких мышц матки беременной женщины - платообразный, а длительность его составляет почти 1 минуту). У сердечной мышцы потенциал действия имеет платообразную форму.
Природа ПД:
При исследовании ПД аксонов и сомы нервной клетки, ПД скелетной мышцы было установлено, что фаза деполяризации обусловлена значительным повышением проницаемости для ионов натрия, которые входят в клетку в начале процесса возбуждения и таким образом уменьшают существующую разность потенциала (деполяризация). При этом чем выше степень деполяризации, тем выше становится проницаемость натриевых каналов, тем больше входит ионов натрия в клетку и тем выше степень деполяризации. В этот период происходит не только снижение разности потенциалов до нуля, но и изменение поляризованности мембраны - на высоте пика ПД внутренняя поверхность мембраны заряжена положительно по отношению к наружной (явление реверсии, или овершута). Однако бесконечно этот процесс идти не может: в результате закрытия инактивационных ворот натриевые каналы закрываются, и приток натрия в клетку прекращается. Затем наступает фаза реполяризации. Она связана с увеличением выхода из клетки ионов калия. Это происходит за счет того, что в результате деполяризации большая часть калиевых каналов, которые в условиях покоя были закрыты, открываются и «+» заряды уходят за пределы клетки. Вначале этот процесс идет очень быстро, потом - медленно, поэтому фаза реполяризации вначале протекает быстро (нисходящая часть пика ПД), а потом медленно (следовая негативность). Этот же процесс лежит в основе фазы следовой гиперполяризации. На фоне следовых потенциалов происходит активация калий-натриевого насоса. Если он работает в электронейтральном режиме (2 иона натрия выносятся из клетки в обмен на 2 вносимых в клетку иона калия), то на форме ПД этот процесс не отражается. Если же насос работает в электрогенном режиме, когда 3 иона натрия выносятся из клетки в обмен на 2 вносимых в клетку иона калия, то в результате на каждый такт работы насоса в клетку вносится на 1 катион меньше, чем выносится, поэтому в клетке постепенно возрастает избыток анионов, т. с. в таком режиме насос способствует появлению дополнительной разности потенциалов. Это явление может лежать в основе фазы следовой гиперполяризации.
В сердечной мышце природа ПД иная: процесс деполяризации обусловлен ионами натрия и кальция - эти ионы входят внутрь клетки в начале фазы деполяризации.
В гладких мышцах сосудов, желудка, кишечника, матки и других образований генерация ПД связана с тем, что в момент возбуждения в клетку входят главным образом не ионы натрия, а ионы кальция.
Соотношение фаз потенциала действия и возбудимости
Уровень возбудимости клетки зависит от фазы ПД. В фазу локального ответа возбудимость возрастает. Это фазу возбудимости называют латентным дополнением.
В фазу реполяризации ПД, когда открываются все натриевые каналы и ионы натрия лавинообразно устремляются в клетку, никакой даже сверхсильный раздражитель не может стимулировать этот процесс. Поэтому фазе деполяризации соответствует фаза полной невозбудимости, или абсолютной рефрактерности.
В фазе реполяризации все большая часть натриевых каналов закрывается. Однако они могут вновь открываться при действии сверхпорогового раздражителя. Т.е. возбудимость начинает вновь повышаться. Этому соответствует фаза относительной невозбудимости, или относительной рефрактерности.
Во время следовой деполяризации МП находится у критического уровня, поэтому даже допороговые стимулы могут вызвать возбуждение клетки. Следовательно, в этот момент ее возбудимость повышена. Эта фаза называется фазой экзальтации, или супернормальной возбудимости.
studfiles.net
